
New Era in the Treatment of Iron Deficiency Anaemia Using Trimaltol Iron and Other Lipophilic Iron Chelator Complexes: Historical Perspectives of Discovery and Future Applications

 ,
,
Abstract
1. Introduction
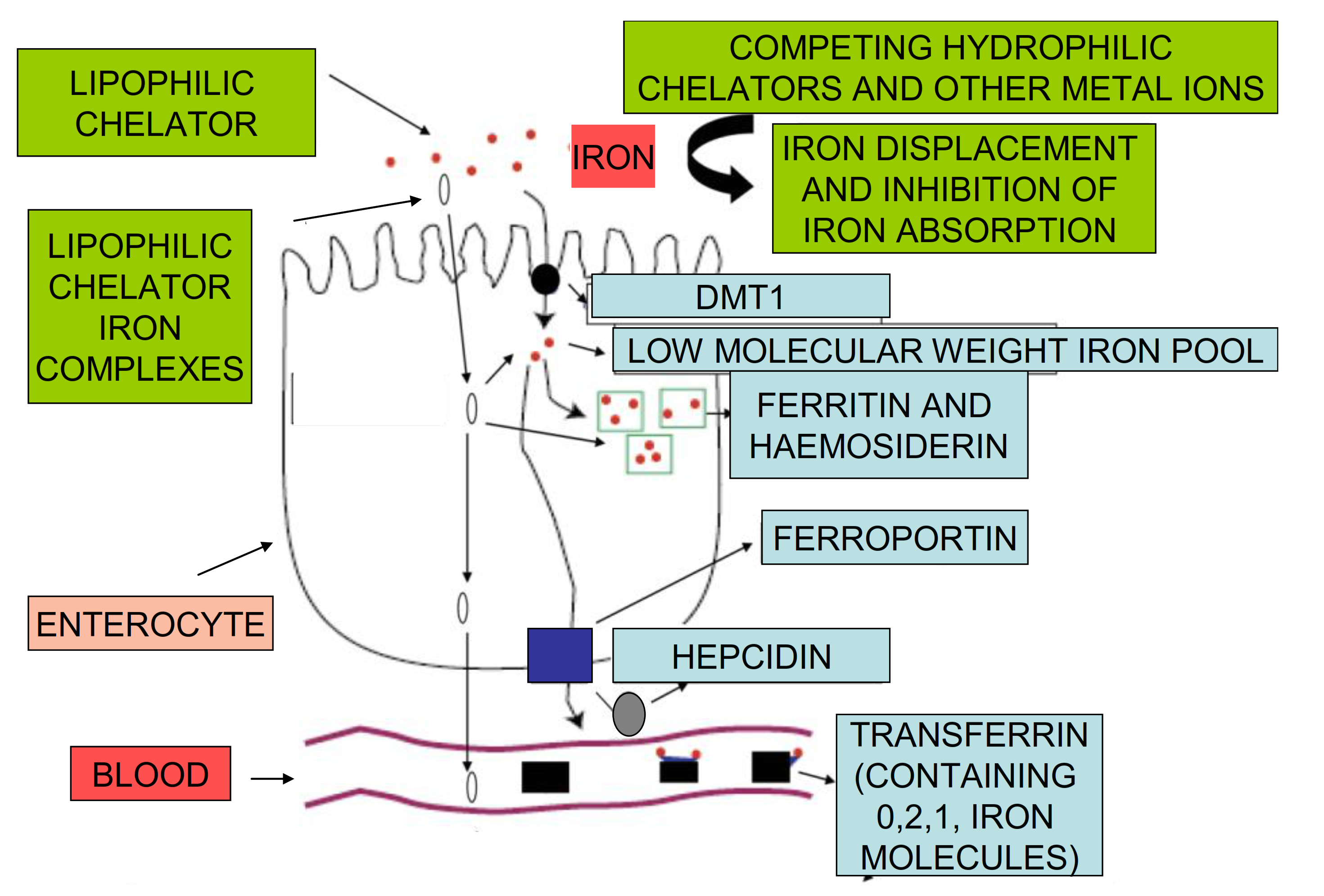
2. Iron Absorption and Distribution Pathways
2.1. Iron Absorption in Humans
2.2. The Role of Proteins of Iron Metabolism in the Regulation of Iron Absorption
2.3. Differences among Individuals in Iron Absorption Requirements
3. Non-Regulatory Mechanisms of Iron Absorption
3.1. Dietary Molecules and Forms of Iron Affecting Its Absorption
3.2. Iron Chelating Drugs and Other Drugs Affecting Iron Absorption
3.3. Quantitative Aspects of Iron Affecting the Rate of Iron Absorption
4. Iron Formulations Used for the Treatment of Iron Deficiency Anaemia
5. The In Vitro Properties of Iron Maltol and Other Lipophilic Iron Complexes
5.1. Physicochemical Properties of Lipophilic Chelators and Their Iron Complexes
5.2. In Vitro Properties of Maltol and the Maltol Iron Complex
5.3. Interactions of Maltol and the Maltol Iron Complex with Proteins
5.4. The Antioxidant Effects of Maltol and Other Iron Chelators
6. Cell Studies Using Maltol and Other Chelator Iron Complexes
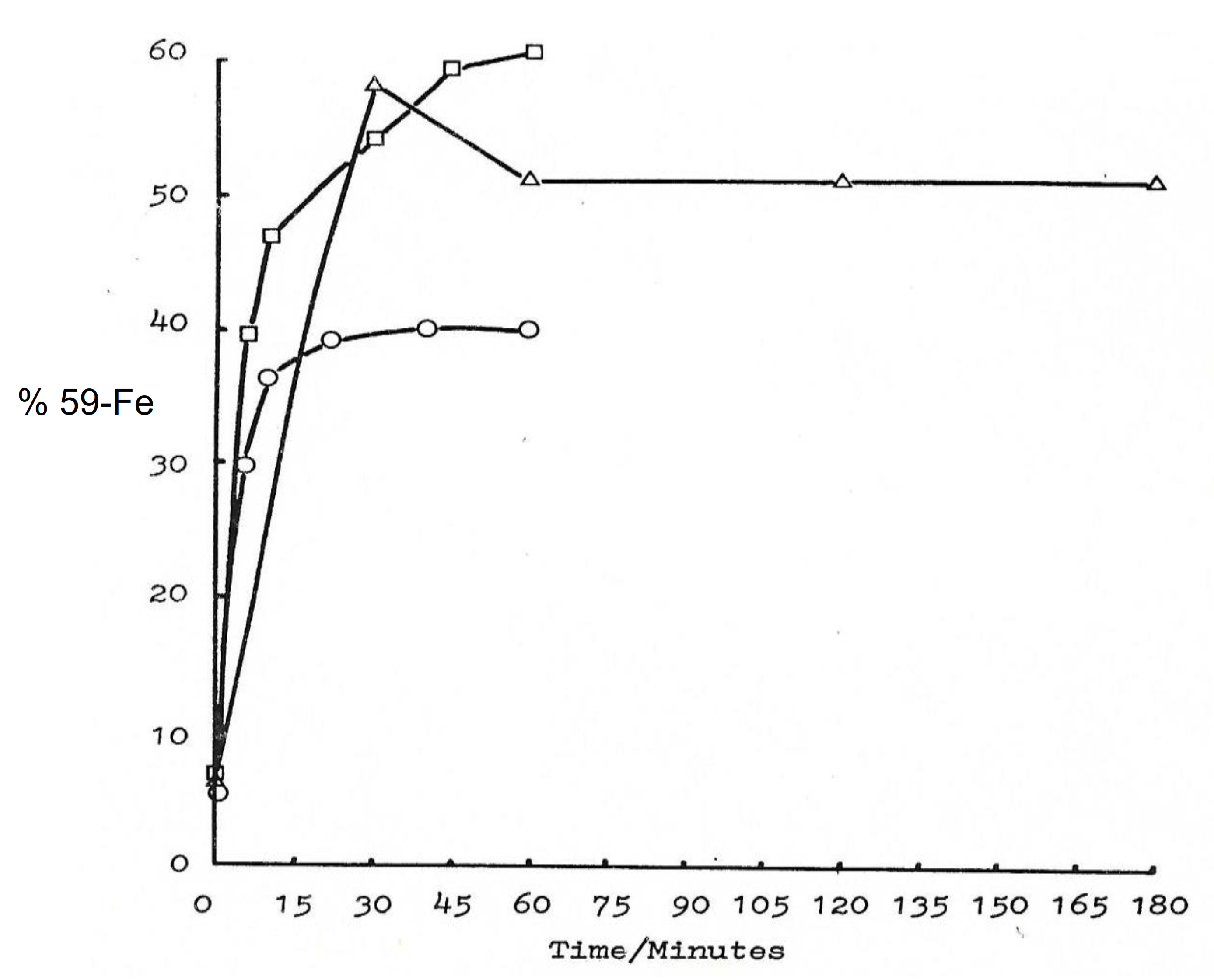
6.1. Iron Transport by Maltol and Other Chelators in Matured Red Blood Cells
6.2. The Role of Chelators in the Uptake and Intracellular Distribution of Iron in Erythroid Cells
6.3. The Antimicrobial Effects of Maltol and Other Chelators
6.4. Uptake of Iron Maltol by the Enterocyte
7. In Vivo Studies of the Effect of Maltol and Other Chelators on Iron Absorption
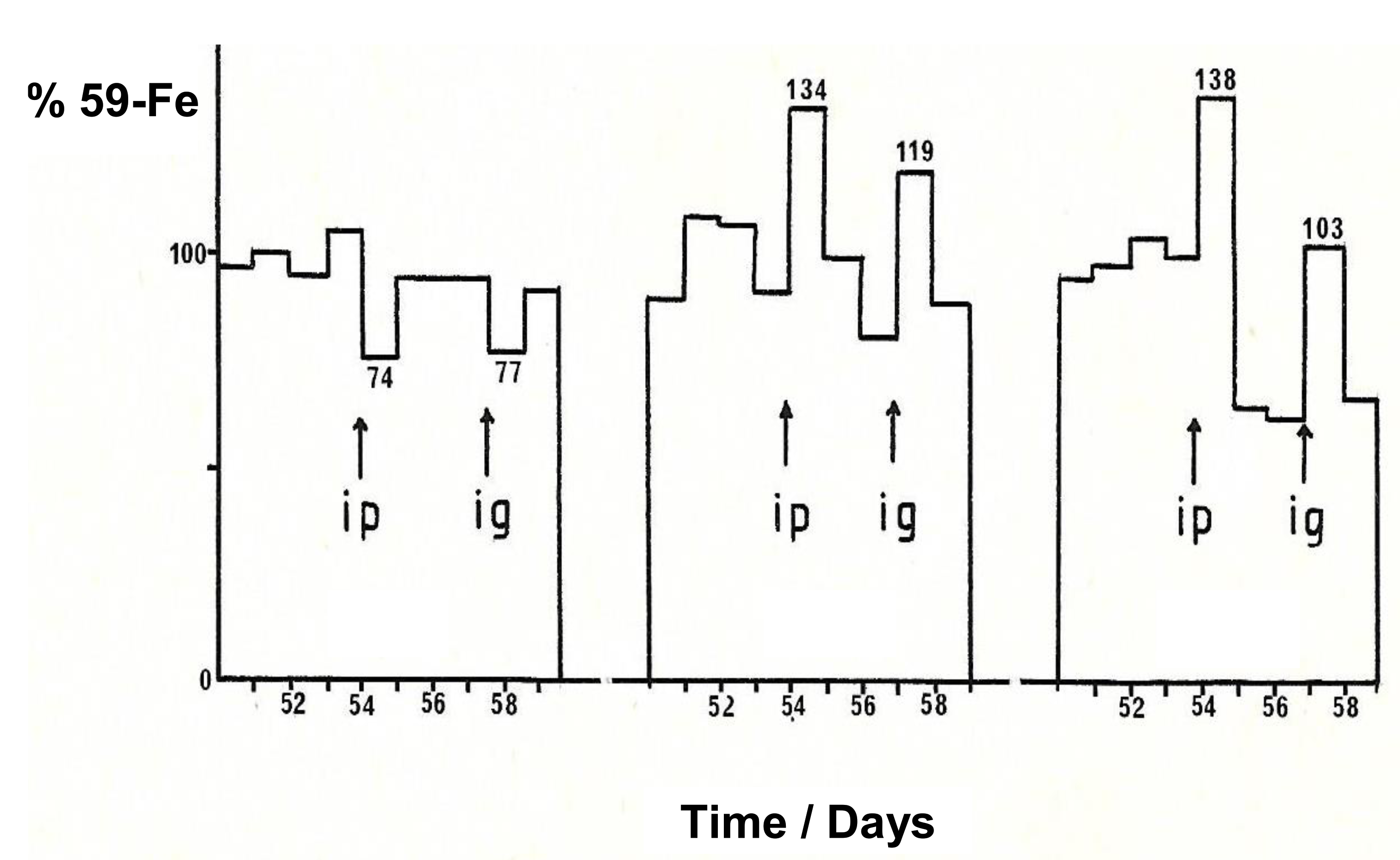
7.1. Increase in Iron Absorption in Animals Using Maltol and Other Lipophilic Chelators
7.2. The Effect of Maltol and Maltol Iron Complex on Iron Excretion
8. Factors Affecting the Absorption of Maltol and the Maltol Iron Complex
9. Clinical Studies Related to the Efficacy of Iron Maltol in Iron Deficiency Anaemia
10. Potential Applications of Maltol and Maltol Metal Complexes in Medicine
11. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 2,4-Dihydroxypyridine-1-oxide | (L3) |
| 2-Hydroxypyridine-1-oxide | (L4) |
| 2-Hydroxy-4- methoxypyridine-1-oxide | (L6) |
| Diferric transferrin | (Fe-T-Fe) |
| Divalent metal transported protein | (DMT1) |
| Ethylenediaminetetraacetic acid | (EDTA) |
| Optical density | (OD) |
| Hydroxyeicosatetraenoic acid | (HETE) |
| Hydroperoxyeicosatetraenoic acid | (HPETE) |
| Investigational new drugs | (IND) |
| Iron deficiency anaemia | (IDA) |
| Red blood cells | (RBC) |
References
- Katsarou, A.; Pantopoulos, K. Basics and principles of cellular and systemic iron homeostasis. Mol. Aspects Med. 2020, 75, 100866. [Google Scholar] [CrossRef] [PubMed]
- Gozzelino, R.; Arosio, P. Iron Homeostasis in Health and Disease. Int. J. Mol. Sci. 2016, 17, 130. [Google Scholar] [CrossRef] [PubMed]
- Cairo, G.; Bernuzzi, F.; Recalcati, S.A. A precious metal: Iron, an essential nutrient for all cells. Genes Nutr. 2006, 1, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Andrews, N.C. Disorders of iron metabolism. N. Engl. J. Med. 1999, 341, 1986–1995, Erratum in 2000, 342, 364. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Zinc: An overview. Nutrition 1995, 11 (Suppl. S1), 93–99. [Google Scholar]
- Coleman, J.E. Zinc proteins: Enzymes, storage proteins, transcription factors, and replication proteins. Annu. Rev. Biochem. 1992, 61, 897–946. [Google Scholar] [CrossRef]
