
Human Genomics and the Biocultural Origin of Music

 , , and
, , and
Abstract
1. Introduction
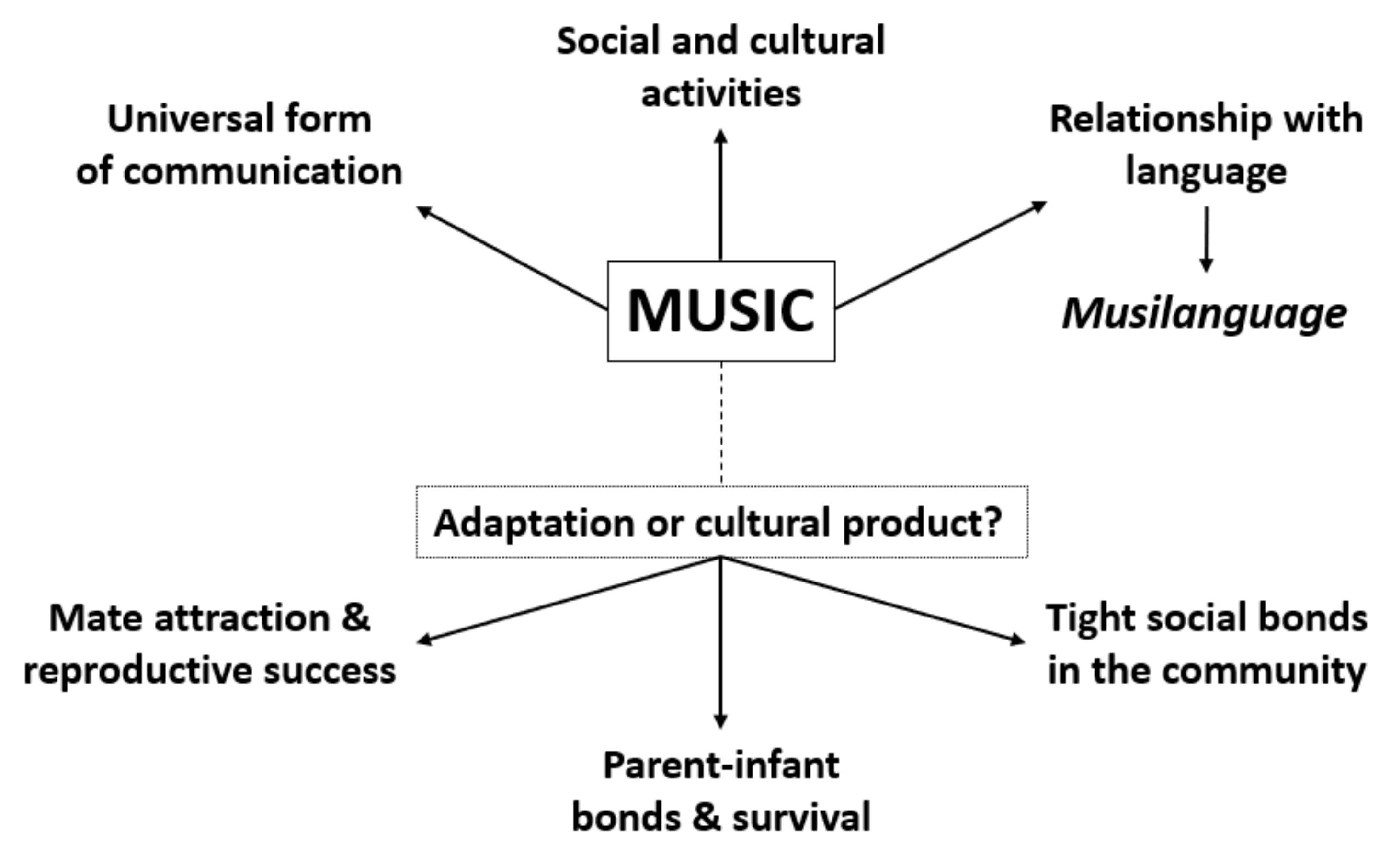
2. Music–Language Universality and the Biocultural Origins of Music
3. Comparative and Cross-Species Behavioral Studies
4. Twin Studies, Heritability and Hints to the Neuropsychology of Music
5. Genetic and Genomic Candidates for Musicality in Humans
6. The Effect of Music on Gene Expression and Brain Activity
7. Concluding Remarks and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Honing, H. On the Biological Basis of Musicality. Ann. N. Y. Acad. Sci. 2018, 1423, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Savage, P.E.; Brown, S.; Sakai, E.; Currie, T.E. Statistical Universals Reveal the Structures and Functions of Human Music. Proc. Natl. Acad. Sci. USA 2015, 112, 8987–8992. [Google Scholar] [CrossRef]
- Brown, S.; Jordania, J. Universals in the World’s Musics. Psychol. Music 2013, 4, 229–248. [Google Scholar] [CrossRef]
- Karma, K. The Ability to Structure Acoustic Material as a Measure of Musical Aptitude: I. Background Theory and Pilot Studies. Res. Bull. 38 1973. [Google Scholar]
- Merker, B.H.; Madison, G.S.; Eckerdal, P. On the Role and Origin of Isochrony in Human Rhythmic Entrainment. Cortex 2009, 45, 4–17. [Google Scholar] [CrossRef]
- Merker, B.; Morley, I.; Zuidema, W. Five Fundamental Constraints on Theories of the Origins of Music. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370, 20140095. [Google Scholar] [CrossRef] [PubMed]
- Fitch, W.T. Four Principles of Bio-Musicology. Philos. Trans. R. Soc. Lond. B. Biol. Sci 2015, 370, 20140091. [Google Scholar] [CrossRef] [PubMed]
- Marler, P. Origins of Music and Speech: Insights from Animals. In The Origins of Music; Wallin, N.L., Brown, S., Merker, B., Eds.; MIT Press: Cambridge, MA, USA, 2000; pp. 31–48. [Google Scholar]
- Honing, H.; ten Cate, C.; Peretz, I.; Trehub, S.E. Without It No Music: Cognition, Biology and Evolution of Musicality. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370, 20140088. [Google Scholar] [CrossRef]
- Morley, I. The Prehistory of Music: Human Evolution, Archaeology, and the Origins of Musicality; Oxford University Press: Oxford, UK, 2013. [Google Scholar] [CrossRef]
- Conard, N.J.; Malina, M.; Münzel, S.C. New Flutes Document the Earliest Musical Tradition in Southwestern Germany. Nature 2009, 460, 737–740. [Google Scholar] [CrossRef]
- Higham, T.; Basell, L.; Jacobi, R.; Wood, R.; Ramsey, C.B.; Conard, N.J. Τesting Models for the Beginnings of the Aurignacian and the Advent of Figurative Art and Music: The Radiocarbon Chronology of Geißenklösterle. J. Hum. Evol. 2012, 62, 664–676. [Google Scholar] [CrossRef]
- Turk, M.; Košir, A. Mousterian osseous artefacts? The case of Divje babe I., Slovenia. Quat. Int. 2017, 450, 103–115. [Google Scholar] [CrossRef]
- Turk, M.; Turk, I.; Dimkaroski, L.; Blackwell, B.A.B.; Zoltan Horusitzky, F.; Otte, M.; Bastiani, G.; Korat, L. Divje babe I cave (Slovenia): Arguments on the Material Evidence for Neanderthal Musical Behaviour. Anthropologie 2018, 122, 679–706. [Google Scholar] [CrossRef]
- Turk, M.; Turk, I.; Otte, M. The Neanderthal Musical Instrument from Divje Babe I Cave (Slovenia): A Critical Review of the Discussion. Appl. Sci. 2020, 10, 1226. [Google Scholar] [CrossRef]
- Fritz, C.; Tosello, G.; Fleury, G.; Kasarhérou, E.; Walter, P.; Duranthon, F.; Gaillard, P.; Tardieu, J. First Record of the Sound Produced by the Oldest Upper Paleolithic Seashell Horn. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef]
- Molino, J.; Nattiez, J.-J. L’unità della musica. In Enciclopedia Della Musica; Nattiez, J.-J., Ed.; V, Einaudi: Torino, Italy, 2005; Volume 5, pp. 331–366. [Google Scholar]
- Harvey, A. Music, Evolution, and the Harmony of Souls; Oxford University Press: Oxford, UK, 2017. [Google Scholar] [CrossRef][Green Version]
- Harvey, A.R. Music and the Meeting of Human Minds. Front. Psychol. 2018, 9, 762. [Google Scholar] [CrossRef]
- Oesch, N. Music and Language in Social Interaction: Synchrony, Antiphony, and Functional Origins. Front. Psychol. 2019, 10, 1514. [Google Scholar] [CrossRef]
- Brown, S. The “Musilanguage” Model of Music Evolution. In The Origins of Music; Wallin, N.L., Brown, S., Merker, B., Eds.; MIT Press: Cambridge, MA, USA, 2000; pp. 271–300. [Google Scholar]
- Brown, S. A Joint Prosodic Origin of Language and Music. Front. Psychol. 2017, 8, 1894. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Martinez, M.J.; Parsons, L.M. Music and Language Side by Side in the Brain: A PET Study of the Generation of Melodies and Sentences. Eur. J. Neurosci. 2006, 23, 2791–2803. [Google Scholar] [CrossRef]
- Koelsch, S. Toward a Neural Basis of Music Perception—A Review and Updated Model. Front. Psychol. 2011, 2, 110. [Google Scholar] [CrossRef] [PubMed]
- Trehub, S.E.; Becker, J.; Morley, I. Cross-Cultural Perspectives on Music and Musicality. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370, 20140096. [Google Scholar] [CrossRef] [PubMed]
