
Molecular Basis of Complement C1q Collagen-Like Region Interaction with the Immunoglobulin-Like Receptor LAIR-1

 , ,
, ,
Abstract
1. Introduction
2. Results
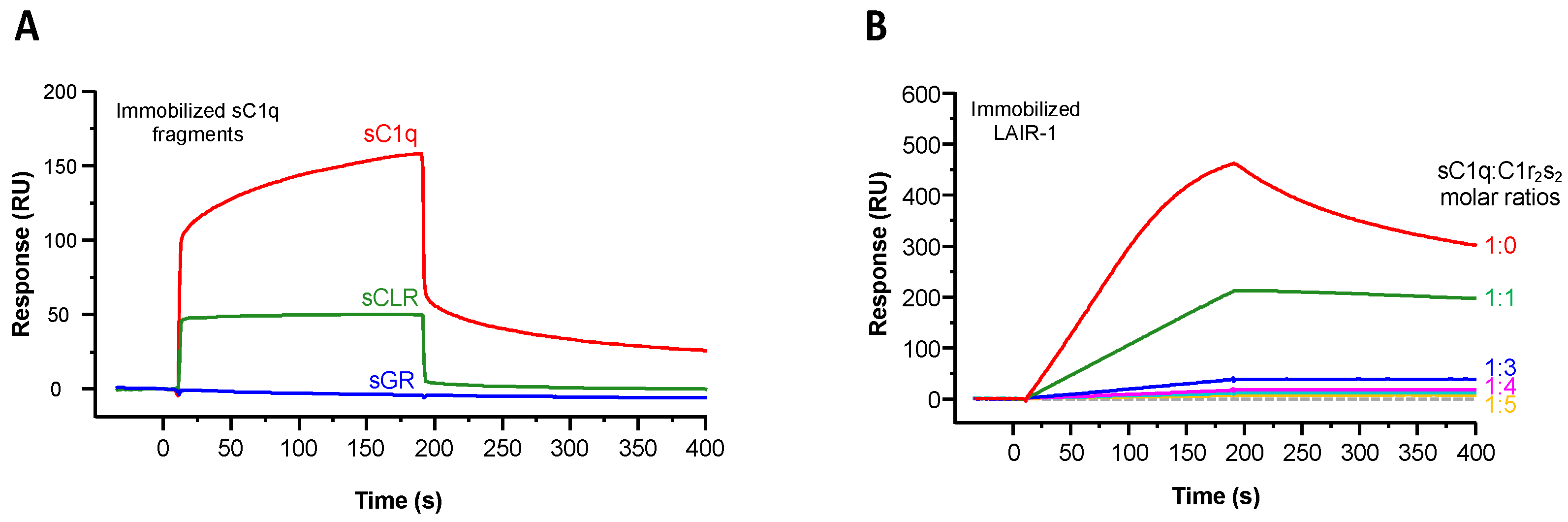
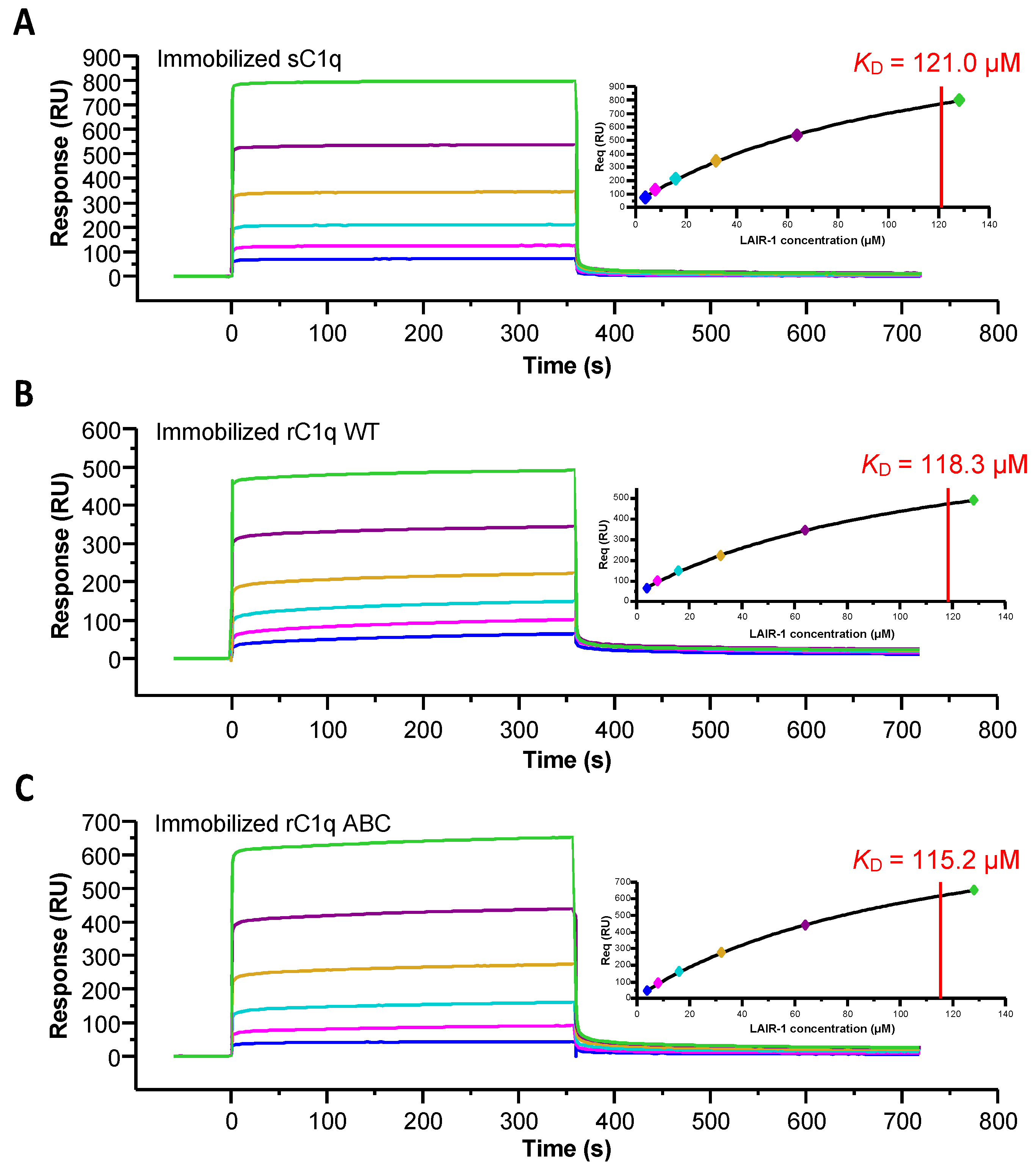
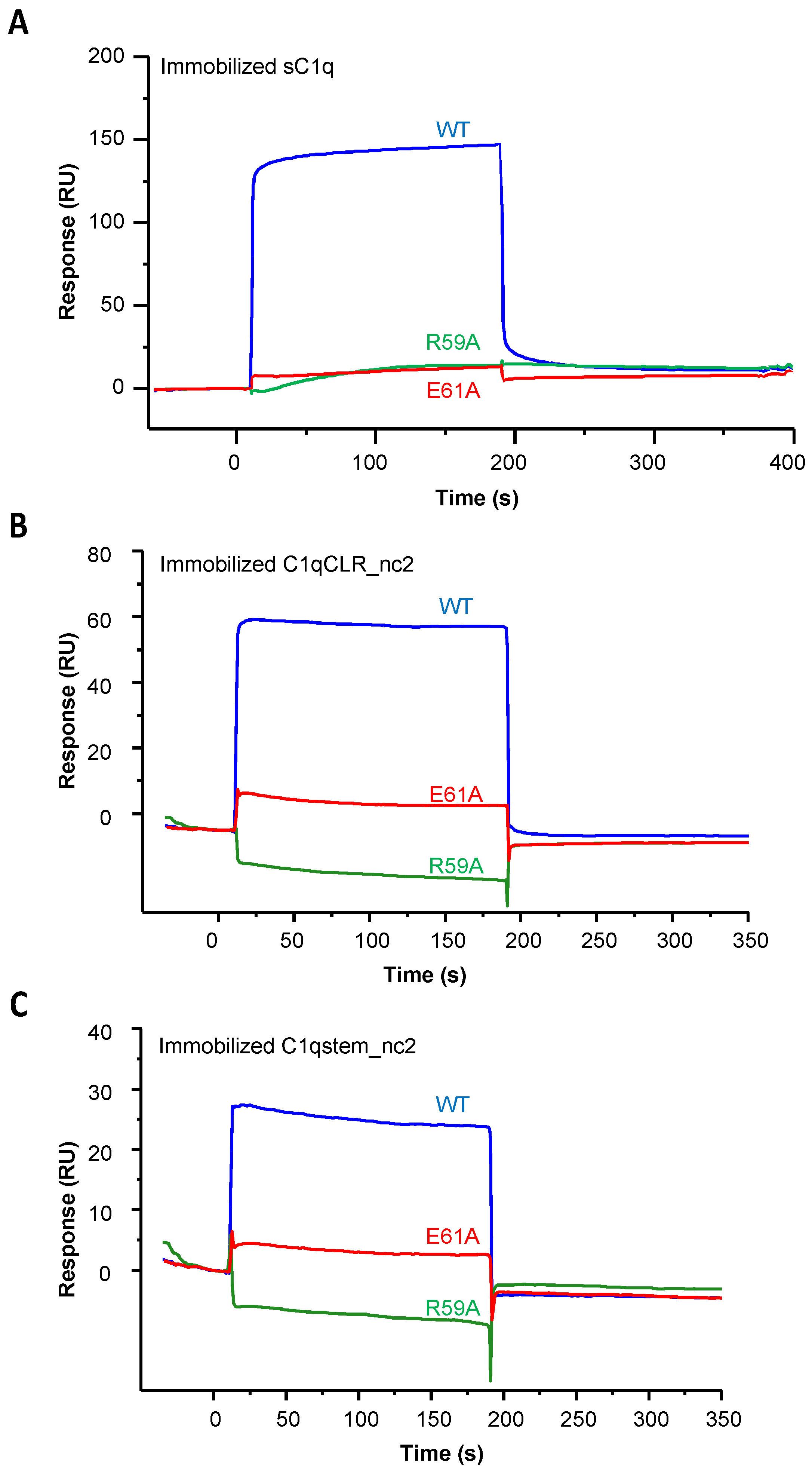
2.1. Recombinant LAIR-1 Ig-Like Domain Interacts with Serum-Derived C1q
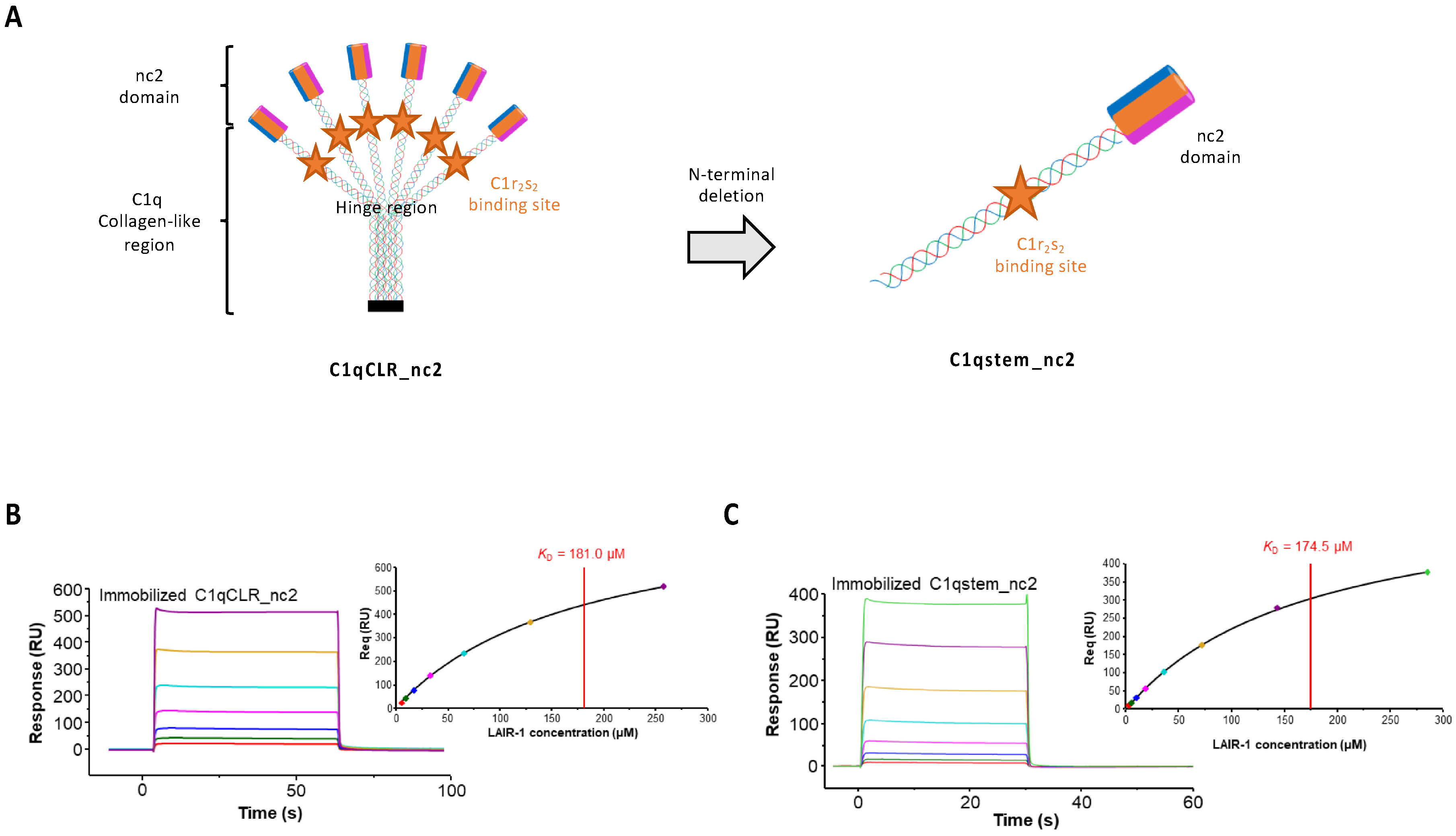
2.2. LAIR-1 Interacts with C1q Collagen Stems Through a Site in Close Proximity but Different from the C1r2s2 Tetramer Binding Site
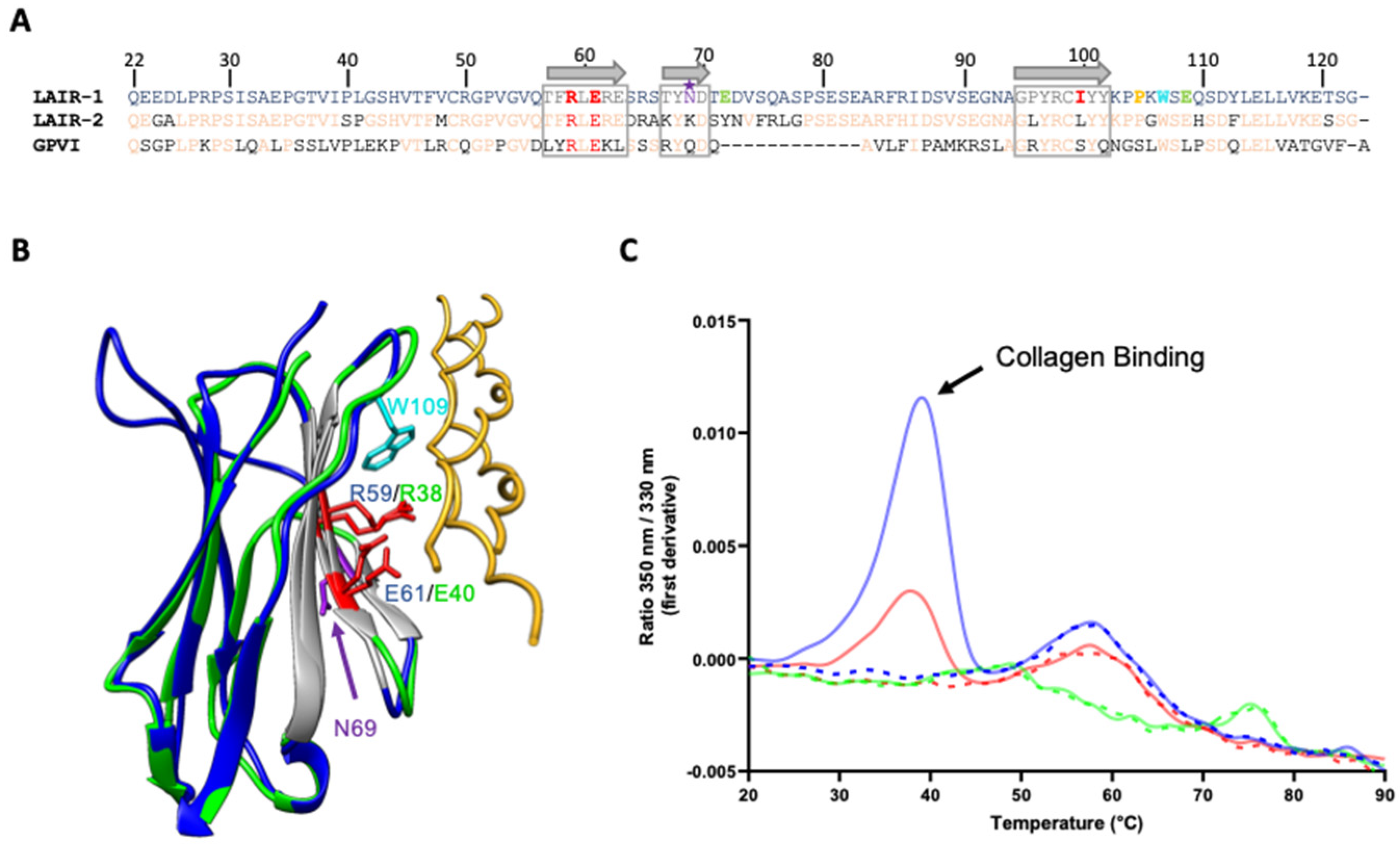
2.3. A Central Binding Groove in LAIR-1 Common for C1q and Generic Collagen Recognition
2.4. Building of a C1q/LAIR-1 Interaction Model
2.5. Exploring the Distal LAIR-1 Residues Involved in C1q Binding
3. Discussion
4. Materials and Methods
4.1. Proteins, Cells and Reagents
4.2. Construction of the C1qCLR_nc2 Protein
4.3. Construction and Purification of the C1qstem_nc2 Protein
4.4. LAIR-1 Ig-Like Domain Expression and Purification
4.5. SDS/PAGE, Circular Dichroism and LC/ESI Mass Spectrometry
4.5.1. SDS/PAGE
4.5.2. Circular Dichroism
4.5.3. LC/ESI Mass Spectrometry
4.6. Surface Plasmon Resonance Analyses and Evaluation
4.7. LAIR-1 Interaction with sC1q, sCLR and sGR and Competition Assay with C1r2s2
4.8. Interaction of LAIR-1 Variants with Immobilized Serum-Derived C1q
4.9. Kinetics of LAIR-1 Interaction with sC1q, rC1q Variants and CLR_nc2 Variants
4.10. NanoDSF Experiments
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Nayak, A.; Pednekar, L.; Reid, K.B.; Kishore, U. Complement and Non-Complement Activating Functions of C1q: A Prototypical Innate Immune Molecule. Innate Immun. 2012, 18, 350–363. [Google Scholar] [CrossRef]
