Suppression of the Reactive Oxygen Response Alleviates Experimental Autoimmune Uveitis in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. ROS Deficiency due to the absence of Ncf1 Decreases the Severity of EAU in Mice with Reduced Levels of Oxidative Stress, Inflammatory Mediators, and NF-ĸB Activation in the Retina
2.1.1. Ncf1 Deficiency Reduces Malondialdehyde Levels in the Retinas and Spleens of Mice with EAU Induction
2.1.2. Absence of Ncf1 Reduces EAU Severity in Mice
2.1.3. The Influence of Ncf1 Deficiency on the Levels of Inflammatory Mediators in the Retinas of Mice with EAU Induction
2.1.4. Absence of Ncf1 Reduces NF-κB Activation in the Retinas of Mice with EAU Induction
2.2. Treatment with the Antioxidant NAC Reduces the Severity of EAU in Mice with Reduced Levels of Oxidative Stress, Inflammatory Mediators, and NF-ĸB Activation in the Retina
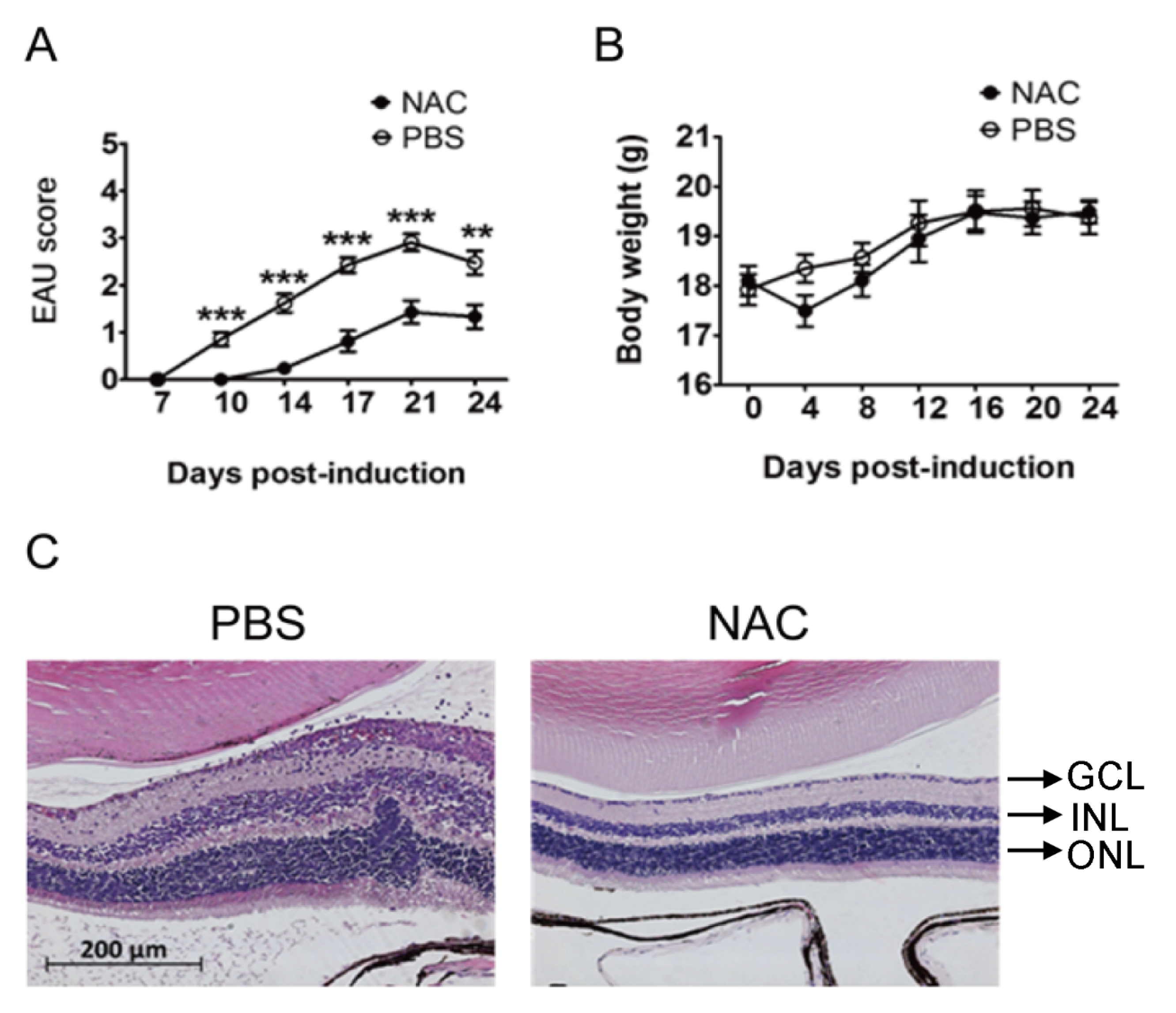
2.2.1. NAC Treatment Reduces EAU Severity in Mice
2.2.2. The Influence of NAC Treatment on the Levels of Inflammatory Mediators and NF-κB Activation in the Retinas of Mice with EAU Induction
3. Discussion
4. Materials and Methods
4.1. EAU Induction and NAC Treatment in Mice
4.2. Malondialdehyde Assays
4.3. Luminex Assay
4.4. Histological, Immunohistochemical, and Immunofluorescence Staining
4.5. EMSA of NF-κB
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ROS | Reactive oxygen species |
| EAU | Experimental autoimmune uveitis |
| NAC | N-acetylcysteine |
| IRBP | Interphotoreceptor retinoid binding protein |
| iNOS | inducible nitric oxide synthase |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NOX2 | NADPH oxidase 2 |
| RNS | Reactive nitrogen species |
| CGD | Chronic granulomatous disease |
| Ncf1 | Neutrophil cytosolic factor 1 |
| IL | Interleukin |
| IFN | Interferon |
| TNF | Tumor necrosis factor |
| MCP | Monocyte chemoattractant protein |
References
- Wakefield, D.; Chang, J.H. Epidemiology of uveitis. Int. Ophthalmol. Clin. 2005, 45, 1–13. [Google Scholar] [CrossRef]
- Gonzalez, M.M.; Solano, M.M.; Porco, T.C.; Oldenburg, C.E.; Acharya, N.R.; Lin, S.C.; Chan, M.F. Epidemiology of uveitis in a US population-based study. J. Ophthalmic Inflamm. Infect. 2018, 8, 6. [Google Scholar] [CrossRef]
- Castiblanco, C.P.; Adelman, R.A. Sympathetic ophthalmia. Graefes Arch. Clin. Exp. Ophthalmol. 2009, 247, 289–302. [Google Scholar] [CrossRef]
- Chang, G.C.; Young, L.H. Sympathetic ophthalmia. Semin. Ophthalmol. 2011, 26, 316–320. [Google Scholar] [CrossRef]