- Prasad, A.S. Zinc deficiency. BMJ 2003, 326, 409–410. [Google Scholar] [CrossRef]
- Daniel, K.G.; Harbach, R.H.; Guida, W.C.; Dou, Q.P. Copper storage diseases: Menkes, Wilsons, and cancer. Front. Biosci. 2004, 9, 2652–2662. [Google Scholar] [CrossRef]
- Baldari, S.; Di Rocco, G.; Toietta, G. Current Biomedical Use of Copper Chelation Therapy. Int. J. Mol. Sci. 2020, 21, 1069. [Google Scholar] [CrossRef]
- Balsano, C.; Porcu, C.; Sideri, S. Is copper a new target to counteract the progression of chronic diseases? Metallomics 2018, 10, 1712–1722. [Google Scholar] [CrossRef]
- Kontoghiorghe, G.J.; Kontoghiorghe, C.N. Iron and Chelation in Biochemistry and Medicine: New Approaches to Controlling Iron Metabolism and Treating Related Diseases. Cells 2020, 9, 1456. [Google Scholar] [CrossRef]
- McLean, E.; Cogswell, M.; Egli, I.; Wojdyla, D.; De Benoist, B. Worldwide Prevalence of Anaemia, WHO Vitamin and Mineral Nutrition Information System, 1993–2005. Public Health Nutr. 2009, 12, 444–454. [Google Scholar] [CrossRef]
- Pasricha, S.R.; Tye-Din, J.; Muckenthaler, M.U.; Swinkels, D.W. Iron deficiency. Lancet 2021, 397, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Tardy, A.L.; Pouteau, E.; Marquez, D.; Yilmaz, C.; Scholey, A. Vitamins and Minerals for Energy, Fatigue and Cognition: A Narrative Review of the Biochemical and Clinical Evidence. Nutrients 2020, 12, 228. [Google Scholar] [CrossRef]
- Moustarah, F.; Mohiuddin, S.S. Dietary iron. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Pantopoulos, K. TfR2 links iron metabolism and erythropoiesis. Blood 2015, 125, 1055–1056. [Google Scholar] [CrossRef]
- Makey, D.G.; Seal, U.S. The detection of four molecular forms of human transferrin during the iron binding process. Biochim. Biophys. Acta 1976, 453, 250–256. [Google Scholar] [CrossRef]
- Gomme, P.T.; McCann, K.B.; Bertolini, J. Transferrin: Structure, function and potential therapeutic actions. Drug Discov. Today 2005, 10, 267–273. [Google Scholar] [CrossRef]
- Sargent, P.J.; Farnaud, S.; Evans, R.W. Structure/function overview of proteins involved in iron storage and transport. Curr. Med. Chem. 2005, 12, 2683–2693. [Google Scholar] [CrossRef]
- Mehlenbacher, M.; Poli, M.; Arosio, P.; Santambrogio, P.; Levi, S.; Chasteen, N.D.; Bou-Abdallah, F. Iron Oxidation and Core Formation in Recombinant Heteropolymeric Human Ferritins. Biochemistry 2017, 56, 3900–3912. [Google Scholar] [CrossRef]
- Theil, E.C. Ferritin: The protein nanocage and iron biomineral in health and in disease. Inorg. Chem. 2013, 52, 12223–12233. [Google Scholar] [CrossRef]
- Iancu, T.C. Ferritin and hemosiderin in pathological tissues. Electron. Microsc Rev. 1992, 5, 209–229. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kolnagou, A. Molecular factors and mechanisms affecting iron and other metal excretion or absorption in health and disease. The role of natural and synthetic chelators. Curr. Med. Chem. 2005, 12, 2695–2709. [Google Scholar] [CrossRef]
- Anderson, F.W.; Hiller, M.C. (Eds.) Development of Iron Chelators for Clinical Use; DHEW Publication No. (NIH) 77-994; DHEW Publication: Bethesda, MD, USA, 1975; pp. 1–275. [Google Scholar]
- Kontoghiorghes, G.J. The Design of Orally Active Iron Chelators for the Treatment of Thalassaemia. Ph.D. Thesis, University of Essex, Colchester, UK, 1982; pp. 1–243, British Library. Microfilm No D66194/86. Available online: https://www.pri.ac.cy/files/KGJ_thesis_1982.pdf (accessed on 20 May 2021).
