The Novel ncRNA OsiR Positively Regulates Expression of katE2 and is Required for Oxidative Stress Tolerance in Deinococcus radiodurans

 ,
,
Abstract
1. Introduction
2. Results
2.1. Expression of osiR Is Induced by Oxidative Stress
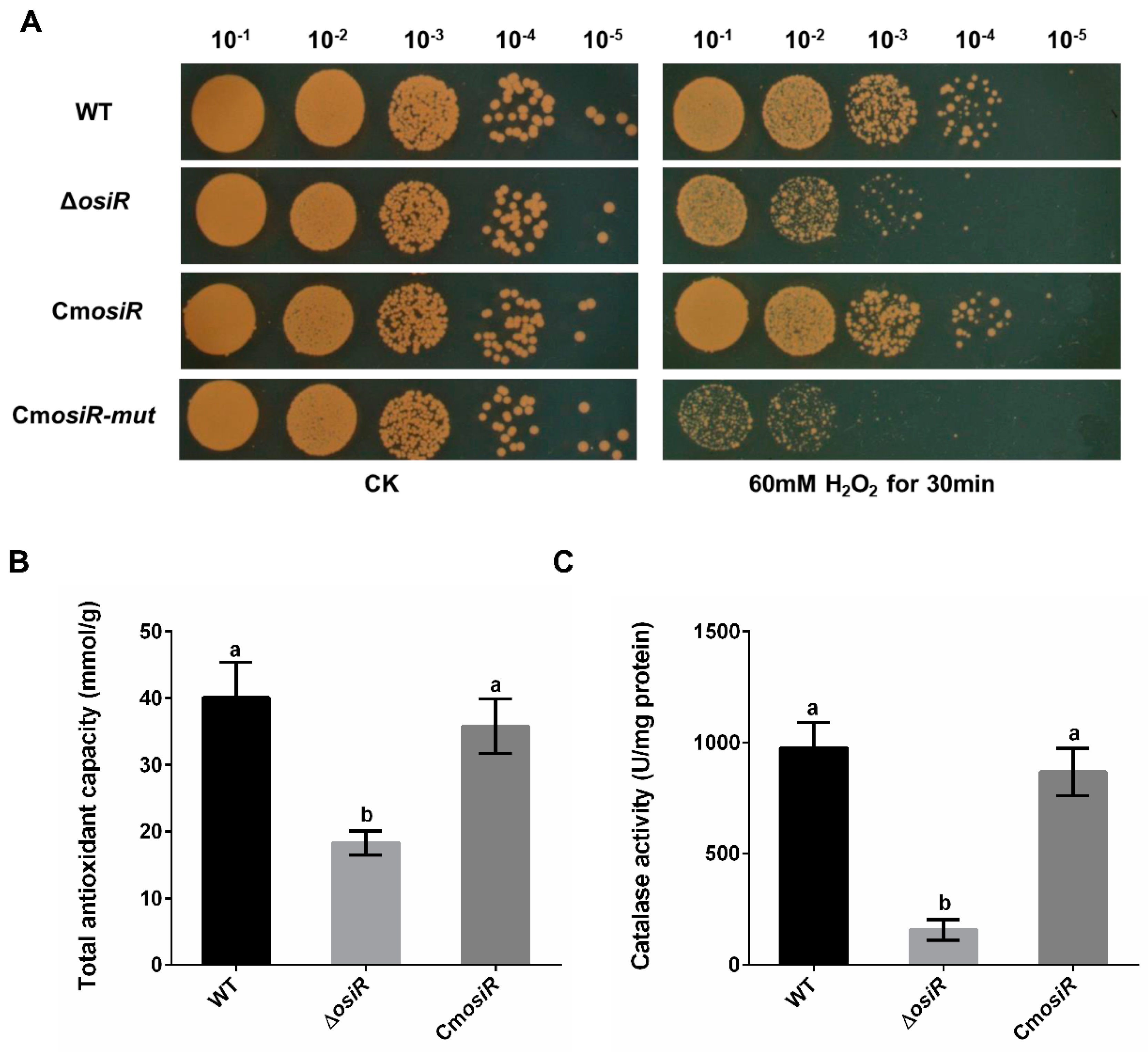
2.2. OsiR Contributes to the Oxidative Stress Tolerance in D. radiodurans
2.3. katE2 Is a Target of OsiR in Response to Oxidative Stress
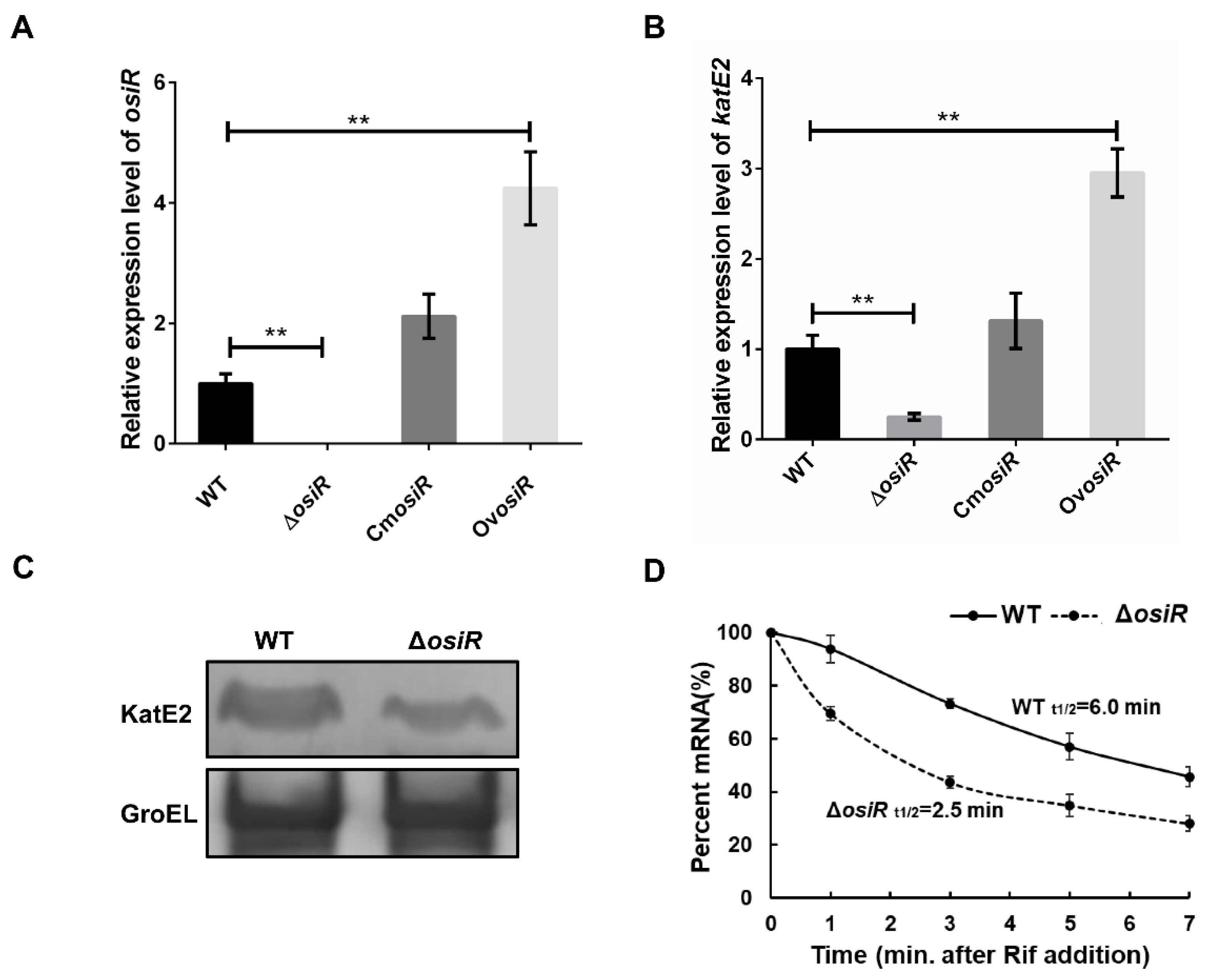
2.4. OsiR Positively Regulates the Expression of katE2
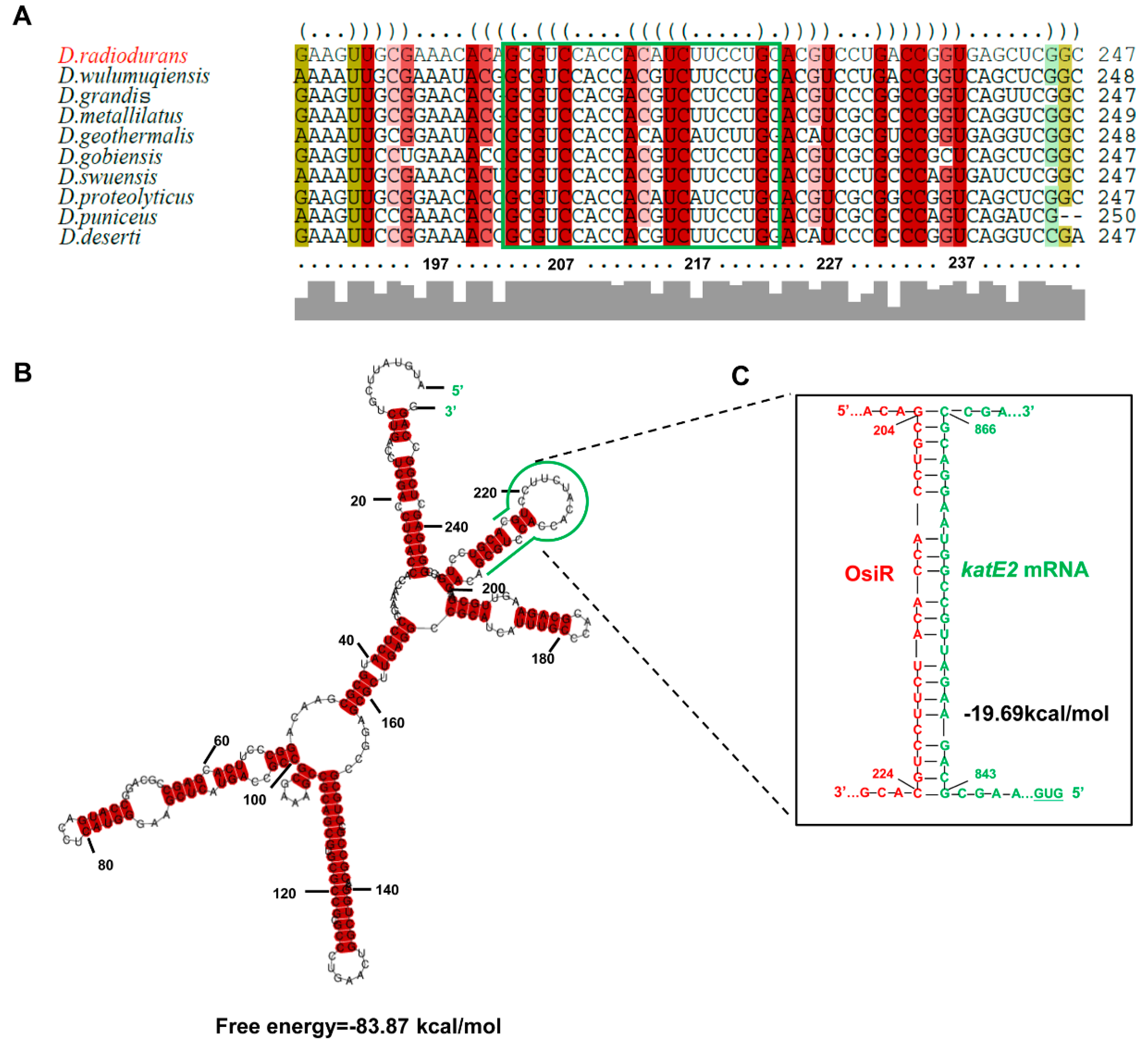
2.5. OsiR Directly Base Pairs With katE2 mRNA to Enhance Oxidative Stress Tolerance in D. radiodurans
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, and Growth Conditions
4.2. Nucleotide Sequence
4.3. Construction of the Deletion Mutants and Complementation Strains
4.4. Quantitative Real-Time PCR (qRT-PCR)
4.5. 5‘RACE
4.6. Northern Blot Analysis
4.7. Western Blot Analysis
4.8. Oxidative Stress Survival Assays
4.9. Half-Life Experiment
4.10. Determination of Total Antioxidant Capacity and Catalase Activity
4.11. Microscale Thermophoresis Measurements (MST)
5. Patents
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Battista, J.R.; Earl, A.M.; Park, M.J. Why is Deinococcus Radiodurans so Resistant to Ionizing Radiation? Trends Microbiol. 1999, 7, 362–365. [Google Scholar] [CrossRef]
- Makarova, K.S.; Aravind, L.; Wolf, Y.I.; Tatusov, R.L.; Minton, K.W.; Koonin, E.V.; Daly, M.J. Genome of the Extremely Radiation-Resistant Bacterium Deinococcus Radiodurans Viewed from the Perspective of Comparative Genomics. Microbiol. Mol. Biol. Rev. 2001, 65, 44–79. [Google Scholar] [CrossRef] [PubMed]
- Englander, J.; Klein, E.; Brumfeld, V.; Sharma, A.K.; Doherty, A.J.; Minsky, A. DNA Toroids: Framework for DNA Repair in Deinococcus Radiodurans and in Germinating Bacterial Spores. J. Bacteriol. 2004, 186, 5973–5977. [Google Scholar] [CrossRef] [PubMed]
