
The Organization of the Golgi Structures during Drosophila Male Meiosis Requires the Citrate Lyase ATPCL




{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
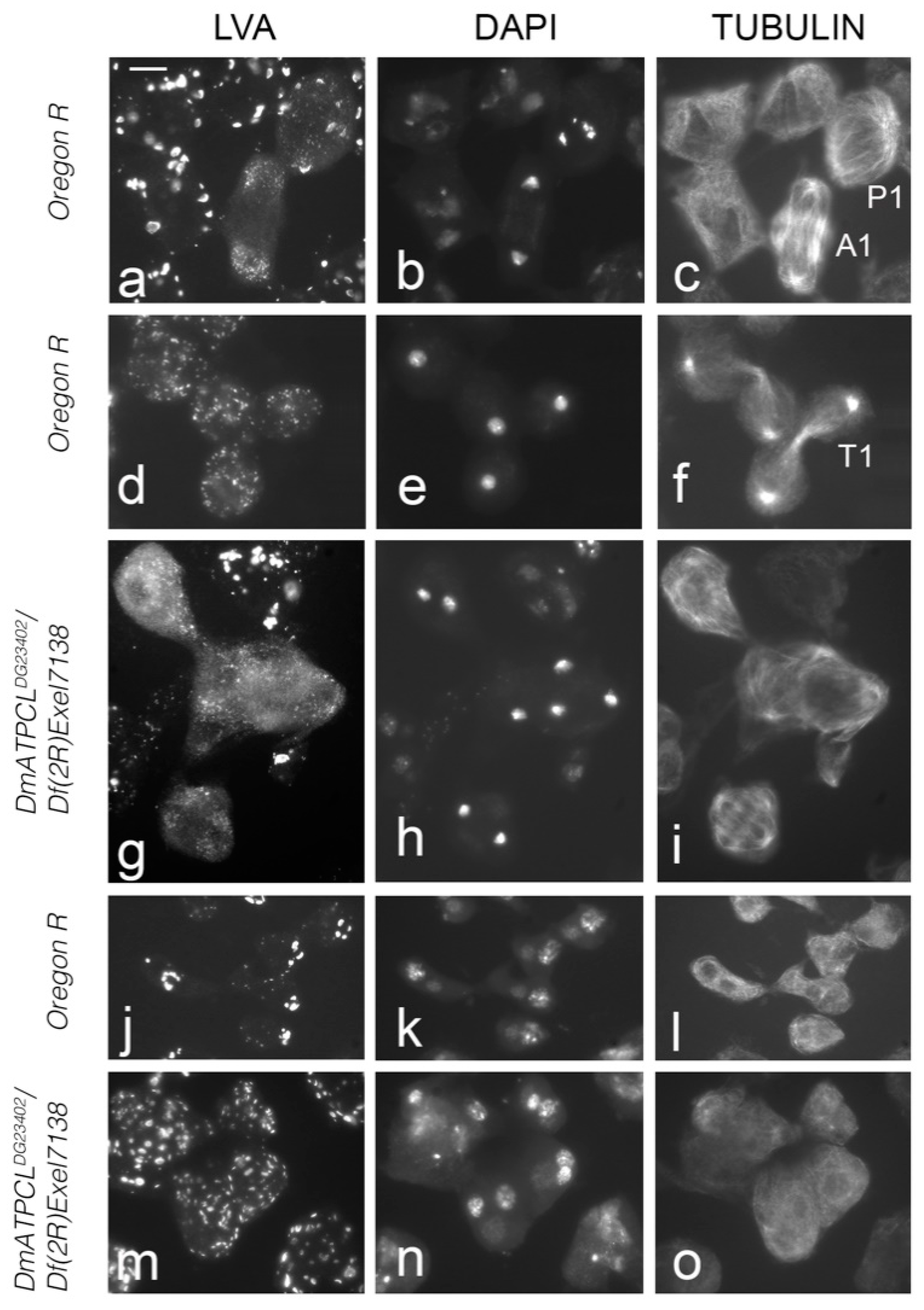
2. Results and Conclusions
ATPCL Is Required for Acroblast Assembly and Golgi Integrity
3. Materials and Methods
3.1. Drosophila Strains and Crosses
3.2. Cromosome Cytology, Immunostaining, and Microscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chypre, M.; Zaidi, N.; Smans, K. ATP-citrate lyase: A mini-review. Biochem. Biophys. Res. Commun. 2012, 422, 1–4. [Google Scholar] [CrossRef]
- Sivanand, S.; Rhoades, S.; Jiang, Q.; Lee, J.V.; Benci, J.; Zhang, J.; Yuan, S.; Viney, I.; Zhao, S.; Carrer, A.; et al. Nuclear Acetyl-CoA Production by ACLY Promotes Homologous Recombination. Mol. Cell 2017, 67, 252–265. [Google Scholar] [CrossRef] [PubMed]
- Wellen, K.E.; Hatzivassiliou, G.; Sachdeva, U.M.; Bui, T.V.; Cross, J.R.; Thompson, C.B. ATP-citrate lyase links cellular metabolism to histone acetylation. Science 2009, 324, 1076–1080. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhang, L.; Xu, S.; Shen, A.Z. ATP-citrate lyase (ACLY) in lipid metabolism and atherosclerosis: An updated review. Prog. Lipid Res. 2020, 77, 101006. [Google Scholar] [CrossRef] [PubMed]
- Icard, P.; Wu, Z.; Fournel, L.; Coquerel, A.; Lincet, H.; Alifano, M. ATP citrate lyase: A central metabolic enzyme in cancer. Cancer Lett. 2020, 471, 125–134. [Google Scholar] [CrossRef]
- Michno, A.; Skibowska, A.; Raszeja-Specht, A.; Cwikowska, J.; Szutowicz, A. The role of adenosine triphosphate citrate lyase in the metabolism of acetyl coenzyme a and function of blood platelets in diabetes mellitus. Metabolism 2004, 53, 66–72. [Google Scholar] [CrossRef]
- Tian, H.; Shi, S.; You, B.; Zhang, Q.; Gu, M.; You, Y. ER resident protein 44 promotes malignant phenotype in nasopharyngeal carcinoma through the interaction with ATP citrate lyase. J. Transl. Med. 2021, 19, 77. [Google Scholar] [CrossRef] [PubMed]
- Di Giorgio, M.L.; Morciano, P.; Bucciarelli, E.; Porrazzo, A.; Cipressa, F.; Saraniero, S.; Manzi, D.; Rong, Y.S.; Cenci, G. The Drosophila Citrate Lyase Is Required for Cell Division during Spermatogenesis. Cells 2020, 9, 206. [Google Scholar] [CrossRef]
- Fuller, L.M. Spermatogenesis. In The Development of Drosophila Melanogaster; Cold Spring Harbor Press: Cold Spring Harbor, NY, USA, 1993. [Google Scholar]