- Kawabata, H. Transferrin and transferrin receptors update. Free Radic. Biol. Med. 2019, 133, 46–54. [Google Scholar] [CrossRef]
- Wang, B.; Timilsena, Y.P.; Blanch, E.; Adhikari, B. Lactoferrin: Structure, function, denaturation and digestion. Crit. Rev. Food Sci. Nutr. 2019, 59, 580–596. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. Advances on Chelation and Chelator Metal Complexes in Medicine. Int. J. Mol. Sci. 2020, 21, 2499. [Google Scholar] [CrossRef]
- Kobayashi, M.; Suhara, T.; Baba, Y.; Kawasaki, N.K.; Higa, J.K.; Matsui, T. Pathological Roles of Iron in Cardiovascular Disease. Curr. Drug Targets 2018, 19, 1068–1076. [Google Scholar] [CrossRef]
- Wessells, K.R.; Young, R.R.; Ferguson, E.L.; Ouédraogo, C.T.; Faye, M.T.; Hess, S.Y. Assessment of Dietary Intake and Nutrient Gaps, and Development of Food-Based Recommendations, Among Pregnant and Lactating Women in Zinder, Niger: An Optifood Linear Programming Analysis. Nutrients 2019, 11, 72. [Google Scholar] [CrossRef]
- Zhang, C.; Rawal, S. Dietary Iron Intake, Iron Status, and Gestational Diabetes. Am. J. Clin. Nutr. 2017, 106, 1672–1680. [Google Scholar] [CrossRef]
- Man, C.D.; Maideen, S.F.K.; Rashid, A. Knowledge, Attitude and Practice towards Dietary Iron among Patients with Thalassemia and Their Caregivers in Peninsular Malaysia. Med. J. Malays 2019, 74, 365–371. [Google Scholar]
- Ganz, T. Anemia of Inflammation. N. Engl. J. Med. 2019, 381, 1148–1157. [Google Scholar] [CrossRef] [PubMed]
- Tolkien, Z.; Stecher, L.; Mander, A.P.; Pereira, D.I.; Powell, J.J. Ferrous sulfate supplementation causes significant gastrointestinal side-effects in adults: A systematic review and meta-analysis. PLoS ONE 2015, 10, e0117383. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bouvard, V.; Loomis, D.; Guyton, K.Z.; Grosse, Y.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Mattock, H.; Straif, K.; Stewart, B.W.; et al. Carcinogenicity of Consumption of Red and Processed Meat. Lancet Oncol. 2015, 16, 1599–1600. [Google Scholar] [CrossRef]
- Kontoghiorghe, C.N.; Kolnagou, A.; Kontoghiorghes, G.J. Dietary and Pharmacological Factors Affecting Iron Absorption in Mice and Man (Comment for a Letter to the Editor). Haematologica 2016, 101, 120–121. [Google Scholar] [CrossRef] [PubMed]
- Sim, M.; Garvican-Lewis, L.A.; Cox, G.R.; Govus, A.; McKay, A.K.A.; Stellingwerff, T.; Peeling, P. Iron considerations for the athlete: A narrative review. Eur. J. Appl. Physiol. 2019, 119, 1463–1478. [Google Scholar] [CrossRef] [PubMed]
- Salvin, H.E.; Pasricha, S.R.; Marks, D.C.; Speedy, J. Iron deficiency in blood donors: A national cross-sectional study. Transfusion 2014, 54, 2434–2444. [Google Scholar] [CrossRef] [PubMed]
- Mantadakis, E.; Chatzimichael, E.; Zikidou, P. Iron Deficiency Anemia in Children Residing in High and Low-Income Countries: Risk Factors, Prevention, Diagnosis and Therapy. Mediterr. J. Hematol. Infect. Dis. 2020, 12, e2020041. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Grover, K.; Kumar, T.; Doda, A.; Bhutani, R.; Yadav, S.; Kaushal, P.; Kapoor, R.; Sharma, S. Prevalence of anaemia and its association with dietary habits among pregnant women in the urban area of Haryana. J. Fam. Med. Prim. Care 2020, 9, 783–787. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Savva, S.C.; Kafatos, A. Is red meat required for the prevention of iron deficiency among children and adolescents? Curr. Pediatr. Rev. 2014, 10, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Spyrou, A.; Kolnagou, A. Iron chelation therapy in hereditary hemochromatosis and thalassemia intermedia: Regulatory and non regulatory mechanisms of increased iron absorption. Hemoglobin 2010, 34, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Girelli, D.; Nemeth, E.; Swinkels, D.W. Hepcidin in the diagnosis of iron disorders. Blood 2016, 127, 2809–2813. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Aschemeyer, S.; Qiao, B.; Stefanova, D.; Valore, E.V.; Sek, A.C.; Alex Ruwe, T.; Vieth, K.R.; Jung, G.; Casu, C.; Rivella, S.; et al. Structure-function analysis of ferroportin defines the binding site and an alternative mechanism of action of hepcidin. Blood 2018, 131, 899–910. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Powers, A.S.; Gonen, S.; Schneider, S.; Arvedson, T.; Dror, R.O.; Cheng, Y.; Manglik, A. Structure of hepcidin-bound ferroportin reveals iron homeostatic mechanisms. Nature 2020, 586, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.L.; Ghosh, M.C.; Ollivierre, H.; Li, Y.; Rouault, T.A. Ferroportin deficiency in erythroid cells causes serum iron deficiency and promotes hemolysis due to oxidative stress. Blood 2018, 132, 2078–2087. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Drakesmith, H.; Nemeth, E.; Ganz, T. Ironing out Ferroportin. Cell Metab. 2015, 22, 777–787. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Pietrangelo, A. Ferroportin disease: Pathogenesis, diagnosis and treatment. Haematologica 2017, 102, 1972–1984. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Byrnes, V.; Barrett, S.; Ryan, E.; Kelleher, T.; O’Keane, C.; Coughlan, B.; Crowe, J. Increased duodenal DMT-1 expression and unchanged HFE mRNA levels in HFE-associated hereditary hemochromatosis and iron deficiency. Blood Cells Mol. Dis. 2002, 29, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Pantopoulos, K. Function of the hemochromatosis protein HFE: Lessons from animal models. World J. Gastroenterol. 2008, 14, 6893–6901. [Google Scholar] [CrossRef]
- Gombart, A.F.; Pierre, A.; Maggini, S. A Review of Micronutrients and the Immune System—Working in Harmony to Reduce the Risk of Infection. Nutrients 2020, 12, 236. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kolnagou, A.; Kontoghiorghe, C.N.; Mourouzidis, L.; Timoshnikov, V.A.; Polyakov, N.E. Trying to Solve the Puzzle of the Interaction of Ascorbic Acid and Iron: Redox, Chelation and Therapeutic Implications. Medicines 2020, 7, 45. [Google Scholar] [CrossRef]
- Sheppard, L.N.; Kontoghiorghes, G.J. Competition between deferiprone, desferrioxamine and other chelators for iron and the effect of other metals. Drug Res. 1993, 43, 659–663. [Google Scholar] [PubMed]
- Dresow, B.; Fischer, R.; Nielsen, P.; Gabbe, E.E.; Piga, A. Effect of oral iron chelator L1 on iron absorption in man. Ann. N. Y. Acad. Sci. 1998, 850, 466–468. [Google Scholar] [CrossRef] [PubMed]
- Jackson, T.W.; Ling, L.J.; Washington, V. The effect of oral deferoxamine on iron absorption in humans. J. Toxicol. Clin. Toxicol. 1995, 33, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Gomez, H.F.; McClafferty, H.H.; Flory, D.; Brent, J.; Dart, R.C. Prevention of gastrointestinal iron absorption by chelation from an orally administered premixed deferoxamine/charcoal slurry. Ann. Emerg Med. 1997, 30, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Eracleous, E.; Economides, C.; Kolnagou, A. Advances in iron overload therapies. Prospects for effective use of deferiprone (L1), deferoxamine, the new experimental chelators ICL670, GT56-252, L1NAll and their combination. Curr. Med. Chem. 2005, 12, 2663–2681. [Google Scholar] [CrossRef]
- Kontoghiorghe, C.N.; Kontoghiorghes, G.J. Efficacy and safety of iron-chelation therapy with deferoxamine, deferiprone, and deferasirox for the treatment of iron-loaded patients with non-transfusion-dependent thalassemia syndromes. Drug Des. Devel. Ther. 2016, 10, 465–481. [Google Scholar] [CrossRef]
- Djaldetti, M.; Fishman, P.; Notti, I.; Bessler, H. The effect of tetracycline administration on iron absorption in mice. Biomedicine 1981, 35, 150–152. [Google Scholar]
- Konstantinou, E.; Pashalidis, I.; Kolnagou, A.; Kontoghiorghes, G.J. Interactions of hydroxycarbamide (hydroxyurea) with iron and copper: Implications on toxicity and therapeutic strategies. Hemoglobin 2011, 35, 237–246. [Google Scholar] [CrossRef]
- Timoshnikov, V.A.; Kobzeva, T.V.; Polyakov, N.E.; Kontoghiorghes, G.J. Redox Interactions of Vitamin C and Iron: Inhibition of the Pro-Oxidant Activity by Deferiprone. Int. J. Mol. Sci. 2020, 21, 3967. [Google Scholar] [CrossRef]
- Senba, M.; Nakamura, T.; Itakura, H. Relationships among iron accumulation, cirrhosis, and hepatitis B virus infection in Bantu siderosis. Ann. Soc. Belg. Med. Trop. 1989, 69, 77–78. [Google Scholar]
- Robertson, A.; Tenenbein, M. Hepatotoxicity in acute iron poisoning. Hum. Exp. Toxicol. 2005, 24, 559–562. [Google Scholar] [CrossRef]
- Baranwal, A.K.; Singhi, S.C. Acute iron poisoning: Management guidelines. Indian Pediatr. 2003, 40, 534–540. [Google Scholar]
