
Paracrine Kynurenic Pathway Activation in the Bone of Young Uremic Rats Can Antagonize Anabolic Effects of PTH on Bone Turnover and Strength through the Disruption of PTH-Dependent Molecular Signaling



Abstract
1. Introduction
2. Results
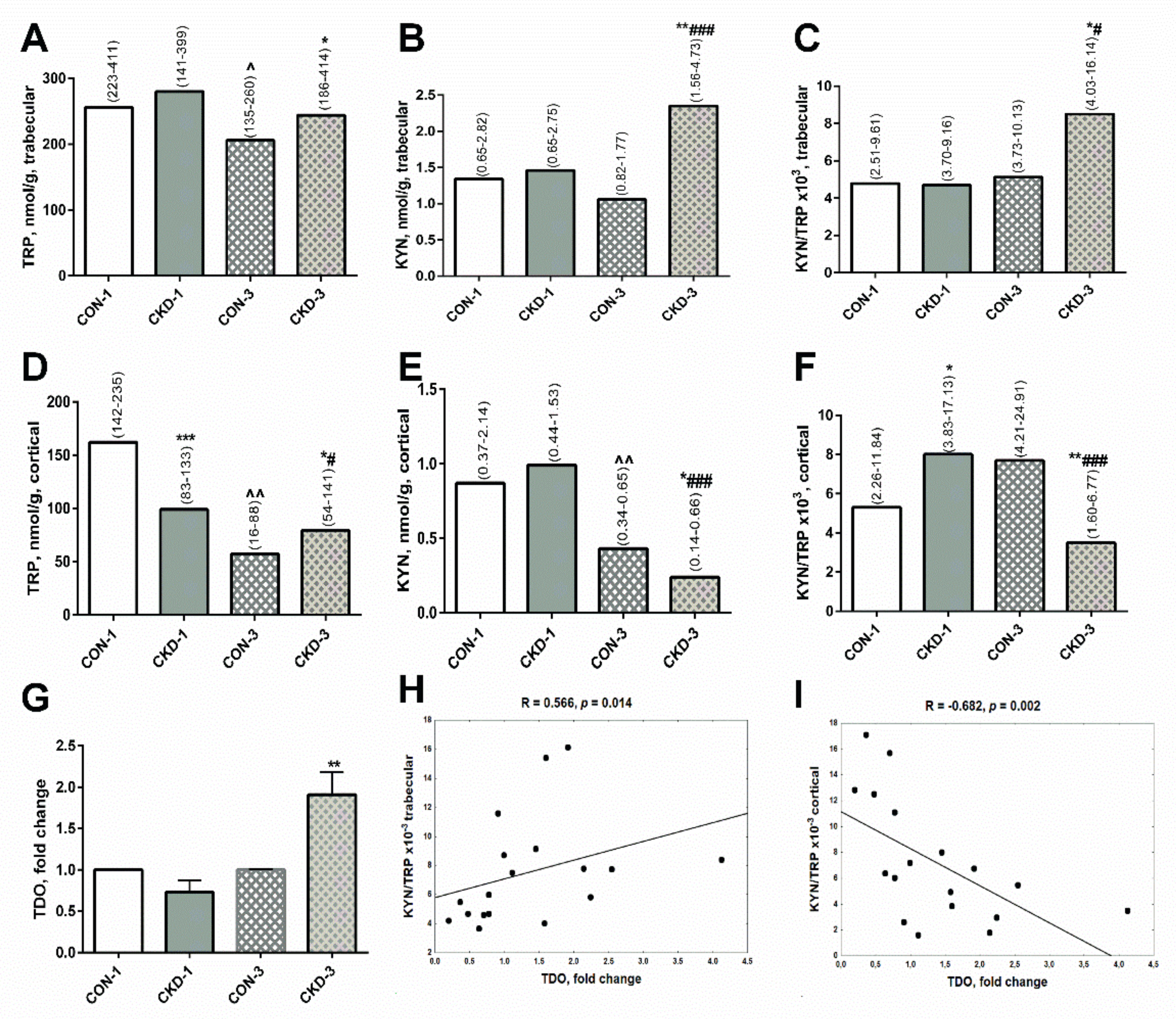
2.1. Paracrine Kynurenic System in Trabecular and Cortical Bone of Young Rats with CKD
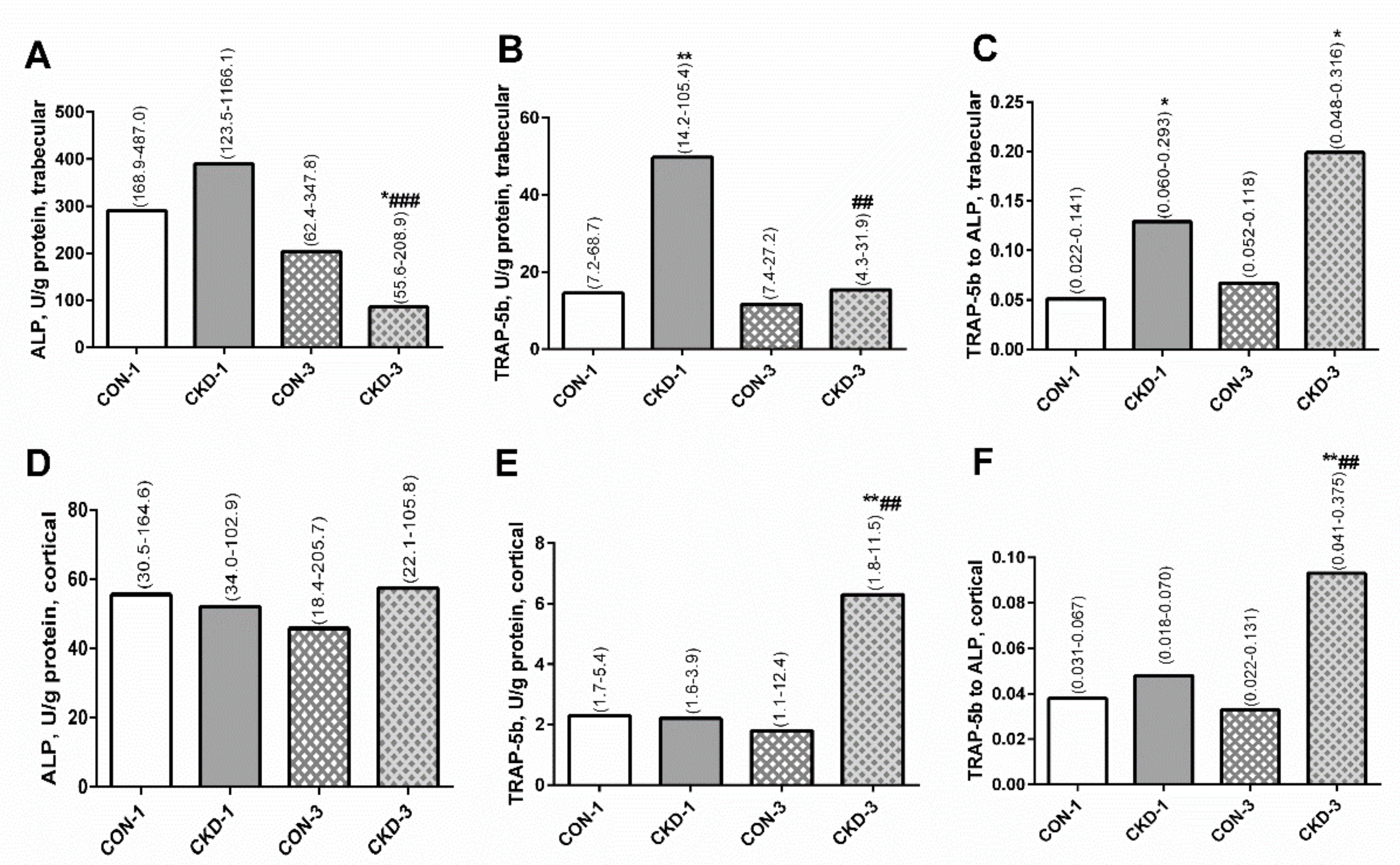
2.2. Bone Turnover in Trabecular and Cortical Bone of Young Rats with CKD
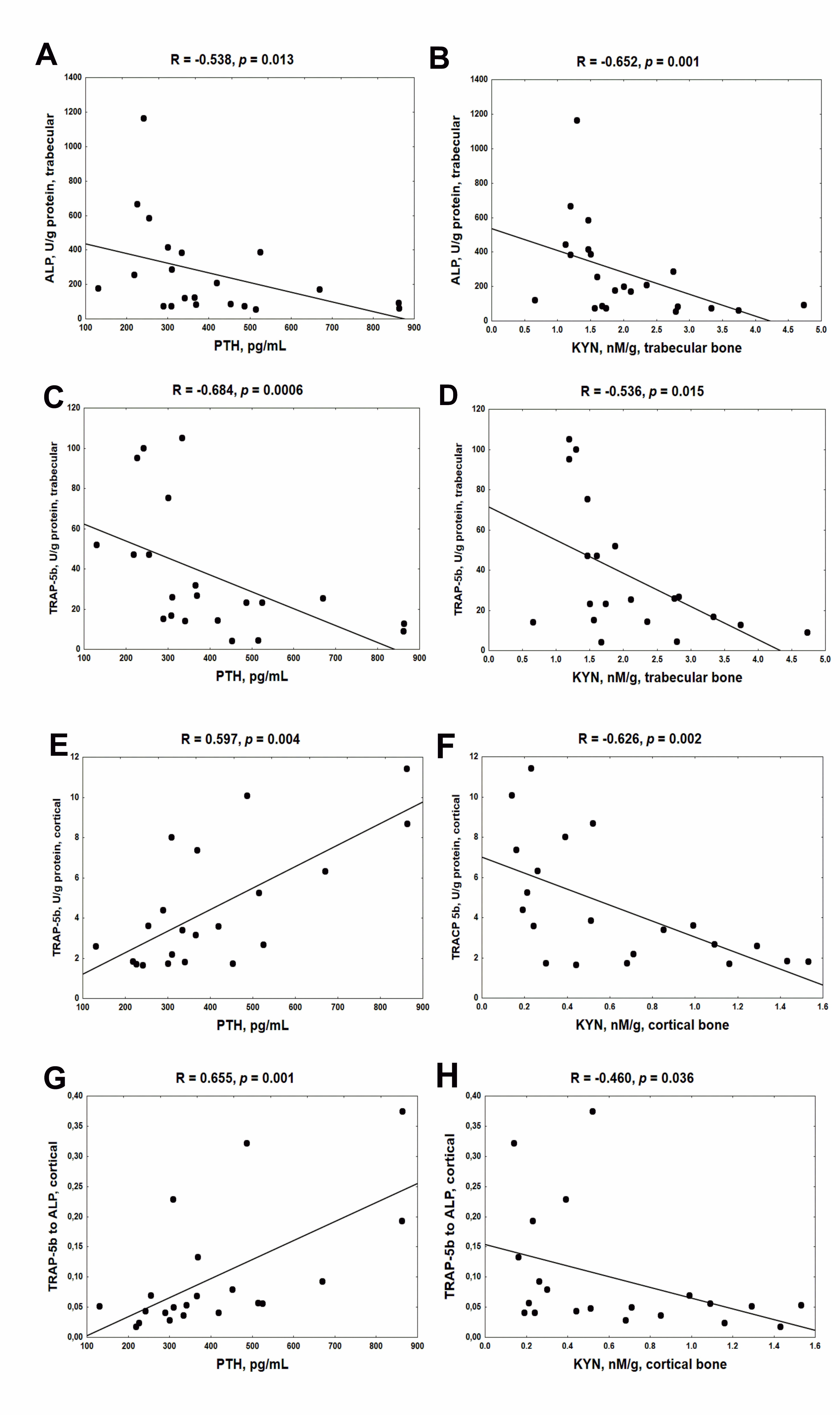
2.3. Association between Serum PTH, Bone KYN and Bone Turnover Markers (BTMs) in Young Rats with CKD
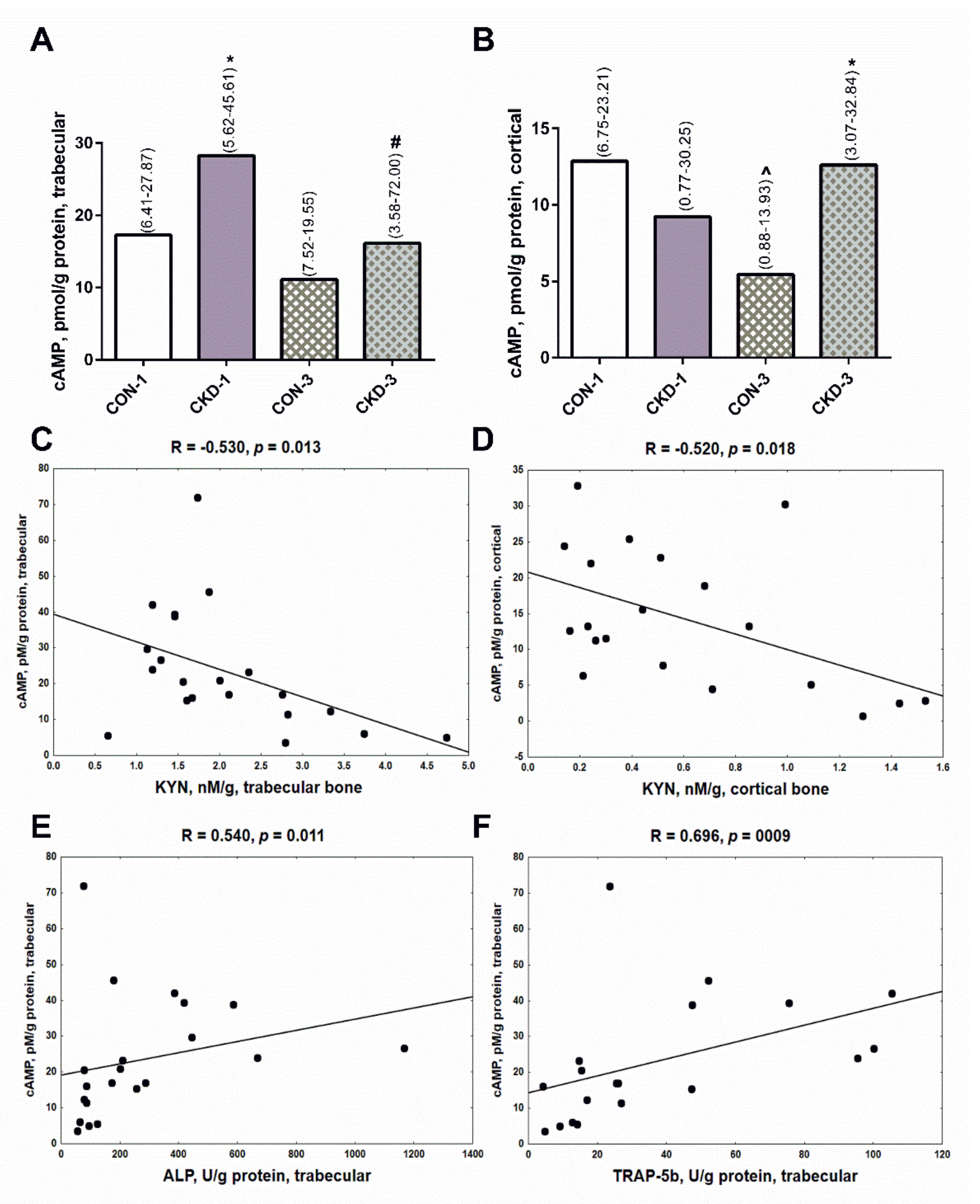
2.4. Bone KYN Can Affect cyclic adenosine monophosphate (cAMP) Levels in Trabecular and Cortical Bone
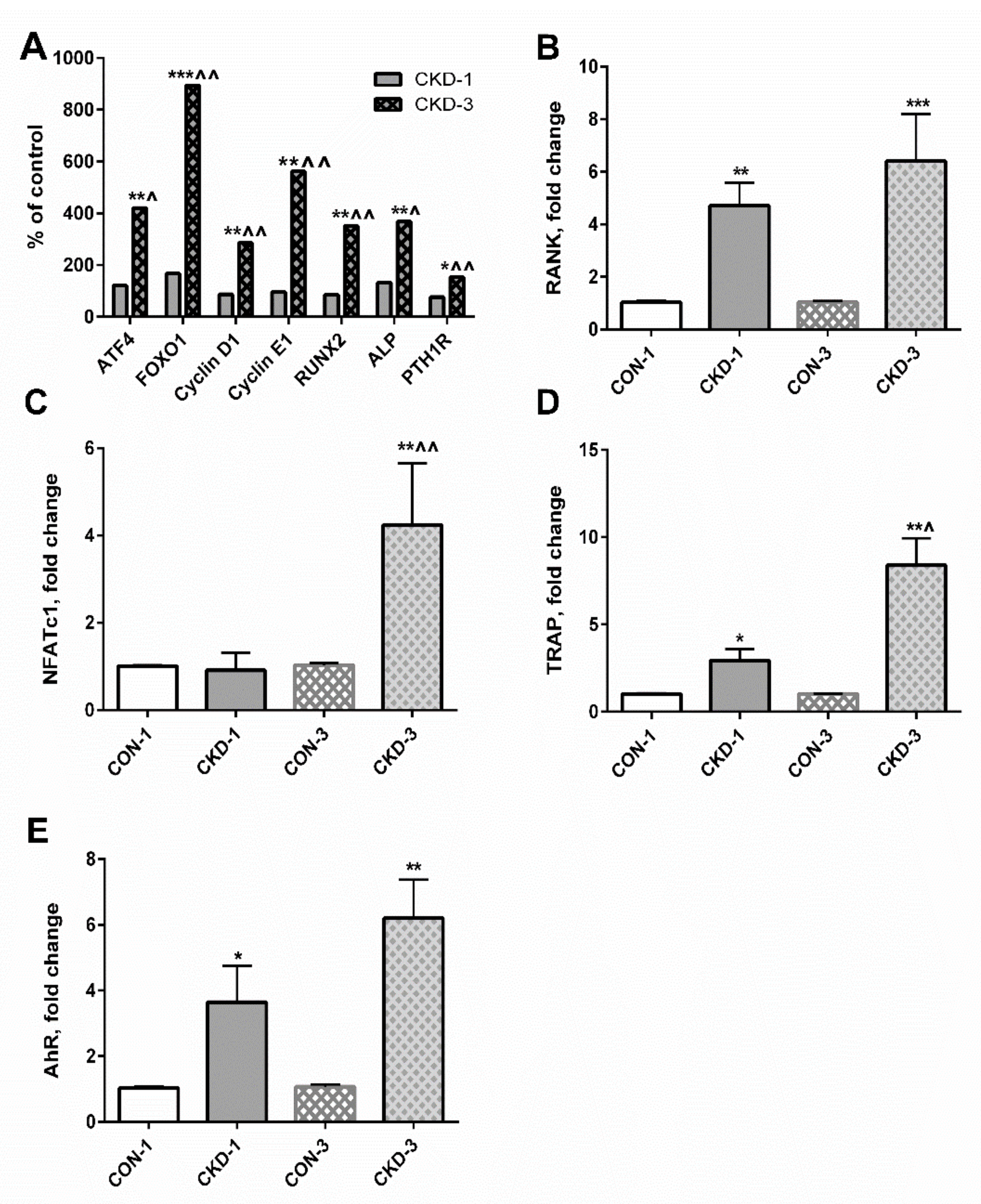
2.5. Expression of Genes Involved in Osteoblasto- and Osteoclastogenesis Can Be Mediated by Both PTH1R and the aryl hydrocarbon receptor (AhR)-Dependent Pathway
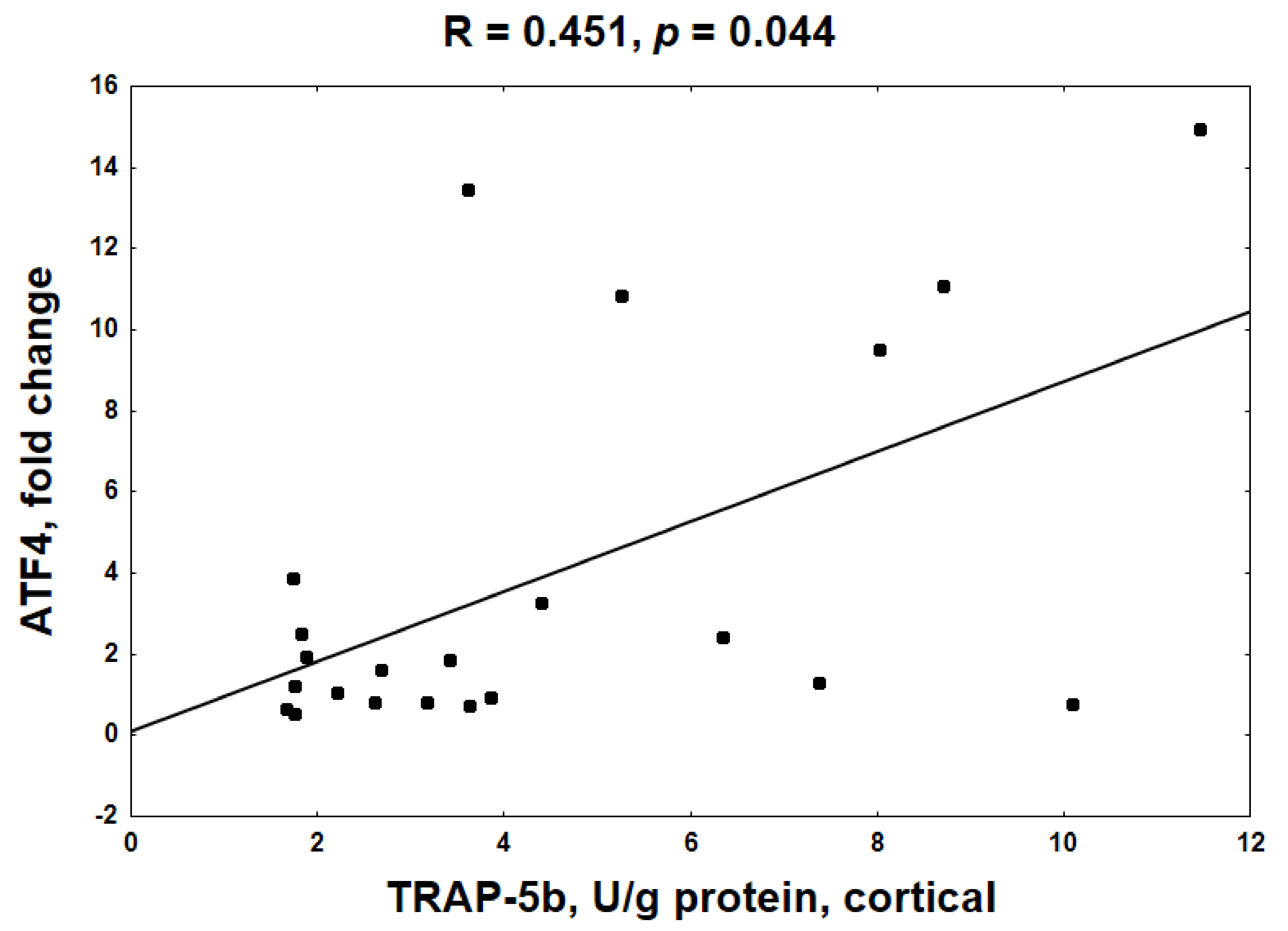
2.6. Mechanistic Relationships between KYN Pathway Activation, Gene Expression, cAMP and BTMs in Bone Tissue of Young Rats with CKD
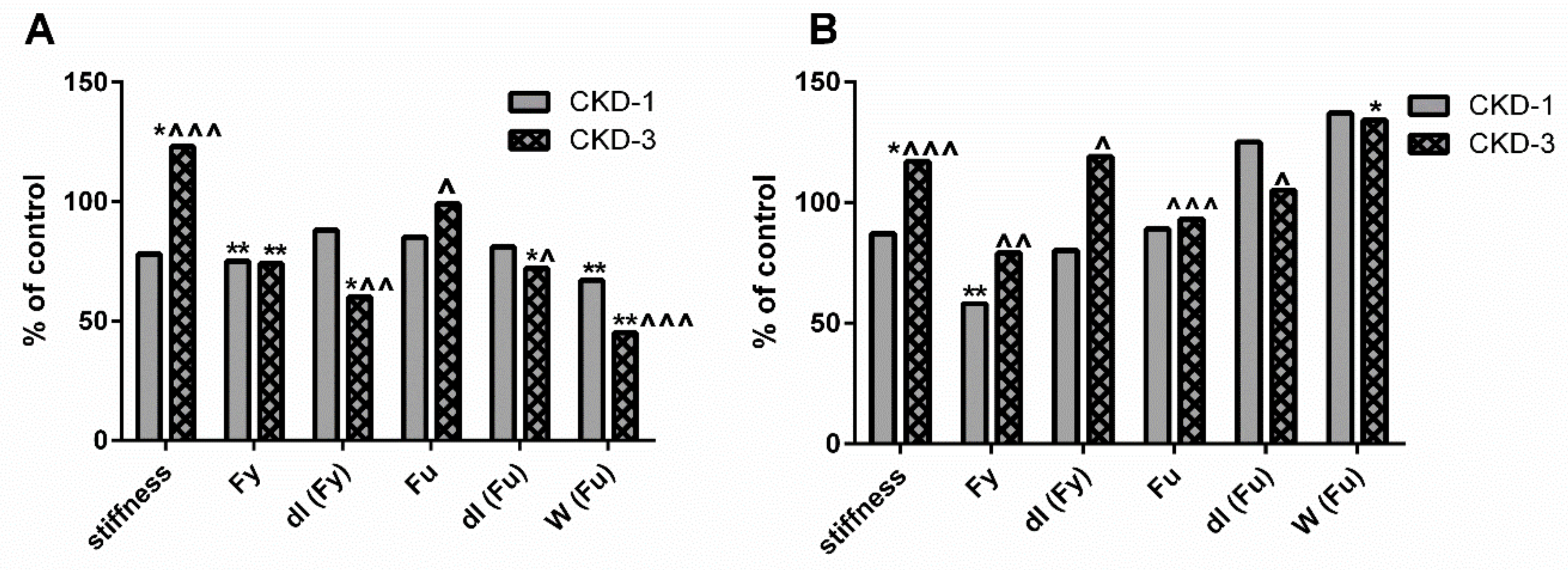
2.7. Endogenous Bone KYN and BTMs Can Affect the Biomechanical Properties of Trabecular and Cortical Bone of Young Rats with CKD
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Sample Preparation
4.3. Determination of TRP and KYN in in Bone Homogenates
4.4. Determination of BTMs Concentrations in Bone Homogenates
4.5. RNA Isolation and RT—qPCR Analysis
4.6. Bone Biomechanical Properties Determination
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kidney Disease: Improving Global Outcomes (KDIGO) CKD-MBD Update Work Group. KDIGO 2017 Clinical Practice Guideline Update for the Diagnosis, Evaluation, Prevention, and Treatment of Chronic Kidney Disease-Mineral and Bone Disorder (CKD-MBD). Kidney Int. Suppl. 2017, 7, 1–59. [Google Scholar] [CrossRef]
- Sidibe, A.; Auguste, D.; Desbiens, L.C.; Fortier, C.; Wang, Y.P.; Jean, S.; Moore, L.; Mac-Way, F. Fracture risk in dialysis and kidney transplanted patients: A systematic review. JBMR Plus 2019, 3, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Spasovski, G.B.; Bervoets, A.R.; Behets, G.J.; Ivanovski, N.; Sikole, A.; Dams, G.; Couttenye, M.M.; De Broe, M.E.; D’Haese, P.C. Spectrum of renal bone disease in end-stage renal failure patients not yet on dialysis. Nephrol. Dial. Transplant. 2003, 18, 1159–1166. [Google Scholar] [CrossRef]