- Ravignani, A. Darwin, Sexual Selection, and the Origins of Music. Trends. Ecol. Evol. 2018, 33, 716–719. [Google Scholar] [CrossRef]
- Ravignani, A.; Madison, G. The Paradox of Isochrony in the Evolution of Human Rhythm. Front. Psychol. 2017, 8, 1820. [Google Scholar] [CrossRef]
- Darwin, C. The Descent of Man, and Selection in Relation to Sex; John Murray: London, UK, 1981. [Google Scholar]
- Marin, M.M.; Gingras, B.; Bhattacharya, J. Crossmodal Transfer of Arousal, but Not Pleasantness, from the Musical to the Visual Domain. Emotion 2012, 12, 618–631. [Google Scholar] [CrossRef]
- Marin, M.M.; Schober, R.; Gingras, B.; Leder, H. Misattribution of Musical Arousal Increases Sexual Attraction towards Opposite-Sex Faces in Females. PLoS ONE 2017, 12, e0183531. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.; Zhang, L.; Ou, Y.; Zhang, X. The Audio-Visual Integration Effect on Music Emotion: Behavioral and Physiological Evidence. PLoS ONE 2019, 14, e0217040. [Google Scholar] [CrossRef]
- Trehub, S. Human Processing Predispositions and Musical Universals. In The Origins of Music; Wallin, N.L., Brown, S., Merker, B., Eds.; MIT Press: Cambridge, MA, USA, 2000; pp. 427–448. [Google Scholar]
- Dissanayake, E. If Music Is the Food of Love, What about Survival and Reproductive Success? Musicae Sci. 2008, 12, 169–195. [Google Scholar] [CrossRef]
- Fischer, R.; Callander, R.; Reddish, P.; Bulbulia, J. How Do Rituals Affect Cooperation? An Experimental Field Study Comparing Nine Ritual Types. Hum. Nat. 2013, 24, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Salimpoor, V.N.; Benovoy, M.; Larcher, K.; Dagher, A.; Zatorre, R.J. Anatomically Distinct Dopamine Release during Anticipation and Experience of Peak Emotion to Music. Nat. Neurosci. 2011, 14, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Rilling, J.; Gutman, D.; Zeh, T.; Pagnoni, G.; Berns, G.; Kilts, C. A Neural Basis for Social Cooperation. Neuron 2002, 35, 395–405. [Google Scholar] [CrossRef]
- Rilling, J.K.; King-Casas, B.; Sanfey, A.G. The Neurobiology of Social Decision-Making. Curr. Opin. Neurobiol. 2008, 18, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, L.; Mas-Herrero, E.; Zatorre, R.J.; Ripolles, P.; Gomez-Andres, A.; Alicart, H.; Olivé, G.; Marco-Pallarés, J.; Antonjioan, R.M.; Valle, M.; et al. Dopamine Modulates the Reward Experiences Elicited by Music. Proc. Natl. Acad. Sci. USA 2019, 116, 3793–3798. [Google Scholar] [CrossRef] [PubMed]
- Goupil, L.; Aucouturier, J.J. Musical Pleasure and Musical Emotions. Proc. Natl. Acad. Sci. USA 2019, 116, 3364–3366. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.R. Links Between the Neurobiology of Oxytocin and Human Musicality. Front. Hum. Neurosci. 2020, 14, 350. [Google Scholar] [CrossRef]
- Marsh, N.; Marsh, A.A.; Lee, M.R.; Hurlemann, R. Oxytocin and the Neurobiology of Prosocial Behavior. Neuroscientist 2020. [Google Scholar] [CrossRef]
- Cardona, G.; Rodriguez-Fornells, A.; Nye, H.; Rifà-Ros, X.; Ferreri, L. The Impact of Musical Pleasure and Musical Hedonia on Verbal Episodic Memory. Sci. Rep. 2020, 10, 16113. [Google Scholar] [CrossRef] [PubMed]
- Gold, B.P.; Pearce, M.T.; Mas-Herrero, E.; Dagher, A.; Zatorre, R.J. Predictability and Uncertainty in the Pleasure of Music: A Reward for Learning? J. Neurosci. 2019, 39, 9397–9409. [Google Scholar] [CrossRef] [PubMed]
- Belfi, A.M.; Loui, P. Musical Anhedonia and Rewards of Music Listening: Current Advances and A Proposed Model. Ann. N. Y. Acad. Sci. 2020, 1464, 99–114. [Google Scholar] [CrossRef]
- Trainor, L.J. The Origins of Music in Auditory Scene Analysis and the Roles of Evolution and Culture in Musical Creation. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370, 20140089. [Google Scholar] [CrossRef]
- Mehr, S.A.; Singh, M.; Knox, D.; Ketter, D.M.; Pickens-Jones, D.; Atwood, S.; Lucas, C.; Jacoby, N.; Egner, A.A.; Hopkins, E.J.; et al. Universality and Diversity in Human Song. Science 2019, 366, eaax0868. [Google Scholar] [CrossRef]
- Kotler, J.; Mehr, S.A.; Egner, A.; Haig, D.; Krasnow, M.M. Response to Vocal Music in Angelman Syndrome Contrasts with Prader-Willi Syndrome. Evol. Hum. Behav. 2019, 40, 420–426. [Google Scholar] [CrossRef]
- Reybrouck, M., Podlipniak. Preconceptual Spectral and Temporal Cues as a Source of Meaning in Speech and Music. Brain Sci. 2019, 9, 53. [Google Scholar] [CrossRef]
- Malloch, S., Trevarthen. The Human Nature of Music. Front. Psychol. 2018, 9, 1680. [Google Scholar] [CrossRef]
- Podlipniak, P. The Role of the Baldwin Effect in the Evolution of Human Musicality. Front Neurosci. 2017, 11, 542. [Google Scholar] [CrossRef] [PubMed]
- Podlipniak, P. The Role of Canalization and Plasticity in the Evolution of Musical Creativity. Front Neurosci. 2021, 15, 607887. [Google Scholar] [CrossRef] [PubMed]
