
Effect of Ecdysterone on the Hepatic Transcriptome and Lipid Metabolism in Lean and Obese Zucker Rats

 , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. Growth Performance and Organ Weights
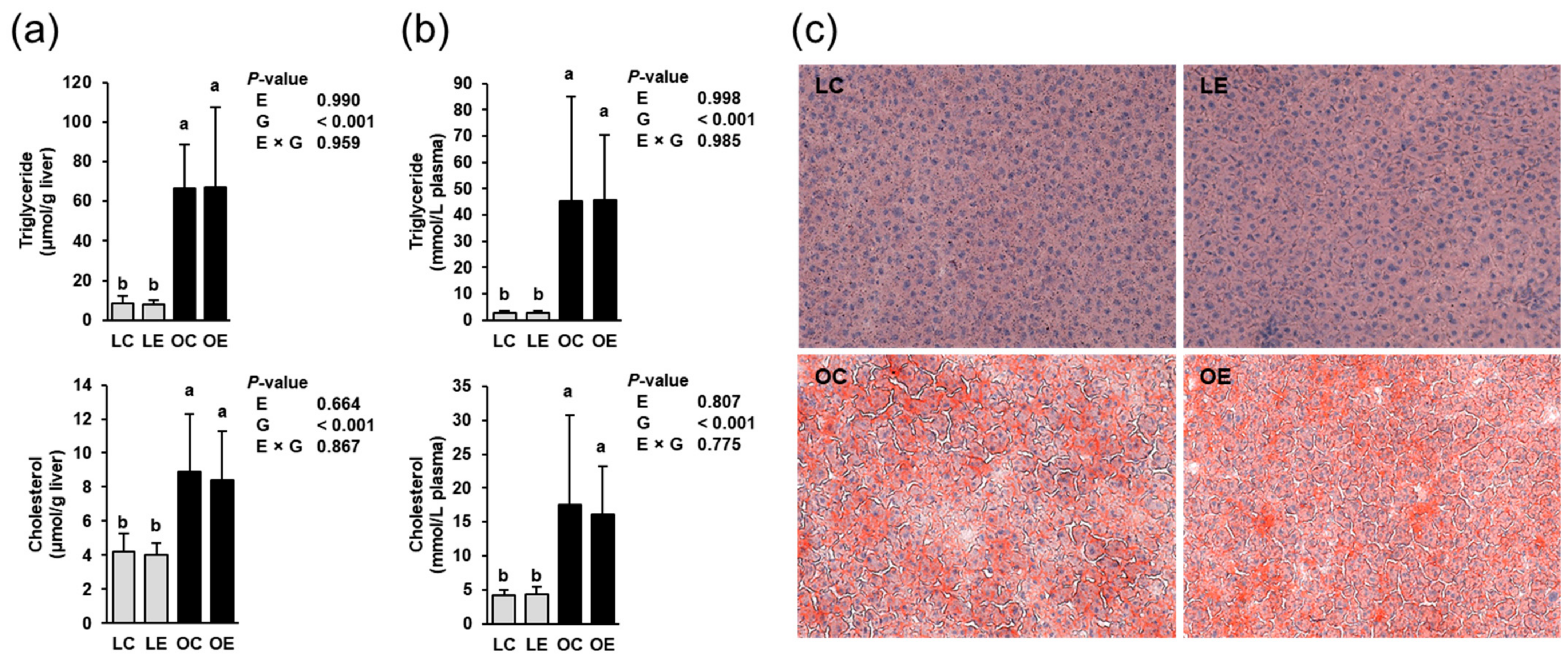
2.2. Hepatic and Plasma Lipid Concentrations
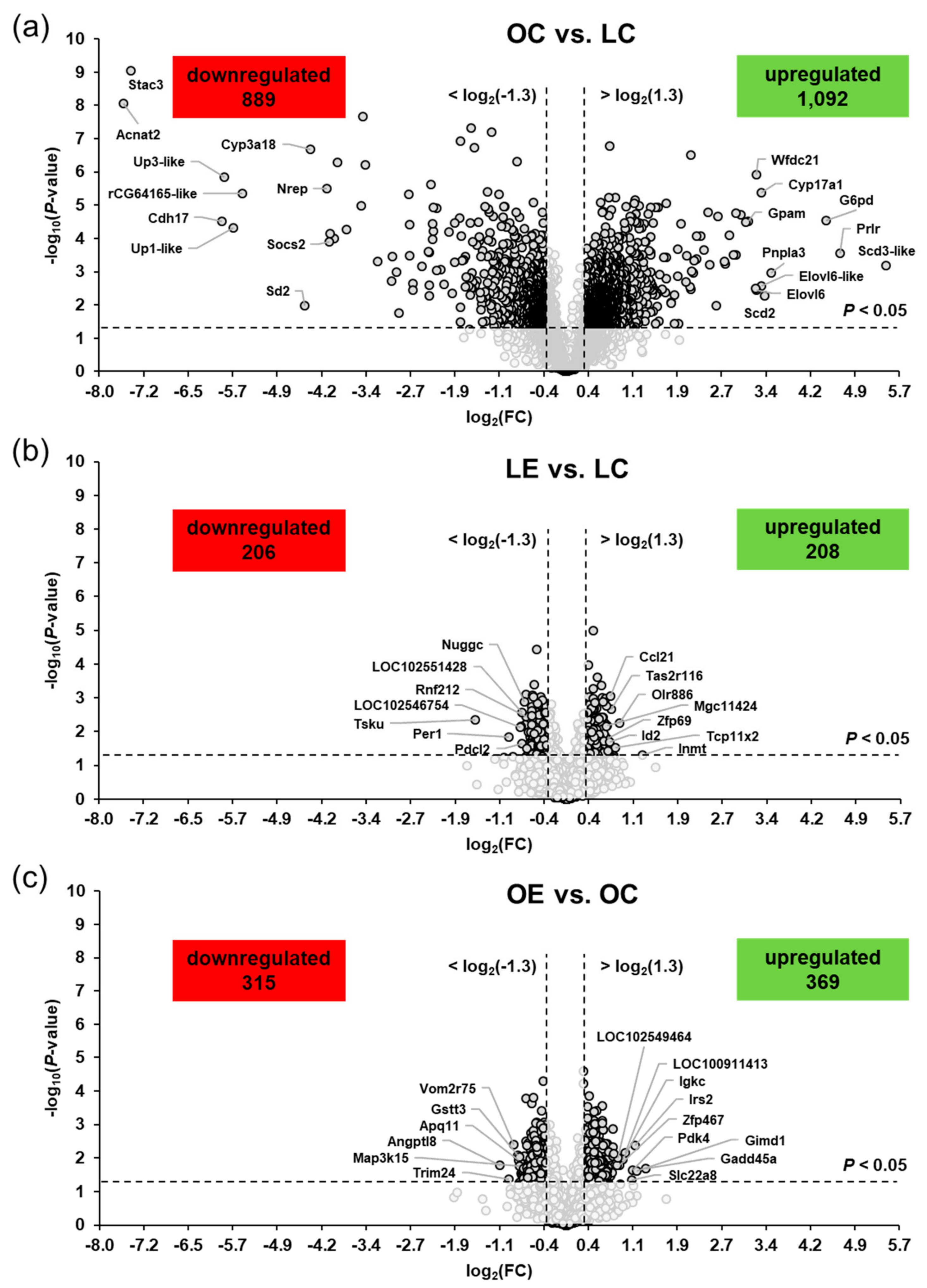
2.3. Genotype- and Ecdysterone-Regulated Transcripts in the Liver
2.4. Technical Validation of Microarray Data
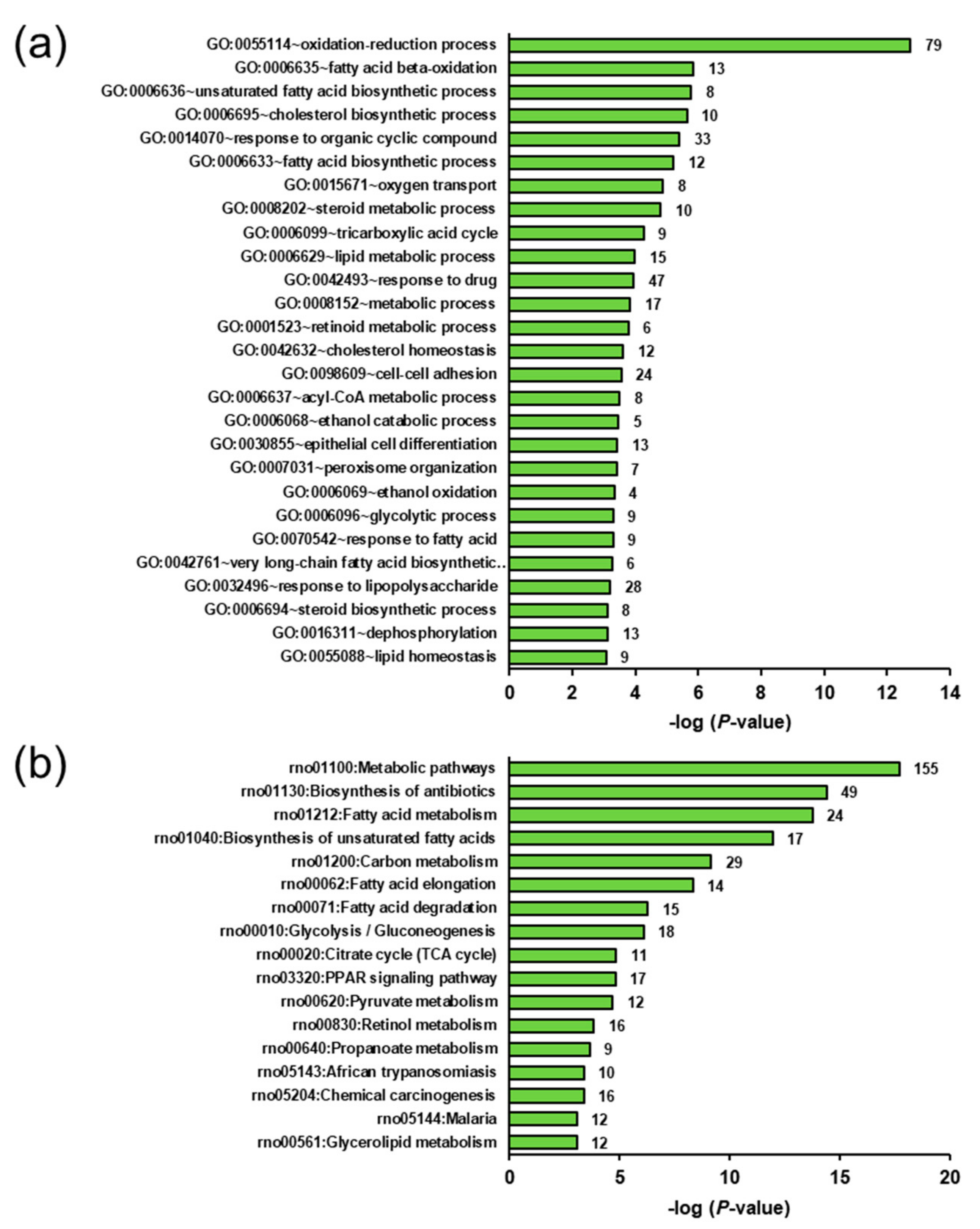
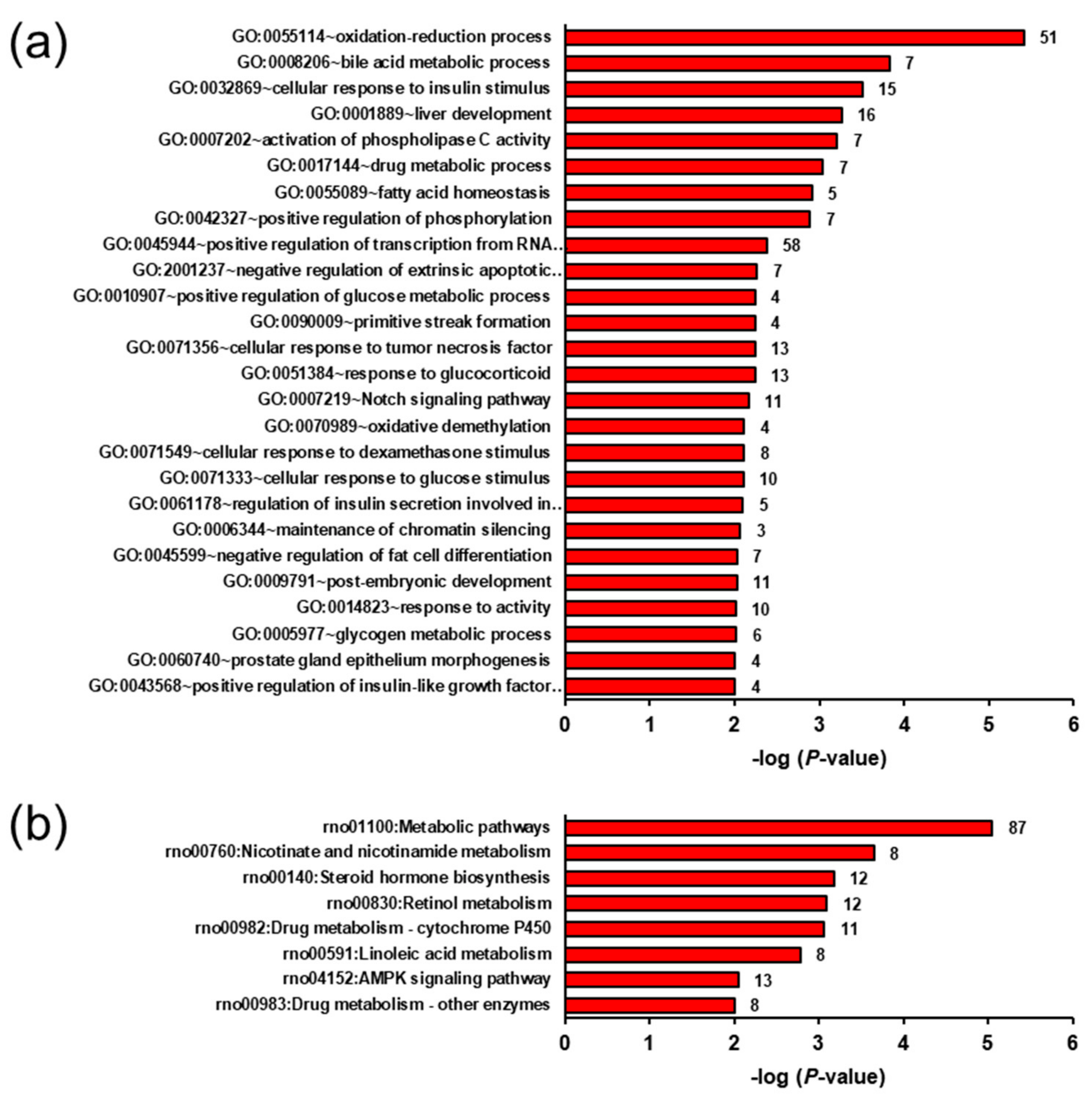
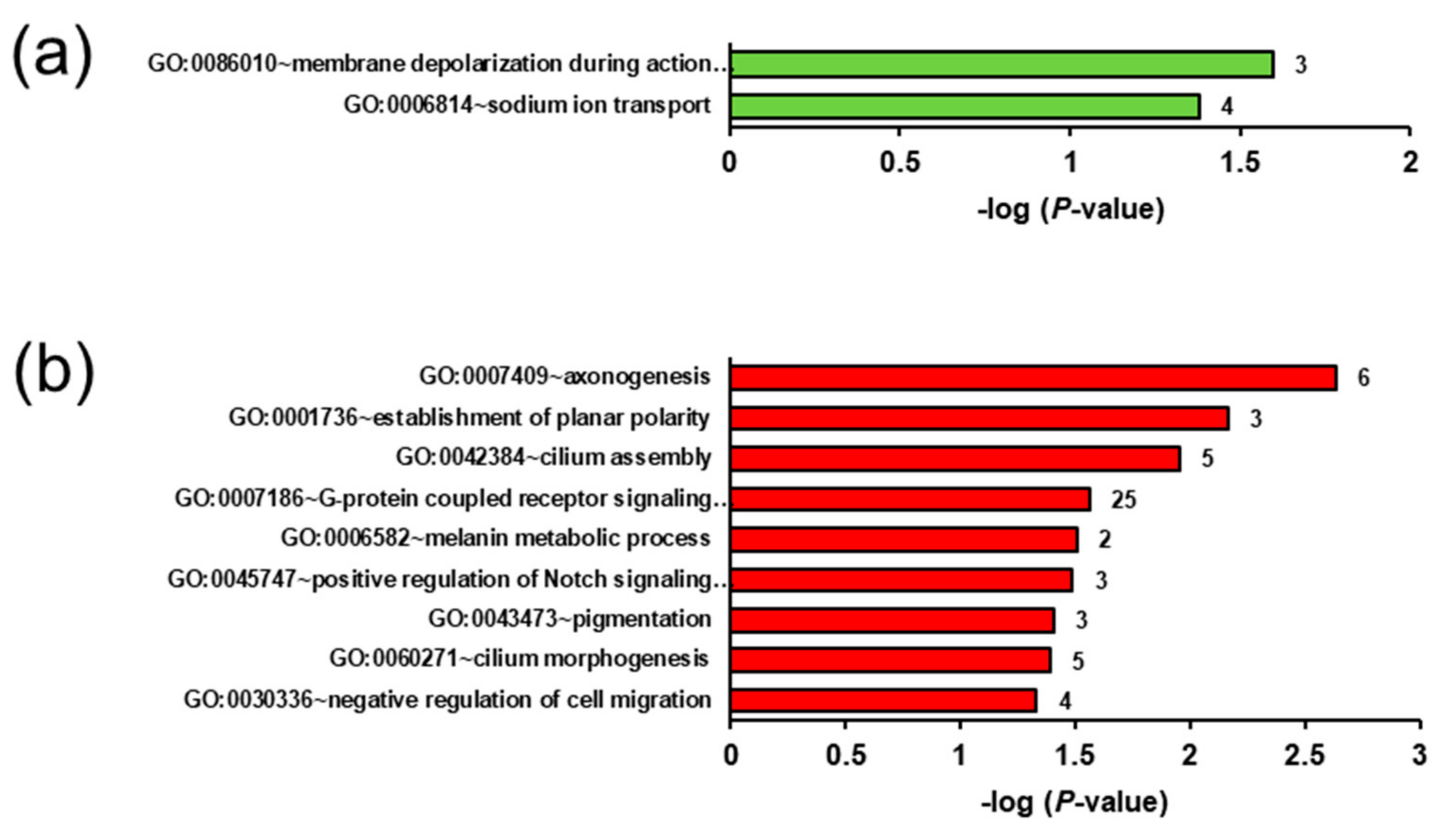
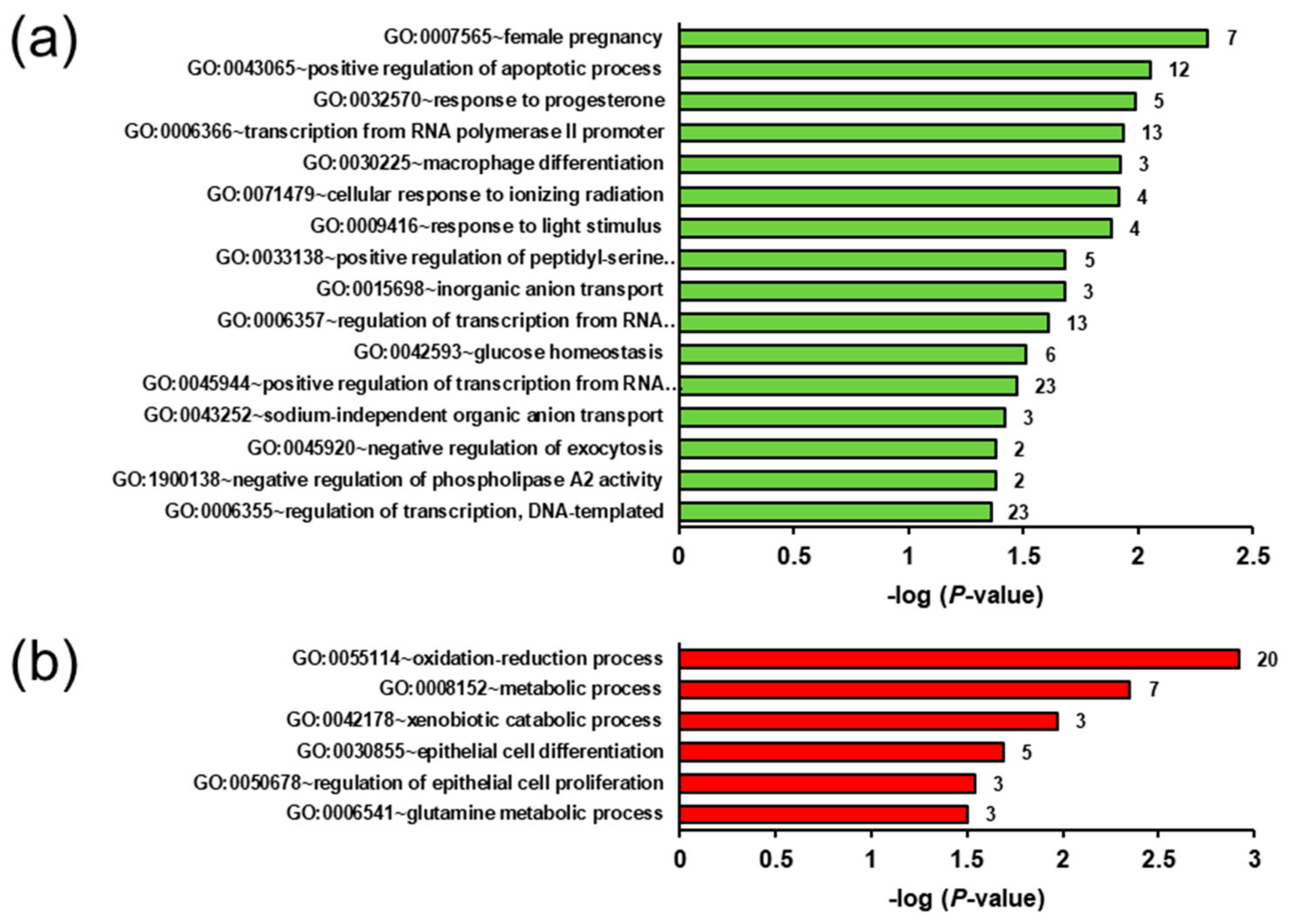
2.5. Biological Processes and Pathways Affected by the Genotype- and Ecdysterone-Regulated Transcripts in the Liver
2.6. Effect of Ecdysterone on Genes Involved in Lipid Synthetic Pathways in the Liver
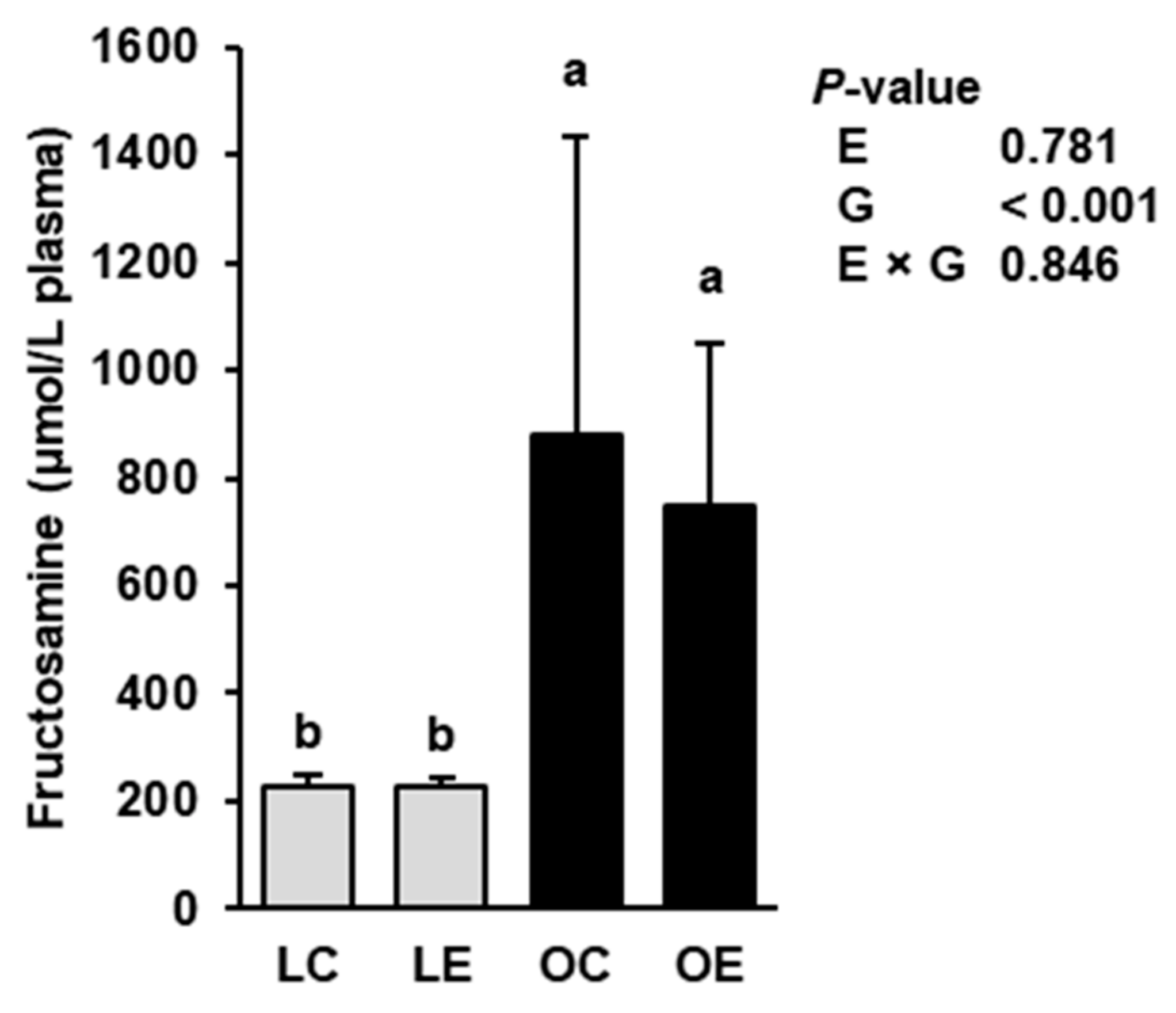
2.7. Plasma Concentration of Fructosamine
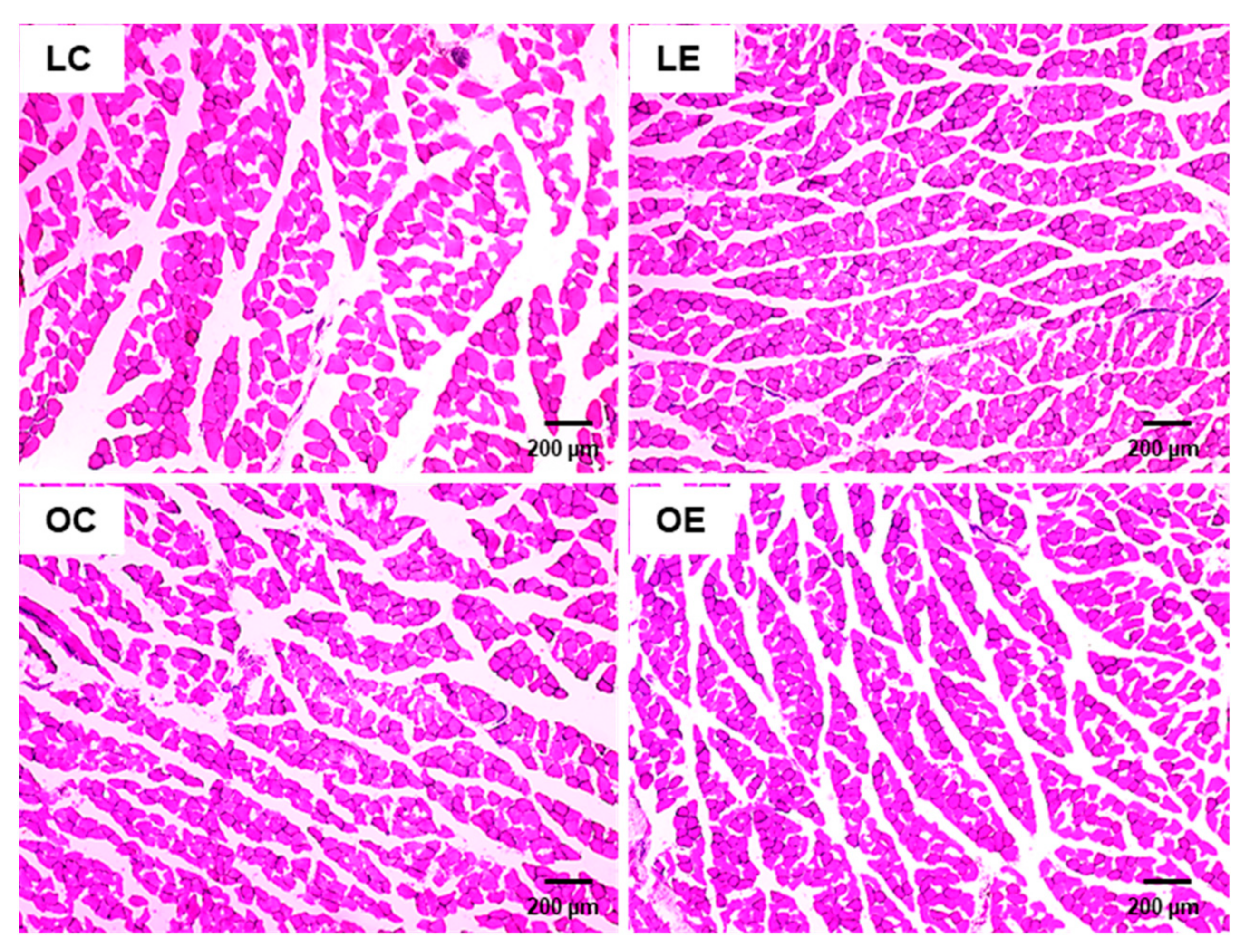
2.8. Histological Analysis of M. rectus Femoris
3. Discussion
4. Materials and Methods
4.1. Animals and Diets
4.2. Sample Collection
4.3. Determination of TG and Cholesterol Concentrations in Liver and Plasma
4.4. Determination of the Concentrations of Fatty Acids of Hepatic Total Lipids
4.5. RNA Extraction
4.6. Microarray Analysis and Bioinformatic Analysis
4.7. Validation of Microarray Data Using qPCR Analysis
4.8. Determination of Fructosamine Concentration in Plasma
4.9. Histological Analysis of Liver and M. rectus Femoris
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilbert, L.I.; Bollenbacher, W.E.; Goodman, W.; Smith, S.L.; Agui, N.; Granger, N.; Sedlak, B.J. Hormones controlling insect metamorphosis. Recent Prog. Horm. Res. 1980, 36, 401–449. [Google Scholar]
- Dinan, L. Phytoecdysteroids: Biological aspects. Phytochemistry 2001, 57, 325–339. [Google Scholar] [CrossRef]
- Delbecque, J.P.; Delachambre, J.; Hirn, M.; De Reggi, M. Abdominal production of ecdysterone and pupal-adult development in Tenebrio molitor (Insecta, Coleoptera). Gen. Comp. Endocrinol. 1978, 35, 436–444. [Google Scholar] [CrossRef]
- Truman, J.W. The Evolution of Insect Metamorphosis. Curr. Biol. 2019, 29, R1252–R1268. [Google Scholar] [CrossRef]