- Sprague, S.M.; Bellorin-Font, E.; Jorgetti, V.; Carvalho, A.B.; Malluche, H.H.; Ferreira, A.; D’Haese, P.C.; Drueke, T.B.; Du, H.; Manley, T.; et al. Diagnostic accuracy of bone turnover markers and bone histology in patients with CKD treated by dialysis. Am. J. Kidney Dis. 2016, 67, 559–566. [Google Scholar] [CrossRef]
- Barreto, F.C.; Barreto, D.V.; Moyses, R.M.; Neves, K.R.; Canziani, M.E.; Draibe, S.A.; Jorgetti, V.; Carvalho, A.B. K/DOQI-recommended intact PTH levels do not prevent low-turnover bone disease in hemodialysis patients. Kidney Int. 2008, 73, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Karsdal, M.A.; Martin, T.; Bollerslev, J.; Christiansen, C.; Henriksen, K. Are nonresorbing osteoclasts sources of bone anabolic activity? J. Bone Miner. Res. 2007, 22, 487–494. [Google Scholar] [CrossRef]
- Henriksen, K.; Neutzsky-Wulff, A.V.; Bonewald, L.F.; Karsdal, M.A. Local communication on and within bone controls bone remodeling. Bone 2009, 44, 1026–1033. [Google Scholar] [CrossRef]
- Drueke, T.B.; Massy, Z.A. Changing bone patterns with progression of chronic kidney disease. Kidney Int. 2016, 89, 289–302. [Google Scholar] [CrossRef]
- Evenepoel, P.; Bover, J.; Ureña Torres, P. Parathyroid hormone metabolism and signaling in health and chronic kidney disease. Kidney Int. 2016, 90, 1184–1190. [Google Scholar] [CrossRef]
- Wronski, T.J.; Yen, C.F.; Qi, H.; Dann, L.M. Parathyroid hormone is more effective than estrogen or bisphosphonates for restoration of lost bone mass in ovariectomized rats. Endocrinology 1993, 132, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Demiralp, B.; Chen, H.L.; Koh, A.J.; Keller, E.T.; McCauley, L.K. Anabolic actions of parathyroid hormone during bone growth are dependent on c-fos. Endocrinology 2002, 143, 4038–4047. [Google Scholar] [CrossRef][Green Version]
- Black, D.M.; Greenspan, S.L.; Ensrud, K.E.; Palermo, L.; McGowan, J.A.; Lang, T.F.; Garnero, P.; Bouxsein, M.L.; Bilezikian, J.P.; Rosen, C.J.; et al. The effects of parathyroid hormone and alendronate alone or in combination in postmenopausal osteoporosis. N. Engl. J. Med. 2003, 349, 1207–1215. [Google Scholar] [CrossRef]
- Finkelstein, J.S.; Hayes, A.; Hunzelman, J.L.; Wyland, J.J.; Lee, H.; Neer, R.M. The effects of parathyroid hormone, alendronate, or both in men with osteoporosis. N. Engl. J. Med. 2003, 349, 1216–1226. [Google Scholar] [CrossRef]