- Hoeschele, M.; Merchant, H.; Kikuchi, Y.; Hattori, Y.; ten Cate, C. Searching for the Origins of Musicality across Species. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370, 20140094. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.D.; Iversen, J.R.; Bregman, M.R.; Schulz, I. Experimental Evidence for Synchronization to a Musical Beat in a Nonhuman Animal. Curr. Biol. 2009, 19, 827–830. [Google Scholar] [CrossRef] [PubMed]
- Cook, P.; Rouse, A.; Wilson, M.; Reichmuth, C. A California Sea Lion (Zalophus Californianus) Can Keep the Beat: Motor Entrainment to Rhythmic Auditory Stimuli in a Non Vocal Mimic. J. Comp. Psychol. 2013, 127, 412–427. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Tomonaga, M.; Matsuzawa, T. Spontaneous Synchronized Tapping to an Auditory Rhythm in a Chimpanzee. Sci. Rep. 2013, 3, 1566. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Tomonaga, M.; Matsuzawa, T. Distractor Effect of Auditory Rhythms on Self-Paced Tapping in Chimpanzees and Humans. PLoS ONE 2015, 10, e0130682. [Google Scholar] [CrossRef]
- Hattori, Y.; Tomonaga, M. Rhythmic Swaying Induced by Sound in Chimpanzees (Pan Troglodytes). Proc. Natl. Acad. Sci. USA 2020, 117, 936–942. [Google Scholar] [CrossRef]
- Large, E.W.; Gray, P.M. Spontaneous Tempo and Rhythmic Entrainment in a Bonobo (Pan Paniscus). J. Comp. Psychol. 2015, 129, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Repp, B.H. Sensorimotor Synchronization: A Review of the Tapping Literature. Psychon. Bull. Rev. 2005, 12, 969–992. [Google Scholar] [CrossRef] [PubMed]
- Repp, B.H.; Su, Y.-H. Sensorimotor Synchronization: A Review of Recent Research (2006–2012). Psychon. Bull. Rev. 2013, 20, 403–452. [Google Scholar] [CrossRef]
- Fujioka, T.; Trainor, L.J.; Large, E.W.; Ross, B. Internalized Timing of Isochronous Sounds Is Represented in Neuromagnetic β Oscillations. J. Neurosci. 2012, 32, 1791–1802. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.D. The Evolutionary Biology of Musical Rhythm: Was Darwin Wrong? PLoS Biol. 2014, 12, e1001821. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.; Cook, P.F. Rhythmic Entrainment: Why Humans Want to, Fireflies Can’t Help It, Pet Birds Try, and Sea Lions Have to Be Bribed. Psychon. Bull. Rev. 2016, 23, 1647–1659. [Google Scholar] [CrossRef]
- Patel, A. Musical Rhythm, Linguistic Rhythm, and Human Evolution. Music Percept. 2006, 24, 99–104. [Google Scholar] [CrossRef]
- Zarco, W.; Merchant, H.; Prado, L.; Mendez, J.C. Subsecond Timing in Primates: Comparison of Interval Production between Human Subjects and Rhesus Monkeys. J. Neurophysiol. 2009, 102, 3191–3202. [Google Scholar] [CrossRef]
- Merchant, H.; Pérez, O.; Bartolo, R.; Méndez, J.C.; Mendoza, G.; Gámez, J.; Yc, K.; Prado, L. Sensorimotor Neural Dynamics during Isochronous Tapping in the Medial Premotor Cortex of the Macaque. Eur. J. Neurosci. 2015, 41, 586–602. [Google Scholar] [CrossRef]
- Honing, H.; Bouwer, F.L.; Prado, L.; Merchant, H. Rhesus Monkeys (Macaca Mulatta) Sense Isochrony in Rhythm, but Not the Beat: Additional Support for the Gradual Audiomotor Evolution Hypothesis. Front. Neurosci. 2018, 12, 475. [Google Scholar] [CrossRef] [PubMed]
- Merchant, H.; Honing, H. Are Non-Human Primates Capable of Rhythmic Entrainment? Evidence for the Gradual Audiomotor Evolution Hypothesis. Front. Neurosci. 2014, 7, 274. [Google Scholar] [CrossRef] [PubMed]
- Goodall, J. The Chimpanzees of Gombe: Patterns of Behavior; Belknap Press of Harvard University Press: Boston, MA, USA, 1986. [Google Scholar]
- Grahn, J.A.; Brett, M. Rhythm and Beat Perception in Motor Areas of the Brain. J. Cogn. Neurosci. 2007, 19, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.D.; Iversen, J.R. The Evolutionary Neuroscience of Musical Beat Perception: The Action Simulation for Auditory Prediction (ASAP) Hypothesis. Front. Syst. Neurosci. 2014, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Merchant, H.; Grahn, J.; Trainor, L.; Rohrmeier, M.; Fitch, W.T. Finding the Beat: A Neural Perspective across Humans and Non-Human Primates. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370, 20140093. [Google Scholar] [CrossRef] [PubMed]
- Mosing, M.A.; Verwein, K.J.H.; Madison, G.; Pedersen, N.L.; Pietsch, B.P.; Ullen, F. Did Sexual Selection Shape Human Music? Testing Predictions from the Sexual Selection Hypothesis of Music Evolution Using a Large Genetically Informative Sample of Over 10,000 Twins. Evol. Hum. Behav. 2015, 5, 359–366. [Google Scholar] [CrossRef]
- Wesseldijk, L.; Gordon, R.L.; Mosing, M.; Ullén, F. Music and Verbal Ability—A Twin Study of Genetic and Environmental Associations. Psychol. Aesthet. Creat. Arts. in press. [CrossRef]
- Gustavson, D.E.; Friedman, N.P.; Stallings, M.; Reynolds, C.A.; Coon, H.; Corley, R.P.; Hewitt, J.K.; Gordon, R.L. Musical Instrument Engagement in Adolescence Predicts Verbal Ability Four Years Later: A Twin and Adoption Study. PsyArXiv 2021. [Google Scholar] [CrossRef]
- Centanni, T.M.; Anchan, D.M.; Beard, M.; Brooks, R.; Thompson, L.A.; Petrill, S.A. Genetic and Environmental Influences on Decoding Skills—Implications for Music and Reading. Front. Psychol. 2019, 10, 2604. [Google Scholar] [CrossRef]