- Walport, M.J.; Davies, K.A.; Botto, M. C1q and Systemic Lupus Erythematosus. Immunobiology 1998, 199, 265–285. [Google Scholar] [CrossRef]
- Kishore, U.; Gaboriaud, C.; Waters, P.; Shrive, A.K.; Greenhough, T.J.; Reid, K.B.M.; Sim, R.B.; Arlaud, G.J. C1q and Tumor Necrosis Factor Superfamily: Modularity and Versatility. Trends Immunol. 2004, 25, 551–561. [Google Scholar] [CrossRef]
- Shelton, E.; Yonemasu, K.; Stroud, R.M. Ultrastructure of the Human Complement Component, Clq. Proc. Natl. Acad. Sci. USA 1972, 69, 65–68. [Google Scholar] [CrossRef]
- Thielens, N.M.; Tedesco, F.; Bohlson, S.S.; Gaboriaud, C.; Tenner, A.J. C1q: A Fresh Look upon an Old Molecule. Mol. Immunol. 2017, 89, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Gaboriaud, C.; Frachet, P.; Thielens, N.M.; Arlaud, G.J. The Human C1q Globular Domain: Structure and Recognition of Non-Immune Self Ligands. Front. Immunol. 2011, 2, 92. [Google Scholar] [CrossRef] [PubMed]
- Ugurlar, D.; Howes, S.C.; de Kreuk, B.-J.; Koning, R.I.; de Jong, R.N.; Beurskens, F.J.; Schuurman, J.; Koster, A.J.; Sharp, T.H.; Parren, P.W.H.I.; et al. Structures of C1-IgG1 Provide Insights into How Danger Pattern Recognition Activates Complement. Science 2018, 359, 794–797. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.C.; Schumaker, V.N. Measurement of the Association Constants of the Complexes Formed between Intact C1q or Pepsin-Treated C1q Stalks and the Unactivated or Activated C1r2C1s2 Tetramers. Mol. Immunol. 1983, 20, 53–66. [Google Scholar] [CrossRef]
