
One Hundred Candidate Genes and Their Roles in Drought and Salt Tolerance in Wheat




Abstract
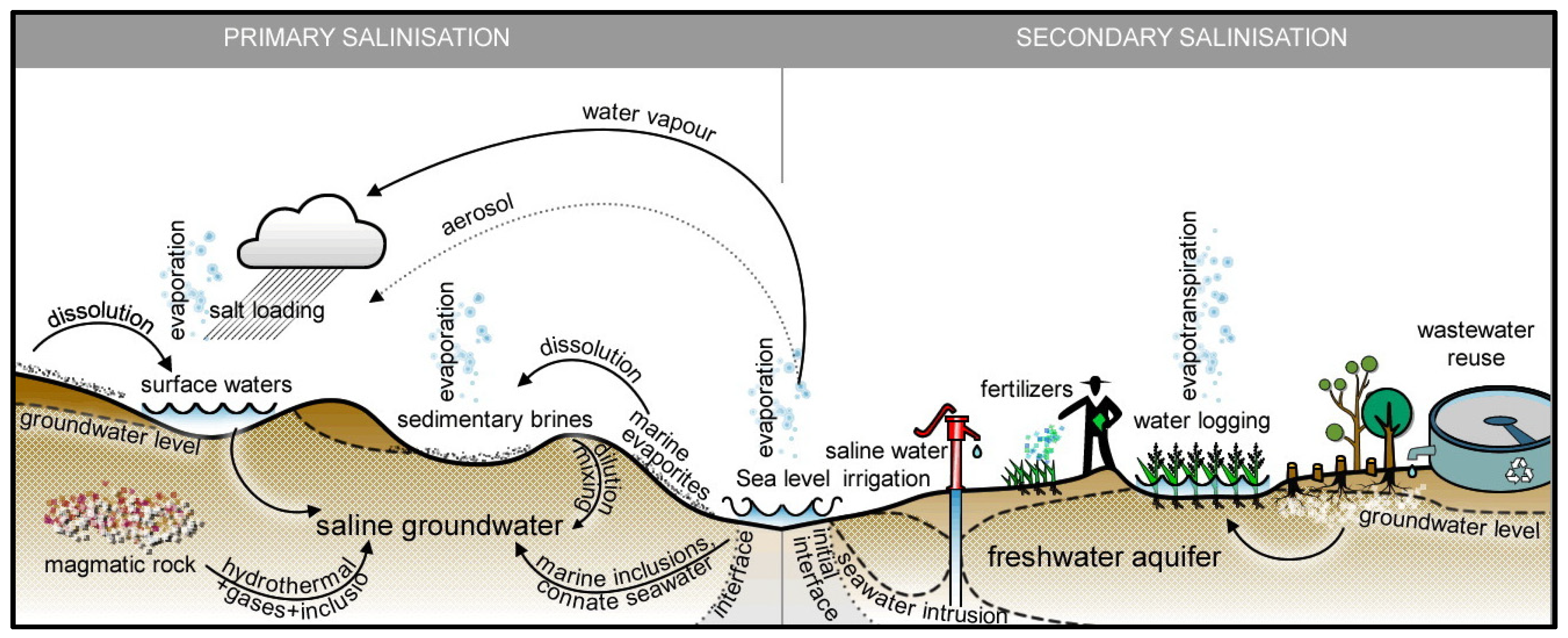
1. Global Situation
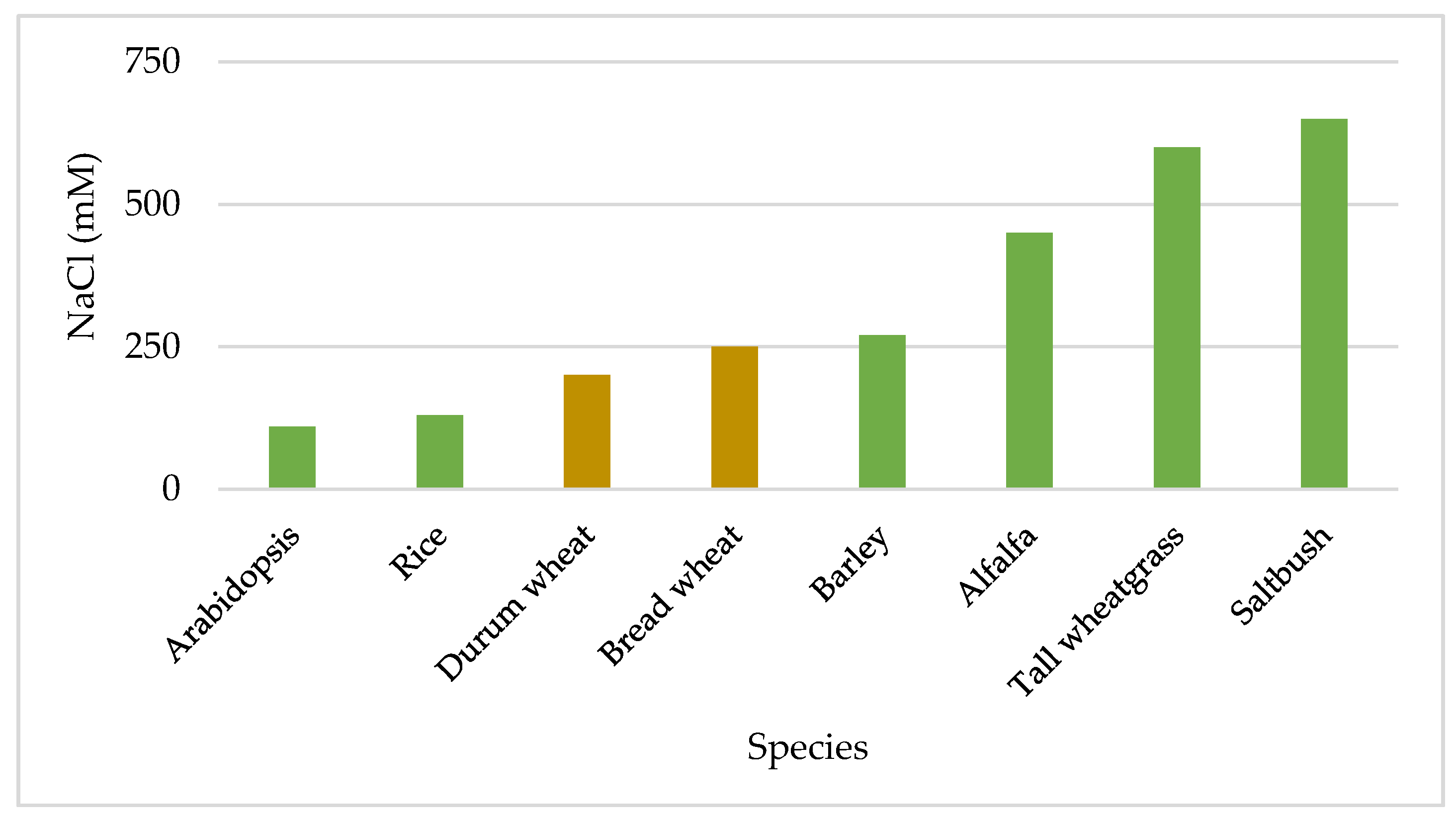
2. Germplasm
2.1. Sources of Resistances in Cultivated Species
2.2. Sources of Resistances in Alien Species

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Genome | Common Name | Reference |
|---|---|---|---|
| Triticum monococcum L. ssp. aegilopoides (Link) Thell. | AA | Wild einkorn | [22,42] |
| Triticum monococcum L. ssp. monococcum | AA | Einkorn | [43] |
| Triticum urartu Thumanjan ex Gandilyan | AA | [42,44,45] | |
| Triticum turgidum L. ssp. dicoccoides (Körn. Ex Asch. and Graebn.) Thell. | AABB | Wild emmer | [33,40,44,45,46] |
| Triticum turgidum ssp. durum L. (Desf.) Husn | AABB | Durum wheat | [22,47] |
| Triticum aestivum L. ssp. aestivum | AABBDD | Bread wheat | [48,49,50] |
| Aegilops markgrafii (Greuter) K. Hammer | CC | [34] | |
| Aegilops cylindrica Host | CCDD | Jointed goat grass | [34,51,52] |
| Aegilops triuncialis L. | CuCuCC | Barb goat grass | [51] |
| Aegilops tauschii Coss. | DD | Goat grass | [34,36,45,53] |
| Elytrigia elongata Host Nevski | EbEb | Tall wheatgrass | [54,55,56,57] |
| Thinopyrum ponticum (Podp.) Barkworth and DR Dewey | EEEEEEEEEE | [55,58,59] | |
| Thinopyrum bessarabicum (Savul and Rayss) Á. Löve | EjEj | Tall wheatgrass | [60] |
| Triticum timopheevii (Zhuk.) Zhuk. ssp. armeniacum (Jakubz.) Slageren | GGAA | [35] | |
| Triticum timopheevii (Zhuk.) Zhuk. ssp. timopheevii | GGAA | ||
| Aegilops bicornis (Forssk.) Jaub. and Spach | SbSb | [34,51] | |
| Aegilops sharonensis Eig | SjSj | [34,51] | |
| Aegilops longissima Schweinf. and Muschl. | SjSj | [34,51] | |
| Aegilops speltoides Tausch var. speltoides | SS | [45] | |
| Aegilops searsii Feldman and Kislev ex K. Hammer | SSSS | [34] | |
| Aegilops umbellulata Zhuk. | UU | Jointed goat grass | [34,51] |
| Aegilops biuncialis Vis. | UUMM | [34] | |
| Aegilops ovata auct. | UUMM | Ovate goat grass | [34,51] |
| Aegilops variabilis Eig | UUSS | [34,51] | |
| Thinopyrum junceiforme (Á. Löve and D. Löve) Á. Löve | J1J1J2J2 | [61] | |
| Thinopyrum scirpeum (K Presl) DR Dewey | JJJJ | [61] | |
| Thinopyrum junceum L. (Á. Löve) | JJJJEE | Sand couch Sea wheatgrass | [62] |
| Aegilops comosa Sm. | MM | [34] |
3. Morphological and Physiological Response
4. Mechanisms of Tolerance
4.1. Osmotic Adjustment
- The proline content increases under salt stress at the intracellular level and acts as a reserve of organic nitrogen during the stress period. Deivanai et al. [87] highlighted how rice treated with proline improves its response under salt stress.