- Cohrdes, C.; Grolig, L.; Schroeder, S. Relating Language and Music Skills in Young Children: A First Approach to Systemize and Compare Distinct Competencies on Different Levels. Front. Psychol. 2016, 7, 1616. [Google Scholar] [CrossRef] [PubMed]
- Politimou, N.; Dalla Bella, S.; Farrugia, N.; Franco, F. Born to Speak and Sing: Musical Predictors of Language Development in Pre-schoolers. Front. Psychol. 2019, 10, 948. [Google Scholar] [CrossRef]
- Swaminathan, S.; Schellenberg, E.G. Musical Ability, Music Training, and Language Ability in Childhood. J. Exp. Psychol. Learn. Mem. Cogn. 2020, 46, 2340–2348. [Google Scholar] [CrossRef]
- Sallat, S.; Jentschke, S. Music Perception Influences Language Acquisition: Melodic and Rhythmic-Melodic Perception in Children with Specific Language Impairment. Behav. Neurol. 2015, 2015, 606470. [Google Scholar] [CrossRef]
- Pfeifer, J.; Hamann, S. The Nature and Nurture of Congenital Amusia: A Twin Case Study. Front. Behav. Neurosci. 2018, 12, 120. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Huang, H.; Haponenko, H.; Sun, H.J. Face Recognition and Memory in Congenital Amusia. PLoS ONE 2019, 14, e0225519. [Google Scholar] [CrossRef]
- Jasmin, K.; Dick, F.; Stewart, L.; Tierney, A.T. Altered Functional Connectivity During Speech Perception in Congenital Amusia. eLife 2020, 9, e53539. [Google Scholar] [CrossRef] [PubMed]
- Jasmin, K.; Dick, F.; Holt, L.L.; Tierney, A. Tailored Perception: Individuals’ Speech and Music Perception Strategies Fit Their Perceptual Abilities. J. Exp. Psychol. Gen. 2020, 149, 914–934. [Google Scholar] [CrossRef]
- Jasmin, K.; Sun, H.; Tierney, A.T. Effects of Language Experience on Domain-General Perceptual Strategies. Cognition 2021, 206, 104481. [Google Scholar] [CrossRef]
- Kunert, R.; Willems, R.M.; Casasanto, D.; Patel, A.D.; Hagoort, P. Music and Language Syntax Interact in Broca’s Area: An fMRI Study. PLoS ONE 2015, 4, e0141069. [Google Scholar] [CrossRef]
- Chiang, J.N.; Rosenberg, M.H.; Bufford, C.A.; Stephens, D.; Lysy, A.; Monti, M.M. The Language of Music: Common Neural Codes for Structured Sequences in Music and Natural Language. Brain Lang. 2018, 185, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Peretz, I. The Biological Foundations of Music: Insights from Congenital Amusia. In The Psychology of Music, 3rd ed.; Deutsch, D., Ed.; Elsevier: San Diego, CA, USA, 2013; pp. 551–564. [Google Scholar] [CrossRef]
- Müllensiefen, D.; Gingras, B.; Musil, J.; Stewart, L. The Musicality of Non-Musicians: An Index for Assessing Musical Sophistication in the General Population. PLoS ONE 2014, 9, e89642. [Google Scholar] [CrossRef]
- Gingras, B.; Honing, H.; Peretz, I.; Trainor, L.J.; Fisher, S.E. Defining the Biological Bases of Individual Differences in Musicality. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370, 20140092. [Google Scholar] [CrossRef]
- Järvelä, I. Genomics Studies on Musical Aptitude, Music Perception, and Practice. Ann. N. Y. Acad. Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S. Initial Impact of the Sequencing of the Human Genome. Nature 2011, 470, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Pulli, K.; Karma, K.; Norio, R.; Sistonen, P.; Göring, H.H.H.; Järvelä, I. Genome-Wide Linkage Scan for Loci of Musical Aptitude in Finnish Families: Evidence for a Major Locus at 4q22. J. Med. Genet. 2008, 45, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Ukkola, L.T.; Onkamo, P.; Raijas, P.; Karma, K.; Järvelä, I. Musical Aptitude Is Associated with AVPR1A-Haplotypes. PLoS ONE 2009, 4, e5534. [Google Scholar] [CrossRef] [PubMed]
- Ukkola-Vuoti, L.; Oikkonen, J.; Onkamo, P.; Karma, K.; Raijas, P.; Järvelä, I. Association of the Arginine Vasopressin Receptor 1A (AVPR1A) Haplotypes with Listening to Music. J. Hum. Genet. 2011, 56, 324–329. [Google Scholar] [CrossRef]
- Ukkola-Vuoti, L.; Kanduri, C.; Oikkonen, J.; Buck, G.; Blancher, C.; Raijas, P.; Karma, K.; Lähdesmäki, H.; Järvelä, I. Genome-Wide Copy Number Variation Analysis in Extended Families and Unrelated Individuals Characterized for Musical Aptitude and Creativity in Music. PLoS ONE 2013, 8, e56356. [Google Scholar] [CrossRef]
- Morley, A.P.; Narayanan, M.; Mines, R.; Molokhia, A.; Baxter, S.; Craig, G.; Lewis, C.M.; Craig, I. AVPR1A and SLC6A4 Polymorphisms in Choral Singers and Non-Musicians: A Gene Association Study. PLoS ONE 2012, 7, e31763. [Google Scholar] [CrossRef]
- Park, H.; Lee, S.; Kim, H.-J.; Ju, Y.S.; Shin, J.-Y.; Hong, D.; von Grotthuss, M.; Lee, D.-S.; Park, C.; Kim, J.H.; et al. Comprehensive Genomic Analyses Associate UGT8 Variants with Musical Ability in a Mongolian Population. J. Med. Genet. 2012, 49, 747–752. [Google Scholar] [CrossRef]