- Martin, T.J. Does bone resorption inhibition affect the anabolic response to parathyroid hormone? Trends Endocrinol. Metab. 2004, 15, 49–50. [Google Scholar] [CrossRef] [PubMed]
- Udagawa, N.; Takahashi, N.; Yasuda, H.; Mizuno, A.; Itoh, K.; Ueno, Y.; Shinki, T.; Gillespie, M.T.; Martin, T.J.; Higashio, K.; et al. Osteoprotegerin produced by osteoblasts is an important regulator in osteoclast development and function. Endocrinology 2000, 141, 3478–3484. [Google Scholar] [CrossRef]
- Znorko, B.; Pawlak, D.; Oksztulska-Kolanek, E.; Domaniewski, T.; Pryczynicz, A.; Roszczenko, A.; Rogalska, J.; Lipowicz, P.; Doroszko, M.; Brzoska, M.M.; et al. RANKL/OPG system regulation by endogenous PTH and PTH1R/ATF4 axis in bone: Implications for bone accrual and strength in growing rats with mild uremia. Cytokine 2018, 106, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Anaya, J.M.; Bollag, W.B.; Hamrick, M.W.; Isales, C.M. The role of tryptophan metabolites in musculoskeletal stem cell aging. Int. J. Mol. Sci. 2020, 21, 6670. [Google Scholar] [CrossRef] [PubMed]
- El Refaey, M.; McGee-Lawrence, M.E.; Fulzele, S.; Kennedy, E.J.; Bollag, W.B.; Elsalanty, M.; Zhong, Q.; Ding, K.H.; Bendzunas, N.G.; Shi, X.M.; et al. Kynurenine, a tryptophan metabolite that accumulates with age, induces bone loss. J. Bone Miner. Res. 2017, 32, 2182–2193. [Google Scholar] [CrossRef]
- Pawlak, D.; Oksztulska-Kolanek, E.; Znorko, B.; Domaniewski, T.; Rogalska, J.; Roszczenko, A.; Brzóska, M.M.; Pryczynicz, A.; Kemona, A.; Pawlak, K. The association between elevated levels of peripheral serotonin and its metabolite—5-hydroxyindoleacetic acid and bone strength and metabolism in growing rats with mild experimental chronic kidney disease. PLoS ONE 2016, 11, e0163526. [Google Scholar] [CrossRef]
- Kalaska, B.; Pawlak, K.; Domaniewski, T.; Oksztulska-Kolanek, E.; Znorko, B.; Roszczenko, A.; Rogalska, J.; Brzoska, M.M.; Lipowicz, P.; Doroszko, M.; et al. Elevated levels of peripheral kynurenine decrease bone strength in rats with chronic kidney disease. Front. Physiol. 2017, 8, 836. [Google Scholar] [CrossRef]
- Mor, A.; Pawlak, K.; Kalaska, B.; Domaniewski, T.; Sieklucka, B.; Zieminska, M.; Cylwik, B.; Pawlak, D. Modulation of the paracrine kynurenic system in bone as a new regulator of osteoblastogenesis and bone mineral status in an animal model of chronic kidney disease treated with LP533401. Int. J. Mol. Sci. 2020, 21, 5979. [Google Scholar] [CrossRef]