- Slade, D.; Radman, M. Oxidative Stress Resistance in Deinococcus Radiodurans. Microbiol. Mol. Biol. Rev. 2011, 75, 133–191. [Google Scholar] [CrossRef] [PubMed]
- Agapov, A.A.; Kulbachinskiy, A.V. Mechanisms of Stress Resistance and Gene Regulation in the Radioresistant Bacterium Deinococcus radiodurans. Biochem. (Moscow) 2015, 80, 1201–1216. [Google Scholar] [CrossRef] [PubMed]
- D’Autreaux, B.; Toledano, M.B. ROS as Signalling Molecules: Mechanisms that Generate Specificity in ROS Homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.J. A New Perspective On Radiation Resistance Based on Deinococcus Radiodurans. Nat. Rev. Microbiol. 2009, 7, 237–245. [Google Scholar] [CrossRef]
- Imlay, J.A. Cellular Defenses Against Superoxide and Hydrogen Peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef]
- Mishra, S.; Imlay, J. Why Do Bacteria Use so Many Enzymes to Scavenge Hydrogen Peroxide? Arch. Biochem. Biophys. 2012, 525, 145–160. [Google Scholar] [CrossRef]
- Battista, J.R. Against All Odds: The Survival Strategies of Deinococcus Radiodurans. Annu. Rev. Microbiol. 1997, 51, 203–224. [Google Scholar] [CrossRef]
- Lim, S.; Jung, J.H.; Blanchard, L.; de Groot, A. Conservation and Diversity of Radiation and Oxidative Stress Resistance Mechanisms in Deinococcus Species. Fems. Microbiol. Rev. 2019, 43, 19–52. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Wu, Y.; Sheng, D.; Zheng, Z.; Gao, G.; Hua, Y. Chemiluminescence Assay for Reactive Oxygen Species Scavenging Activities and Inhibition on Oxidative Damage of DNA in Deinococcus Radiodurans. Lumin. 2004, 19, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Schellhorn, H.E. Induction of Resistance to Hydrogen Peroxide and Radiation in Deinococcus Radiodurans. Can. J. Microbiol. 