- Oikkonen, J.; Huang, Y.; Onkamo, P.; Ukkola-Vuoti, L.; Raijas, P.; Karma, K.; Vieland, V.J.; Järvelä, I. A Genome-Wide Linkage and Association Study of Musical Aptitude Identifies Loci Containing Genes Related to Inner Ear Development and Neurocognitive Functions. Mol. Psychiatry 2015, 20, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Kanduri, C.; Oikkonen, J.; Karma, K.; Raijas, P.; Ukkola-Vuoti, L.; Teo, Y.-Y.; Järvelä, I. Detecting Signatures of Positive Selection Associated with Musical Aptitude in the Human Genome. Sci. Rep. 2016, 6, 21198. [Google Scholar] [CrossRef]
- Mariath, L.M.; da Silva, A.M.; Kowalski, T.W.; Gattino, G.S.; de Araujo, G.A.; Figueiredo, F.G.; Tagliani-Ribeiro, A.; Roman, T.; Vianna, F.S.L.; Schuler-Faccini, L.; et al. Music Genetics Research: Association with Musicality of a Polymorphism in the AVPR1A Gene. Genet. Mol. Biol. 2017, 40, 421–429. [Google Scholar] [CrossRef]
- Bachner-Melman, R.; Dina, C.; Zohar, A.H.; Constantini, N.; Lerer, E.; Hoch, S.; Sella, S.; Nemanov, L.; Gritsenko, I.; Lichtenberg, P.; et al. AVPR1a and SLC6A4 Gene Polymorphisms Are Associated with Creative Dance Performance. PLoS Genet. 2005, 1, e42. [Google Scholar] [CrossRef]
- Fink, S.; Excoffier, L.; Heckel, G. High Variability and Non-Neutral Evolution of the Mammalian Avpr1a Gene. BMC Evol. Biol. 2007, 7, 176. [Google Scholar] [CrossRef]
- Knafo, A.; Israel, S.; Darvasi, A.; Bachner-Melman, R.; Uzefovsky, F.; Cohen, L.; Feldman, E.; Lerer, E.; Laiba, E.; Raz, Y.; et al. Individual Differences in Allocation of Funds in the Dictator Game Associated with Length of the Arginine Vasopressin 1a Receptor RS3 Promoter Region and Correlation between RS3 Length and Hippocampal MRNA. Genes Brain Behav. 2008, 7, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Wassink, T.H.; Piven, J.; Vieland, V.J.; Pietila, J.; Goedken, R.J.; Folstein, S.E.; Sheffield, V.C. Examination of AVPR1a as an Autism Susceptibility Gene. Mol. Psychiatry 2004, 9, 968–972. [Google Scholar] [CrossRef] [PubMed]
- Granot, R.; Frankel, Y.; Gritsenko, V.; Lerer, E.; Gritsenko, I.; Bachner-Melman, R.; Israel, S.; Ebstein, R. Provisional Evidence that the Arginine Vasopressin 1a Receptor Gene Is Associated with Musical Memory. Evol. Hum. Behav. 2007, 28, 313–318. [Google Scholar] [CrossRef]
- Lesch, K.P.; Bengel, D.; Heils, A.; Sabol, S.Z.; Greenberg, B.D.; Petri, S.; Benjamin, J.; Müller, C.R.; Hamer, D.H.; Murphy, D.L. Association of Anxiety-Related Traits with a Polymorphism in the Serotonin Transporter Gene Regulatory Region. Science 1996, 274, 1527–1531. [Google Scholar] [CrossRef] [PubMed]
- Kremer, I.; Bachner-Melman, R.; Reshef, A.; Broude, L.; Nemanov, L.; Gritsenko, I.; Heresco-Levy, U.; Elizur, Y.; Ebstein, R.P. Association of the Serotonin Transporter Gene with Smoking Behavior. Am. J. Psychiatry 2005, 162, 924–930. [Google Scholar] [CrossRef]
- Sarosi, A.; Gonda, X.; Balogh, G.; Domotor, E.; Szekely, A.; Hejjas, K.; Sasvari-Szekely, M.; Faludi, G. Association of the STin2 Polymorphism of the Serotonin Transporter Gene with a Neurocognitive Endophenotype in Major Depressive Disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 1667–1672. [Google Scholar] [CrossRef]
- Garcia, L.F.; Aluja, A.; Fibla, J.; Cuevas, L.; García, O. Incremental Effect for Antisocial Personality Disorder Genetic Risk Combining 5-HTTLPR and 5-HTTVNTR Polymorphisms. Psychiatry Res. 2010, 177, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Oikkonen, J.; Onkamo, P.; Järvelä, I.; Kanduri, C. Convergent Evidence for the Molecular Basis of Musical Traits. Sci. Rep. 2016, 6, 39707. [Google Scholar] [CrossRef]
- Stein, E.; Tessier-Lavigne, M. Hierarchical Organization of Guidance Receptors: Silencing of Netrin Attraction by Slit through a Robo/DCC Receptor Complex. Science 2001, 291, 1928–1938. [Google Scholar] [CrossRef]
- Williams, J.; O’Donovan, M.C. The Genetics of Developmental Dyslexia. Eur. J. Hum. Genet. 2006, 14, 681–689. [Google Scholar] [CrossRef]
- Dugas, J.C.; Tai, Y.C.; Speed, T.P.; Ngai, J.; Barres, B.A. Functional Genomic Analysis of Oligodendrocyte Differentiation. J. Neurosci. 2006, 26, 10967–10983. [Google Scholar] [CrossRef]
- Xiao, H.; Sun, Z.; Wan, J.; Hou, S.; Xiong, Y. Overexpression of Protocadherin 7 Inhibits Neuronal Survival by Downregulating BIRC5 in Vitro. Exp. Cell Res. 2018, 366, 71–80. [Google Scholar] [CrossRef]
- Lin, J.; Yan, X.; Wang, C.; Guo, Z.; Rolfs, A.; Luo, J. Anatomical Expression Patterns of Delta-Protocadherins in Developing Chicken Cochlea. J. Anat. 2012, 221, 598–608. [Google Scholar] [CrossRef] [PubMed]
- Hertel, N.; Redies, C.; Medina, L. Cadherin Expression Delineates the Divisions of the Postnatal and Adult Mouse Amygdala. J. Comp. Neurol. 2012, 520, 3982–4012. [Google Scholar] [CrossRef]
- Gosselin, N.; Peretz, I.; Johnsen, E.; Adolphs, R. Amygdala Damage Impairs Emotion Recognition from Music. Neuropsychologia 2007, 45, 236–244. [Google Scholar] [CrossRef]