- Dinan, L.; Savchenko, T.; Whiting, P. On the distribution of phytoecdysteroids in plants. Cell. Mol. Life Sci. CMLS 2001, 58, 1121–1132. [Google Scholar] [CrossRef]
- Wang, J.L.; Ruan, D.C.; Cheng, Z.Y.; Yang, C.R. The dynamic variation of 20-hydroxyecdysone in Cyanotis arachnoidea. Acta Bot. Yunnanica 1996, 18, 459–464. [Google Scholar]
- Bandara, B.M.R.; Jayasinghe, L.; Karunaratne, V.; Wannigama, G.P.; Bokel, M.; Kraus, W.; Sotheeswaran, S. Ecdysterone from stem of Diploclisia glaucescens. Phytochemistry 1989, 28, 1073–1075. [Google Scholar] [CrossRef]
- Grebenok, R.J.; Ripa, P.V.; Adler, J.H. Occurrence and levels of ecdysteroids in spinach. Lipids 1991, 26, 666–668. [Google Scholar] [CrossRef]
- Schmelz, E.A.; Grebenok, R.J.; Galbraith, D.W.; Bowers, W.S. Damage-induced accumulation of phytoecdysteroids in spinach: A rapid root response involving the octadecanoic acid pathway. J. Chem. Ecol. 1998, 24, 339–360. [Google Scholar] [CrossRef]
- Schmelz, E.A.; Grebenok, R.J.; Galbraith, D.W.; Bowers, W.S. Insect-induced synthesis of phytoecdysteroids in spinach, Spinacia oleracea. J. Chem. Ecol. 1999, 25, 1739–1757. [Google Scholar] [CrossRef]
- Kizelsztein, P.; Govorko, D.; Komarnytsky, S.; Evans, A.; Wang, Z.; Cefalu, W.T.; Raskin, I. 20-Hydroxyecdysone decreases weight and hyperglycemia in a diet-induced obesity mice model. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E433–E439. [Google Scholar] [CrossRef]
- Foucault, A.S.; Even, P.; Lafont, R.; Dioh, W.; Veillet, S.; Tomé, D.; Huneau, J.F.; Hermier, D.; Quignard-Boulangé, A. Quinoa extract enriched in 20-hydroxyecdysone affects energy homeostasis and intestinal fat absorption in mice fed a high-fat diet. Physiol. Behav. 2014, 128, 226–231. [Google Scholar] [CrossRef]
- Parr, M.K.; Zhao, P.; Haupt, O.; Ngueu, S.T.; Hengevoss, J.; Fritzemeier, K.H.; Piechotta, M.; Schlörer, N.; Muhn, P.; Zheng, W.Y.; et al. Estrogen receptor beta is involved in skeletal muscle hypertrophy induced by the phytoecdysteroid ecdysterone. Mol. Nutr. Food Res. 2014, 58, 1861–1872. [Google Scholar] [CrossRef]