- Eggleton, P.; Tenner, A.J.; Reid, K.B.M. C1q Receptors. Clin. Exp. Immunol. 2000, 120, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Meyaard, L.; Adema, G.J.; Chang, C.; Woollatt, E.; Sutherland, G.R.; Lanier, L.L.; Phillips, J.H. LAIR-1, a Novel Inhibitory Receptor Expressed on Human Mononuclear Leukocytes. Immunity 1997, 7, 283–290. [Google Scholar] [CrossRef]
- Meyaard, L. The Inhibitory Collagen Receptor LAIR-1 (CD305). J. Leukoc. Biol. 2008, 83, 799–803. [Google Scholar] [CrossRef]
- Lebbink, R.J.; de Ruiter, T.; Adelmeijer, J.; Brenkman, A.B.; van Helvoort, J.M.; Koch, M.; Farndale, R.W.; Lisman, T.; Sonnenberg, A.; Lenting, P.J.; et al. Collagens Are Functional, High Affinity Ligands for the Inhibitory Immune Receptor LAIR-1. J. Exp. Med. 2006, 203, 1419–1425. [Google Scholar] [CrossRef]
- Lebbink, R.J.; Raynal, N.; de Ruiter, T.; Bihan, D.G.; Farndale, R.W.; Meyaard, L. Identification of Multiple Potent Binding Sites for Human Leukocyte Associated Ig-like Receptor LAIR on Collagens II and III. Matrix Biol. 2009, 28, 202–210. [Google Scholar] [CrossRef]
- Son, M.; Santiago-Schwarz, F.; Al-Abed, Y.; Diamond, B. C1q Limits Dendritic Cell Differentiation and Activation by Engaging LAIR-1. Proc. Natl. Acad. Sci. USA 2012, 109, E3160–E3167. [Google Scholar] [CrossRef]
- Olde Nordkamp, M.J.M.; van Eijk, M.; Urbanus, R.T.; Bont, L.; Haagsman, H.P.; Meyaard, L. Leukocyte-Associated Ig-like Receptor-1 Is a Novel Inhibitory Receptor for Surfactant Protein D. J. Leukoc. Biol. 2014, 96, 105–111. [Google Scholar] [CrossRef]
- Son, M.; Diamond, B. C1q-Mediated Repression of Human Monocytes Is Regulated by Leukocyte-Associated Ig-Like Receptor 1 (LAIR-1). Mol. Med. 2015, 20, 559–568. [Google Scholar] [CrossRef]