- Glycine betaine, known also as trimethyl glycine (TMG), is a quaternary ammonium compound with three methyl groups derived from glycine found in many plants and microbes. The TMG is electrically neutral on a wide range of pH and highly water-soluble, but it also contains groups of non-polar methylins. Due to its unique structural characteristics, it interacts with both hydrophobic domains and hydrophilic macromolecules, such as enzymes and protein complexes. Glycine betaine increases the osmolarity of the cell during the period of stress [88], stabilizes the proteins [89], protects the photosynthetic apparatus from stress damage [90], and then plays an important role in stress mitigation [91].
- Sugars: Plants under saline stress tend to accumulate carbohydrates that play a role in osmo-protection and energy reserves during the stress phases [92].
- Polyols are chemical compounds composed of multiple oxydrilic groups available for organic reactions. They are classified into two types: cyclical (e.g., pinitol) and acyclic (e.g., mannitol). Polyols acts as protectors or enzyme stabilizers when stress related to dehydration occurs [93].
4.2. Ionic Homeostasis
4.3. Redox Homeostasis
5. Genes and Transcription Factors Involved
5.1. Genes Involved in Hydric Stress Tolerance
5.2. Stress Tolerance-Related Genes and Functional Proteins
5.3. The Specific Genes Involved in Ionic Homeostasis
5.4. The Genes Associated with Reduce Reactive Oxygen Species (ROS)
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mann, M.E.; Gleick, P.H. Climate change and California drought in the 21st century. Proc. Natl. Acad. Sci. USA 2015, 112, 3858–3859. [Google Scholar] [CrossRef]
- Nam, W.H.; Hayes, M.J.; Svoboda, M.D.; Tadesse, T.; Wilhite, D.A. Drought hazard assessment in the context of climate change for South Korea. Agric. Water Manag. 2015, 160, 106–117. [Google Scholar] [CrossRef]
- Bot, A.; Benites, J. Drought-resistant soils: Optimization of Soil Moisture for Sustainable Plant Production. In Proceedings of the Electronic Conference ‘Drought-Resistant Soils. Optimization of Soil Moisture for Sustainable Plant Production.’ FAO Land and Water Development Division, 15 November–18 December 2004; Volume 11. [Google Scholar]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Mitchell, R.A. Crop ecosystem responses to climatic change. In Climate Change and Global Crop Productivity; Nobel, P.S., Reddy, K.R., Hodges, H.F., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 57–80. [Google Scholar]
- Huang, J.; Zhang, W.; Zuo, J.; Bi, J.; Shi, J.; Wang, X.; Chang, Z.; Huang, Z.; Yang, S.; Zhang, B.; et al. An overview of the semi-arid climate and environment research observatory over the loess plateau. Adv. Atmos. Sci. 2008, 25, 906–921. [Google Scholar] [CrossRef]
- Houghton, J.T.; Albritton, D.L.; Meira Filho, L.G.; Cubasch, U.; Dai, X.; Ding, Y.; Karl, T. Technical summary of working group 1. In Intergovernmental Panel on Climate Change (IPCC), Climate Change 2001: The Scientific Basis; Cambridge University Press: Cambridge, UK, 2001; pp. 3–45. [Google Scholar]
- Rani, G.; Kaur, J.; Kumar, A.; Yogalakshmi, K.N. Ecosystem health and dynamics: An indicator of global climate change. In Contemporary Environmental Issues and Challenges in Era of Climate Change; Springer: Singapore, 2019; pp. 1–32. [Google Scholar]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Wegulo, S.; Giesler, L.; Harveson, R.; Jackson-Ziems, T.; Liu, B.; Korus, K. 2013 Crop Production Clinic Proceedings, Impacts of Drought on Disease Development and Management. Pap. Plant Pathol. 2013, 125–127. [Google Scholar]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 153–188. [Google Scholar]
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The threat of soil salinity: A European scale review. Sci. Total Environ. 2016, 573, 727–739. [Google Scholar] [CrossRef]
- Allen, M.R.; Ingram, W.J. Constraints on future changes in climate and the hydrologic cycle. Nature 2002, 419, 228–232. [Google Scholar] [CrossRef]
- Rozema, J.; Flowers, T.J. Crops for salinized world. Science 2008, 322, 1478–1480. [Google Scholar] [CrossRef]
- Konapala, G.; Mishra, A.K.; Wada, Y.; Mann, M.E. Climate change will affect global water availability through compounding changes in seasonal precipitation and evaporation. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Rizwan, M.; Qayyum, M.F.; Ok, Y.S.; Ibrahim, M.; Riaz, M.; Arif, M.S.; Hafeez, F.; Al-Wabel, M.I.; Shahzad, A.N. Biochar soil amendment on alleviation of drought and salt stress in plants: A critical review. Environ. Sci. Pollut. Res. 2017, 24, 12700–12712. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Prasad, M.N.V. Abiotic Stress Responses in Plants: Metabolism, Productivity and Sustainability; Springer Science + Business Media, LLC: New York, NY, USA, 2012. [Google Scholar] [CrossRef]
- Lawlor, D.W. Stress metabolism: Its implication in breeding programmes. In Drought Tolerance in Winter Cereals; Srivastava, E.J.P., Porceddu, E., Acevedo, S., Eds.; Varma. Icarda: Aleppo, Syria, 1987. [Google Scholar]
- Hatfield, J.L.; Dold, C. Water-use efficiency: Advances and challenges in a changing climate. Front. Plant Sci. 2019, 10, 103. [Google Scholar] [CrossRef]
- Richards, R.A.; Passioura, J.B. Seminal Root Morphology and Water Use of Wheat I. Environmental Effects 1. Crop Sci. 1981, 21, 249–252. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A. Screening methods for salinity tolerance: A case study with tetraploid wheat. Plant Soil 2003, 253, 201–218. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef]
- Wei, B.; Jing, R.; Wang, C.; Chen, J.; Mao, X.; Chang, X.; Jia, J. Dreb1 genes in wheat (Triticum aestivum L.): Development of functional markers and gene mapping based on SNPs. Mol. Breed. 2009, 23, 13–22. [Google Scholar] [CrossRef]
- Mondini, L.; Nachit, M.M.; Pagnotta, M.A. Allelic variants in durum wheat (Triticum turgidum L. var. durum) DREB genes conferring tolerance to abiotic stresses. Mol. Genet. Genom. 2015, 290, 531–544. [Google Scholar] [CrossRef]
- Mondini, L.; Nachit, M.M.; Porceddu, E.; Pagnotta, M.A. HRM technology for the identification and characterization of INDEL and SNPs mutations in genes involved in drought and salt tolerance of durum wheat. Plant Genet. Resour. Characterisation Util. 2011, 9, 166. [Google Scholar] [CrossRef]