- Lucarelli, G.; Angelucci, E.; Giardini, C.; Baronciani, D.; Galimberti, M.; Polchi, P.; Bartolucci, M.; Muretto, P.; Albertini, F. Fate of iron stores in thalassaemia after bone-marrow transplantation. Lancet 1993, 342, 1388–1391. [Google Scholar] [CrossRef] [PubMed]
- Lucarelli, G.; Galimberti, M.; Polchi, P.; Angelucci, E.; Baronciani, D.; Giardini, C.; Andreani, M.; Agostinelli, F.; Albertini, F.; Clift, R.A. Marrow transplantation in patients with thalassemia responsive to iron chelation therapy. N. Engl. J. Med. 1993, 329, 840–844. [Google Scholar] [CrossRef] [PubMed]
- Lopez, A.; Cacoub, P.; Macdougall, I.C.; Peyrin-Biroulet, L. Iron deficiency anaemia. Lancet 2016, 387, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Barad, A.; Clark, A.G.; Wang, Y.; Lin, X.; Gu, Z.; O’Brien, K.O. Ethnic Differences in Iron Status. Adv. Nutr. 2021, nmab035. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghe, C.N.; Andreou, N.; Constantinou, K.; Kontoghiorghes, G.J. World health dilemmas: Orphan and rare diseases, orphan drugs and orphan patients. World J. Methodol. 2014, 4, 163–188. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rao, C.P.; Geetha, K.; Raghavan, M.S.S.; Sreedhara, A.; Tokunaga, K.; Yamaguchi, T.; Jadhav, V.; Ganesh, K.N.; Krishnamoorthy, T.; Ramaiah, K.V.A.; et al. Transition Metal Saccharide Chemistry and Biology: Syntheses, Characterization, Solution Stability and Putative Bio-Relevant Studies of Iron-Saccharide Complexes. Inorg. Chim. Acta 2000, 297, 373–382. [Google Scholar] [CrossRef]
- Chandra, J. Treating Iron Deficiency Anemia. Indian J. Pediatr. 2019, 86, 1085–1086. [Google Scholar] [CrossRef]
- Pachuta Węgier, L.; Kubiak, M.; Liebert, A.; Clavel, T.; Montagne, A.; Stennevin, A.; Roye, S.; Boudribila, A. Ferrous Sulfate Oral Solution in Young Children with Iron Deficiency Anemia. Pediatr. Int. 2020. [Google Scholar] [CrossRef]
- Valenzuela, C.; Olivares, M.; Brito, A.; Hamilton-West, C.; Pizarro, F. Is a 40% Absorption of Iron from a Ferrous ascorbate Reference Dose Appropriate to Assess Iron Absorption Independent of Iron Status? Biol. Trace Elem. Res. 2013, 155, 322–326. [Google Scholar] [CrossRef]
- Patil, P.; Geevarghese, P.; Khaire, P.; Joshi, T.; Suryawanshi, A.; Mundada, S.; Pawar, S.; Farookh, A. Comparison of Therapeutic Efficacy of Ferrous Ascorbate and Iron Polymaltose Complex in Iron Deficiency Anemia in Children: A Randomized Controlled Trial. Indian J. Pediatr. 2019, 86, 1112–1117. [Google Scholar] [CrossRef]
- Tarng, D.C.; Huang, T.P.; Wei, Y.H. Erythropoietin and Iron: The Role of Ascorbic Acid. Nephrol. Dial. Transplant. 2001, 16, 35–39. [Google Scholar] [CrossRef][Green Version]
- Sourabh, S.; Bhatia, P.; Jain, R. Favourable Improvement in Haematological Parameters in Response to Oral Iron and Vitamin C Combination in Children with Iron Refractory Iron Deficiency Anemia (IRIDA) Phenotype. Blood Cells Mol. Dis. 2019, 75, 26–29. [Google Scholar] [CrossRef]
- Scheers, N.; Sandberg, A.S. Iron Transport through Ferroportin Is Induced by Intracellular Ascorbate and Involves IRP2 and HIF2α. Nutrients 2014, 3, 249–260. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Barr, J.; Nortey, P.; Sheppard, L. Selection of a new generation of orally active alpha-ketohydroxypyridine iron chelators intended for use in the treatment of iron overload. Am. J. Hematol. 1993, 42, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghe, C.N.; Kontoghiorghes, G.J. New developments and controversies in iron metabolism and iron chelation therapy. World J. Methodol. 2016, 6, 1–19. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kontoghiorghe, C.N.; Kolnagou, A.; Kontoghiorghes, G.J. Phytochelators Intended for Clinical Use in Iron Overload, Other Diseases of Iron Imbalance and Free Radical Pathology. Molecules 2015, 20, 20841–20872. [Google Scholar] [CrossRef] [PubMed]
- Rennhard, H.H. The metabolism of ethyl maltol and maltol in the dog. J. Agric. Food Chem. 1971, 19, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Arosio, P.; Elia, L.; Poli, M. Ferritin, cellular iron storage and regulation. IUBMB Life 2017, 69, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Saito, H. Storage Iron Turnover from a New Perspective. Acta Haematol. 2019, 141, 201–208. [Google Scholar] [CrossRef]
- La, A.; Nguyen, T.; Tran, K.; Sauble, E.; Tu, D.; Gonzalez, A.; Kidane, T.Z.; Soriano, C.; Morgan, J.; Doan, M.; et al. Mobilization of iron from ferritin: New steps and details. Metallomics 2018, 10, 154–168. [Google Scholar] [CrossRef]
- Wang, W.; Knovich, M.A.; Coffman, L.G.; Torti, F.M.; Torti, S.V. Serum ferritin: Past, present and future. Biochim. Biophys. Acta 2010, 1800, 760–769. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. The study of iron mobilisation from transferrin using α-ketohydroxy heteroaromatic chelators. Biochim. Biophys. Acta 1986, 869, 141–146. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. Iron mobilisation from lactoferrin by chelators at physiological pH. Biochim. Biophys. Acta 1986, 882, 267–270. [Google Scholar] [CrossRef]
- Kontoghiorghe, C.N.; Kolnagou, A.; Kontoghiorghes, G.J. Potential clinical applications of chelating drugs in diseases targeting transferrin-bound iron and other metals. Expert Opin. Investig. Drugs 2013, 22, 591–618. [Google Scholar] [CrossRef]
- Levina, A.; Lay, P.A. Transferrin Cycle and Clinical Roles of Citrate and Ascorbate in Improved Iron Metabolism. ACS Chem. Biol. 2019, 14, 893–900. [Google Scholar] [CrossRef]
- Pratt, R.; Handelman, G.J.; Edwards, T.E.; Gupta, A. Ferric pyrophosphate citrate: Interactions with transferrin. Biometals 2018, 31, 1081–1089. [Google Scholar] [CrossRef]
- Evans, R.W.; Sharma, M.; Ogwang, W.; Patel, K.J.; Bartlett, A.N.; Kontoghiorghes, G.J. The effect of α-ketohydroxypyridine chelators on transferrin saturation in vitro and in vivo. Drugs Today 1992, 28, 9–23. [Google Scholar]
- Kontoghiorghes, G.J.; Goddard, J.G.; Bartlett, A.N.; Sheppard, L. Pharmacokinetic studies in humans with the oral iron chelator 1,2-dimethyl-3-hydroxypyrid-4-one. Clin. Pharmacol. Ther. 1990, 48, 255–261. [Google Scholar] [CrossRef]
- Vreugdenhil, G.; Kontoghiorghes, G.J.; Van Eijk, H.G.; Swaak, A.J.G. Impaired erythropoietin responsiveness to the anemia in rheumatoid arthritis. A possible inverse relationship with iron stores and effects of the oral iron chelator 1,2-dimethyl-3-hydroxypyrid-4-one. Clin. Exp. Rheumatol. 1991, 9, 35–40. [Google Scholar]
- Vreughtenhil, G.; Kontoghiorghes, G.J.; Van Eijk, H.G.; Swaak, A.J.G. Efficacy and safety of the oral chelator L1 in anaemic rheumadoit arthritis patients. Lancet 1989, II, 1398–1399. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. Iron mobilization from transferrin and non-transferrin bound iron by deferiprone. Implications in the treatment of thalassaemia, anaemia of chronic disease, cancer and other conditions. Hemoglobin 2006, 30, 183–200. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. Iron mobilization from ferritin using alpha-oxohydroxy heteroaromatic chelators. Biochem. J. 1986, 233, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Chambers, S.; Hoffbrand, A.V. Comparative study of iron mobilization from haemosiderin, ferritin and iron(III) precipitates by chelators. Biochem. J. 1987, 241, 87–92. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. Decreased solubilisation of ferritin iron and fresh iron (III) precipitate following repeated chelator treatments. Inorg. Chim. Acta 1987, 138, 36–40. [Google Scholar] [CrossRef]
- Mostert, L.J.; Van Dorst, J.A.L.M.; Koster, J.F.; Van Eijk, H.G.; Kontoghiorghes, G.J. Free radical and cytotoxic effects of chelators and their iron complexes in the hepatocyte. Free Rad. Res. Comms. 1987, 3, 379–388. [Google Scholar] [CrossRef]
- Jeremy, J.Y.; Gill, J.; Prior, T.; Sifaksi, G.; Barradas, M.A.; Kontoghiorghes, G.J. Inhibition of cycloxygenase and lipoxygenase activity by iron chelators: Possible use in the treatment of eicosanoid-related disorders. Drugs Today 1992, 28 (Suppl. SA), 35–43. [Google Scholar]
- Jeremy, J.Y.; Kontoghiorghes, G.J.; Hoffbrand, A.V.; Dandona, P. The iron chelators desferrioxamine and 1-alkyl-2-methyl-3-hydroxypyrid-4-ones inhibit vascular prostacyclin synthesis in vitro. Biochem. J. 1988, 254, 239–244. [Google Scholar] [CrossRef]
- Barradas, M.A.; Jeremy, J.Y.; Kontoghiorghes, G.J.; Mikhailidis, D.P.; Hoffbrand, A.V.; Dandona, P. Iron chelators inhibit human platelet aggregation, thromboxane A2 synthesis and lipoxygenase activity. FEBS Lett. 1989, 245, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Denisov, E.T.; Afanas’ev, I.B. Oxidation and Antioxidants in Organic Chemistry and Biology; CRC Press: Boca Raton, FL, USA; Taylor and Francis Group: Boca Raton, FL, USA, 2005. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C.; Cross, C.E. Free radicals, antioxidants and human disease: Where are we now? J. Lab. Clin. Med. 1992, 119, 598–620. [Google Scholar]