1995, 41, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, G.; Chen, H.; Zhao, Y.; Xu, N.; Tian, B.; Hua, Y. DrRRA: A Novel Response Regulator Essential for the Extreme Radioresistance of Deinococcus Radiodurans. Mol. Microbiol. 2008, 67, 1211–1222. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xu, G.; Zhao, Y.; Tian, B.; Lu, H.; Yu, X.; Xu, Z.; Ying, N.; Hu, S.; Hua, Y. A Novel OxyR Sensor and Regulator of Hydrogen Peroxide Stress with One Cysteine Residue in Deinococcus Radiodurans. PLoS ONE 2008, 3, e1602. [Google Scholar] [CrossRef]
- Jeong, S.W.; Seo, H.S.; Kim, M.K.; Choi, J.I.; Lim, H.M.; Lim, S. PprM is Necessary for Up-Regulation of katE1, Encoding the Major Catalase of Deinococcus Radiodurans, under Unstressed Culture Conditions. J. Microbiol. 2016, 54, 426–431. [Google Scholar] [CrossRef]
- Jeong, S.W.; Jung, J.H.; Kim, M.K.; Seo, H.S.; Lim, H.M.; Lim, S. The Three Catalases in Deinococcus Radiodurans: Only Two Show Catalase Activity. Biochem. Biophys. Res. Commun. 2016, 469, 443–448. [Google Scholar] [CrossRef]
- Nitzan, M.; Rehani, R.; Margalit, H. Integration of Bacterial Small RNAs in Regulatory Networks. Annu. Rev. Biophys. 2017, 46, 131–148. [Google Scholar] [CrossRef]
- Bouche, F.; Bouche, J.P. Genetic Evidence that DicF, a Second Division Inhibitor Encoded by the Escherichia coli dicB Operon, is Probably RNA. Mol. Microbiol. 1989, 3, 991–994. [Google Scholar] [CrossRef]
- Holmqvist, E.; Wagner, E.G.H. Impact of Bacterial sRNAs in Stress Responses. Biochem. Soc. Trans. 2017, 45, 1203–1212. [Google Scholar] [CrossRef]