- Mondini, L.; Nachit, M.; Porceddu, E.; Pagnotta, M.A. Identification of SNP mutations in DREB1, HKT1, and WRKY1 genes involved in drought and salt stress tolerance in durum wheat (Triticum turgidum L. var durum). Omi. A J. Integr. Biol. 2012, 16, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Dubcovsky, J.; Santa María, G.; Epstein, E.; Luo, M.C.; Dvořák, J. Mapping of the K+/Na+ discrimination locus Kna1 in wheat. Theor. Appl. Genet. 1996, 92, 448–454. [Google Scholar] [CrossRef]
- Colmer, T.D.; Flowers, T.J.; Munns, R. Use of wild relatives to improve salt tolerance in wheat. J. Exp. Bot. 2006, 57, 1059–1078. [Google Scholar] [CrossRef]
- Nevo, E.; Chen, G. Drought and salt tolerances in wild relatives for wheat and barley improvement. Plant Cell Environ. 2010, 33, 670–685. [Google Scholar] [CrossRef] [PubMed]
- Farooq, S.; Farooq, E.A. Production of low input and stress tolerant wheat germplasm through the use of biodiversity residing in the wild relatives. Hereditas 2001, 135, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Peleg, Z.; Fahima, T.; Abbo, S.; Krugman, T.; Nevo, E.; Yakir, D.; Saranga, Y. Genetic diversity for drought resistance in wild emmer wheat and its ecogeographical associations. Plant Cell Environ. 2005, 28, 176–191. [Google Scholar] [CrossRef]
- Gorham, J. Salt tolerance in the triticeae: K/Na discrimination in Aegilops species. J. Exp. Bot. 1990, 41, 615–621. [Google Scholar] [CrossRef]
- Gorham, J. Salt tolerance in the triticeae: Ion discrimination in rye and triticale. J. Exp. Bot. 1990, 41, 609–614. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Munns, R.; Whitecross, M.I. Variation in Sodium Exclusion and Salt Tolerance in Triticum tauschii. Crop Sci. 1991, 31, 992–997. [Google Scholar] [CrossRef]
- Dvořak, J.; Noaman, M.M.; Goyal, S.; Gorham, J. Enhancement of the salt tolerance of Triticum turgidum L. by the Kna1 locus transferred from the Triticum aestivum L. chromosome 4D by homoeologous recombination. Theor. Appl. Genet. 1994, 87, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Dvořak, J.; Gorham, J. Methodology of gene transfer by homoeologous recombinatiod into Triticum turgidum: Transfer of K+/Na+ discrimination from Triticum aestivum. Genome 1992, 35, 639–646. [Google Scholar] [CrossRef]
- Nevo, E.; Gorham, J.; Beiles, A. Variation for 11Na uptake in wild emmer wheat, Triticum dicoccoides in israel: Salt tolerance resources for wheat improvement. J. Exp. Bot. 1992, 43, 511–518. [Google Scholar] [CrossRef]
- Nevo, E.; Krugman, T.; Beiles, A. Genetic Resources for Salt Tolerance in the Wild Progenitors of Wheat (Triticum dicoccoides) and Barley (Hordeum spontaneum) in Israel. Plant Breed. 1993, 110, 338–341. [Google Scholar] [CrossRef]
- Lindsay, M.P.; Lagudah, E.S.; Hare, R.A.; Munns, R. A locus for sodium exclusion (Nax1), a trait for salt tolerance, mapped in durum wheat. Funct. Plant Biol. 2004, 31, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Gorham, J.; Bristol, A.; Young, E.M.; Wyn Jones, R.G. The presence of the enhanced K/Na discrimination trait in diploid Triticum species. Theor. Appl. Genet. 1991, 82, 236–729. [Google Scholar] [CrossRef]
- Kumar, A.; Varma, S.K.; Angrish, R. Differentiation of chloride and sulphate salinity on the basis of ionic distribution in genetically diverse cultivars of wheat. J. Plant Nutr. 1995, 18, 2199–2212. [Google Scholar] [CrossRef]
- Kara, Y.; Martin, A.; Souyris, I.; Rekika, D.; Monneveux, P. Root characteristics in durum wheat (T. turgidum conv. durum) and some wild Triticeae species. Genetic variation and relationship with plant architecture. Cereal Res. Commun. 2000, 28, 247–254. [Google Scholar] [CrossRef]
- Valkoun, J.J. Wheat pre-breeding using wild progenitors. Euphytica 2001, 119, 17–23. [Google Scholar] [CrossRef]
- Peleg, Z.; Fahima, T.; Krugman, T.; Abbo, S.; Yakir, D.; Korol, A.B.; Saranga, Y. Genomic dissection of drought resistance in durum wheat × wild emmer wheat recombinant inbreed line population. Plant Cell Environ. 2009, 32, 758–779. [Google Scholar] [CrossRef]
- Forster, B.P.; Gorham, J.; Miller, T.E. Salt Tolerance of an Amphiploid between Triticum aestivum and Agropyron junceum. Plant Breed. 1987, 98, 1–8. [Google Scholar] [CrossRef]
- Islam, S.; Malik, A.I.; Islam, A.K.M.R.; Colmer, T.D. Salt tolerance in a Hordeum marinum-Triticum aestivum amphiploid, and its parents. J. Exp. Bot. 2007, 58, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Kingsbury, R.W.; Epstein, E. Selection for Salt-Resistant Spring Wheat1. Crop Sci. 1984, 24, 310–315. [Google Scholar] [CrossRef]
- Slafer, G.A.; Araus, J.L.; Royo, C.; García Del Moral, L.F. Promising eco-physiological traits for genetic improvement of cereal yields in Mediterranean environments. Ann. Appl. Biol. 2005, 146, 61–70. [Google Scholar] [CrossRef]
- Farooq, S.; Niazi, M.L.K.; Iqbal, N.; Shah, T.M. Salt tolerance potential of wild resources of the tribe Triticeae-II. Screening of species of the genus Aegilops. Plant Soil 1989, 119, 255–260. [Google Scholar] [CrossRef]
- Farooq, S.; Iqbal, N.; Asghar, M.; Shah, T.M. Intergeneric hybridization for wheat improvement-IV: Expression of salt tolerance gene(s) of Aegilops cylindrica in hybrids with hexaploid wheat. Cereal Res. Commun. 1992, 20, 111–118. [Google Scholar]
- Mujeeb-Kazi, A.; Rosas, V.; Roldan, S. Conservation of the genetic variation of Triticum tauschii (Coss.) Schmalh. (Aegilops squarrosa auct. non L.) in synthetic hexaploid wheats (T. turgidum L. s.lat. × T. tauschii; 2n = 6x = 42, AABBDD) and its potential utilization for wheat improvement. Genet. Resour. Crop Evol. 1996, 43, 129–134. [Google Scholar] [CrossRef]
- Dvořák, J.; Ross, K. Expression of Tolerance of Na+, K+, Mg 2+, Cl− and SO2− 4 Ions and Sea Water in the Amphiploid of Triticum aestivum × Elytrigia elongata 1. Crop Sci. 1986, 26, 658–660. [Google Scholar] [CrossRef]
- McGuire, P.E.; Dvôrák, J. High Salt-Tolerance Potential in Wheatgrasses1. Crop Sci. 1981, 21, 702–705. [Google Scholar] [CrossRef]
- Omielan, J.A.; Epstein, E.; Dvorak, J. Salt tolerance and ionic relations of wheat as affected by individual chromosomes of salt-tolerant Lophopyrum elongatum. Genome 1991, 34, 961–974. [Google Scholar] [CrossRef]
- Storey, R.; Graham, R.D.; Shepherd, K.W. Modification of the salinity response of wheat by the genome of Elytrigia elongatum. Plant Soil 1985, 83, 327–330. [Google Scholar] [CrossRef]
- Suiyun, C.; Guangmin, X.; Taiyong, Q.; Fengnin, X.; Yan, J.; Huimin, C. Introgression of salt-tolerance from somatic hybrids between common wheat and Thinopyrum ponticum. Plant Sci. 2004, 167, 773–779. [Google Scholar] [CrossRef]