- Fisher, S.E.; Francks, C.; Marlow, A.J.; MacPhie, I.L.; Newbury, D.F.; Cardon, L.R.; Ishikawa-Brush, Y.; Richardson, A.J.; Talcott, J.B.; Gayán, J.; et al. Independent Genome-Wide Scans Identify a Chromosome 18 Quantitative-Trait Locus Influencing Dyslexia. Nat. Genet. 2002, 30, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, Y.; Hirabayashi, T.; Katori, S.; Kawamura, Y.; Sanbo, M.; Hirabayashi, M.; Kiyonari, H.; Nakao, K.; Uchimura, A.; Yagi, T. Total Expression and Dual Gene-Regulatory Mechanisms Maintained in Deletions and Duplications of the Pcdha Cluster. J. Biol. Chem. 2009, 284, 32002–32014. [Google Scholar] [CrossRef] [PubMed]
- Katori, S.; Hamada, S.; Noguchi, Y.; Fukuda, E.; Yamamoto, T.; Yamamoto, H.; Hasegawa, S.; Yagi, T. Protocadherin-Alpha Family Is Required for Serotonergic Projections to Appropriately Innervate Target Brain Areas. J. Neurosci. 2009, 29, 9137–9147. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, E.; Hamada, S.; Hasegawa, S.; Katori, S.; Sanbo, M.; Miyakawa, T.; Yamamoto, T.; Yamamoto, H.; Hirabayashi, T.; Yagi, T. Down-Regulation of Protocadherin-Alpha a Isoforms in Mice Changes Contextual Fear Conditioning and Spatial Working Memory. Eur. J. Neurosci. 2008, 28, 1362–1376. [Google Scholar] [CrossRef] [PubMed]
- Theusch, E.; Basu, A.; Gitschier, J. Genome-Wide Study of Families with Absolute Pitch Reveals Linkage to 8q24.21 and Locus Heterogeneity. Am. J. Hum. Genet. 2009, 85, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Niarchou, M.; Gustavson, D.E.; Fah Sathirapongsasuti, J.; Anglada-Tort, M.; Eising, E.; Bell, E.; McArthur, E.; Straub, P.; The 23andMe Research Team; Devin McAuley, J.; et al. Unravelling the genetic architecture of musical rhythm: A large-scale genome-wide association study of beat synchronization. bioRxiv 836197. [CrossRef]
- Blakely, R.D.; Ramamoorthy, S.; Schroeter, S.; Qian, Y.; Apparsundaram, S.; Galli, A.; DeFelice, L.J. Regulated Phosphorylation and Trafficking of Antidepressant-Sensitive Serotonin Transporter Proteins. Biol. Psychiatry 1998, 44, 169–178. [Google Scholar] [CrossRef]
- Rodrigues, A.C.; Loureiro, M.A.; Caramelli, P. Long-Term Musical Training May Improve Different Forms of Visual Attention Ability. Brain Cogn. 2013, 82, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Kanduri, C.; Kuusi, T.; Ahvenainen, M.; Philips, A.K.; Lähdesmäki, H.; Järvelä, I. The Effect of Music Performance on the Transcriptome of Professional Musicians. Sci. Rep. 2015, 5, 9506. [Google Scholar] [CrossRef]
- Pfenning, A.R.; Hara, E.; Whitney, O.; Rivas, M.V.; Wang, R.; Roulhac, P.L.; Howard, J.T.; Wirthlin, M.; Lovell, P.V.; Ganapathy, G.; et al. Convergent Transcriptional Specializations in the Brains of Humans and Song-Learning Birds. Science 2014, 346, 1256846. [Google Scholar] [CrossRef]
- Kimpo, R.R.; Doupe, A.J. FOS Is Induced by Singing in Distinct Neuronal Populations in a Motor Network. Neuron 1997, 18, 315–325. [Google Scholar] [CrossRef]
- Polymeropoulos, M.H.; Lavedan, C.; Leroy, E.; Ide, S.E.; Dehejia, A.; Dutra, A.; Pike, B.; Root, H.; Rubenstein, J.; Boyer, R.; et al. Mutation in the Alpha-Synuclein Gene Identified in Families with Parkinson’s Disease. Science 1997, 276, 2045–2047. [Google Scholar] [CrossRef] [PubMed]
- Spillantini, M.G.; Goedert, M. The Alpha-Synucleinopathies: Parkinson’s Disease, Dementia with Lewy Bodies, and Multiple System Atrophy. Ann. N. Y. Acad. Sci. 2000, 920, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Al-Wandi, A.; Ninkina, N.; Millership, S.; Williamson, S.J.M.; Jones, P.A.; Buchman, V.L. Absence of Alpha-Synuclein Affects Dopamine Metabolism and Synaptic Markers in the Striatum of Aging Mice. Neurobiol. Aging 2010, 31, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Scherzer, C.R.; Grass, J.A.; Liao, Z.; Pepivani, I.; Zheng, B.; Eklund, A.C.; Ney, P.A.; Ng, J.; McGoldrick, M.; Mollenhauer, B.; et al. GATA Transcription Factors Directly Regulate the Parkinson’s Disease-Linked Gene Alpha-Synuclein. Proc. Natl. Acad. Sci. USA 2008, 105, 10907–10912. [Google Scholar] [CrossRef]
- Haugas, M.; Lilleväli, K.; Hakanen, J.; Salminen, M. Gata2 Is Required for the Development of Inner Ear Semicircular Ducts and the Surrounding Perilymphatic Space. Dev. Dyn. 2010, 239, 2452–2469. [Google Scholar] [CrossRef]
- Lahti, L.; Achim, K.; Partanen, J. Molecular Regulation of GABAergic Neuron Differentiation and Diversity in the Developing Midbrain. Acta Physiol. (Oxf.) 2013, 207, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Vernes, S.C.; Spiteri, E.; Nicod, J.; Groszer, M.; Taylor, J.M.; Davies, K.E.; Geschwind, D.H.; Fisher, S.E. High-Throughput Analysis of Promoter Occupancy Reveals Direct Neural Targets of FOXP2, a Gene Mutated in Speech and Language Disorders. Am. J. Hum. Genet. 2007, 81, 1232–1250. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.S.; Fisher, S.E.; Hurst, J.A.; Vargha-Khadem, F.; Monaco, A.P. A Forkhead-Domain Gene Is Mutated in a Severe Speech and Language Disorder. Nature 2001, 413, 519–523. [Google Scholar] [CrossRef]
- Haesler, S.; Wada, K.; Nshdejan, A.; Morrisey, E.E.; Lints, T.; Jarvis, E.D.; Scharff, C. FoxP2 Expression in Avian Vocal Learners and Non-Learners. J. Neurosci. 2004, 24, 3164–3175. [Google Scholar] [CrossRef] [PubMed]