- Hsu, S.M.; Mathew, R.; Taylor, A.W.; Stein-Streilein, J. Ex-vivo tolerogenic F4/80(+) antigen-presenting cells (APC) induce efferent CD8(+) regulatory T cell-dependent suppression of experimental autoimmune uveitis. Clin. Exp. Immunol. 2014, 176, 37–48. [Google Scholar] [CrossRef]
- Hsu, S.M.; Yang, C.H.; Shen, F.H.; Chen, S.H.; Lin, C.J.; Shieh, C.C. Proteasome inhibitor bortezomib suppresses nuclear factor-kappa B activation and ameliorates eye inflammation in experimental autoimmune uveitis. Mediat. Inflamm. 2015, 2015, 847373. [Google Scholar] [CrossRef]
- Gritz, D.C.; Wong, I.G. Incidence and prevalence of uveitis in Northern California; the Northern California epidemiology of uveitis study. Ophthalmology 2004, 111, 491–500, discussion 500. [Google Scholar] [CrossRef]
- Namba, K.; Kitaichi, N.; Nishida, T.; Taylor, A.W. Induction of regulatory T cells by the immunomodulating cytokines alpha-melanocyte-stimulating hormone and transforming growth factor-beta2. J. Leukoc. Biol. 2002, 72, 946–952. [Google Scholar]
- Avichezer, D.; Liou, G.I.; Chan, C.C.; Lewis, G.M.; Wiggert, B.; Donoso, L.A.; Nickerson, J.M.; Crawford, M.A.; Caspi, R.R. Interphotoreceptor retinoid-binding protein (IRBP)-deficient C57BL/6 mice have enhanced immunological and immunopathogenic responses to IRBP and an altered recognition of IRBP epitopes. J. Autoimmun. 2003, 21, 185–194. [Google Scholar] [CrossRef]
- Rajendram, R.; Saraswathy, S.; Rao, N.A. Photoreceptor mitochondrial oxidative stress in early experimental autoimmune uveoretinitis. Br. J. Ophthalmol. 2007, 91, 531–537. [Google Scholar] [CrossRef]
- Nguyen, A.M.; Rao, N.A. Oxidative photoreceptor cell damage in autoimmune uveitis. J. Ophthalmic Inflamm. Infect. 2011, 1, 7–13. [Google Scholar] [CrossRef]
- Saraswathy, S.; Rao, N.A. Photoreceptor mitochondrial oxidative stress in experimental autoimmune uveitis. Ophthalmic Res. 2008, 40, 160–164. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef]
- Yeh, P.T.; Huang, H.W.; Yang, C.M.; Yang, W.S.; Yang, C.H. Astaxanthin Inhibits Expression of Retinal Oxidative Stress and Inflammatory Mediators in Streptozotocin-Induced Diabetic Rats. PLoS ONE 2016, 11, e0146438. [Google Scholar] [CrossRef]
- Fang, F.C. Antimicrobial reactive oxygen and nitrogen species: Concepts and controversies. Nat. Rev. Microbiol. 2004, 2, 820–832. [Google Scholar] [CrossRef]
- Rada, B.; Leto, T.L. Oxidative innate immune defenses by Nox/Duox family NADPH oxidases. Contrib. Microbiol. 2008, 15, 164–187. [Google Scholar] [CrossRef]
- Segal, B.H.; Leto, T.L.; Gallin, J.I.; Malech, H.L.; Holland, S.M. Genetic, biochemical, and clinical features of chronic granulomatous disease. Medicine 2000, 79, 170–200. [Google Scholar] [CrossRef]