- Hime, G.R.; Brill, J.A.; Fuller, M.T. Assembly of ring canals in the male germ line from structural components of the contractile ring. J. Cell Sci. 1996, 109 Pt 12, 2779–2788. [Google Scholar] [CrossRef]
- Cenci, G.; Bonaccorsi, S.; Pisano, C.; Verni, F.; Gatti, M. Chromatin and microtubule organization during premeiotic, meiotic and early postmeiotic stages of Drosophila melanogaster spermatogenesis. J. Cell Sci. 1994, 107 Pt 12, 3521–3534. [Google Scholar] [CrossRef]
- Farkas, R.M.; Giansanti, M.G.; Gatti, M.; Fuller, M.T. The Drosophila Cog5 homologue is required for cytokinesis, cell elongation, and assembly of specialized Golgi architecture during spermatogenesis. Mol. Biol. Cell 2003, 14, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Yasuno, Y.; Kawano, J.; Inoue, Y.H.; Yamamoto, M.T. Distribution and morphological changes of the Golgi apparatus during Drosophila spermatogenesis. Dev. Growth Differ. 2013, 55, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Morciano, P.; Di Giorgio, M.L.; Porrazzo, A.; Licursi, V.; Negri, R.; Rong, Y.; Cenci, G. Depletion of ATP-Citrate Lyase (ATPCL) Affects Chromosome Integrity Without Altering Histone Acetylation in Drosophila Mitotic Cells. Front. Physiol. 2019, 10, 383. [Google Scholar] [CrossRef] [PubMed]
- Tates, A.D. Cytodifferentiation During Spermatogenesis in Drosophila Melanogaster: An Electron Microscope Study. Ph.D. Thesis, Rijksuniversiteit, Leiden, The Netherlands, 1971. [Google Scholar]
- Giansanti, M.G.; Bonaccorsi, S.; Kurek, R.; Farkas, R.M.; Dimitri, P.; Fuller, M.T.; Gatti, M. The class I PITP giotto is required for Drosophila cytokinesis. Curr. Biol. 2006, 16, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Giansanti, M.G.; Belloni, G.; Gatti, M. Rab11 is required for membrane trafficking and actomyosin ring constriction in meiotic cytokinesis of Drosophila males. Mol. Biol. Cell 2007, 18, 5034–5047. [Google Scholar] [CrossRef] [PubMed]
- Dieterich, I.A.; Cui, Y.; Braun, M.M.; Lawton, A.J.; Robinson, N.H.; Peotter, J.L.; Yu, Q.; Casler, J.C.; Glick, B.S.; Audhya, A.; et al. Acetyl-CoA flux from the cytosol to the ER regulates engagement and quality of the secretory pathway. Sci. Rep. 2021, 11, 2013. [Google Scholar] [CrossRef]
- Egea, G.; Serra-Peinado, C.; Salcedo-Sicilia, L.; Gutierrez-Martinez, E. Actin acting at the Golgi. Histochem. Cell Biol. 2013, 140, 347–360. [Google Scholar] [CrossRef]
- Gurel, P.S.; Hatch, A.L.; Higgs, H.N. Connecting the cytoskeleton to the endoplasmic reticulum and Golgi. Curr. Biol. 2014, 24, R660–R672. [Google Scholar] [CrossRef]