- Lebbink, R.J.; van den Berg, M.C.W.; de Ruiter, T.; Raynal, N.; van Roon, J.A.G.; Lenting, P.J.; Jin, B.; Meyaard, L. The Soluble Leukocyte-Associated Ig-like Receptor (LAIR)-2 Antagonizes the Collagen/LAIR-1 Inhibitory Immune Interaction. J. Immunol. 2008, 180, 1662–1669. [Google Scholar] [CrossRef]
- Son, M.; Porat, A.; He, M.; Suurmond, J.; Santiago-Schwarz, F.; Andersson, U.; Coleman, T.R.; Volpe, B.T.; Tracey, K.J.; Al-Abed, Y.; et al. C1q and HMGB1 Reciprocally Regulate Human Macrophage Polarization. Blood 2016, 128, 2218–2228. [Google Scholar] [CrossRef]
- Son, M.; Diamond, B.; Volpe, B.T.; Aranow, C.B.; Mackay, M.C.; Santiago-Schwarz, F. Evidence for C1q-Mediated Crosslinking of CD33/LAIR-1 Inhibitory Immunoreceptors and Biological Control of CD33/LAIR-1 Expression. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Bally, I.; Ancelet, S.; Moriscot, C.; Gonnet, F.; Mantovani, A.; Daniel, R.; Schoehn, G.; Arlaud, G.J.; Thielens, N.M. Expression of Recombinant Human Complement C1q Allows Identification of the C1r/C1s-Binding Sites. Proc. Natl. Acad. Sci. USA 2013, 110, 8650–8655. [Google Scholar] [CrossRef]
- Fouët, G.; Bally, I.; Signor, L.; Häußermann, K.; Thielens, N.M.; Rossi, V.; Gaboriaud, C. Headless C1q: A New Molecular Tool to Decipher Its Collagen-like Functions. FEBS J. 2020. [Google Scholar] [CrossRef] [PubMed]
- Brondijk, T.H.C.; de Ruiter, T.; Ballering, J.; Wienk, H.; Lebbink, R.J.; van Ingen, H.; Boelens, R.; Farndale, R.W.; Meyaard, L.; Huizinga, E.G. Crystal Structure and Collagen-Binding Site of Immune Inhibitory Receptor LAIR-1: Unexpected Implications for Collagen Binding by Platelet Receptor GPVI. Blood 2010, 115, 1364–1373. [Google Scholar] [CrossRef] [PubMed]