- Arnold, D.E.; Heimall, J.R. A Review of Chronic Granulomatous Disease. Adv Ther 2017, 34, 2543–2557. [Google Scholar] [CrossRef]
- Magnani, A.; Brosselin, P.; Beaute, J.; de Vergnes, N.; Mouy, R.; Debre, M.; Suarez, F.; Hermine, O.; Lortholary, O.; Blanche, S.; et al. Inflammatory manifestations in a single-center cohort of patients with chronic granulomatous disease. J. Allergy Clin. Immunol. 2014, 134, 655–662.e8. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Hultqvist, M.; Olofsson, P.; Holmberg, J.; Backstrom, B.T.; Tordsson, J.; Holmdahl, R. Enhanced autoimmunity, arthritis, and encephalomyelitis in mice with a reduced oxidative burst due to a mutation in the Ncf1 gene. Proc. Natl. Acad. Sci. USA 2004, 101, 12646–12651. [Google Scholar] [CrossRef]
- Allan, E.R.; Tailor, P.; Balce, D.R.; Pirzadeh, P.; McKenna, N.T.; Renaux, B.; Warren, A.L.; Jirik, F.R.; Yates, R.M. NADPH oxidase modifies patterns of MHC class II-restricted epitopic repertoires through redox control of antigen processing. J. Immunol. 2014, 192, 4989–5001. [Google Scholar] [CrossRef]
- Jackson, S.H.; Gallin, J.I.; Holland, S.M. The p47phox mouse knock-out model of chronic granulomatous disease. J. Exp. Med. 1995, 182, 751–758. [Google Scholar] [CrossRef]
- Rio, D.D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Huang, C.K.; Zhan, L.; Hannigan, M.O.; Ai, Y.; Leto, T.L. P47(phox)-deficient NADPH oxidase defect in neutrophils of diabetic mouse strains, C57BL/6J-m db/db and db/+. J. Leukoc. Biol. 2000, 67, 210–215. [Google Scholar] [CrossRef]
- Yang, D.; Elner, S.G.; Bian, Z.M.; Till, G.O.; Petty, H.R.; Elner, V.M. Pro-inflammatory cytokines increase reactive oxygen species through mitochondria and NADPH oxidase in cultured RPE cells. Exp. Eye Res. 2007, 85, 462–472. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Caspi, R.R. Th1 and Th2 responses in pathogenesis and regulation of experimental autoimmune uveoretinitis. Int. Rev. Immunol. 2002, 21, 197–208. [Google Scholar] [CrossRef]
- Gloire, G.; Legrand-Poels, S.; Piette, J. NF-kappaB activation by reactive oxygen species: Fifteen years later. Biochem. Pharmacol. 2006, 72, 1493–1505. [Google Scholar] [CrossRef]
- Horai, R.; Caspi, R.R. Cytokines in autoimmune uveitis. J. Interferon Cytokine Res. 2011, 31, 733–744. [Google Scholar] [CrossRef]
- Awane, M.; Andres, P.G.; Li, D.J.; Reinecker, H.C. NF-kappa B-inducing kinase is a common mediator of IL-17-, TNF-alpha-, and IL-1 beta-induced chemokine promoter activation in intestinal epithelial cells. J. Immunol. 1999, 162, 5337–5344. [Google Scholar] [PubMed]