- Kontoghiorghes, G.J. Iron Chelation in Biochemistry and Medicine. In Free Radicals, Oxidant Stress and Drug Action; Rice-Evans, C., Ed.; Rechelieu Press: London, UK, 1987; pp. 277–303. [Google Scholar]
- Kontoghiorghes, G.J.; Kontoghiorghe, C.N. Prospects for the introduction of targeted antioxidant drugs for the prevention and treatment of diseases related to free radical pathology. Expert Opin. Investig. Drugs 2019, 28, 593–603. [Google Scholar] [CrossRef]
- Perron, N.R.; Brumaghim, J.L. A review of the antioxidant mechanisms of polyphenol compounds related to iron binding. Cell Biochem. Biophys. 2009, 53, 75–100. [Google Scholar] [CrossRef]
- Korkina, L.G.; Afanas’ev, I.B. Antioxidant and chelating properties of flavonoids. Adv. Pharmacol. 1997, 38, 151–163. [Google Scholar]
- Nkhili, E.; Loonis, M.; Mihai, S.; El Hajji, H.; Dangles, O. Reactivity of food phenols with iron and copper ions: Binding, dioxygen activation and oxidation mechanisms. Food Funct. 2014, 5, 1186–1220. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Jackson, M.I.; Lunec, J. In vitro screening of iron chelators using models of free radical damage. Free Rad. Res. Commun. 1986, 2, 115–124. [Google Scholar] [CrossRef]
- Hong, Y.L.; Pan, H.Z.; Scott, M.D.; Meshnick, S.R. Activated oxygen generation by a primaquine metabolite: Inhibition by antioxidants derived from Chinese herbal remedies. Free Radic. Biol. Med. 1992, 12, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.G.; Shibamoto, T. Antioxidant properties of aroma compounds isolated from soybeans and mung beans. J. Agric. Food Chem. 2000, 48, 4290–4293. [Google Scholar] [CrossRef] [PubMed]
- Wei, A.; Mura, K.; Shibamoto, T. Antioxidative activity of volatile chemicals extracted from beer. J. Agric. Food Chem. 2001, 49, 4097–4101. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Kim, H.Y.; Baek, S.H.; Yoo, H.H.; Park, J.H.; Yokozawa, T. Study on the hydroxyl radical scavenging activity changes of ginseng and ginsenoside-Rb2 by heat processing. Biol. Pharm. Bull. 2007, 30, 724–728. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jenner, A.M.; Lee, C.Y.J.; Shui, G.; Tang, S.Y.; Whiteman, M.; Wenk, M.R.; Halliwell, B. The identification of antioxidants in dark soy sauce. Free Radic. Res. 2007, 41, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Yamabe, N.; Kim, H.Y.; Yokozawa, T. Role of maltol in advanced glycation end products and free radicals: In-vitro and in-vivo studies. J. Pharm. Pharmacol. 2008, 60, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Minaiyan, M.; Mostaghel, E.; Mahzouni, P. Preventive Therapy of Experimental Colitis with Selected iron Chelators and Anti-oxidants. Int. J. Prev. Med. 2012, 3 (Suppl. S1), S162–S169. [Google Scholar] [PubMed] [PubMed Central]
- Han, Y.; Xu, Q.; Hu, J.N.; Han, X.Y.; Li, W.; Zhao, L.C. Maltol, a food flavoring agent, attenuates acute alcohol-induced oxidative damage in mice. Nutrients 2015, 7, 682–696. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Anwar-Mohamed, A.; El-Kadi, A.O. Induction of cytochrome P450 1a1 by the food flavoring agent, maltol. Toxicol. In Vitro 2007, 21, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, Z.; Hou, J.G.; Zhou, Y.D.; He, Y.F.; Jiang, S.; Wang, Y.P.; Ren, S.; Li, W. The Liver Protection Effects of Maltol, a Flavoring Agent, on Carbon Tetrachloride-Induced Acute Liver Injury in Mice via Inhibiting Apoptosis and Inflammatory Response. Molecules 2018, 23, 2120. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Guo, N.; Li, C.; Liu, Q.; Liu, S.; Huan, Y.; Wang, X.; Bai, G.; Yang, M.; Sun, S.; Xu, C.; et al. Maltol, a food flavor enhancer, attenuates diabetic peripheral neuropathy in streptozotocin-induced diabetic rats. Food Funct. 2018, 9, 6287–6297. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Hao, W.; Hu, J.; Mi, X.; Han, Y.; Ren, S.; Jiang, S.; Wang, Y.; Li, X.; Li, W. Maltol Improves APAP-Induced Hepatotoxicity by Inhibiting Oxidative Stress and Inflammation Response via NF-κB and PI3K/Akt Signal Pathways. Antioxidants 2019, 8, 395. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Singh, R.K.; Barrand, M.A. Lipid peroxidation effects of a novel iron compound, ferric maltol. A comparison with ferrous sulphate. J. Pharm. Pharmacol. 1990, 42, 276–279. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. Structure/red blood cell permeability activity of iron (III) chelator complexes. Inorg. Chim. Acta 1988, 151, 101–106. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Sheppard, L. Simple synthesis of the potent iron chelators 1-alkyl-3-hydroxy-2-methylpyrid-4-ones. Inorg. Chim. Acta Bioinorg. Chem. 1987, 136, L11–L12. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; May, A. Uptake and intracellular distribution of iron from transferrin and chelators in erythroid cells. Biol. Met. 1990, 3, 183–187. [Google Scholar] [CrossRef]
- Forsbeck, K.; Nilsson, K.; Kontoghiorghes, G.J. Variation in iron accumulation, transferrin membrane binding and DNA synthesis in the K-562 and U-937 cell lines induced by chelators and their iron complexes. Eur. J. Haematol. 1987, 39, 318–325. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Piga, A.; Hoffbrand, A.V. Cytotoxic and DNA-inhibitory effects of iron chelators on human leukaemic cell lines. Hematol. Oncol. 1986, 4, 195–204. [Google Scholar] [CrossRef]
- Neilands, J.B. Siderophores: Structure and function of microbial iron transport compounds. J. Biol. Chem. 1995, 270, 26723–26726. [Google Scholar] [CrossRef]
- Weinberg, E.D. Iron depletion: A defense against intracellular infection and neoplasm. Life Sci. 1992, 50, 1289–1297. [Google Scholar] [CrossRef]
- Andrews, S.C.; Robinson, A.K.; Rodríguez-Quiñones, F. Bacterial iron homeostasis. FEMS Microbiol. Rev. 2003, 27, 215–237. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Weinberg, E.D. Iron: Mammalian defense systems, mechanisms of disease, and chelation therapy approaches. Blood Rev. 1995, 9, 33–45. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kolnagou, A.; Skiada, A.; Petrikkos, G. The role of iron and chelators on infections in iron overload and non iron loaded conditions: Prospects for the design of new antimicrobial therapies. Hemoglobin 2010, 34, 227–239. [Google Scholar] [CrossRef]
- Robins-Browne, R.M.; Pipic, J.K. Desferrioxamine and systemic yersiniosis. Lancet 1983, II, 1372. [Google Scholar] [CrossRef]
- Boelaert, J.R.; Fenves, A.Z.; Coburn, J.W. Deferoxamine therapy and mucormycosis in dialysis patients: Report of an international registry. Am. J. Kidney Dis. 1991, 18, 660–667. [Google Scholar] [CrossRef]
- Wilson, B.R.; Bogdan, A.R.; Miyazawa, M.; Hashimoto, K.; Tsuji, Y. Siderophores in Iron Metabolism: From Mechanism to Therapy Potential. Trends Mol. Med. 2016, 22, 1077–1090. [Google Scholar] [CrossRef] [PubMed]
- Page, M.G.P. The Role of Iron and Siderophores in Infection, and the Development of Siderophore Antibiotics. Clin. Infect. Dis. 2019, 69 (Suppl. S7), S529–S537. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.S.; Edwards, J.E., Jr.; Fu, Y.; Spellberg, B. Deferiprone iron chelation as a novel therapy for experimental mucormycosis. J. Antimicrob. Chemother. 2006, 58, 1070–1073. [Google Scholar] [CrossRef] [PubMed]
- Brock, J.H.; Licéaga, J.; Kontoghiorghes, G.J. The effect of synthetic iron chelators on bacterial growth in human serum. FEMS Microbiol. Immunol. 1988, 1, 55–60. [Google Scholar] [CrossRef]
- Heppner, D.G.; Hallaway, P.E.; Kontoghiorghes, G.J.; Eaton, J.W. Antimalarial properties of orally active iron chelators. Blood 1988, 72, 358–361. [Google Scholar] [CrossRef]
- Brock, J.H.; Licéaga, J.; Arthur, H.M.; Kontoghiorghes, G.J. Effect of novel 1-alkyl-3-hydroxy-2-methylpyrid-4-one chelators on uptake and release of iron from macrophages. Am. J. Hematol. 1990, 34, 21–25. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Kleanthous, M.; Kontoghiorghe, C.N. The History of Deferiprone (L1) and the Paradigm of the Complete Treatment of Iron Overload in Thalassaemia. Mediterr. J. Hematol. Infect. Dis. 2020, 12, e2020011. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. Oral iron chelation is here. Br. Med. J. 1991, 303, 1279–1280. [Google Scholar] [CrossRef][Green Version]
- Kontoghiorghes, G.J. Chelators affecting iron absorption in mice. Drug Res. 1990, 40, 1332–1335. [Google Scholar]