- Soeiro, E.M.D.; Castro, L.; Menezes, R.; Elias, R.M.; Dos Reis, L.M.; Jorgetti, V.; Moysés, R.M.A. Association of parathormone and alkaline phosphatase with bone turnover and mineralization in children with CKD on dialysis: Effect of age, gender, and race. Pediatr. Nephrol. 2020, 35, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Singh, R.; Divieti, P.; Guo, J.; Bouxsein, M.L.; Bringhurst, F.R. Contributions of parathyroid hormone (PTH)/PTH-related peptide receptor signaling pathways to the anabolic effect of PTH on bone. Bone 2007, 40, 1453–1461. [Google Scholar] [CrossRef]
- Rixon, R.H.; Whitfield, J.F.; Gagnon, L.; Isaacs, R.J.; Maclean, S.; Chakravarthy, B.; Durkin, J.P.; Neugebauer, W.; Ross, V.; Sung, W.; et al. Parathyroid hormone fragments may stimulate bone growth in ovariectomized rats by activating adenylyl cyclase. J. Bone. Miner. Res. 1994, 9, 1179–1189. [Google Scholar] [CrossRef]
- Pawlak, D.; Domaniewski, T.; Znorko, B.; Oksztulska-Kolanek, E.; Lipowicz, P.; Doroszko, M.; Karbowska, M.; Pawlak, K. The impact of peripheral serotonin on leptin-brain serotonin axis, bone metabolism and strength in growing rats with experimental chronic kidney disease. Bone 2017, 105, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, H.; Parker, E.; Hamrick, M.W. Kynurenine signaling through the aryl hydrocarbon receptor: Implications for aging and healthspan. Exp. Gerontol. 2020, 130, 110797. [Google Scholar] [CrossRef]
- Langub, M.C.; Monier-Faugere, M.C.; Qi, Q.; Geng, Z.; Koszewski, N.J.; Malluche, H.H. Parathyroid hormone/parathyroid hormone-related peptide type 1 receptor in human bone. J. Bone. Miner. Res. 2001, 16, 448–456. [Google Scholar] [CrossRef]
- Kim, B.J.; Hamrick, M.W.; Yoo, H.J.; Lee, S.H.; Kim, S.J.; Koh, J.M.; Isales, C.M. The detrimental effects of kynurenine, a tryptophan metabolite, on human bone metabolism. J. Clin. Endocrinol. Metab. 2019, 104, 2334–2342. [Google Scholar] [CrossRef] [PubMed]
- Abou-Samra, A.B.; Juppner, T.H.; Force, M.W.; Freeman, X.F.; Kong, E.; Schipani, E.; Urena, P.; Richards, J.; Bonventre, J.V.; Potts, J.T., Jr.; et al. Expression cloning of a common receptor for parathyroid hormone and parathyroid hormone-related peptide from rat osteoblastlike cells: A single receptor stimulates intracellular accumulation of both cAMP and inositol trisphosphates and increases intracellular free calcium. Proc. Natl. Acad. Sci. USA 1992, 89, 2732–2736. [Google Scholar] [CrossRef]