- Xia, G.; Xiang, F.; Zhou, A.; Wang, H.; Chen, H. Asymmetric somatic hybridization between wheat (Triticum aestivum L.) and Agropyron elongatum (Host) Nevishi. Theor. Appl. Genet. 2003, 107, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Gorham, J.; Mcdonnell, E.; Budrewicz, E.; Jones, R.G.W. Salt tolerance in the Triticeae: Growth and solute accumulation in leaves of Thinopyrum bessarabicum. J. Exp. Bot. 1985, 36, 1021–1031. [Google Scholar] [CrossRef]
- Gorham, J.; Budrewicz, E.; Mcdonnell, E.; Jones, R.G.W. Salt tolerance in the triticeae: Salinity-induced changes in the leaf solute composition of some perennial Triticeae. J. Exp. Bot. 1986, 37, 1114–1128. [Google Scholar] [CrossRef]
- Wang, R.R.C.; Li, X.M.; Hu, Z.M.; Zhang, J.Y.; Larson, S.R.; Zhang, X.Y.; Grieve, C.M.; Shannon, M.C. Development of salinity-tolerant wheat recombinant lines from a wheat disomic addition line carrying a Thinopyrum junceum chromosome. Int. J. Plant Sci. 2003, 164, 25–33. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Springer: Cham, Switzerland, 2009; pp. 153–188. ISBN 9789048126651. [Google Scholar]
- Rawson, H.M.; Richards, R.A.; Munns, R. An examination of selection criteria for salt tolerance in wheat, barley and Triticale genotypes. Aust. J. Agric. Res. 1988, 39, 759–772. [Google Scholar] [CrossRef]
- Thiyagarajan, K.; Latini, A.; Cantale, C.; Galeffi, P. Structural characterization of the DRF1 gene of Aegilops speltoides and comparison of its sequence with those of B and other Triticeae genomes. Euphytica 2020, 216, 1–17. [Google Scholar] [CrossRef]
- Zhong, G.-Y.; Dvořák, J. Evidence for common genetic mechanisms controlling the tolerance of sudden salt stress in the tribe Triticeae. Plant Breed. 1995, 114, 297–302. [Google Scholar] [CrossRef]
- Qin, P.; Lin, Y.; Hu, Y.; Liu, K.; Mao, S.; Li, Z.; Wang, J.; Liu, Y.; Wei, Y.; Zheng, Y. Genome-wide association study of drought-related resistance traits in Aegilops tauschii. Genet. Mol. Biol. 2016, 39, 398–407. [Google Scholar] [CrossRef]
- Gill, R.S.; Dhaliwal, H.S.; Multani, D.S. Synthesis and evaluation of Triticum durum-T. monococcum amphiploids. Theor. Appl. Genet. 1988, 75, 912–916. [Google Scholar] [CrossRef]
- Limin, A.E.; Fowler, D.B. Cold hardiness expression in interspecific hybrids and amphiploids of the Triticeae. Genome 1988, 30, 361–365. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Kurishbayev, A.; Jatayev, S.; Shvidchenko, V.; Zotova, L.; Koekemoer, F.; De Groot, S.; Soole, K.; Langridge, P. Early flowering as a drought escape mechanism in plants: How can it aid wheat production? Front. Plant Sci. 2017, 8, 1950. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; García-Sánchez, F.; Pérez-Pérez, J.G.; Colmenero-Flores, J.M.; Rubio, F.; Rosales, M.A. Coping With Water Shortage: An Update on the Role of K+, Cl-, and Water Membrane Transport Mechanisms on Drought Resistance. Front. Plant Sci. 2019, 10, 1619. [Google Scholar] [CrossRef]
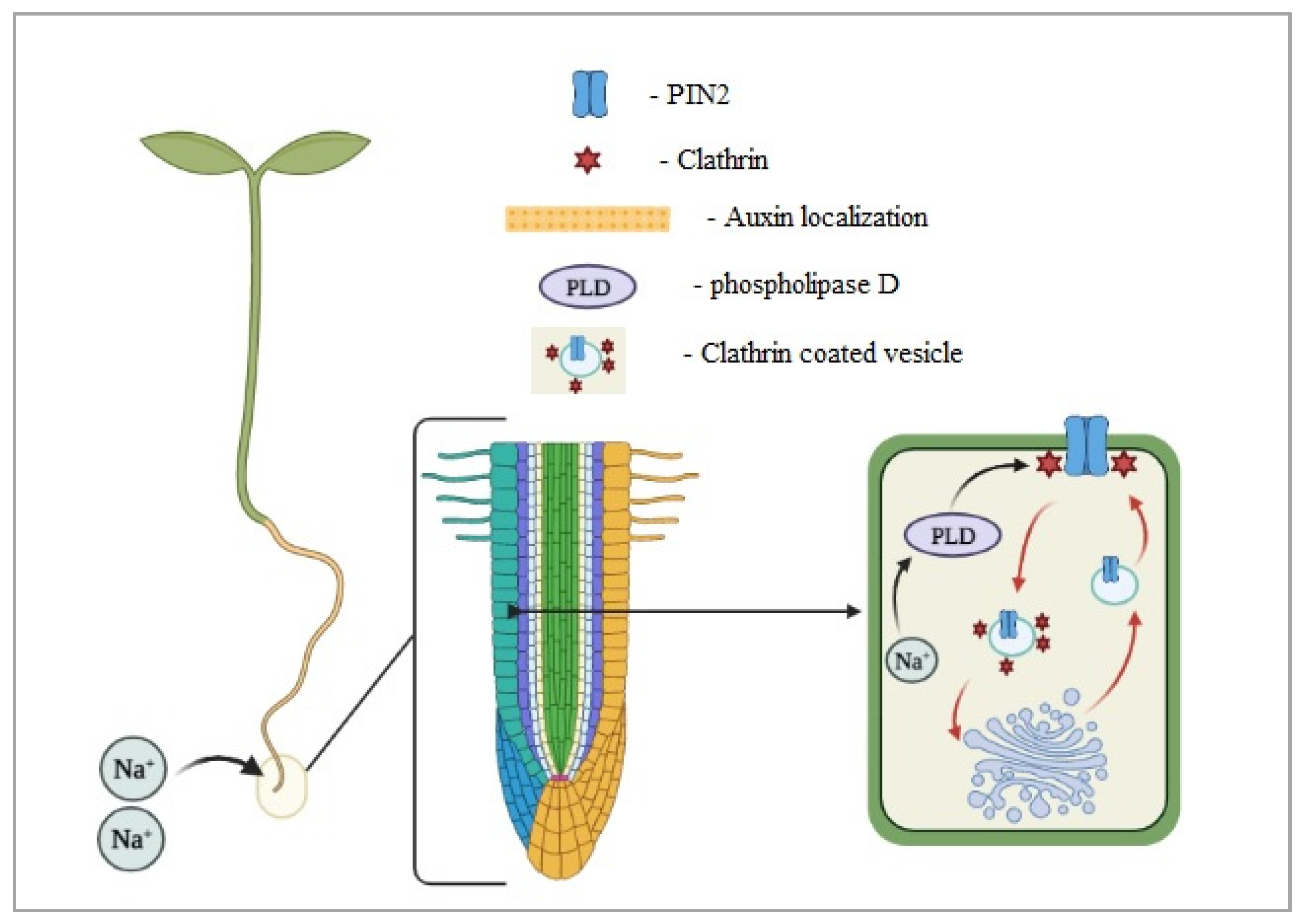
- Galvan-Ampudia, C.S.; Julkowska, M.M.; Darwish, E.; Gandullo, J.; Korver, R.A.; Brunoud, G.; Haring, M.A.; Munnik, T.; Vernoux, T.; Testerink, C. Halotropism is a response of plant roots to avoid a saline environment. Curr. Biol. 2013, 23, 2044–2050. [Google Scholar] [CrossRef]
- Dai, M.; Zhang, C.; Kania, U.; Chen, F.; Xue, Q.; McCray, T.; Li, G.; Qin, G.; Wakeley, M.; Terzaghi, W.; et al. A PP6-type phosphatase holoenzyme directly regulates PIN phosphorylation and auxin efflux in Arabidopsis. Plant Cell 2012, 24, 2497–2514. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zhang, W.; Hu, H.; Li, B.; Wang, Y.; Zhao, Y.; Li, K.; Liu, M.; Li, X. Salt modulates gravity signaling pathway to regulate growth direction of primary roots in arabidopsis. Plant Physiol. 2008, 146, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Julkowska, M.M.; Koevoets, I.T.; Mol, S.; Hoefsloot, H.; Feron, R.; Tester, M.A.; Keurentjes, J.J.B.; Korte, A.; Haring, M.A.; De Boer, G.J.; et al. Genetic components of root architecture remodeling in response to salt stress. Plant Cell 2017, 29, 3198–3213. [Google Scholar] [CrossRef] [PubMed]
- Mondini, L.; Pagnotta, M.A. Drought and Salt Stress in Cereals; Springer: Cham, Switzerland, 2015; pp. 1–31. [Google Scholar]
- Sun, T.; Wang, Y.; Wang, M.; Li, T.; Zhou, Y.; Wang, X.; Wei, S.; He, G.; Yang, G. Identification and comprehensive analyses of the CBL and CIPK gene families in wheat (Triticum aestivum L.). BMC Plant Biol. 2015, 15, 1–17. [Google Scholar] [CrossRef]
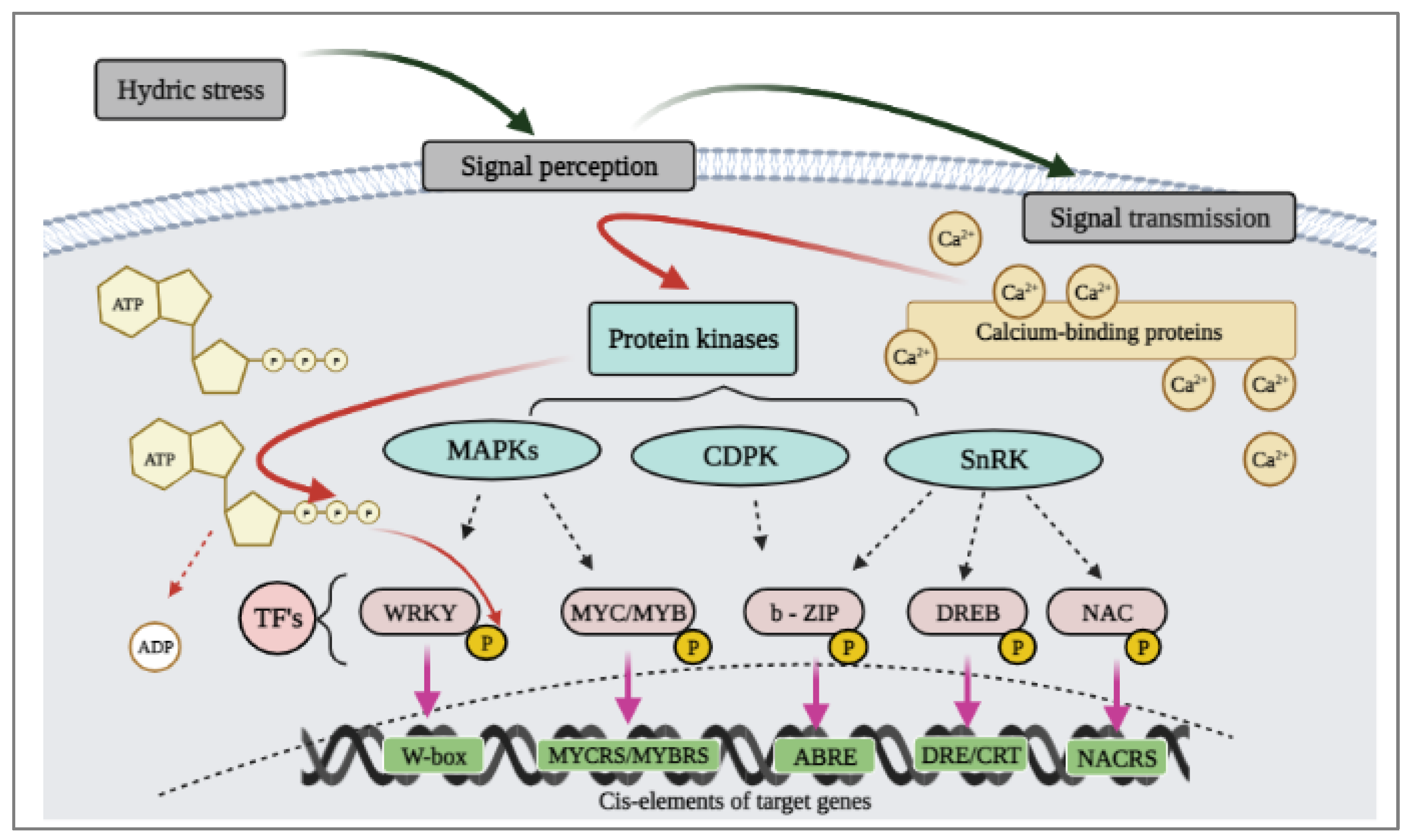
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. CRC. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef]
- Faïc Xal Brini, F.; Hanin, M.; Mezghani, I.; Berkowitz, G.A.; Masmoudi, K. Overexpression of wheat Na+/H+ antiporter TNHX1 and H+-pyrophosphatase TVP1 improve salt-and drought-stress tolerance in Arabidopsis thaliana plants. J. Exp. Bot. 2007, 58, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, A.; Niazi, A.; Abolimoghadam, A.A.; Zamani Babgohari, M.; Deihimi, T.; Ebrahimi, M.; Akhtardanesh, H.; Ebrahimie, E. Quantitative expression analysis of TaSOS1 and TaSOS4 genes in cultivated and wild wheat plants under salt stress. Mol. Biotechnol. 2013, 53, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.M.; Zhang, H.M.; Liu, Z.H.; Li, H.C.; Guo, X.L.; Li, G.L. The wheat NHX antiporter gene TaNHX2 confers salt tolerance in transgenic alfalfa by increasing the retention capacity of intracellular potassium. Plant Mol. Biol. 2015, 87, 317–327. [Google Scholar] [CrossRef]
- Tavakoli, M.; Poustini, K.; Alizadeh, H. Proline accumulation and related genes in wheat leaves under salinity stress. J. Agric. Sci. Technol. 2016, 18, 707–716. [Google Scholar]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef]
- Blum, A. Plant Breeding for Water-Limited Environments; Springer: New York, NY, USA, 2011; pp. 27–28. ISBN 9781441974914. [Google Scholar]
- Deivanai, S.; Xavier, R.; Vinod, V.; Timalata, K.; Lim, O.F. Role of Exogenous Proline in Ameliorating Salt Stress at Early Stage in Two Rice Cultivars. J. Stress Physiol. Biochem. 2011, 7, 157–174. [Google Scholar]
- Gadallah, M.A.A. Effects of proline and glycinebetaine on Vicia faba responses to salt stress. Biol. Plant. 1999, 42, 249–257. [Google Scholar] [CrossRef]
- Mäkelä, P.; Karkkainen, J.; Somersalo, S. Effect of glycinebetaine on chloroplast ultrastructure, chlorophyll and protein content, and RuBPCO activities in tomato grown under drought or salinity. Biol. Plant. 2000, 43, 471–475. [Google Scholar] [CrossRef]