- Makowski, S.L.; Tran, T.T.; Field, S.J. Emerging themes of regulation at the Golgi. Curr. Opin. Cell Biol. 2017, 45, 17–23. [Google Scholar] [CrossRef]
- Kondylis, V.; van Nispen tot Pannerden, H.E.; Herpers, B.; Friggi-Grelin, F.; Rabouille, C. The golgi comprises a paired stack that is separated at G2 by modulation of the actin cytoskeleton through Abi and Scar/WAVE. Dev. Cell 2007, 12, 901–915. [Google Scholar] [CrossRef] [PubMed]
- Bard, F.; Casano, L.; Mallabiabarrena, A.; Wallace, E.; Saito, K.; Kitayama, H.; Guizzunti, G.; Hu, Y.; Wendler, F.; Dasgupta, R.; et al. Functional genomics reveals genes involved in protein secretion and Golgi organization. Nature 2006, 439, 604–607. [Google Scholar] [CrossRef]
- Zanetti, S.R.; Monclus Mde, L.; Rensetti, D.E.; Fornes, M.W.; Aveldano, M.I. Differential involvement of rat sperm choline glycerophospholipids and sphingomyelin in capacitation and the acrosomal reaction. Biochimie 2010, 92, 1886–1894. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, M.; Gupta, G.; Jain, S.K.; Setty, B.S. Lipid metabolising enzymes in isolated rat testicular germ cells and changes associated with meiosis. Andrologia 1998, 30, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.Y.; Yang, Y.H.; Chen, S.R. Molecular genetics of infertility: Loss-of-function mutations in humans and corresponding knockout/mutated mice. Hum. Reprod. Update 2021, 27, 154–189. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morciano, P.; Di Giorgio, M.L.; Tullo, L.; Cenci, G. The Organization of the Golgi Structures during Drosophila Male Meiosis Requires the Citrate Lyase ATPCL. Int. J. Mol. Sci. 2021, 22, 5745. https://doi.org/10.3390/ijms22115745
Morciano P, Di Giorgio ML, Tullo L, Cenci G. The Organization of the Golgi Structures during Drosophila Male Meiosis Requires the Citrate Lyase ATPCL. International Journal of Molecular Sciences. 2021; 22(11):5745. https://doi.org/10.3390/ijms22115745
Chicago/Turabian StyleMorciano, Patrizia, Maria Laura Di Giorgio, Liliana Tullo, and Giovanni Cenci. 2021. "The Organization of the Golgi Structures during Drosophila Male Meiosis Requires the Citrate Lyase ATPCL" International Journal of Molecular Sciences 22, no. 11: 5745. https://doi.org/10.3390/ijms22115745
APA StyleMorciano, P., Di Giorgio, M. L., Tullo, L., & Cenci, G. (2021). The Organization of the Golgi Structures during Drosophila Male Meiosis Requires the Citrate Lyase ATPCL. International Journal of Molecular Sciences, 22(11), 5745. https://doi.org/10.3390/ijms22115745