- Nagy, B.M.; Nagaraj, C.; Meinitzer, A.; Sharma, N.; Papp, R.; Foris, V.; Ghanim, B.; Kwapiszewska, G.; Kovacs, G.; Klepetko, W.; et al. Importance of kynurenine in pulmonary hypertension. Am. J. Physiol. Lung Cell Mol. Physiol. 2017, 313, L741–L751. [Google Scholar] [CrossRef]
- Kanatani, M.; Sugimoto, T.; Takahashi, Y.; Kaji, H.; Kitazawa, R.; Chihara, K. Estrogen via the estrogen receptor blocks cAMP-mediated parathyroid hormone (PTH)-stimulated osteoclast formation. J. Bone Miner. Res. 1998, 13, 854–862. [Google Scholar] [CrossRef]
- Sahbani, K.; Cardozo, C.P.; Bauman, W.A.; Tawfeek, H.A. Abaloparatide exhibits greater osteoanabolic response and higher cAMP stimulation and beta-arrestin recruitment than teriparatide. Physiol. Rep. 2019, 19, e14225. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, N.K.; Lee, S.Y. Current understanding of RANK signaling in osteoclast differentiation and maturation. Mol. Cells 2017, 40, 706–713. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Jiang, L.; Wan, B.; Zhang, W.; Yao, L.; Che, T.; Gan, C.; Su, N.; He, J.; Huang, J.; et al. The role of aryl hydrocarbon receptor in bone remodeling. Prog. Biophys. Mol. Biol. 2018, 134, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.C.; Delany, A.M.; Khouzam, N.M.; Bowen, R.E.; Freymiller, E.G.; Salusky, I.B.; Wesseling-Perry, K. Primary osteoblast-like cells from patients with end-stage kidney disease reflect gene expression, proliferation and mineralization characteristics ex vivo. Kidney Int. 2015, 87, 593–601. [Google Scholar] [CrossRef]
- Yamamoto, T.; Hatabayashi, K.; Arita, M.; Yajima, N.; Takenaka, C.; Suzuki, T.; Takahashi, M.; Oshima, Y.; Hara, K.; Kagawa, K.; et al. Kynurenine signaling through the aryl hydrocarbon receptor maintains the undifferentiated state of human embryonic stem cells. Sci. Signal. 2019, 12, eaaw3306. [Google Scholar] [CrossRef]
- El Refaey, M.; Watkins, C.P.; Kennedy, E.J.; Chang, A.; Zhong, Q.; Ding, K.H.; Shi, X.; Xu, J.; Bollag, W.B.; Hill, W.D.; et al. Oxidation of the aromatic amino acids tryptophan and tyrosine disrupts their anabolic effects on bone marrow mesenchymal stem cells. Mol. Cell. Endocrinol. 2015, 410, 87–96. [Google Scholar] [CrossRef]
- Pierce, J.L.; Roberts, R.L.; Yu, K.; Kendall, R.K.; Kaiser, H.; Davis, C.; Johnson, M.H.; Hill, W.D.; Isales, C.M.; Bollag, W.B.; et al. Kynurenine suppresses osteoblastic cell energetics in vitro and osteoblast numbers in vivo. Exp. Gerontol. 2020, 130, 110818. [Google Scholar] [CrossRef] [PubMed]