- Moroi, M.; Jung, S.M. Platelet Glycoprotein VI: Its Structure and Function. Thromb. Res. 2004, 114, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Persikov, A.V.; Ramshaw, J.A.M.; Kirkpatrick, A.; Brodsky, B. Electrostatic Interactions Involving Lysine Make Major Contributions to Collagen Triple-Helix Stability. Biochemistry 2005, 44, 1414–1422. [Google Scholar] [CrossRef]
- Persikov, A.V.; Ramshaw, J.A.M.; Brodsky, B. Prediction of Collagen Stability from Amino Acid Sequence. J. Biol. Chem. 2005, 280, 19343–19349. [Google Scholar] [CrossRef]
- Olde Nordkamp, M.J.M.; Boross, P.; Yildiz, C.; Jansen, J.H.M.; Leusen, J.H.W.; Wouters, D.; Urbanus, R.T.; Hack, C.E.; Meyaard, L. Inhibition of the Classical and Lectin Pathway of the Complement System by Recombinant LAIR-2. J. Innate Immun. 2014, 6, 284–292. [Google Scholar] [CrossRef]
- Reid, K.B. Complete Amino Acid Sequences of the Three Collagen-like Regions Present in Subcomponent C1q of the First Component of Human Complement. Biochem. J. 1979, 179, 367–371. [Google Scholar] [CrossRef]
- Pflieger, D.; Przybylski, C.; Gonnet, F.; Le Caer, J.-P.; Lunardi, T.; Arlaud, G.J.; Daniel, R. Analysis of Human C1q by Combined Bottom-up and Top-down Mass Spectrometry. Mol. Cell Proteom. 2010, 9, 593–610. [Google Scholar] [CrossRef]
- Pieper, K.; Tan, J.; Piccoli, L.; Foglierini, M.; Barbieri, S.; Chen, Y.; Silacci-Fregni, C.; Wolf, T.; Jarrossay, D.; Anderle, M.; et al. Public Antibodies to Malaria Antigens Generated by Two LAIR1 Insertion Modalities. Nature 2017, 548, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Pieper, K.; Piccoli, L.; Abdi, A.; Foglierini, M.; Geiger, R.; Maria Tully, C.; Jarrossay, D.; Maina Ndungu, F.; Wambua, J.; et al. A LAIR1 Insertion Generates Broadly Reactive Antibodies against Malaria Variant Antigens. Nature 2016, 529, 105–109. [Google Scholar] [CrossRef]