- Papenfort, K.; Silpe, J.E.; Schramma, K.R.; Cong, J.P.; Seyedsayamdost, M.R.; Bassler, B.L. A Vibrio Cholerae Autoinducer-Receptor Pair that Controls Biofilm Formation. Nat. Chem. Biol. 2017, 13, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.F.; Micheva-Viteva, S.; Li, N.; Hong-Geller, E. Small RNA-mediated Regulation of Host-Pathogen Interactions. Virulence 2013, 4, 785–795. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Papenfort, K.; Vogel, J. Small RNA Functions in Carbon Metabolism and Virulence of Enteric Pathogens. Front Cell Infect. Microbiol. 2014, 4, 91. [Google Scholar] [CrossRef] [PubMed]
- Storz, G.; Vogel, J.; Wassarman, K.M. Regulation by Small RNAs in Bacteria: Expanding Frontiers. Mol. Cell 2011, 43, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.G.H.; Romby, P. Small RNAs in Bacteria and Archaea: Who They are, What They Do, and How They Do It. Adv. Genet. 2015, 90, 133–208. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Vogel, J. Multiple Target Regulation by Small Noncoding RNAs Rewires Gene Expression at the Post-Transcriptional Level. Res. Microbiol. 2009, 160, 278–287. [Google Scholar] [CrossRef]
- Lu, X.; Goodrich-Blair, H.; Tjaden, B. Assessing Computational Tools for the Discovery of Small RNA Genes in Bacteria. RNA 2011, 17, 1635–1647. [Google Scholar] [CrossRef]
- Waters, L.S.; Storz, G. Regulatory RNAs in Bacteria. Cell 2009, 136, 615–628. [Google Scholar] [CrossRef]
- Lalaouna, D.; Simoneau-Roy, M.; Lafontaine, D.; Masse, E. Regulatory RNAs and Target mRNA Decay in Prokaryotes. Biochim. Et Biophys. Acta (BBA)—Gene Regul. Mech. 2013, 1829, 742–747. [Google Scholar] [CrossRef]
- Altuvia, S.; Weinstein-Fischer, D.; Zhang, A.; Postow, L.; Storz, G. A Small, Stable RNA Induced by Oxidative Stress: Role as a Pleiotropic Regulator and Antimutator. Cell 1997, 90, 43–53. [Google Scholar] [CrossRef]
- De Lay, N.; Gottesman, S. A Complex Network of Small Non-Coding RNAs Regulate Motility in Escherichia Coli. Mol. Microbiol. 2012, 86, 524–538. [Google Scholar] [CrossRef] [PubMed]