- Teramitsu, I.; White, S.A. FoxP2 Regulation during Undirected Singing in Adult Songbirds. J. Neurosci. 2006, 26, 7390–7394. [Google Scholar] [CrossRef]
- Zatorre, R.J.; Salimpoor, V.N. From Perception to Pleasure: Music and Its Neural Substrates. Proc. Natl. Acad. Sci. USA 2013, 110 (Suppl. 2), 10430–10437. [Google Scholar] [CrossRef]
- Zatorre, R.J.; Chen, J.L.; Penhune, V.B. When the Brain Plays Music: Auditory-Motor Interactions in Music Perception and Production. Nat. Rev. Neurosci. 2007, 8, 547–558. [Google Scholar] [CrossRef]
- Herholz, S.C.; Zatorre, R.J. Musical Training as a Framework for Brain Plasticity: Behavior, Function, and Structure. Neuron 2012, 76, 486–502. [Google Scholar] [CrossRef]
- Gaser, C.; Schlaug, G. Brain Structures Differ between Musicians and Non-Musicians. J. Neurosci. 2003, 23, 9240–9245. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, K.; Williamon, A.; Rothwell, J.C. Motorcortical Excitability and Synaptic Plasticity Is Enhanced in Professional Musicians. J. Neurosci. 2007, 27, 5200–5206. [Google Scholar] [CrossRef] [PubMed]
- Schlaug, G.; Norton, A.; Overy, K.; Winner, E. Effects of Music Training on the Child’s Brain and Cognitive Development. Ann. N. Y. Acad. Sci. 2005, 1060, 219–230. [Google Scholar] [CrossRef]
- Sluming, V.; Brooks, J.; Howard, M.; Downes, J.J.; Roberts, N. Broca’s Area Supports Enhanced Visuospatial Cognition in Orchestral Musicians. J. Neurosci. 2007, 27, 3799–3806. [Google Scholar] [CrossRef]
- Kanduri, C.; Raijas, P.; Ahvenainen, M.; Philips, A.K.; Ukkola-Vuoti, L.; Lähdesmäki, H.; Järvelä, I. The Effect of Listening to Music on Human Transcriptome. PeerJ 2015, 3, e830. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.S.; Kuusi, T.; Ahvenainen, M.; Philips, A.K.; Järvelä, I. Music-Performance Regulates MicroRNAs in Professional Musicians. PeerJ 2019, 7, e6660. [Google Scholar] [CrossRef]
- Nair, P.S.; Raijas, P.; Ahvenainen, M.; Philips, A.K.; Ukkola-Vuoti, L.; Järvelä, I. Music-Listening Regulates Human MicroRNA Expression. Epigenetics 2020, 1–13. [Google Scholar] [CrossRef]
- Glatt, S.J.; Everall, I.P.; Kremen, W.S.; Corbeil, J.; Sásik, R.; Khanlou, N.; Han, M.; Liew, C.-C.; Tsuang, M.T. Comparative Gene Expression Analysis of Blood and Brain Provides Concurrent Validation of SELENBP1 Up-Regulation in Schizophrenia. Proc. Natl. Acad. Sci. USA 2005, 102, 15533–15538. [Google Scholar] [CrossRef]
- Liew, C.-C.; Ma, J.; Tang, H.-C.; Zheng, R.; Dempsey, A.A. The Peripheral Blood Transcriptome Dynamically Reflects System Wide Biology: A Potential Diagnostic Tool. J. Lab. Clin. Med. 2006, 147, 126–132. [Google Scholar] [CrossRef]
- George, J.M.; Jin, H.; Woods, W.S.; Clayton, D.F. Characterization of a Novel Protein Regulated during the Critical Period for Song Learning in the Zebra Finch. Neuron 1995, 15, 361–372. [Google Scholar] [CrossRef]
- Clayton, D.F.; George, J.M. The Synucleins: A Family of Proteins Involved in Synaptic Function, Plasticity, Neurodegeneration and Disease. Trends Neurosci. 1998, 21, 249–254. [Google Scholar] [CrossRef]
- Horita, H.; Kobayashi, M.; Liu, W.-C.; Oka, K.; Jarvis, E.D.; Wada, K. Specialized Motor-Driven Dusp1 Expression in the Song Systems of Multiple Lineages of Vocal Learning Birds. PLoS ONE 2012, 7, e42173. [Google Scholar] [CrossRef]
- Chen, Q.; Heston, J.B.; Burkett, Z.D.; White, S.A. Expression Analysis of the Speech-Related Genes FoxP1 and FoxP2 and Their Relation to Singing Behavior in Two Songbird Species. J. Exp. Biol. 2013, 216, 3682–3692. [Google Scholar] [CrossRef]
- Gunaratne, P.H.; Lin, Y.-C.; Benham, A.L.; Drnevich, J.; Coarfa, C.; Tennakoon, J.B.; Creighton, C.J.; Kim, J.H.; Milosavljevic, A.; Watson, M.; et al. Song Exposure Regulates Known and Novel MicroRNAs in the Zebra Finch Auditory Forebrain. BMC Genom. 2011, 12, 277. [Google Scholar] [CrossRef]
- Wang, J.; Lu, M.; Qiu, C.; Cui, Q. TransmiR: A Transcription Factor-MicroRNA Regulation Database. Nucleic Acids Res. 2010, 38, D119–D122. [Google Scholar] [CrossRef]
- Shi, Z.; Luo, G.; Fu, L.; Fang, Z.; Wang, X.; Li, X. MiR-9 and MiR-140-5p Target FoxP2 and Are Regulated as a Function of the Social Context of Singing Behavior in Zebra Finches. J. Neurosci. 2013, 33, 16510–16521. [Google Scholar] [CrossRef]
- Drnevich, J.; Replogle, K.L.; Lovell, P.; Hahn, T.P.; Johnson, F.; Mast, T.G.; Nordeen, E.; Nordeen, K.; Strand, C.; London, S.E.; et al. Impact of Experience-Dependent and -Independent Factors on Gene Expression in Songbird Brain. Proc. Natl. Acad. Sci. USA 2012, 109 (Suppl. 2), 17245–17252. [Google Scholar] [CrossRef] [PubMed]
- Heston, J.B.; White, S.A. Behavior-Linked FoxP2 Regulation Enables Zebra Finch Vocal Learning. J. Neurosci. 2015, 35, 2885–2894. [Google Scholar] [CrossRef]
- Liu, X.; Cannon, D.M.; Akula, N.; Moya, P.R.; Knudsen, G.M.; Arentzen, T.E.; Steele, J.; Laje, G.; Drevets, W.C.; McMahon, F.J. A Non-Synonymous Polymorphism in Galactose Mutarotase (GALM) Is Associated with Serotonin Transporter Binding Potential in the Human Thalamus: Results of a Genome-Wide Association Study. Mol. Psychiatry 2011, 16, 584–585. [Google Scholar] [CrossRef] [PubMed]