- Cha-Um, S.; Kirdmanee, C. Effect of glycinebetaine on proline, water use, and photosynthetic efficiencies, and growth of rice seedlings under salt stress. Turkish J. Agric. For. 2010, 34, 517–527. [Google Scholar] [CrossRef]
- Sakamoto, A.; Murata, N. The role of glycine betaine in the protection of plants from stress: Clues from transgenic plants. Plant Cell Environ. 2002, 25, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Lim, C.J.; Kwon, S.Y. Glycine betaine: A versatile compound with great potential for gene pyramiding to improve crop plant performance against environmental stresses. Plant Biotechnol. Rep. 2013, 7, 49–57. [Google Scholar] [CrossRef]
- Bohnert, H.J.; Nelson, D.E.; Jensen, R.G. Adaptations to environmental stresses. Plant Cell 1995, 7, 1099. [Google Scholar] [CrossRef] [PubMed]
- Tisi, A.; Angelini, R.; Cona, A. Wound healing in plants: Cooperation of copper amine oxidase and flavin-containing polyamine oxidase. Plant Signal. Behav. 2008, 3, 204–206. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Shi, H.; Xiong, L.; Stevenson, B.; Lu, T.; Zhu, J.K. The Arabidopsis salt overly sensitive 4 mutants uncover a critical role for vitamin B6 in plant salt tolerance. Plant Cell 2002, 14, 575–588. [Google Scholar] [CrossRef]
- Liu, W.; Schachtman, D.P.; Zhang, W. Partial deletion of a loop region in the high affinity K+ transporter HKT1 changes ionic permeability leading to increased salt tolerance. J. Biol. Chem. 2000, 275, 27924–27932. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, J.K. A calcium sensor homolog required for plant salt tolerance. Science 1998, 280, 1943–1945. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Comparative physiology of salt and water stress. Plant. Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Rodríguez, H.G.; Roberts, J.K.M.; Jordan, W.R.; Drew, M.C. Growth, water relations, and accumulation of organic and inorganic solutes in roots of maize seedlings during salt stress. Plant Physiol. 1997, 113, 881–893. [Google Scholar] [CrossRef]
- Groß, F.; Durner, J.; Gaupels, F. Nitric oxide, antioxidants and prooxidants in plant defence responses. Front. Plant Sci. 2013, 4, 419. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 167, 645–663. [Google Scholar] [CrossRef]
- Zhang, S.J.; Song, G.Q.; Li, Y.L.; Gao, J.; Liu, J.J.; Fan, Q.Q.; Huang, C.Y.; Sui, X.X.; Chu, X.S.; Guo, D.; et al. Cloning of 9-cis-epoxycarotenoid dioxygenase gene (TaNCED1) from wheat and its heterologous expression in tobacco. Biol. Plant. 2014, 58, 89–98. [Google Scholar] [CrossRef]
- Nambara, E.; Marion-Poll, A. Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef]
- Laxalt, A.M.; Munnik, T. Phospholipid signalling in plant defence. Curr. Opin. Plant Biol. 2002, 5, 332–338. [Google Scholar] [CrossRef]
- Canonne, J.; Froidure-Nicolas, S.; Rivas, S. Phospholipases in action during plant defense signaling. Plant Signal. Behav. 2011, 6, 13–18. [Google Scholar] [CrossRef]
- Pokotylo, I.; Kolesnikov, Y.; Kravets, V.; Zachowski, A.; Ruelland, E. Plant phosphoinositide-dependent phospholipases C: Variations around a canonical theme. Biochimie 2014, 96, 144–157. [Google Scholar] [CrossRef] [PubMed]
- Gillaspy, G.E. The cellular language of myo-inositol signaling. New Phytologist. 2011, 192, 823–839. [Google Scholar] [CrossRef] [PubMed]
- Munnik, T.; Vermeer, J.E.M. Osmotic stress-induced phosphoinositide and inositol phosphate signalling in plants. Plant Cell Environ. 2010, 33, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Jin, C.; Wu, L.; Hou, M.; Dou, S.; Pan, Y. Expression analysis of a stress-related phosphoinositide-specific phospholipase c gene in wheat (Triticum aestivum L.). PLoS ONE 2014, 9, e105061. [Google Scholar] [CrossRef]
- Aggarwal, S.; Shukla, V.; Bhati, K.K.; Kaur, M.; Sharma, S.; Singh, A.; Mantri, S.; Pandey, A.K. Hormonal regulation and expression profiles of wheat genes involved during phytic acid biosynthesis pathway. Plants 2015, 4, 298–319. [Google Scholar] [CrossRef]
- Mills, L.N.; Hunt, L.; Leckie, C.P.; Aitken, F.L.; Wentworth, M.; McAinsh, M.R.; Gray, J.E.; Hetherington, A.M. The effects of manipulating phospholipase C on guard cell ABA-signalling. J. Exp. Bot. 2004, 55, 199–204. [Google Scholar] [CrossRef][Green Version]
- Zhao, Y.; Dong, W.; Zhang, N.; Ai, X.; Wang, M.; Huang, Z.; Xiao, L.; Xia, G. A wheat allene oxide cyclase gene enhances salinity tolerance via jasmonate signaling. Plant Physiol. 2014, 164, 1068–1076. [Google Scholar] [CrossRef]
- Batistič, O.; Kudla, J. Analysis of calcium signaling pathways in plants. Biochim. Biophys. Acta-Gen. Subj. 2012, 1820, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Wang, X.; Sun, Y.; Wang, X.; Chen, X.; Guo, J.; Duan, Y.; Huang, L.; Kang, Z. Cloning and characterization of a calcium binding EF-hand protein gene TaCab1 from wheat and its expression in response to Puccinia striiformis f. sp. tritici and abiotic stresses. Mol. Biol. Rep. 2011, 38, 3857–3866. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Hai Lu, Y.; Song, M.; Wang, Y.; Xu, W.; Wu, L.; Wang, H.; Ma, Z. Overexpression of a triticum aestivum calreticulin gene (TaCRT1) improves salinity tolerance in tobacco. PLoS ONE 2015, 10, e0140591. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.Y.; Xu, C.Y.; Jing, R.L.; Li, R.Z.; Mao, X.G.; Wang, J.P.; Chang, X.P. Molecular cloning and characterization of wheat calreticulin (CRT) gene involved in drought-stressed responses. J. Exp. Bot. 2008, 59, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.S.; Liu, L.; Ni, Z.Y.; Liu, P.; Chen, M.; Li, L.C.; Chen, Y.F.; Ma, Y.Z. W55a encodes a novel protein kinase that is involved in multiple stress responses. J. Integr. Plant Biol. 2009, 51, 58–66. [Google Scholar] [CrossRef]
- McLoughlin, F.; Galvan-Ampudia, C.S.; Julkowska, M.M.; Caarls, L.; Van Der Does, D.; Laurière, C.; Munnik, T.; Haring, M.A.; Testerink, C. The Snf1-related protein kinases SnRK2.4 and SnRK2.10 are involved in maintenance of root system architecture during salt stress. Plant J. 2012, 72, 436–449. [Google Scholar] [CrossRef]
- Zhang, H.; Mao, X.; Wang, C.; Jing, R. Overexpression of a common wheat gene Tasnrk2.8 enhances tolerance to drought, salt and low temperature in Arabidopsis. PLoS ONE 2010, 5, e16041. [Google Scholar] [CrossRef]
- Christov, N.K.; Christova, P.K.; Kato, H.; Liu, Y.; Sasaki, K.; Imai, R. TaSK5, an abiotic stress-inducible GSK3/shaggy-like kinase from wheat, confers salt and drought tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2014, 84, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.P.; Ma, W.S.; Huang, Z.J.; Xu, T.; Xue, Y.B.; Shen, Y.Z. Isolation and characterization of TaGSK1 involved in wheat salt tolerance. Plant Sci. 2003, 165, 1369–1375. [Google Scholar] [CrossRef]