- Tjaden, B.; Goodwin, S.S.; Opdyke, J.A.; Guillier, M.; Fu, D.X.; Gottesman, S.; Storz, G. Target Prediction for Small, Noncoding RNAs in Bacteria. Nucleic Acids Res. 2006, 34, 2791–2802. [Google Scholar] [CrossRef] [PubMed]
- Barshishat, S.; Elgrably-Weiss, M.; Edelstein, J.; Georg, J.; Govindarajan, S.; Haviv, M.; Wright, P.R.; Hess, W.R.; Altuvia, S. OxyS Small RNA Induces Cell Cycle Arrest to Allow DNA Damage Repair. EMBO J. 2018, 37, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhan, Y.; Yan, Y.; Liu, Y.; Hu, G.; Wang, S.; Yang, H.; Qiu, X.; Liu, Y.; Li, J.; et al. The Pseudomonas stutzeri-Specific Regulatory Noncoding RNA NfiS Targets katB mRNA Encoding a Catalase Essential for Optimal Oxidative Resistance and Nitrogenase Activity. J. Bacteriol. 2019, 201, e00334-19. [Google Scholar] [CrossRef]
- Chen, X.; Quinn, A.M.; Wolin, S.L. Ro Ribonucleoproteins Contribute to the Resistance of Deinococcus Radiodurans to Ultraviolet Irradiation. Genes Dev. 2000, 14, 777–782. [Google Scholar] [CrossRef]
- Chen, X.; Taylor, D.W.; Fowler, C.C.; Galan, J.E.; Wang, H.W.; Wolin, S.L. An RNA Degradation Machine Sculpted by Ro Autoantigen and Noncoding RNA. Cell 2013, 153, 166–177. [Google Scholar] [CrossRef]
- Tsai, C.H.; Liao, R.; Chou, B.; Contreras, L.M. Transcriptional Analysis of Deinococcus Radiodurans Reveals Novel Small RNAs that are Differentially Expressed under Ionizing Radiation. Appl. Environ. Microbiol. 2015, 81, 1754–1764. [Google Scholar] [CrossRef]
- Xue, D.; Chen, Y.; Li, J.; Han, J.; Liu, Y.; Jiang, S.; Zhou, Z.; Zhang, W.; Chen, M.; Lin, M.; et al. Targeting Hsp20 Using the Novel Small Non-Coding RNA DnrH Regulates Heat Tolerance in Deinococcus radiodurans. Front. Microbiol. 2019, 10, 2354. [Google Scholar] [CrossRef]
- Will, S.; Joshi, T.; Hofacker, I.L.; Stadler, P.F.; Backofen, R. LocARNA-P: Accurate Boundary Prediction and Improved Detection of Structural RNAs. RNA 2012, 18, 900–914. [Google Scholar] [CrossRef]