- Jacquet, M.; Cioci, G.; Fouet, G.; Bally, I.; Thielens, N.M.; Gaboriaud, C.; Rossi, V. C1q and Mannose-Binding Lectin Interact with CR1 in the Same Region on CCP24-25 Modules. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Fouët, G.; Gout, E.; Wicker-Planquart, C.; Bally, I.; De Nardis, C.; Dedieu, S.; Chouquet, A.; Gaboriaud, C.; Thielens, N.M.; Kleman, J.-P.; et al. Complement C1q Interacts with LRP1 Clusters II and IV Through a Site Close but Different From the Binding Site of Its C1r and C1s-Associated Proteases. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Sim, R.B.; Arlaud, G.J.; Colomb, M.G. C1 Inhibitor-Dependent Dissociation of Human Complement Component C1 Bound to Immune Complexes. Biochem. J. 1979, 179, 449–457. [Google Scholar] [CrossRef]
- Kouser, L.; Madhukaran, S.P.; Shastri, A.; Saraon, A.; Ferluga, J.; Al-Mozaini, M.; Kishore, U. Emerging and Novel Functions of Complement Protein C1q. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef]
- Bohlson, S.S.; O’Conner, S.D.; Hulsebus, H.J.; Ho, M.-M.; Fraser, D.A. Complement, C1q, and C1q-Related Molecules Regulate Macrophage Polarization. Front. Immunol. 2014, 5, 402. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Xiang, A.; Peng, T.; Doran, A.C.; Tracey, K.J.; Barnes, B.J.; Tabas, I.; Son, M.; Diamond, B. HMGB1-C1q Complexes Regulate Macrophage Function by Switching between Leukotriene and Specialized Proresolving Mediator Biosynthesis. Proc. Natl. Acad. Sci. USA 2019, 116, 23254–23263. [Google Scholar] [CrossRef]