- Vidal, C.; Li, W.; Santner-Nanan, B.; Lim, C.K.; Guillemin, G.J.; Ball, H.J.; Hunt, N.H.; Nanan, R.; Duqueet, G. The kynurenine pathway of tryptophan degradation is activated during osteoblastogenesis. Stem Cells 2015, 33, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Park, R.; Madhavaram, S.; Ji, J.D. The role of Aryl-Hydrocarbon Receptor (AhR) in osteoclast differentiation and function. Cells 2020, 9, 2294. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yu, S.; Galson, D.L.; Luo, M.; Fan, J.; Zhang, J.; Guan, Y.; Xiao, G. Activating transcription factor 4 is critical for proliferation and survival in primary bone marrow stromal cells and calvarial osteoblasts. J. Cell Biochem. 2008, 105, 885–895. [Google Scholar] [CrossRef]
- Yu, S.; Franceschi, R.T.; Luo, M.; Fan, J.; Jiang, D.; Cao, H.; Kwon, T.G.; Lai, Y.; Zhang, J.; Patrene, K.; et al. Critical role of activating transcription factor 4 in the anabolic actions of parathyroid hormone in bone. PLoS ONE 2009, 4, e7583. [Google Scholar] [CrossRef]
- Cao, H.; Yu, S.; Yao, Z.; Galson, D.L.; Jiang, Y.; Zhang, X.; Fan, J.; Lu, B.; Guan, Y.; Luo, M.; et al. Activating transcription factor 4 regulates osteoclast differentiation in mice. J. Clin. Investig. 2010, 120, 2755–2766. [Google Scholar] [CrossRef]
- Herlin, M.; Finnilä, M.A.; Zioupos, P.; Aula, A.; Risteli, J.; Miettinen, H.M.; Jämsä, T.; Tuukkanen, J.; Korkalainen, M.; Håkansson, H.; et al. New insights to the role of aryl hydrocarbon receptor in bone phenotype and in dioxin-induced modulation of bone microarchitecture and material properties. Toxicol. Appl. Pharmacol. 2013, 273, 219–226. [Google Scholar] [CrossRef]
- Locklin, R.M.; Khosla, S.; Turner, R.T.; Riggs, B.L. Mediators of the biphasic responses of bone to intermittent and continuously administered parathyroid hormone. J. Cell. Biochem. 2003, 89, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Lasco, A.; Catalano, A.; Morabito, N.; Gaudio, A.; Basile, G.; Trifiletti, A.; Atteritano, M. Adrenal effects of teriparatide in the treatment of severe postmenopausal osteoporosis. Osteoporos. Int. 2011, 22, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Wesseling-Perry, K.; Pereira, R.C.; Tseng, C.H.; Elashoff, R.; Zaritsky, J.J.; Yadin, O.; Sahney, S.; Gales, B.; Juppner, H.; Salusky, I.B. Early skeletal and biochemical alterations in pediatric chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2012, 7, 146–152. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FOXO1 | ATF4 | Cyclin D1 | Cyclin E1 | RUNX2 | ALP | RANK | NFATc1 | TRAP | |