- Bernhart, S.H.; Hofacker, I.L.; Will, S.; Gruber, A.R.; Stadler, P.F. RNAalifold: Improved Consensus Structure Prediction for RNA Alignments. BMC Bioinform. 2008, 9, 474. [Google Scholar] [CrossRef]
- Zapf, R.L.; Wiemels, R.E.; Keogh, R.A.; Holzschu, D.L.; Howell, K.M.; Trzeciak, E.; Caillet, A.R.; King, K.A.; Selhorst, S.A.; Naldrett, M.J.; et al. The Small RNA Teg41 Regulates Expression of the Alpha Phenol-Soluble Modulins and is Required for Virulence in Staphylococcus aureus. mBio 2019, 10, e02484-18. [Google Scholar] [CrossRef] [PubMed]
- Amin, S.V.; Roberts, J.T.; Patterson, D.G.; Coley, A.B.; Allred, J.A.; Denner, J.M.; Johnson, J.P.; Mullen, G.E.; O’Neal, T.K.; Smith, J.T.; et al. Novel Small RNA (sRNA) Landscape of the Starvation-Stress Response Transcriptome of Salmonella Enterica Serovar Typhimurium. RNA Biol. 2016, 13, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Michaux, C.; Verneuil, N.; Hartke, A.; Giard, J.C. Physiological Roles of Small RNA Molecules. Microbiology 2014, 160, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Grenga, L.; Little, R.H.; Malone, J.G. Quick Change: Post-Transcriptional Regulation in Pseudomonas. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed]
- Lenz, D.H.; Mok, K.C.; Lilley, B.N.; Kulkarni, R.V.; Wingreen, N.S.; Bassler, B.L. The Small RNA Chaperone Hfq and Multiple Small RNAs Control Quorum Sensing in Vibrio Harveyi and Vibrio Cholerae. Cell 2004, 118, 69–82. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, J.; Liu, X.; Liu, Y.; Guo, C.; Zhang, L.; Han, J.; Wu, X.; Xue, D.; Gomaa, A.E.; et al. DrwH, a Novel WHy Domain-Containing Hydrophobic LEA5C Protein from Deinococcus radiodurans, Protects Enzymatic Activity under Oxidative Stress. Sci. Rep. 2017, 7, 9281. [Google Scholar] [CrossRef]