- Huang, L.; Yasir, T.A.; Phillips, A.L.; Hu, Y.G. Isolation and characterization of ERECTA genes and their expression patterns in common wheat (Triticum aestivum L.). Aust. J. Crop Sci. 2013, 7, 381–390. [Google Scholar]
- Wang, C.; Jing, R.; Mao, X.; Chang, X.; Li, A. TaABC1, a member of the activity of bc 1 complex protein kinase family from common wheat, confers enhanced tolerance to abiotic stresses in Arabidopsis. J. Exp. Bot. 2011, 62, 1299–1311. [Google Scholar] [CrossRef]
- Gahlaut, V.; Jaiswal, V.; Kumar, A.; Gupta, P.K. Transcription factors involved in drought tolerance and their possible role in developing drought tolerant cultivars with emphasis on wheat (Triticum aestivum L.). Theor. Appl. Genet. 2016, 129, 2019–2042. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Sun, F.Y.; Ji, C.; Hu, Q.W.; Wang, C.Y.; Wu, D.X.; Sun, G. Nucleotide diversity patterns at the DREB1 transcriptional factor gene in the genome donor species of wheat (Triticum aestivum L). PLoS ONE 2019, 14, e0217081. [Google Scholar] [CrossRef]
- Egawa, C.; Kobayashi, F.; Ishibashi, M.; Nakamura, T.; Nakamura, C.; Takumi, S. Differential regulation of transcript accumulation and alternative splicing of a DREB2 homolog under abiotic stress conditions in common wheat. Genes Genet. Syst. 2006, 81, 77–91. [Google Scholar] [CrossRef]
- Pellegrineschi, A.; Reynolds, M.; Pacheco, M.; Brito, R.M.; Almeraya, R.; Yamaguchi-Shinozaki, K.; Hoisington, D. Stress-induced expression in wheat of the Arabidopsis thaliana DREB1A gene delays water stress symptoms under greenhouse conditions. Genome 2004, 47, 493–500. [Google Scholar] [CrossRef]
- Wang, J.; Li, Q.; Mao, X.; Li, A.; Jing, R. Wheat transcription factor TaAREB3 participates in drought and freezing tolerances in Arabidopsis. Int. J. Biol. Sci. 2016, 12, 257. [Google Scholar] [CrossRef]
- Kobayashi, F.; Maeta, E.; Terashima, A.; Takumi, S. Positive role of a wheat HvABI5 ortholog in abiotic stress response of seedlings. Physiol. Plant. 2008, 134, 74–86. [Google Scholar] [CrossRef]
- Kobayashi, F.; Maeta, E.; Terashima, A.; Kawaura, K.; Ogihara, Y.; Takumi, S. Development of abiotic stress tolerance via bZIP-type transcription factor LIP19 in common wheat. J. Exp. Bot. 2008, 59, 891–905. [Google Scholar] [CrossRef]
- Xu, D.B.; Gao, S.Q.; Ma, Y.Z.; Xu, Z.S.; Zhao, C.P.; Tang, Y.M.; Li, X.Y.; Li, L.C.; Chen, Y.F.; Chen, M. ABI-like transcription factor gene TaABL1 from wheat improves multiple abiotic stress tolerances in transgenic plants. Funct. Integr. Genom. 2014, 14, 717–730. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, G.; Zhao, G.; Xia, C.; Jia, J.; Liu, X.; Kong, X. Characterization of a wheat R2R3-MYB transcription factor gene, TaMYB19, involved in enhanced abiotic stresses in Arabidopsis. Plant Cell Physiol. 2014, 55, 1802–1812. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, G.; Xia, C.; Jia, J.; Liu, X.; Kong, X. Overexpression of a wheat MYB transcription factor gene, TaMYB56-B, enhances tolerances to freezing and salt stresses in transgenic Arabidopsis. Gene 2012, 505, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Rahaie, M.; Xue, G.P.; Naghavi, M.R.; Alizadeh, H.; Schenk, P.M. A MYB gene from wheat (Triticum aestivum L.) is up-regulated during salt and drought stresses and differentially regulated between salt-tolerant and sensitive genotypes. Plant Cell Rep. 2010, 29, 835–844. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, G.; Jia, J.; Liu, X.; Kong, X. Molecular characterization of 60 isolated wheat MYB genes and analysis of their expression during abiotic stress. J. Exp. Bot. 2012, 63, 203–214. [Google Scholar] [CrossRef]
- Qin, Y.; Wang, M.; Tian, Y.; He, W.; Han, L.; Xia, G. Over-expression of TaMYB33 encoding a novel wheat MYB transcription factor increases salt and drought tolerance in Arabidopsis. Mol. Biol. Rep. 2012, 39, 7183–7192. [Google Scholar] [CrossRef]
- He, Y.; Li, W.; Lv, J.; Jia, Y.; Wang, M.; Xia, G. Ectopic expression of a wheat MYB transcription factor gene, TaMYB73, improves salinity stress tolerance in Arabidopsis thaliana. J. Exp. Bot. 2012, 63, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Zhang, L.; Xia, C.; Fu, S.; Zhao, G.; Jia, J.; Kong, X. The wheat transcription factor, TabHLH39, improves tolerance to multiple abiotic stressors in transgenic plants. Biochem. Biophys. Res. Commun. 2016, 473, 1321–1327. [Google Scholar] [CrossRef] [PubMed]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Niu, C.F.; Wei, W.; Zhou, Q.Y.; Tian, A.G.; Hao, Y.J.; Zhang, W.K.; Ma, B.; Lin, Q.; Zhang, Z.B.; Zhang, J.S.; et al. Wheat WRKY genes TaWRKY2 and TaWRKY19 regulate abiotic stress tolerance in transgenic Arabidopsis plants. Plant Cell Environ. 2012, 35, 1156–1170. [Google Scholar] [CrossRef]
- Wang, C.; Deng, P.; Chen, L.; Wang, X.; Ma, H.; Hu, W.; Yao, N.; Feng, Y.; Chai, R.; Yang, G.; et al. A Wheat WRKY Transcription Factor TaWRKY10 Confers Tolerance to Multiple Abiotic Stresses in Transgenic Tobacco. PLoS ONE 2013, 8, e65120. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, J.; Li, Y.; Rong, X.; Sun, J.; Sun, T.; Li, M.; Wang, L.; Feng, Y.; Chai, R.; et al. Expression of TaWRKY44, a wheat WRKY gene, in transgenic tobacco confers multiple abiotic stress tolerances. Front. Plant Sci. 2015, 6, 615. [Google Scholar] [CrossRef]
- Zhou, S.; Zheng, W.J.; Liu, B.H.; Zheng, J.C.; Dong, F.S.; Liu, Z.F.; Wen, Z.Y.; Yang, F.; Wang, H.B.; Xu, Z.S.; et al. Characterizing the role of TaWRKY13 in salt tolerance. Int. J. Mol. Sci. 2019, 20, 5712. [Google Scholar] [CrossRef]
- Qin, Y.; Tian, Y.; Liu, X. A wheat salinity-induced WRKY transcription factor TaWRKY93 confers multiple abiotic stress tolerance in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2015, 464, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wang, Y.; Li, B.; Chang, J.; Chen, M.; Li, K.; Yang, G.; He, G. TaNAC29, a NAC transcription factor from wheat, enhances salt and drought tolerance in transgenic Arabidopsis. BMC Plant Biol. 2015, 15, 1–15. [Google Scholar] [CrossRef]
- Mao, X.; Zhang, H.; Qian, X.; Li, A.; Zhao, G.; Jing, R. TaNAC2, a NAC-type wheat transcription factor conferring enhanced multiple abiotic stress tolerances in Arabidopsis. J. Exp. Bot. 2012, 63, 2933–2946. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Chen, S.; Li, A.; Zhai, C.; Jing, R. Novel NAC transcription factor TaNAC67 confers enhanced multi-abiotic stress tolerances in Arabidopsis. PLoS ONE 2014, 9, e84359. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, L.; Xia, C.; Zhao, G.; Jia, J.; Kong, X. The novel wheat transcription factor TaNAC47 enhances multiple abiotic stress tolerances in transgenic plants. Front. Plant Sci. 2016, 6, 1174. [Google Scholar] [CrossRef]