- Yue, Y.; Stone, S.; Lin, W. Role of nuclear factor kappaB in multiple sclerosis and experimental autoimmune encephalomyelitis. Neural Regen. Res. 2018, 13, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Moldeus, P.; Cotgreave, I.A.; Berggren, M. Lung protection by a thiol-containing antioxidant: N-acetylcysteine. Respir. Int. Rev. Thorac. Dis. 1986, 50, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Decramer, M.; Rutten-van Molken, M.; Dekhuijzen, P.N.; Troosters, T.; van Herwaarden, C.; Pellegrino, R.; van Schayck, C.P.; Olivieri, D.; Donno, M.D.; de Backer, W.; et al. Effects of N-acetylcysteine on outcomes in chronic obstructive pulmonary disease (bronchitis randomized on NAC cost-utility study, BRONCUS): A randomised placebo-controlled trial. Lancet 2005, 365, 1552–1560. [Google Scholar] [CrossRef]
- van Zandwijk, N. N-acetylcysteine (NAC) and glutathione (GSH): Antioxidant and chemopreventive properties, with special reference to lung cancer. J. Cell. Biochem. Suppl. 1995, 22, 24–32. [Google Scholar] [CrossRef]
- Ljubisavljevic, S.; Stojanovic, I.; Pavlovic, D.; Sokolovic, D.; Stevanovic, I. Aminoguanidine and N-acetyl-cysteine supress oxidative and nitrosative stress in EAE rat brains. Redox Rep. 2011, 16, 166–172. [Google Scholar] [CrossRef]
- de Senzi Moraes Pinto, R.; Ferretti, R.; Moraes, L.H.; Neto, H.S.; Marques, M.J.; Minatel, E. N-acetylcysteine treatment reduces TNF-alpha levels and myonecrosis in diaphragm muscle of mdx mice. Clin. Nutr. 2013, 32, 472–475. [Google Scholar] [CrossRef]
- Cao, L.; Li, L.; Zuo, Z. N-acetylcysteine reverses existing cognitive impairment and increased oxidative stress in glutamate transporter type 3 deficient mice. Neuroscience 2012, 220, 85–89. [Google Scholar] [CrossRef]
- Lehmann, D.; Karussis, D.; Misrachi-Koll, R.; Shezen, E.; Ovadia, H.; Abramsky, O. Oral administration of the oxidant-scavenger N-acetyl-L-cysteine inhibits acute experimental autoimmune encephalomyelitis. J. Neuroimmunol. 1994, 50, 35–42. [Google Scholar] [CrossRef]
- Stanislaus, R.; Gilg, A.G.; Singh, A.K.; Singh, I. N-acetyl-L-cysteine ameliorates the inflammatory disease process in experimental autoimmune encephalomyelitis in Lewis rats. J. Autoimmune Dis. 2005, 2, 4. [Google Scholar] [CrossRef]
- Khurana, R.N.; Parikh, J.G.; Saraswathy, S.; Wu, G.S.; Rao, N.A. Mitochondrial oxidative DNA damage in experimental autoimmune uveitis. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3299–3304. [Google Scholar] [CrossRef] [PubMed]
- Silver, P.B.; Tarrant, T.K.; Chan, C.C.; Wiggert, B.; Caspi, R.R. Mice deficient in inducible nitric oxide synthase are susceptible to experimental autoimmune uveoretinitis. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1280–1284. [Google Scholar]