- Xu, M.J.; Zhao, R.; Zhao, Z.J. Identification and Characterization of Leukocyte-Associated Ig-like Receptor-1 as a Major Anchor Protein of Tyrosine Phosphatase SHP-1 in Hematopoietic Cells. J. Biol. Chem. 2000, 275, 17440–17446. [Google Scholar] [CrossRef] [PubMed]
- Sathish, J.G.; Johnson, K.G.; Fuller, K.J.; LeRoy, F.G.; Meyaard, L.; Sims, M.J.; Matthews, R.J. Constitutive Association of SHP-1 with Leukocyte-Associated Ig-Like Receptor-1 in Human T Cells. J. Immunol. 2001, 166, 1763–1770. [Google Scholar] [CrossRef]
- Jin, J.; Wang, Y.; Ma, Q.; Wang, N.; Guo, W.; Jin, B.; Fang, L.; Chen, L. LAIR-1 Activation Inhibits Inflammatory Macrophage Phenotype in Vitro. Cell. Immunol. 2018, 331, 78–84. [Google Scholar] [CrossRef]
- Arlaud, J.; Sim, B.; Colomb, G. Differential Elution of Clq, Clr and Cls from Human Cl Bound to Immune Aggregates. Use in the Rapid Purification of Cl Subcomponents. Mol. Immunol. 1979, 16, 445–450. [Google Scholar] [CrossRef]
- Tacnet-Delorme, P.; Chevallier, S.; Arlaud, G.J. β-Amyloid Fibrils Activate the C1 Complex of Complement Under Physiological Conditions: Evidence for a Binding Site for Aβ on the C1q Globular Regions. J. Immunol. 2001, 167, 6374–6381. [Google Scholar] [CrossRef]
- Bally, I.; Inforzato, A.; Dalonneau, F.; Stravalaci, M.; Bottazzi, B.; Gaboriaud, C.; Thielens, N.M. Interaction of C1q With Pentraxin 3 and IgM Revisited: Mutational Studies with Recombinant C1q Variants. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.; Veit, G.; Stricker, S.; Bhatt, P.; Kutsch, S.; Zhou, P.; Reinders, E.; Hahn, R.A.; Song, R.; Burgeson, R.E.; et al. Expression of Type XXIII Collagen MRNA and Protein. J. Biol. Chem. 2006, 281, 21546–21557. [Google Scholar] [CrossRef] [PubMed]