- Xue, G.P.; Way, H.M.; Richardson, T.; Drenth, J.; Joyce, P.A.; McIntyre, C.L. Overexpression of TaNAC69 leads to enhanced transcript levels of stress up-regulated genes and dehydration tolerance in bread wheat. Mol. Plant 2011, 4, 697–712. [Google Scholar] [CrossRef]
- Xia, N.; Zhang, G.; Sun, Y.F.; Zhu, L.; Xu, L.S.; Chen, X.M.; Liu, B.; Yu, Y.T.; Wang, X.J.; Huang, L.L.; et al. TaNAC8, a novel NAC transcription factor gene in wheat, responds to stripe rust pathogen infection and abiotic stresses. Physiol. Mol. Plant Pathol. 2010, 74, 394–402. [Google Scholar] [CrossRef]
- Xia, N.; Zhang, G.; Liu, X.Y.; Deng, L.; Cai, G.L.; Zhang, Y.; Wang, X.J.; Zhao, J.; Huang, L.L.; Kang, Z.S. Characterization of a novel wheat NAC transcription factor gene involved in defense response against stripe rust pathogen infection and abiotic stresses. Mol. Biol. Rep. 2010, 37, 3703–3712. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, X.; Tian, Y.; Zhang, D.; Zhang, L. The functional analysis of a wheat group 3 late embryogenesis abundant protein in Escherichia coli and Arabidopsis under abiotic stresses. Plant Signal. Behav. 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Dure, L. A repeating 11-mer amino acid motif and plant desiccation. Plant J. 1993, 3, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhao, S.; Zhao, B.; Li, C. Overexpression of TaLEA3 induces rapid stomatal closure under drought stress in Phellodendron amurense Rupr. Plant Sci. 2018, 277, 100–109. [Google Scholar] [CrossRef]
- Liang, Y.; Kang, K.; Gan, L.; Ning, S.; Xiong, J.; Song, S.; Xi, L.; Lai, S.; Yin, Y.; Gu, J.; et al. Drought-responsive genes, late embryogenesis abundant group3 (LEA3) and vicinal oxygen chelate, function in lipid accumulation in Brassica napus and Arabidopsis mainly via enhancing photosynthetic efficiency and reducing ROS. Plant Biotechnol. J. 2019, 17, 2123–2142. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Chen, S.; Liu, G. Enhanced drought tolerance in transgenic Leymus chinensis plants with constitutively expressed wheat TaLEA 3. Biotechnol. Lett. 2009, 31, 313–319. [Google Scholar] [CrossRef]
- Koubaa, S.; Brini, F. Functional analysis of a wheat group 3 late embryogenesis abundant protein (TdLEA3) in Arabidopsis thaliana under abiotic and biotic stresses. Plant Physiol. Biochem. 2020, 156, 396–406. [Google Scholar] [CrossRef]
- Bhagi, P.; Zhawar, V.K.; Gupta, A.K. Antioxidant response and Lea genes expression under salt stress and combined salt plus water stress in two wheat cultivars contrasting in drought tolerance. Indian J. Exp. Biol. 2013, 51, 746–757. [Google Scholar]
- Wang, Y.; Xu, H.; Zhu, H.; Tao, Y.; Zhang, G.; Zhang, L.; Zhang, C.; Zhang, Z.; Ma, Z. Classification and expression diversification of wheat dehydrin genes. Plant Sci. 2014, 214, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Brini, F.; Hanin, M.; Lumbreras, V.; Amara, I.; Khoudi, H.; Hassairi, A.; Pagès, M.; Masmoudi, K. Overexpression of wheat dehydrin DHN-5 enhances tolerance to salt and osmotic stress in Arabidopsis thaliana. Plant Cell Rep. 2007, 26, 2017–2026. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C. Plant aquaporins: Novel functions and regulation properties. FEBS Lett. 2007, 581, 2227–2236. [Google Scholar] [CrossRef]
- Gao, Z.; He, X.; Zhao, B.; Zhou, C.; Liang, Y.; Ge, R.; Shen, Y.; Huang, Z. Overexpressing a putative aquaporin gene from wheat, TaNIP, enhances salt tolerance in transgenic Arabidopsis. Plant Cell Physiol. 2010, 51, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, M.; Zhou, L.; Quan, T.; Xia, G. Heterologous expression of the wheat aquaporin gene TaTIP2;2 compromises the abiotic stress tolerance of Arabidopsis thaliana. PLoS ONE 2013, 8, e79618. [Google Scholar] [CrossRef]
- Zhou, S.; Hu, W.; Deng, X.; Ma, Z.; Chen, L.; Huang, C.; Wang, C.; Wang, J.; He, Y.; Yang, G.; et al. Overexpression of the Wheat Aquaporin Gene, TaAQP7, Enhances Drought Tolerance in Transgenic Tobacco. PLoS ONE 2012, 7, e52439. [Google Scholar] [CrossRef]
- Hu, W.; Yuan, Q.; Wang, Y.; Cai, R.; Deng, X.; Wang, J.; Zhou, S.; Chen, M.; Chen, L.; Huang, C.; et al. Overexpression of a wheat aquaporin gene, TaAQP8, enhances salt stress tolerance in transgenic tobacco. Plant Cell Physiol. 2012, 53, 2127–2141. [Google Scholar] [CrossRef]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef]
- Yokoi, S.; Bressan, R.; Hasegawa, P. Salt stress tolerance of plants. JIRCAS Work. Rep. 2002, 23, 25–33. [Google Scholar]
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J.K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Spielmeyer, W.; Lagudah, E.S.; Munns, R. Comparative mapping of HKT genes in wheat, barley, and rice, key determinants of Na+ transport, and salt tolerance. J. Exp. Bot. 2008, 59, 927–937. [Google Scholar] [CrossRef]
- Horie, T.; Schroeder, J.I. Sodium transporters in plants. Diverse genes and physiological functions. Plant Physiol. 2004, 136, 2457–2462. [Google Scholar] [CrossRef]
- Ma, B.; Xiang, Y.; An, L. Structural bases of physiological functions and roles of the vacuolar H+-ATPase. Cell. Signal. 2011, 23, 1244–1256. [Google Scholar] [CrossRef]
- He, X.; Huang, X.; Shen, Y.; Huang, Z. Wheat V-H+-ATPase Subunit Genes Significantly Affect Salt Tolerance in Arabidopsis thaliana. PLoS ONE 2014, 9, e86982. [Google Scholar] [CrossRef] [PubMed]
- Baxter, I.; Brazelton, J.N.; Yu, D.; Huang, Y.S.; Lahner, B.; Yakubova, E.; Li, Y.; Bergelson, J.; Borevitz, J.O.; Nordborg, M.; et al. A coastal cline in sodium accumulation in Arabidopsis thaliana is driven by natural variation of the sodium transporter AtHKT1;1. PLoS Genet. 2010, 6, e1001193. [Google Scholar] [CrossRef]
- Cotsaftis, O.; Plett, D.; Shirley, N.; Tester, M.; Hrmova, M. A two-staged model of Na+ exclusion in rice explained by 3d modeling of HKT transporters and alternative splicing. PLoS ONE 2012, 7, e39865. [Google Scholar] [CrossRef]