- Charlotte, F.; Ito, S.; Wu, G.; Kimoto, T.; Rao, N. Highly selective inhibitor of inducible nitric oxide synthase enhances S-antigen-induced uveitis. Curr. Eye Res. 2003, 26, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Avichezer, D.; Chan, C.C.; Silver, P.B.; Wiggert, B.; Caspi, R.R. Residues 1-20 of IRBP and whole IRBP elicit different uveitogenic and immunological responses in interferon gamma deficient mice. Exp. Eye Res. 2000, 71, 111–118. [Google Scholar] [CrossRef]
- Weinstein, J.E.; Pepple, K.L. Cytokines in uveitis. Curr. Opin. Ophthalmol. 2018, 29, 267–274. [Google Scholar] [CrossRef]
- Daudin, J.B.; Monnet, D.; Kavian, N.; Espy, C.; Wang, A.; Chereau, C.; Goulvestre, C.; Omri, S.; Brezin, A.; Weill, B.; et al. Protective effect of pristane on experimental autoimmune uveitis. Immunol. Lett. 2011, 141, 83–93. [Google Scholar] [CrossRef]
- Freitas, E.C.; de Oliveira, M.S.; Monticielo, O.A. Pristane-induced lupus: Considerations on this experimental model. Clin. Rheumatol. 2017, 36, 2403–2414. [Google Scholar] [CrossRef]
- Tuncel, J.; Haag, S.; Hoffmann, M.H.; Yau, A.C.; Hultqvist, M.; Olofsson, P.; Backlund, J.; Nandakumar, K.S.; Weidner, D.; Fischer, A.; et al. Animal models of rheumatoid arthritis (I): Pristane-induced arthritis in the rat. PLoS ONE 2016, 11, e0155936. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, S.-M.; Yang, C.-H.; Teng, Y.-T.; Tsai, H.-Y.; Lin, C.-Y.; Lin, C.-J.; Shieh, C.-C.; Chen, S.-H. Suppression of the Reactive Oxygen Response Alleviates Experimental Autoimmune Uveitis in Mice. Int. J. Mol. Sci. 2020, 21, 3261. https://doi.org/10.3390/ijms21093261
Hsu S-M, Yang C-H, Teng Y-T, Tsai H-Y, Lin C-Y, Lin C-J, Shieh C-C, Chen S-H. Suppression of the Reactive Oxygen Response Alleviates Experimental Autoimmune Uveitis in Mice. International Journal of Molecular Sciences. 2020; 21(9):3261. https://doi.org/10.3390/ijms21093261
Chicago/Turabian StyleHsu, Sheng-Min, Chang-Hao Yang, Yu-Ti Teng, Hsien-Yang Tsai, Chieh-Yu Lin, Chia-Jhen Lin, Chi-Chang Shieh, and Shun-Hua Chen. 2020. "Suppression of the Reactive Oxygen Response Alleviates Experimental Autoimmune Uveitis in Mice" International Journal of Molecular Sciences 21, no. 9: 3261. https://doi.org/10.3390/ijms21093261
APA StyleHsu, S.-M., Yang, C.-H., Teng, Y.-T., Tsai, H.-Y., Lin, C.-Y., Lin, C.-J., Shieh, C.-C., & Chen, S.-H. (2020). Suppression of the Reactive Oxygen Response Alleviates Experimental Autoimmune Uveitis in Mice. International Journal of Molecular Sciences, 21(9), 3261. https://doi.org/10.3390/ijms21093261