- Naresh Kumar, R.; Sundaram, R.; Shanthi, P.; Sachdanandam, P. Protective role of 20-OH ecdysone on lipid profile and tissue fatty acid changes in streptozotocin induced diabetic rats. Eur. J. Pharmacol. 2013, 698, 489–498. [Google Scholar] [CrossRef]
- Buniam, J.; Chukijrungroat, N.; Rattanavichit, Y.; Surapongchai, J.; Weerachayaphorn, J.; Bupha-Intr, T.; Saengsirisuwan, V. 20-Hydroxyecdysone ameliorates metabolic and cardiovascular dysfunction in high-fat-high-fructose-fed ovariectomized rats. BMC Complement. Med. Ther. 2020, 20, 140. [Google Scholar] [CrossRef]
- Foucault, A.S.; Mathé, V.; Lafont, R.; Even, P.; Dioh, W.; Veillet, S.; Tomé, D.; Huneau, J.F.; Hermier, D.; Quignard-Boulangé, A. Quinoa extract enriched in 20-hydroxyecdysone protects mice from diet-induced obesity and modulates adipokines expression. Obesity 2011, 20, 270–277. [Google Scholar] [CrossRef]
- Gessner, D.K.; Schwarz, A.; Meyer, S.; Wen, G.; Most, E.; Zorn, H.; Ringseis, R.; Eder, K. Insect Meal as Alternative Protein Source Exerts Pronounced Lipid-Lowering Effects in Hyperlipidemic Obese Zucker Rats. J. Nutr. 2019, 149, 566–577. [Google Scholar] [CrossRef]
- Meyer, S.; Gessner, D.K.; Wen, G.; Most, E.; Liebisch, G.; Zorn, H.; Ringseis, R.; Eder, K. The Antisteatotic and Hypolipidemic Effect of Insect Meal in Obese Zucker Rats is Accompanied by Profound Changes in Hepatic Phospholipid and 1-Carbon Metabolism. Mol. Nutr. Food Res. 2019, 63, e1801305. [Google Scholar] [CrossRef]
- Stopka, P.; Stancl, J.; Slama, K. Effect of insect hormone, 20-hydroxyecdysone on growth and reproduction in mice. Acta Soc. Zoo. Bohem. 1999, 63, 367–378. [Google Scholar]
- Todorov, I.; Mitrokin, Y.; Efremova, O.; Sidorenko, L. The effect of ecdysterone on the biosynthesis of proteins and nucleic acids in mice. Pharm. Chem. J. 2000, 34, 455–458. [Google Scholar] [CrossRef]
- Syrov, V.; Kurmukov, A. On the anabolic activity of the phytoecdysone-ecdysterone isolated from Rhaponticum carthamoides (Willd.) Iljin. Farmakol. Toksikol. 1976, 39, 690–693. [Google Scholar] [PubMed]
- Kratky, F.; Opletal, L.; Hejhalek, J.; Kucharova, S. Effect of 20-hydroxyecdysone on the protein synthesis of pigs. Zivocisna Vyrob. 1997, 42, 445–451. [Google Scholar]