- Ren, Z.H.; Gao, J.P.; Li, L.G.; Cai, X.L.; Huang, W.; Chao, D.Y.; Zhu, M.Z.; Wang, Z.Y.; Luan, S.; Lin, H.X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Xu, B.; Athman, A.; Conn, S.J.; Jordans, C.; Byrt, C.S.; Hare, R.A.; Tyerman, S.D.; Tester, M.; et al. Wheat grain yield on saline soils is improved by an ancestral Na+ transporter gene. Nat. Biotechnol. 2012, 30, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Waters, S.; Gilliham, M.; Hrmova, M. Plant High-Affinity Potassium (HKT) Transporters Involved in Salinity Tolerance: Structural insights to Probe Differences in ion Selectivity. Int. J. Mol. Sci. 2013, 14, 7660–7680. [Google Scholar] [CrossRef]
- Laurie, S.; Feeney, K.A.; Maathuis, F.J.M.; Heard, P.J.; Brown, S.J.; Leigh, R.A. A role for HKT1 in sodium uptake by wheat roots. Plant J. 2002, 32, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, D.; Liu, C.; Zhang, A. The pyridoxal kinase gene TaPdxK from wheat complements vitamin B 6 synthesis-defective Escherichia coli. J. Plant Physiol. 2004, 161, 1053–1060. [Google Scholar] [CrossRef]
- Trchounian, A.; Petrosyan, M.; Sahakyan, N. Plant Cell Redox Homeostasis and Reactive Oxygen Species in Redox State as a Central Regulator of Plant-Cell Stress Responses; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Switzerland, 2016; pp. 25–50. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Sairam, R.K.; Srivastava, G.C. Changes in antioxidant activity in sub-cellular fractions of tolerant and susceptible wheat genotypes in response to long term salt stress. Plant Sci. 2002, 162, 897–904. [Google Scholar] [CrossRef]
- Sairam, R.K.; Srivastava, G.C.; Agarwal, S.; Meena, R.C. Differences in antioxidant activity in response to salinity stress in tolerant and susceptible wheat genotypes. Biol. Plant. 2005, 49, 85. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and Glutathione: Keeping Active Oxygen under Control. Annu. Rev. Plant Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Sairam, R.K.; Saxena, D.C. Oxidative stress and antioxidants in wheat genotypes: Possible mechanism of water stress tolerance. J. Agron. Crop Sci. 2000, 184, 55–61. [Google Scholar] [CrossRef]
- Zhai, C.Z.; Zhao, L.; Yin, L.J.; Chen, M.; Wang, Q.Y.; Li, L.C.; Xu, Z.S.; Ma, Y.Z. Two Wheat Glutathione Peroxidase Genes Whose Products Are Located in Chloroplasts Improve Salt and H2O2 Tolerances in Arabidopsis. PLoS ONE 2013, 8, e73989. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, S.; Wang, M.; Wei, T.; Meng, C.; Wang, M.; Xia, G. A wheat SIMILAR TO RCD-ONE gene enhances seedling growth and abiotic stress resistance by modulating redox homeostasis and maintaining genomic integrity. Plant Cell 2014, 26, 164–180. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Yan, J.; Li, W.; Wang, L.; Zhao, J.; Ma, X.; Li, A.; Wang, H.; Kong, L. A member of wheat class III peroxidase gene family, TaPRX-2A, enhanced the tolerance of salt stress. BMC Plant Biol. 2020, 20, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Osipova, S.V.; Permyakov, A.V.; Permyakova, M.D.; Pshenichnikova, T.A.; Börner, A. Leaf dehydroascorbate reductase and catalase activity is associated with soil drought tolerance in bread wheat. Acta Physiol. Plant. 2011, 33, 2169–2177. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, Y.; Pu, Z.; Wang, J.; Zheng, Y.; Li, Y.; Wei, Y. Regulation, evolution, and functionality of flavonoids in cereal crops. Biotechnol. Lett. 2013, 35, 1765–1780. [Google Scholar] [CrossRef] [PubMed]
- Shoeva, O.Y.; Khlestkina, E.K. Differently expressed ‘early’ flavonoid Synthesis genes in wheat seedlings become to be co-regulated under salinity stress. Cereal Res. Commun. 2015, 43, 537–543. [Google Scholar] [CrossRef][Green Version]
- Wei, D.; Wang, M.; Xu, F.; Quan, T.; Peng, K.; Xiao, L.; Xia, G. Wheat oxophytodienoate reductase gene TaOPR1 confers salinity tolerance via enhancement of abscisic acid signaling and reactive oxygen species scavenging. Plant Physiol. 2013, 161, 1217–1228. [Google Scholar] [CrossRef]
- Liu, C.; Li, S.; Wang, M.; Xia, G. A transcriptomic analysis reveals the nature of salinity tolerance of a wheat introgression line. Plant Mol. Biol. 2012, 78, 159–169. [Google Scholar] [CrossRef]





| Osmotic Stress | Ionic Stress | ||||
|---|---|---|---|---|---|
| Process | Candidate Genes | Osmotic Tolerance | Na+ Excluding | Tissue Tolerance | References |
| Signaling | SOS3, SnRKs | Signaling regulation | Activation of ion antiporter | Regulation of vacuolar loading | [77] |
| Photosynthesis | ERA1, PP2C, AAPK, PKS3 | Stomatal closure regulation | Protection of chloroplast from ion toxicity | Delay Na+ toxicity effect in chloroplast | [78,79] |
| Accumulation of Na+ in shoots | HKT, SOS1 | - | Decreasing long distance transport of Na+ | Decreasing energy used on Na+ exclusion | [80,81,82] |
| Accumulation of Na+in vacuoles | NHX, AVP | - | Increased sequestration of Na+ into root vacuoles | Increased sequestration of Na+ into leaf vacuoles | [80,83] |
| Accumulation of organic solutes | P5CS, OTS, MT1D, M6PR, S6PDH, IMT1 | Increasing osmotic adjustment | Reduction of Na+ accumulation | Accumulation of organic solutes in cytoplasm | [82,84] |
| Time | Water Stress Effect (Salt-Tolerant Plants) | Salt-Specific Effects Salt-Sensitive Plants |
|---|---|---|
| Minutes | Immediate reduction in leaf and root elongation rate and then rapid partial recovery | |
| Hours | Constant but reduced rate of leaf and root elongation | |
| Days | Leaf growth more affected than root growth; Reduced rate of leaf emergence | Visible injury in the oldest leaf |
| Weeks | Reduced the final size of the leaves and/or the number of side shoots | Death of older leaves |
| Months | Altered flowering time, reduced seed production | Younger leaves dead, plants may die before the seed matures |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbanavičiūtė, I.; Bonfiglioli, L.; Pagnotta, M.A. One Hundred Candidate Genes and Their Roles in Drought and Salt Tolerance in Wheat. Int. J. Mol. Sci. 2021, 22, 6378. https://doi.org/10.3390/ijms22126378
Urbanavičiūtė I, Bonfiglioli L, Pagnotta MA. One Hundred Candidate Genes and Their Roles in Drought and Salt Tolerance in Wheat. International Journal of Molecular Sciences. 2021; 22(12):6378. https://doi.org/10.3390/ijms22126378
Chicago/Turabian StyleUrbanavičiūtė, Ieva, Luca Bonfiglioli, and Mario A. Pagnotta. 2021. "One Hundred Candidate Genes and Their Roles in Drought and Salt Tolerance in Wheat" International Journal of Molecular Sciences 22, no. 12: 6378. https://doi.org/10.3390/ijms22126378
APA StyleUrbanavičiūtė, I., Bonfiglioli, L., & Pagnotta, M. A. (2021). One Hundred Candidate Genes and Their Roles in Drought and Salt Tolerance in Wheat. International Journal of Molecular Sciences, 22(12), 6378. https://doi.org/10.3390/ijms22126378