- Barrand, M.A.; Callingham, B.A.; Hider, R.C. Effects of the pyrones, maltol and ethyl maltol, on iron absorption from the rat small intestine. J. Pharm. Pharmacol. 1987, 39, 203–211. [Google Scholar] [CrossRef]
- Barrand, M.A.; Callingham, B.A.; Dobbin, P.; Hider, R.C. Dissociation of a ferric maltol complex and its subsequent metabolism during absorption across the small intestine of the rat. Br. J. Pharmacol. 1991, 102, 723–729. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yamamoto, R.S.; Williams, G.M.; Frangel, H.H.; Weisburger, J.H. 8-hydroxyquinoline: Chronic toxicity and inhibitory effect on the carcinogenicity of N-2- fluorenylacetamide. Toxicol. Appl. Pharmacol. 1971, 19, 687–698. [Google Scholar] [CrossRef]
- Williams, G.M.; Yamamoto, R.S. Absence of stainable iron from preneoplastic and neoplastic lesions in rat liver with 8-hydroxyquinoline-induced siderosis. J. Natl. Cancer Inst. 1972, 49, 685–692. [Google Scholar] [PubMed]
- Hershko, C.; Konijn, A.M.; Nick, H.P.; Breuer, W.; Cabantchik, Z.I.; Link, G. ICL670A: A new synthetic oral chelator: Evaluation in hypertransfused rats with selective radioiron probes of hepatocellular and reticuloendothelial iron stores and in iron-loaded rat heart cells in culture. Blood 2001, 97, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Eghbali, A.; Shokri, P.; Afzal, R.R.; Bagheri, B. A 1-year randomized trial of deferasirox alone versus deferasirox and deferoxamine combination for the treatment of iron overload in thalassemia major. Transfus. Apher. Sci. 2019, 58, 429–433. [Google Scholar] [CrossRef]
- Karimi, M.; Haghpanah, S.; Bahoush, G.; Ansari, S.; Azarkeivan, A.; Shahsavani, A.; Bazrafshan, A.; Jangjou, A. Evaluation of Efficacy, Safety, and Satisfaction Taking Deferasirox Twice Daily Versus Once Daily in Patients With Transfusion-Dependent Thalassemia. J. Pediatr. Hematol. Oncol. 2020, 42, 23–26. [Google Scholar] [CrossRef]
- Pepe, A.; Meloni, A.; Pistoia, L.; Cuccia, L.; Gamberini, M.R.; Lisi, R.; D’Ascola, D.G.; Rosso, R.; Allò, M.; Spasiano, A.; et al. MRI multicentre prospective survey in thalassaemia major patients treated with deferasirox versus deferiprone and desferrioxamine. Br. J. Haematol. 2018, 183, 783–795. [Google Scholar] [CrossRef]
- Kolnagou, A.; Kleanthous, M.; Kontoghiorghes, G.J. Reduction of body iron stores to normal range levels in thalassaemia by using a deferiprone/deferoxamine combination and their maintenance thereafter by deferiprone monotherapy. Eur. J. Haematol. 2010, 85, 430–438. [Google Scholar] [CrossRef]
- Kolnagou, A.; Kontoghiorghe, C.N.; Kontoghiorghes, G.J. Prevention of Iron Overload and Long Term Maintenance of Normal Iron Stores in Thalassaemia Major Patients using Deferiprone or Deferiprone Deferoxamine Combination. Drug Res. 2017, 67, 404–411. [Google Scholar] [CrossRef]
- Lin, C.H.; Chen, X.; Wu, C.C.; Wu, K.H.; Song, T.S.; Weng, T.F.; Hsieh, Y.W.; Peng, C.T. Therapeutic mechanism of combined oral chelation therapy to maximize efficacy of iron removal in transfusion-dependent thalassemia major—A pilot study. Expert Rev. Hematol. 2019, 12, 265–272. [Google Scholar] [CrossRef]
- Boddaert, N.; Le Quan Sang, K.H.; Rötig, A.; Leroy-Willig, A.; Gallet, S.; Brunelle, F.; Sidi, D.; Thalabard, J.C.; Munnich, A.; Cabantchik, Z.I. Selective iron chelation in Friedreich ataxia: Biologic and clinical implications. Blood 2007, 110, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Martin-Bastida, A.; Ward, R.J.; Newbould, R.; Piccini, P.; Sharp, D.; Kabba, C.; Patel, M.C.; Spino, M.; Connelly, J.; Tricta, F.; et al. Brain iron chelation by deferiprone in a phase 2 randomised double-blinded placebo controlled clinical trial in Parkinson’s disease. Sci. Rep. 2017, 7, 1398. [Google Scholar] [CrossRef] [PubMed]
- Zorzi, G.; Zibordi, F.; Chiapparini, L.; Bertini, E.; Russo, L.; Piga, A.; Longo, F.; Garavaglia, B.; Aquino, D.; Savoiardo, M.; et al. Iron-related MRI images in patients with pantothenate kinase-associated neurodegeneration (PKAN) treated with deferiprone: Results of a phase II pilot trial. Mov. Disord. 2011, 26, 1756–1759. [Google Scholar] [CrossRef] [PubMed]
- Forni, G.L.; Balocco, M.; Cremonesi, L.; Abbruzzese, G.; Parodi, R.C.; Marchese, R. Regression of symptoms after selective iron chelation therapy in a case of neurodegeneration with brain iron accumulation. Mov. Disord. 2008, 23, 904–907. [Google Scholar] [CrossRef]
- Kolnagou, A.; Kleanthous, M.; Kontoghiorghes, G.J. Efficacy, compliance and toxicity factors are affecting the rate of normalization of body iron stores in thalassemia patients using the deferiprone and deferoxamine combination therapy. Hemoglobin 2011, 35, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Petry, N.; Egli, I.; Zeder, C.; Walczy, K.T.; Hurrell, R. Polyphenols and phytic acid contribute to the low iron bioavailability from common beans in young women. J. Nutr. 2010, 140, 1977–1982. [Google Scholar] [CrossRef]
- Sotelo, A.; González-Osnaya, L.; Sánchez-Chinchillas, A.; Trejo, A. Role of oxate, phytate, tannins and cooking on iron bioavailability from foods commonly consumed in Mexico. Int. J. Food Sci. Nutr. 2010, 61, 29–39. [Google Scholar] [CrossRef]
- Campbell, N.R.; Hasinoff, B.B. Iron supplements: A common cause of drug interactions. Br. J. Clin. Pharmacol. 1991, 31, 251–255. [Google Scholar] [CrossRef]
- Jaramillo, Á.; Briones, L.; Andrews, M.; Arredondo, M.; Olivares, M.; Brito, A.; Pizarro, F. Effect of phytic acid, tannic acid and pectin on fasting iron bioavailability both in the presence and absence of calcium. J. Trace Elem. Med. Biol. 2015, 30, 112–117. [Google Scholar] [CrossRef]
- Kojima, Y.; Yoshikawa, Y.; Ueda, E.; Kondo, M.; Takahashi, S.; Matsukura, T.; Sakurai, H.; Hiroi, T.; Imaoka, S.; Funae, Y. Blood glucose lowering and toxicological effects of zinc (II) complexes with maltol, threonine, and picolinic acid. Res. Commun. Mol. Pathol. Pharmacol. 2002, 112, 91–104. [Google Scholar] [PubMed]
- Wang, H.; Shao, B.; Yu, H.; Xu, F.; Wang, P.; Yu, K.; Han, Y.; Song, M.; Li, Y.; Cao, Z. Neuroprotective role of hyperforin on aluminum maltolate-induced oxidative damage and apoptosis in PC12 cells and SH-SY5Y cells. Chem. Biol. Interact. 2019, 299, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Ott, D.B.; Hartwig, A.; Stillman, M.J. Competition between Al3+ and Fe3+ binding to human transferrin and toxicological implications: Structural investigations using ultra-high resolution ESI MS and CD spectroscopy. Metallomics 2019, 11, 968–981. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Barr, J.; Baillod, R.A. Studies of aluminium mobilisation in renal dialysis patients using the oral chelator 1, 2-dimethyl-3-hydroxypyrid-4-one. Arzn Forsch Drug Res. 1994, 44, 522–526. [Google Scholar]
- Pergola, P.E.; Fishbane, S.; Ganz, T. Novel Oral Iron Therapies for Iron Deficiency Anemia in Chronic Kidney Disease. Adv. Chronic. Kidney Dis. 2019, 26, 272–291. [Google Scholar] [CrossRef] [PubMed]
- Murawska, N.; Fabisiak, A.; Fichna, J. Anemia of Chronic Disease and Iron Deficiency Anemia in Inflammatory Bowel Diseases: Pathophysiology, Diagnosis, and Treatment. Inflamm. Bowel Dis. 2016, 22, 1198–1208. [Google Scholar] [CrossRef] [PubMed]
- Khoury, A.; Pagan, K.A.; Farland, M.Z. Ferric Maltol: A New Oral Iron Formulation for the Treatment of Iron Deficiency in Adults. Ann. Pharmacother. 2021, 55, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Stallmach, A.; Büning, C. Ferric maltol (ST10): A novel oral iron supplement for the treatment of iron deficiency anemia in inflammatory bowel disease. Expert Opin. Pharmacother. 2015, 16, 2859–2867. [Google Scholar] [CrossRef] [PubMed]
- Bokemeyer, B.; Krummenerl, A.; Maaser, C.; Howaldt, S.; Mroß, M.; Mallard, N. Randomized Open-Label Phase 1 Study of the Pharmacokinetics of Ferric Maltol in Inflammatory Bowel Disease Patients with Iron Deficiency. Eur. J. Drug Metab. Pharmacokinet. 2017, 42, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.S.; Reffitt, D.M.; Doig, L.A.; Meenan, J.; Ellis, R.D.; Thompson, R.P.; Powell, J.J. Ferric trimaltol corrects iron deficiency anaemia in patients intolerant of iron. Aliment Pharmacol. Ther. 1998, 12, 845–848. [Google Scholar] [CrossRef]