- Koudela, K.; Tenora, J.; Bajer, J.; Mathova, A.; Slama, K. Stimulation of growth and development in Japanese quails after oral administration of ecdysteroid-containing diet. Eur. J. Entomol. 1995, 92, 349–354. [Google Scholar]
- Gorelick-Feldman, J.; Maclean, D.; Ilic, N.; Poulev, A.; Lila, M.A.; Cheng, D.; Raskin, I. Phytoecdysteroids increase protein synthesis in skeletal muscle cells. J. Agric. Food Chem. 2008, 56, 3532–3537. [Google Scholar] [CrossRef]
- Guttridge, D.C. Signaling pathways weigh in on decisions to make or break skeletal muscle. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 443–450. [Google Scholar] [CrossRef]
- Trostler, N.; Amin, R.; Shafrir, E. Increased protease activity in muscles of obese-(ob/ob) mice. Int. J. Obes. 1981, 6, 557–566. [Google Scholar]
- Durschlag, R.; Layman, D. Skeletal muscle growth in lean and obese Zucker rats. Growth 1983, 47, 282. [Google Scholar]
- Couturier, A.; Ringseis, R.; Mooren, F.C.; Krüger, K.; Most, E.; Eder, K. Carnitine supplementation to obese Zucker rats prevents obesity-induced type II to type I muscle fiber transition and favors an oxidative phenotype of skeletal muscle. Nutr. Metab. 2013, 10, 48. [Google Scholar] [CrossRef]
- Acevedo, L.M.; Raya, A.I.; Martínez-Moreno, J.M.; Aguilera-Tejero, E.; Rivero, J.L. Mangiferin protects against adverse skeletal muscle changes and enhances muscle oxidative capacity in obese rats. PLoS ONE 2017, 12, e0173028. [Google Scholar] [CrossRef]
- Meyer, S.; Schäfer, L.; Röhrig, J.; Maheshwari, G.; Most, E.; Zorn, H.; Ringseis, R.; Eder, K.; Gessner, D.K. Supplementation of Sulfur-Containing Amino Acids or Essential Amino Acids Does Not Reverse the Hepatic Lipid-Lowering Effect of a Protein-Rich Insect Meal in Obese Zucker Rats. Nutrients 2020, 12, 987. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Laboratory Animals, 4th ed.; National Academy Press: Washington, DC, USA, 1995. [Google Scholar]
- Sreejit, C.M.; Bose, C.; Banerji, A.; Matthew, P.T. Isolation, quantification and chemical characterization of ecdysterone from medicinal plants of Kerala, Western Ghats. J. Pharmacogn. Phytochem. 2019, 8, 254–257. [Google Scholar]