|---|---|---|---|---|---|---|---|---|---|
| PTH1R | R = 0.805 p < 0.001 | R = 0.826 p < 0.001 | R = 0.616 p = 0.002 | R = 0.807 p < 0.001 | R = 0.829 p < 0.001 | R = 0.823 p < 0.001 | R = 0.795 p < 0.001 | R = 0.116 NS | R = 0.057 NS |
| AhR | R = 0.787 p < 0.001 | R = 0.545 p = 0.010 | R = 0.079 NS | R = 0.626 p = 0.002 | R = 0.400 p = 0.065 | R = 0.407 p = 0.060 | R = 0.732 p = 0.001 | R = 0.596 p = 0.007 | R = 0.539 p = 0.017 |
| ALP | RUNX2 | Cyclin E1 | Cyclin D1 | ATF4 | RANK | NFATc1 | TRAP | |
|---|---|---|---|---|---|---|---|---|
| FOXO1 | R = 0.728 p = 0.0001 | R = 0.719 p = 0.0003 | R = 0.866 p < 0.001 | R= 0.388 p = 0.072 | R = 0.772 p < 0.001 | R = 0.909 p < 0.001 | R = 0.340 NS | R = 0.298 NS |
| Cyclin D1 | R = 0.721 p = 0.0001 | R = 0.725 p = 0.0001 | R=0.521 p = 0.013 | R = 0.681 p < 0.001 | R = 0.681 p = 0.0005 | R = −0.299 NS | R = −0.389 NS | |
| Cyclin E1 | R=0.748 p < 0.001 | R = 0.808 p < 0.001 | R = 0.521 p = 0.013 | R = 0.866 p < 0.001 | R = 0.842 p < 0.001 | R = 0.289 NS | R = 0.211 NS | |
| RUNX2 | R=0.895 p < 0.001 | R = 0.808 p < 0.001 | R = 0.725 p = 0.0001 | R = 0.877 p < 0.001 | R = 0.895 p < 0.001 | R = −0.085 NS | R = −0.038 NS | |
| ALP | R = 0.895 p < 0.001 | R = 0.748 p < 0.001 | R = 0.721 p = 0.0001 | R = 0.872 p < 0.001 | R = 0.758 p < 0.001 | R = −0.185 NS | R = −0.166 NS | |
| RANK | R = 0.758 p < 0.001 | R = 0.706 p = 0.0002 | R = 0.842 p < 0.001 | R = 0.389 p = 0.074 | R = 0.756 p < 0.001 | R = 0.302 NS | R = 0.211 NS | |
| NFATc1 | R = −0.185 NS | R = −0.085 NS | R = 0.289 NS | R = −0.299 NS | R = 0.140 NS | R = 0.302 NS | R = 0.699 p = 0.0006 |
| Genes | KYN | KYN/TRP | cAMP | ALP | TRAP−5b |
|---|---|---|---|---|---|
| ATF4 | R = 0.465 p = 0.049 | R = 0.383 p = 0.086 | R = −0.687 p = 0.002 | R = −0.461 p = 0.031 | R = −0.597 p = 0.004 |
| FOXO1 | R = 0.165 NS | R = 0.452 p = 0.046 | R = −0.275 NS | R = −0.199 NS | R = −0.336 NS |
| Cyclin D1 | R = 0.649 p = 0.002 | R = 0.615 p = 0.004 | R = −0.584 p = 0.008 | R = −0.435 p = 0.048 | R = −0.432 p = 0.057 |
| Cyclin E1 | R = 0.206 NS | R = 0.427 p = 0.060 | R = −0.416 p = 0.076 | R = −0.119 NS | R = −0.349 NS |
| RUNX2 | R = 0.357 NS | R = 0.411 p = 0.071 | R = −0.540 p = 0.017 | R = −0.358 NS | R = −0.444 p = 0.049 |
| ALP | R = 0.367 NS | R = 0.493 p = 0.027 | R = −0.635 p = 0.003 | R = −0.336 NS | R = −0.480 p = 0.032 |