- Kelsey, S.M.; Hider, R.; Bloor, J.R.C.; Blake, D.R.; Gutteridge, C.N.; Newland, A.C. Absorption of low and therapeutic doses of ferric maltol, a novel ferric iron compound, in iron deficient subjects using a single dose iron absorption test. J. Clin. Pharm. Ther. 1991, 16, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Gasche, C.; Ahmad, T.; Tulassay, Z.; Baumgart, D.C.; Bokemeyer, B.; Büning, C.; Howaldt, S.; Stallmach, A.; AEGIS Study Group. Ferric maltol is effective in correcting iron deficiency anemia in patients with inflammatory bowel disease: Results from a phase-3 clinical trial program. Inflamm. Bowel Dis. 2015, 21, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Reffitt, D.M.; Burden, T.J.; Seed, P.T.; Wood, J.; Thompson, R.P.; Powell, J.J. Assessment of iron absorption from ferric trimaltol. Ann. Clin. Biochem. 2000, 37, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.; Ahmad, T.; Tulassay, Z.; Baumgart, D.C.; Bokemeyer, B.; Howaldt, S.; Stallmach, A.; Büning, C.; The AEGIS Study Group. Ferric maltol therapy for iron deficiency anaemia in patients with inflammatory bowel disease: Long-term extension data from a Phase 3 study. Aliment. Pharmacol Ther. 2016, 44, 259–270. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Olsson, K.M.; Fuge, J.; Brod, T.; Kamp, J.C.; Schmitto, J.; Kempf, T.; Bauersachs, J.; Hoeper, M.M. Oral iron supplementation with ferric maltol in patients with pulmonary hypertension. Eur. Respir. J. 2020, 56, 2000616. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kopyt, N.P.; AEGIS-CKD Study Group. Efficacy and safety of oral ferric maltol (FM) in treating iron-deficiency anemia (IDA) in patients with CKD: Randomized controlled trial [FR-OR120]. J. Am. Soc. Nephrol. 2018, 29, 70–71. [Google Scholar]
- Punj, S.; Ghafourian, K.; Ardehali, H. Iron deficiency and supplementation in heart failure and chronic kidney disease. Mol. Aspects Med. 2020, 75, 100873. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nataatmadja, M.S.; Francis, R. Recurrent severe hypophosphatemia following intravenous iron administration. Clin. Case Rep. 2020, 8, 243–246. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kshirsagar, A.V.; Li, X. Long-Term Risks of Intravenous Iron in End-Stage Renal Disease Patients. Adv. Chronic Kidney Dis. 2019, 26, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Liabeuf, S.; Gras, V.; Moragny, J.; Laroche, M.L.; Andrejak, M.; French National Network of Pharmacovigilance Centers. Ulceration of the oral mucosa following direct contact with ferrous sulfate in elderly patients: A case report and a review of the French National Pharmacovigilance Database. Clin. Interv. Aging. 2014, 9, 737–740. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ferrari, P.; Nicolini, A.; Manca, M.L.; Rossi, G.; Anselmi, L.; Conte, M.; Carpi, A.; Bonino, F. Treatment of mild non-chemotherapy-induced iron deficiency anemia in cancer patients: Comparison between oral ferrous bisglycinate chelate and ferrous sulfate. Biomed. Pharmacother. 2012, 66, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Schümann, K.; Kroll, S.; Weiss, G.; Frank, J.; Biesalski, H.K.; Daniel, H.; Friel, J.; Solomons, N.W. Monitoring of hematological, inflammatory and oxidative reactions to acute oral iron exposure in human volunteers: Preliminary screening for selection of potentially-responsive biomarkers. Toxicology 2005, 212, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.H.; Barta, C.A.; Orvig, C. Metal complexes of maltol and close analogues in medicinal inorganic chemistry. Chem. Soc. Rev. 2006, 35, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J.; Efstathiou, A.; Ioannou-Loucaides, S.; Kolnagou, A. Chelators controlling metal metabolism and toxicity pathways: Applications in cancer prevention, diagnosis and treatment. Hemoglobin 2008, 32, 217–227. [Google Scholar] [CrossRef]
- Barve, A.; Kumbhar, A.; Bhat, M.; Joshi, B.; Butcher, R.; Sonawane, U.; Joshi, R. Mixed-ligand copper(II) maltolate complexes: Synthesis, characterization, DNA binding and cleavage, and cytotoxicity. Inorg. Chem. 2009, 48, 9120–9132. [Google Scholar] [CrossRef] [PubMed]
- Chukwuma, C.I.; Mashele, S.S.; Eze, K.C.; Matowane, G.R.; Islam, S.M.; Bonnet, S.L.; Noreljaleel, A.E.M.; Ramorobi, L.M. A comprehensive review on zinc(II) complexes as anti-diabetic agents: The advances, scientific gaps and prospects. Pharmacol. Res. 2020, 155, 104744. [Google Scholar] [CrossRef] [PubMed]
- Kostrzewa, A.; Páli, T.; Froncisz, W.; Marsh, D. Membrane location of spin-labeled cytochrome c determined by paramagnetic relaxation agents. Biochemistry 2000, 39, 6066–6074. [Google Scholar] [CrossRef] [PubMed]
- Kandioller, W.; Hartinger, C.G.; Nazarov, A.A.; Bartel, C.; Skocic, M.; Jakupec, M.A.; Arion, V.B.; Keppler, B.K. Maltol-derived ruthenium-cymene complexes with tumor inhibiting properties: The impact of ligand-metal bond stability on anticancer activity in vitro. Chemistry 2009, 15, 12283–12291. [Google Scholar] [CrossRef] [PubMed]
- Mehtab, S.; Gonçalves, G.; Roy, S.; Tomaz, A.I.; Santos-Silva, T.; Santos, M.F.A.; Romão, M.J.; Jakusch, T.; Kiss, T.; Pessoa, J.C. Interaction of vanadium(IV) with human serum apo-transferrin. J. Inorg. Biochem. 2013, 121, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Satoh, E.; Yasuda, I.; Yamada, T.; Suzuki, Y.; Ohyashiki, T. Involvement of NO generation in aluminum-induced cell death. Biol. Pharm. Bull. 2007, 30, 1390–1394. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, E.; Nakano, K.; Nakayachi, T.; Morshed, S.R.M.; Hashimoto, K.; Kikuchi, H.; Nishikawa, H.; Kawase, M.; Sakagami, H. Cytotoxic activity of deferiprone, maltol and related hydroxyketones against human tumor cell lines. Anticancer. Res. 2004, 24, 755–762. [Google Scholar] [PubMed]
- Chitambar, C.R. The therapeutic potential of iron-targeting gallium compounds in human disease: From basic research to clinical application. Pharmacol. Res. 2017, 115, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Li, Y.; Zhang, J.; Zhou, Y.; Li, L.; Xue, X.; Li, H.; Niu, Q. Role of mGluR 1 in synaptic plasticity impairment induced by maltol aluminium in rats. Environ. Toxicol. Pharmacol. 2020, 78, 103406. [Google Scholar] [CrossRef] [PubMed]
- Enyedy, É.A.; Dömötör, O.; Bali, K.; Hetényi, A.; Tuccinardi, T.; Keppler, B.K. Interaction of the anticancer gallium(III) complexes of 8-hydroxyquinoline and maltol with human serum proteins. J. Biol. Inorg. Chem. 2015, 20, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, T.W.; Lessmann, G.M.; Saleh, J.; Liu, X.; Chitambar, C.R.; Hwang, S.T. Gallium maltolate inhibits human cutaneous T-cell lymphoma tumor development in mice. J. Investig. Dermatol. 2015, 135, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Chitambar, C.R.; Al-Gizawiy, M.M.; Alhajala, H.S.; Pechman, K.R.; Wereley, J.P.; Wujek, R.; Clark, P.A.; Kuo, J.S.; Antholine, W.E.; Schmainda, K.M. Gallium Maltolate Disrupts Tumor Iron Metabolism and Retards the Growth of Glioblastoma by Inhibiting Mitochondrial Function and Ribonucleotide Reductase. Mol. Cancer Ther. 2018, 17, 1240–1250. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Merli, D.; Profumo, A.; Bloise, N.; Risi, G.; Momentè, S.; Cucca, L.; Visai, L. Indium/Gallium Maltolate Effects on Human Breast Carcinoma Cells: In Vitro Investigation on Cytotoxicity and Synergism with Mitoxantrone. ACS Omega 2018, 3, 4631–4640. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bernstein, L.R.; van der Hoeven, J.J.; Boer, R.O. Hepatocellular carcinoma detection by gallium scan and subsequent treatment by gallium maltolate: Rationale and case study. Anticancer Agents Med. Chem. 2011, 11, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Arnold, C.E.; Bordin, A.; Lawhon, S.D.; Libal, M.C.; Bernstein, L.R.; Cohen, N.D. Antimicrobial activity of gallium maltolate against Staphylococcus aureus and methicillin-resistant S. aureus and Staphylococcus pseudintermedius: An in vitro study. Vet. Microbiol. 2012, 155, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Fecteau, M.E.; Aceto, H.W.; Bernstein, L.R.; Sweeney, R.W. Comparison of the antimicrobial activities of gallium nitrate and gallium maltolate against Mycobacterium avium subsp. paratuberculosis in vitro. Vet. J. 2014, 202, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Hijazi, S.; Visaggio, D.; Pirolo, M.; Frangipani, E.; Bernstein, L.; Visca, P. Antimicrobial Activity of Gallium Compounds on ESKAPE Pathogens. Front. Cell Infect. Microbiol. 2018, 8, 316. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ball, K.R.; Sampieri, F.; Chirino, M.; Hamilton, D.L.; Blyth, R.I.; Sham, T.K.; Dowling, P.M.; Thompson, J. Synchrotron X-ray fluorescence microscopy of gallium in bladder tissue following gallium maltolate administration during urinary tract infection. Antimicrob Agents Chemother. 2013, 57, 5197–5201. [Google Scholar] [CrossRef] [PubMed] [PubMed Central][Green Version]