- VDLUFA (Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten). Die chemische Untersuchung von Futtermitteln. VDLUFA-Methodenbuch. Band III, Ergänzungslieferungen von 1983, 1988, 1992, 1997, 2004, 2006, 2007; VDLUFA-Verlag: Darmstadt, Germany, 2007. [Google Scholar]
- Hara, A.; Radin, N.S. Lipid extraction of tissues with a low toxicity solvent. Anal. Biochem 1978, 90, 420–426. [Google Scholar] [CrossRef]
- Eder, K.; Kirchgessner, M. Dietary fat influences the effect of zinc deficiency on liver lipids and fatty acids in rats force-fed equal quantities of diet. J. Nutr. 1994, 124, 1917–1926. [Google Scholar] [CrossRef]
- Schlegel, G.; Ringseis, R.; Shibani, M.; Most, E.; Schuster, M.; Schwarz, F.J.; Eder, K. Influence of a rumen-protected conjugated linoleic acid mixture on carcass traits and meat quality in young Simmental heifers. J. Anim. Sci. 2012, 90, 1532–1540. [Google Scholar] [CrossRef]
- Gessner, D.K.; Winkler, A.; Koch, C.; Dusel, G.; Liebisch, G.; Ringseis, R.; Eder, K. Analysis of hepatic transcript profile and plasma lipid profile in early lactating dairy cows fed grape seed and grape marc meal extract. BMC Genom. 2017, 18, 253. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef]
- Ringseis, R.; Zeitz, J.O.; Weber, A.; Koch, C.; Eder, K. Hepatic transcript profiling in early-lactation dairy cows fed rumen-protected niacin during the transition from late pregnancy to lactation. J. Dairy Sci. 2019, 102, 365–376. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef]
- Chiappisi, E.; Ringseis, R.; Eder, K.; Gessner, D.K. Effect of endoplasmic reticulum stress on metabolic and stress signaling and kidney-specific functions in Madin-Darby bovine kidney cells. J. Dairy Sci. 2017, 100, 6689–6706. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Lean | Obese | Two-Way ANOVA p-Value | ||||
|---|---|---|---|---|---|---|---|
| Ecdysterone (g/kg Diet) | 0 | 0.5 | 0 | 0.5 | E | G | E × G |
| Body weight, g | |||||||
| Initial | 441 ± 29 b | 446 ± 49 b | 570 ± 74 a | 561 ± 29 a | 0.919 | <0.001 | 0.706 |
| Final | 465 ± 34 b | 476 ± 51 b | 611 ± 79 a | 609 ± 32 a | 0.821 | <0.001 | 0.746 |
| Daily body weight gain, g | 0.86 ± 0.35 b | 1.06 ± 0.41 b | 1.46 ± 0.36 a | 1.69 ± 0.93 a | 0.294 | 0.05 | 0.928 |