- Szyfter, K.; Witt, M.P. How Far Musicality and Perfect Pitch Are Derived from Genetic Factors? J. Appl. Genet. 2020, 61, 407–414. [Google Scholar] [CrossRef]
- Atkinson, E.G.; Audesse, A.J.; Palacios, J.A.; Bobo, D.M.; Webb, A.E.; Ramachandran, S.; Henn, B.M. No Evidence for Recent Selection at FOXP2 among Diverse Human Populations. Cell 2018, 174, 1424.e15–1435.e15. [Google Scholar] [CrossRef] [PubMed]
- Furlong, R. FOXP2 Tells a Cautionary Tale. Nat. Rev. Genet. 2018, 19, 592–593. [Google Scholar] [CrossRef]
- Fisher, S.E. Human Genetics: The Evolving Story of FOXP2. Curr. Biol. 2019, 29, R65–R67. [Google Scholar] [CrossRef] [PubMed]
- Mosing, M.A.; Madison, G.; Pedersen, N.L.; Ullén, F. Investigating Cognitive Transfer Within the Framework of Music Practice: Genetic Pleiotropy Rather than Causality. Dev. Sci. 2016, 19, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Wesseldijk, L.W.; Mosing, M.A.; Ullén, F. Gene-Environment Interaction in Expertise: The Importance of Childhood Environment for Musical Achievement. Dev. Psychol. 2019, 55, 1473–1479. [Google Scholar] [CrossRef]
- Wesseldijk, L.W.; Mosing, M.A.; Ullén, F. Why Is an Early Start of Training Related to Musical Skills in Adulthood? A Genetically Informative Study. Psychol. Sci. 2021, 32, 3–13. [Google Scholar] [CrossRef]
- Whitney, O.; Pfenning, A.R.; Howard, J.T.; Blatti, C.A.; Liu, F.; Ward, J.M.; Wang, R.; Audet, J.-N.; Kellis, M.; Mukherjee, S.; et al. Core and Region-Enriched Networks of Behaviorally Regulated Genes and the Singing Genome. Science 2014, 346, 1256780. [Google Scholar] [CrossRef] [PubMed]
- Wassiliwizky, E.; Koelsch, S.; Wagner, V.; Jacobsen, T.; Menninghaus, W. The Emotional Power of Poetry: Neural Circuitry, Psychophysiology and Compositional Principles. Soc. Cogn. Affect. Neurosci. 2017, 12, 1229–1240. [Google Scholar] [CrossRef]
- Zaidel, D.W. Coevolution of Language and Symbolic Meaning: Co-Opting Meaning Underlying the Initial Arts in Early Human Culture. Wiley Interdiscip. Rev. Cogn. Sci. 2020, 11, e1520. [Google Scholar] [CrossRef] [PubMed]
- Basso, J.C.; Satyal, M.K.; Rugh, R. Dance on the Brain: Enhancing Intra- and Inter-Brain Synchrony. Front. Hum. Neurosci. 2021, 14, 584312. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene | Chr Position | Association | Function | Refs. |
|---|---|---|---|---|
| AVPR1A | 12q14.2 | Dance performance, active listening to music, musical aptitude and creativity | Memory, learning, social behaviors | [93,96,100,101,102,103] |
| SLC6A4 | 17q11.2 | Dance performance, short-term music memory | Implicated in emotions, neuropsychiatric disorders and depression | [96,101,105,106,107,108,109] |
| UNC5C | 4q22.3 | Musical aptitude | Axon extension, cell migration in developmental nervous system | [92,111] |
| UGT8 | 4q23 | Musical aptitude | Complex lipids synthesis in myelinating oligodendrocytes, ceramides clearance in neurons | [98,114] |
| PCDH7 | 4p15.1 | Musical aptitude | Cochlea development in chicken, post-natal and adult amygdala | [115,116,117] |
| GALM | 2p22.1 | Musical creativity | Involved in serotonin release and binding potential of serotonin transporter in thalamus | [95,123,125] |
| PCDHA 1–9 | 5q31.3 | Musical aptitude | Synaptogenesis, serotoninergic projections’ maturation, learning, memory | [95,120,121] |
| SNCA | 4q22.1 | Upregulated after musical performance and listening to music | Dopamine homeostasis, synaptic plasticity, Parkinson’s disease, song learning in songbirds | [126,127,128,129,130,131,132] |
| GATA2 | 3q21.3 | Musical aptitude | Inner ear development, inferior colliculus development, determination of GABAergic neurons, expressed in dopaminergic neurons | [99,133,134,135] |
| FOXP2 | 7q31.1 | Targeted by upregulated miRNAs after listen to and music performance | Language development, song learning and singing in songbirds | [136,137,138,139] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beccacece, L.; Abondio, P.; Cilli, E.; Restani, D.; Luiselli, D. Human Genomics and the Biocultural Origin of Music. Int. J. Mol. Sci. 2021, 22, 5397. https://doi.org/10.3390/ijms22105397
Beccacece L, Abondio P, Cilli E, Restani D, Luiselli D. Human Genomics and the Biocultural Origin of Music. International Journal of Molecular Sciences. 2021; 22(10):5397. https://doi.org/10.3390/ijms22105397
Chicago/Turabian StyleBeccacece, Livia, Paolo Abondio, Elisabetta Cilli, Donatella Restani, and Donata Luiselli. 2021. "Human Genomics and the Biocultural Origin of Music" International Journal of Molecular Sciences 22, no. 10: 5397. https://doi.org/10.3390/ijms22105397
APA StyleBeccacece, L., Abondio, P., Cilli, E., Restani, D., & Luiselli, D. (2021). Human Genomics and the Biocultural Origin of Music. International Journal of Molecular Sciences, 22(10), 5397. https://doi.org/10.3390/ijms22105397