- Monk, C.S.; Sweeney, R.W.; Bernstein, L.R.; Fecteau, M.E. Serum and tissue concentrations of gallium after oral administration of gallium nitrate and gallium maltolate to neonatal calves. Am. J. Vet. Res. 2016, 77, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Sampieri, F.; Alcorn, J.; Allen, A.L.; Clark, C.R.; Vannucci, F.A.; Pusterla, N.; Mapes, S.; Ball, K.R.; Dowling, P.M.; Thompson, J.; et al. Pharmacokinetics of gallium maltolate in Lawsonia intracellularis-infected and uninfected rabbits. J. Vet. Pharmacol. Ther. 2014, 37, 486–499. [Google Scholar] [CrossRef] [PubMed]
- Pollina, G.F.; Pepe, M.; Dean, A.; Di Marco, V.; Marton, D. Reduction in absorption of gallium maltolate in adult horses following oral administration with food: Chemistry and pharmacokinetics. J. Vet. Pharmacol. Ther. 2013, 36, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Martens, R.J.; Mealey, K.; Cohen, N.D.; Harrington, J.R.; Chaffin, M.K.; Taylor, R.J.; Bernstein, L.R. Pharmacokinetics of gallium maltolate after intragastric administration in neonatal foals. Am. J. Vet. Res. 2007, 68, 1041–1044. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.R. Molecular mechanisms of iron uptake by cells and the use of iron chelators for the treatment of cancer. Curr. Med. Chem. 2005, 12, 2711–2729. [Google Scholar] [CrossRef] [PubMed]
- Markowicz-Piasecka, M.; Skupień, A.; Mikiciuk-Olasik, E.; Sikora, J. Biocompatibility Studies of Gadolinium Complexes with Iminodiacetic Acid Derivatives. Biol. Trace Elem. Res. 2019, 189, 426–436. [Google Scholar] [CrossRef]
- Parghane, R.V.; Basu, S. Bilateral Orbital Soft-Tissue Metastases from Renal Neuroendocrine Tumor: Successful Theranostic Application of 68Ga/177Lu-DOTATATE with Improvement of Vision. J. Nucl. Med. Technol. 2019, 47, 171–172. [Google Scholar] [CrossRef]
- Rund, D. Intravenous iron: Do we adequately understand the short- and long-term risks in clinical practice? Br. J. Haematol. 2020. [Google Scholar] [CrossRef]
- Auerbach, M.; Gafter-Gvili, A.; Macdougall, I.C. Intravenous iron: A framework for changing the management of iron deficiency. Lancet Haematol. 2020, 7, e342–e350. [Google Scholar] [CrossRef] [PubMed]
- Manz, D.H.; Blanchette, N.L.; Paul, B.T.; Torti, F.M.; Torti, S.V. Iron and cancer: Recent insights. Ann. N. Y. Acad. Sci. 2016, 1368, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Voest, E.E.; Vreugdenhil, G.; Marx, J.J.M. Iron-chelating agents in non-iron overload conditions. Ann. Intern. Med. 1994, 120, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.E.; Weeks, K.; Carter, D.A. Lactoferrin Is Broadly Active against Yeasts and Highly Synergistic with Amphotericin B. Antimicrob Agents Chemother. 2020, 64, e02284-19. [Google Scholar] [CrossRef]
- Rosa, L.; Cutone, A.; Lepanto, M.S.; Paesano, R.; Valenti, P. Lactoferrin: A Natural Glycoprotein Involved in Iron and Inflammatory Homeostasis. Int. J. Mol. Sci. 2017, 18, 1985. [Google Scholar] [CrossRef]
- González-Chávez, S.A.; Arévalo-Gallegos, S.; Rascón-Cruz, Q. Lactoferrin: Structure, function and applications. Int. J. Antimicrob. Agents 2009, 33, 301.e1–301.e8. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Quantitative aspects The quantity of iron present in the diet, e.g., excess iron intake in African siderosis or insufficient iron intake in vegetarian populations |
| Qualitative aspects The form of dietary iron present in food, e.g., haem, ferrous, ferric, ferritin, hemosiderin |
| Role of reducing agents The presence of reducing agents, e.g., ascorbic acid converts Fe (III) to Fe (II) and increases iron absorption |
| Effect of dietary molecules The presence of dietary molecules with chelating properties, e.g., phytates and tannins decrease iron absorption |
| Effect of drugs with chelating properties The presence of drugs with chelating properties, e.g., deferiprone, deferoxamine, tetracycline, hydroxyurea inhibit iron absorption |
| Effect of fluids The quantity of water, alcohol, and other fluid intake can influence the dissolution of iron supplements or other components in the gastrointestinal tract |
| Effect of molecules affecting cellular iron transport Dietary molecules and drugs affecting iron transport across the enterocyte (e.g., Nifedipine, which is an L-type calcium channel blocker) |
| Dietary factors affecting iron solubilisation Dietary factors affecting the solubilization or precipitation of iron. Insoluble iron is not readily absorbed |
| Effect of pH on iron solubilisation pH of the stomach and intestine, e.g., the higher the pH the lower the solubility of iron; Antacids decrease iron absorption |
| Anatomical changes and iron absorption Gastrectomy and other surgical interventions, which can affect gastrointestinal iron absorption. Body weight, e.g., obese people absorb less iron than normal body weight individuals |
| Effect of diseases on iron absorption Malignancy, infectious and other diseases, haemoglobinopathies |
| Changes in iron absorption levels Malnutrition, food poisoning, ageing |
| Iron deficiency anaemia due to increased iron requirements (e.g., pregnant and menstruating women, young children) |
| Iron deficiency anaemia due to insufficient dietary iron (e.g., vegetarian populations, malnutrition) |
| Anaemia of chronic disease or anaemia of inflammation, in neoplastic, infectious and inflammatory diseases, mainly in cases with concurrent iron deficiency or in cases of combination with erythropoietin treatment (e.g., Inflammatory bowel disease, cancer, rheumatoid arthritis) |
| Chronic kidney disease including haemodialysis patients |
| Chronic cardiac failure |
| Ferrous iron formulations (Ferrous sulphate, ferrous ascorbate, ferrous fumarate, ferrous gluconate, ferroglycine sulphate |
| Ferric iron formulations (Ferric fumarate, ferric polymaltose, iron dextran, ferric gluconate, ferric iron sucrose, ferric saccharate, iron bis-glycinate chelate |
| Ferric intravenous iron formulations (Iron sucrose, ferric carboxymaltose, ferric gluconate, ferumoxytol, iron isomaltoside-1000, iron dextran (low-molecular-weight forms) |
| Lipophilic (hetero)aromatic iron complex formulations (Ferric maltol, ferric 8-hydroxyquinoline, ferric tropolone, ferric 2-hydroxy-4-methoxypyridine-1-oxide (L6) |
| Chelator | Log β | MWt | Kpar | Charge | Kpar Iron | Charge Complex |
|---|---|---|---|---|---|---|
| Maltol | 30 | 126 | 1.23 | neutral | 0.32 | neutral |
| Tropolone | 32 | 122 | 3.04 | neutral | 4.50 | neutral |
| 8-Hydroxyquinoline | 37 | 145 | 28.30 | neutral | 10.00 | neutral |
| L3 | 30 | 127 | 0.09 | zwitterionic | 0.04 | neutral |
| L4 | NA | 111 | 0.09 | zwitterionic | 0.95 | neutral |
| L6 | 29 | 155 | 0.37 | zwitterionic | 4.85 | neutral |
| Omadine | NA | 127 | 0.04 | zwitterionic | 2.67 | neutral |
| Mimosine | 36 | 198 | 0.01 | zwitterionic | 0.01 | zwitterionic |
| Deferoxamine | 31 | 561 | 0.02 | positive | 0.02 | positive |
| Deferiprone | 35 | 139 | 0.18 | neutral | 0.05 | neutral |
| Deferasirox | 27 | 373 | 6.30 | negative | NA | negative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kontoghiorghes, G.J.; Kolnagou, A.; Demetriou, T.; Neocleous, M.; Kontoghiorghe, C.N. New Era in the Treatment of Iron Deficiency Anaemia Using Trimaltol Iron and Other Lipophilic Iron Chelator Complexes: Historical Perspectives of Discovery and Future Applications. Int. J. Mol. Sci. 2021, 22, 5546. https://doi.org/10.3390/ijms22115546
Kontoghiorghes GJ, Kolnagou A, Demetriou T, Neocleous M, Kontoghiorghe CN. New Era in the Treatment of Iron Deficiency Anaemia Using Trimaltol Iron and Other Lipophilic Iron Chelator Complexes: Historical Perspectives of Discovery and Future Applications. International Journal of Molecular Sciences. 2021; 22(11):5546. https://doi.org/10.3390/ijms22115546
Chicago/Turabian StyleKontoghiorghes, George J., Annita Kolnagou, Theodora Demetriou, Marina Neocleous, and Christina N. Kontoghiorghe. 2021. "New Era in the Treatment of Iron Deficiency Anaemia Using Trimaltol Iron and Other Lipophilic Iron Chelator Complexes: Historical Perspectives of Discovery and Future Applications" International Journal of Molecular Sciences 22, no. 11: 5546. https://doi.org/10.3390/ijms22115546
APA StyleKontoghiorghes, G. J., Kolnagou, A., Demetriou, T., Neocleous, M., & Kontoghiorghe, C. N. (2021). New Era in the Treatment of Iron Deficiency Anaemia Using Trimaltol Iron and Other Lipophilic Iron Chelator Complexes: Historical Perspectives of Discovery and Future Applications. International Journal of Molecular Sciences, 22(11), 5546. https://doi.org/10.3390/ijms22115546