| Daily feed intake, g | 20.3 ± 1.2 b | 20.8 ± 1.3 b | 23.4 ± 1.3 a | 25.0 ± 0.9 a | 0.103 | <0.001 | 0.361 |
| Feed:gain ratio, g/g | 27.9 ± 9.7 a | 26.2 ± 13.3 a | 16.6 ± 3.7 b | 16.0 ± 3.4 b | 0.796 | 0.027 | 0.905 |
| Organ weights, g | |||||||
| Heart | 1.40 ± 0.08 a,b | 1.39 ± 0.08 b | 1.52 ± 0.13 a | 1.51 ± 0.07 a,b | 0.657 | 0.001 | 0.948 |
| Kidney right | 1.80 ± 0.21 b | 1.74 ± 0.30 b | 1.97 ± 0.17 a | 2.22 ± 0.19 a | 0.242 | <0.001 | 0.059 |
| Kidney left | 1.79 ± 0.20 b | 1.71 ± 0.31 b | 1.97 ± 0.14 a | 2.26 ± 0.24 a | 0.197 | <0.001 | 0.028 |
| Liver | 17.9 ± 1.9 b | 17.7 ± 2.1 b | 33.0 ± 3.6 a | 32.5 ± 4.5 a | 0.802 | <0.001 | 0.898 |
| M. soleus | 0.17 ± 0.02 a | 0.17 ± 0.01 a | 0.12 ± 0.02 b | 0.13 ± 0.01 b | 0.457 | <0.001 | 0.327 |
| M. vastus medialis | 0.50 ± 0.09 a | 0.54 ± 0.13 a | 0.32 ± 0.11 b | 0.30 ± 0.09 b | 0.717 | <0.001 | 0.424 |
| M. gastrocnemius | 2.18 ± 0.19 a | 2.28 ± 0.15 a | 1.58 ± 0.14 b | 1.56 ± 0.05 b | 0.362 | <0.001 | 0.237 |
| M. rectus femoris | 1.48 ± 0.56 a | 1.39 ± 0.29 a | 0.85 ± 0.11 b | 0.88 ± 0.07 b | 0.802 | <0.001 | 0.609 |
| M. vastus intermedius | 1.36 ± 0.12 a | 1.33 ± 0.17 a | 0.87 ± 0.07 b | 0.84 ± 0.09 b | 0.468 | <0.001 | 0.979 |
| Genotype | Lean | Obese | Two-Way ANOVA p-Value | ||||
|---|---|---|---|---|---|---|---|
| Ecdysterone (g/kg Diet) | 0 | 0.5 | 0 | 0.5 | E | G | E × G |
| µmol/g liver | |||||||
| 14:0 | 0.66 ± 0.23 b | 0.52 ± 0.13 b | 3.30 ± 1.11 a | 4.08 ± 1.64 a | 0.666 | <0.001 | 0.414 |
| 14:1 n-5 | 0.12 ± 0.04 b | 0.07 ± 0.03 b | 0.46 ± 0.13 a | 0.38 ± 0.17 a | 0.124 | <0.001 | 0.616 |
| 16:0 | 18.0 ± 3.7 b | 14.7 ± 1.9 b | 62.2 ± 19.4 a | 70.8 ± 28.3 a | 0.662 | <0.001 | 0.340 |
| 16:1 n-7 | 2.8 ± 1.4 b | 2.2 ± 0.7 b | 15.7 ± 5.5 a | 17.2 ± 8.3 a | 0.819 | <0.001 | 0.552 |
| 17:0 | 0.23 ± 0.07 | 0.16 ± 0.04 | 0.52 ± 0.68 | 0.29 ± 0.11 | 0.206 | 0.088 | 0.482 |
| 18:0 | 11.6 ± 2.2 b | 9.9 ± 2.1 b | 16.1 ± 2.2 a | 17.9 ± 3.0 a | 0.604 | <0.001 | 0.026 |
| 18:1 n-9 | 8.5 ± 2.1 b | 6.4 ± 1.4 b | 48.8 ± 18.6 a | 56.0 ± 25.3 a | 0.656 | <0.001 | 0.415 |
| 18:2 n-6 | 12.5 ± 3.2 | 9.1 ± 2.3 | 10.7 ± 2.7 | 11.5 ± 3.5 | 0.752 | 0.357 | 0.030 |
| 18:3 n-3 | 0.37 ± 0.13 b | 0.25 ± 0.08 b | 0.51 ± 0.38 a | 0.52 ± 0.30 a | 0.531 | 0.028 | 0.470 |