- Studier, F.W. Protein Production by Auto-Induction in High-Density Shaking Cultures. Protein Expr. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immobilized Ligands | LAIR-1 WT KD (µM) |
|---|---|
| sC1q | 112.3 ± 6.2 (n = 4) |
| rC1q WT | 99.5 ± 26.6 (n = 2) |
| rC1q ABC | 131.8 ± 23.5 (n = 2) |
| C1qCLR_nc2 | 179.3 ± 4.8 (n = 3) |
| C1qCLR_nc2 AR72E | 295.0 ± 10.6 (n = 3) |
| C1qCLR_nc2 BK65E | 188.8 ± 5.3 (n = 3) |
| C1qstem_nc2 | 174.9 ± 0.6 (n = 2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fouët, G.; Bally, I.; Chouquet, A.; Reiser, J.-B.; Thielens, N.M.; Gaboriaud, C.; Rossi, V. Molecular Basis of Complement C1q Collagen-Like Region Interaction with the Immunoglobulin-Like Receptor LAIR-1. Int. J. Mol. Sci. 2021, 22, 5125. https://doi.org/10.3390/ijms22105125
Fouët G, Bally I, Chouquet A, Reiser J-B, Thielens NM, Gaboriaud C, Rossi V. Molecular Basis of Complement C1q Collagen-Like Region Interaction with the Immunoglobulin-Like Receptor LAIR-1. International Journal of Molecular Sciences. 2021; 22(10):5125. https://doi.org/10.3390/ijms22105125
Chicago/Turabian StyleFouët, Guillaume, Isabelle Bally, Anne Chouquet, Jean-Baptiste Reiser, Nicole M. Thielens, Christine Gaboriaud, and Véronique Rossi. 2021. "Molecular Basis of Complement C1q Collagen-Like Region Interaction with the Immunoglobulin-Like Receptor LAIR-1" International Journal of Molecular Sciences 22, no. 10: 5125. https://doi.org/10.3390/ijms22105125
APA StyleFouët, G., Bally, I., Chouquet, A., Reiser, J.-B., Thielens, N. M., Gaboriaud, C., & Rossi, V. (2021). Molecular Basis of Complement C1q Collagen-Like Region Interaction with the Immunoglobulin-Like Receptor LAIR-1. International Journal of Molecular Sciences, 22(10), 5125. https://doi.org/10.3390/ijms22105125