| Trabecular Bone | Cortical Bone | |||||||
|---|---|---|---|---|---|---|---|---|
| KYN | ALP | TRAP−5b | TRAP−5b/ALP | KYN | ALP | TRAP−5b | TRAP−5b/ALP | |
| stiffness | R = 0.458 p = 0.032 | R = −0.690 p = 0.0002 | R = −0.714 p = 0.0003 | R = −0.534 p = 0.013 | R = −0.752 p < 0.0001 | R = −0.005 NS | R = 0.591 p = 0.004 | R = 0.518 p = 0.014 |
| Fy | R = −0.155 NS | R = 0.089 NS | R = 0.192 NS | R = 0.109 NS | R = −0.755 p < 0.0001 | R = −0.132 NS | R = 0.525 p = 0.012 | R = 0.570 p = 0.005 |
| dl (Fy) | R = −0.499 p = 0.018 | R = 0.655 p = 0.007 | R = 0.694 p = 0.0005 | R = 0.499 p = 0.021 | R = −0.467 p = 0.028 | R = −0.185 NS | R = 0.418 p = 0.052 | R = 0.514 p = 0.014 |
| Fu | R = 0.432 p = 0.044 | R = −0.438 p = 0.037 | R = −0.475 p = 0.029 | R = −0.394 p = 0.078 | R = −0.543 p = 0.009 | R = −0.078 NS | R = 0.527 p = 0.012 | R = 0.598 p = 0.003 |
| dl (Fu) | R = −0.004 NS | R = 0.279 NS | R = 0.364 NS | R = 0.242 NS | R = 0.616 p = 0.002 | R = 0.087 NS | R = −0.531 p = 0.011 | R = −0.404 p = 0.062 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pawlak, K.; Sieklucka, B.; Pawlak, D. Paracrine Kynurenic Pathway Activation in the Bone of Young Uremic Rats Can Antagonize Anabolic Effects of PTH on Bone Turnover and Strength through the Disruption of PTH-Dependent Molecular Signaling. Int. J. Mol. Sci. 2021, 22, 6563. https://doi.org/10.3390/ijms22126563
Pawlak K, Sieklucka B, Pawlak D. Paracrine Kynurenic Pathway Activation in the Bone of Young Uremic Rats Can Antagonize Anabolic Effects of PTH on Bone Turnover and Strength through the Disruption of PTH-Dependent Molecular Signaling. International Journal of Molecular Sciences. 2021; 22(12):6563. https://doi.org/10.3390/ijms22126563
Chicago/Turabian StylePawlak, Krystyna, Beata Sieklucka, and Dariusz Pawlak. 2021. "Paracrine Kynurenic Pathway Activation in the Bone of Young Uremic Rats Can Antagonize Anabolic Effects of PTH on Bone Turnover and Strength through the Disruption of PTH-Dependent Molecular Signaling" International Journal of Molecular Sciences 22, no. 12: 6563. https://doi.org/10.3390/ijms22126563
APA StylePawlak, K., Sieklucka, B., & Pawlak, D. (2021). Paracrine Kynurenic Pathway Activation in the Bone of Young Uremic Rats Can Antagonize Anabolic Effects of PTH on Bone Turnover and Strength through the Disruption of PTH-Dependent Molecular Signaling. International Journal of Molecular Sciences, 22(12), 6563. https://doi.org/10.3390/ijms22126563