| 18:3 n-6 | 0.15 ± 0.02 b | 0.11 ± 0.05 b | 0.28 ± 0.26 a | 0.30 ± 0.12 a | 0.908 | 0.003 | 0.528 |
| 20:3 n-6 | 0.49 ± 0.09 b | 0.41 ± 0.14 b | 0.68 ± 0.24 a | 0.83 ± 0.28 a | 0.659 | <0.001 | 0.113 |
| 20:4 n-6 | 13.4 ± 2.4 a | 10.6 ± 1.8 b | 9.7 ± 1.9 b | 10.5 ± 2.7 b | 0.221 | 0.027 | 0.034 |
| 22:5 n-3 | 0.40 ± 0.08 a | 0.28 ± 0.06 b | 0.26 ± 0.09 b | 0.25 ± 0.05 b | 0.017 | 0.057 | 0.120 |
| 22:6 n-3 | 3.31 ± 0.68 a | 2.60 ± 0.54 a | 1.82 ± 0.46 b | 1.94 ± 0.52 b | 0.140 | <0.001 | 0.042 |
| ∑ total fatty acids | 72.4 ± 12.3 b | 57.3 ± 8.8 b | 170.7 ± 50.2 a | 194.5 ± 74.4 a | 0.790 | <0.001 | 0.236 |
| OC vs. LC | OE vs. OC | ||||
|---|---|---|---|---|---|
| Gene Symbol | Lipid Synthetic Pathway | FC | p-Value | FC | p-Value |
| Scd3-like | Fatty acids | 44.50 | 0.0007 | not regulated | n.s. |
| G6pd | Fatty acids | 21.80 | <0.0001 | not regulated | n.s. |
| Scd2 | Fatty acids | 10.50 | 0.0053 | not regulated | n.s. |
| Elovl6-like | Fatty acids | 10.14 | 0.0026 | not regulated | n.s. |
| Elovl6 | Fatty acids | 9.49 | 0.0032 | not regulated | n.s. |
| Gpam | Triglycerides | 8.64 | <0.0001 | not regulated | n.s. |
| Cd36 | Triglycerides | 8.42 | <0.000 | not regulated | n.s. |
| Fabp4 | Triglycerides | 6.96 | 0.0001 | not regulated | n.s. |
| Me1 | Fatty acids | 6.61 | 0.0006 | not regulated | n.s. |
| Fabp5 | Triglycerides | 4.72 | 0.0001 | not regulated | n.s. |
| Fabp2 | Triglycerides | 3.92 | <0.0001 | not regulated | n.s. |
| Fasn | Fatty acids | 3.64 | 0.0051 | −1.39 | 0.0381 |
| Dgat2 | Triglycerides | 3.26 | 0.0043 | not regulated | n.s. |
| Acss2 | Fatty acids | 2.62 | 0.0095 | not regulated | n.s. |
| Acsl5 | Fatty acids | 2.57 | 0.0001 | not regulated | n.s. |
| Agpat3 | Phospholipids | 2.54 | <0.0001 | not regulated | n.s. |
| Degs1 | Fatty acids | 2.38 | <0.0001 | not regulated | n.s. |
| Fdps | Cholesterol | 2.37 | 0.0350 | not regulated | n.s. |
| Srebf1 | Fatty acids | 2.36 | 0.0003 | not regulated | n.s. |
| Pctp | Phospholipids | 2.31 | 0.0008 | not regulated | n.s. |
| Lss | Cholesterol | 2.18 | 0.0037 | not regulated | n.s. |
| Acaca | Fatty acids | 2.08 | 0.0264 | not regulated | n.s. |
| Fads2 | Fatty acids | 2.07 | <0.0001 | not regulated | n.s. |
| Dhcr7 | Cholesterol | 2.05 | 0.0447 | not regulated | n.s. |
| Acly | Fatty acids | 1.92 | 0.0042 | not regulated | n.s. |
| Acat2 | Fatty acids | 1.87 | 0.0026 | not regulated | n.s. |
| Ldlr | Cholesterol | 1.63 | 0.0268 | not regulated | n.s. |
| Elovl2 | Fatty acids | 1.62 | 0.0011 | not regulated | n.s. |
| Pmvk | Cholesterol | 1.62 | 0.0256 | not regulated | n.s. |
| Elovl5 | Fatty acids | 1.62 | 0.0013 | not regulated | n.s. |
| Scd1 | Fatty acids | 1.61 | 0.0055 | not regulated | n.s. |
| Aacs | Cholesterol | 1.56 | 0.0143 | not regulated | n.s. |
| Components (g/kg) | Basal Diet |
|---|---|
| Cornstarch | 555 |
| Casein | 200 |
| Sucrose | 100 |
| Soybean oil | 50 |
| Cellulose | 50 |
| Mineral mix 1 | 35 |
| Vitamin mix 2 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marschall, M.J.M.; Ringseis, R.; Gessner, D.K.; Grundmann, S.M.; Most, E.; Wen, G.; Maheshwari, G.; Zorn, H.; Eder, K. Effect of Ecdysterone on the Hepatic Transcriptome and Lipid Metabolism in Lean and Obese Zucker Rats. Int. J. Mol. Sci. 2021, 22, 5241. https://doi.org/10.3390/ijms22105241
Marschall MJM, Ringseis R, Gessner DK, Grundmann SM, Most E, Wen G, Maheshwari G, Zorn H, Eder K. Effect of Ecdysterone on the Hepatic Transcriptome and Lipid Metabolism in Lean and Obese Zucker Rats. International Journal of Molecular Sciences. 2021; 22(10):5241. https://doi.org/10.3390/ijms22105241
Chicago/Turabian StyleMarschall, Magdalena J. M., Robert Ringseis, Denise K. Gessner, Sarah M. Grundmann, Erika Most, Gaiping Wen, Garima Maheshwari, Holger Zorn, and Klaus Eder. 2021. "Effect of Ecdysterone on the Hepatic Transcriptome and Lipid Metabolism in Lean and Obese Zucker Rats" International Journal of Molecular Sciences 22, no. 10: 5241. https://doi.org/10.3390/ijms22105241
APA StyleMarschall, M. J. M., Ringseis, R., Gessner, D. K., Grundmann, S. M., Most, E., Wen, G., Maheshwari, G., Zorn, H., & Eder, K. (2021). Effect of Ecdysterone on the Hepatic Transcriptome and Lipid Metabolism in Lean and Obese Zucker Rats. International Journal of Molecular Sciences, 22(10), 5241. https://doi.org/10.3390/ijms22105241