- Appukuttan, D.; Seo, H.S.; Jeong, S.; Im, S.; Joe, M.; Song, D.; Choi, J.; Lim, S. Expression and Mutational Analysis of DinB-Like Protein DR0053 in Deinococcus radiodurans. PLoS ONE 2015, 10, e118275. [Google Scholar] [CrossRef]
- Zhan, Y.; Yan, Y.; Deng, Z.; Chen, M.; Lu, W.; Lu, C.; Shang, L.; Yang, Z.; Zhang, W.; Wang, W.; et al. The Novel Regulatory ncRNA, NfiS, Optimizes Nitrogen Fixation Via Base Pairing with the Nitrogenase Gene nifK mRNA in Pseudomonas Stutzeri A1501. Proc. Natl. Acad. Sci. USA 2016, 113, E4348–E4356. [Google Scholar] [CrossRef]
- Lippok, S.; Seidel, S.A.; Duhr, S.; Uhland, K.; Holthoff, H.P.; Jenne, D.; Braun, D. Direct Detection of Antibody Concentration and Affinity in Human Serum Using Microscale Thermophoresis. Anal. Chem. 2012, 84, 3523–3530. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains/Plasmids | Relevant Characteristics | Source |
|---|---|---|
| Deinococcus radiodurans | Wild type | Lab stock |
osiR | D. radiodurans osiR-deletion mutant, Kmr | This study |
katE2 | D. radiodurans katE2-deletion mutant, Kmr | This study |
| CmosiR | osiR containing the complementation plasmid pRADZ3-osiR, Kmr, Cmr | This study |
| CmosiR-mut | osiR containing the complementation plasmid pRADZ3-osiR-mut, Kmr, Cmr | This study |
| CmkatE2 | katE2 containing the complementation plasmid pRADZ3-katE2, Kmr, Cmr | This study |
| OvosiR | D. radiodurans containing the plasmid pRADZ3-osiR, Cmr | This study |
| Escherichia coli | ||
| Trans 10 Trans 10 Z3-osiR Trans 10 Z3-katE2 Trans 10 Z3-osiR-mut | Host for cloning vectors As trans 10 with pZ3-osiR As trans 10 with pZ3-katE2 As trans 10 with pZ3-osiR-mut | TransGen This study This study This study |
| Plasmids pRADZ3 | Shuttle plasmid between E. coli and D. radiodurans, ampicillin in E. coli chloromycetin in D. radiodurans | Lab stock |
| posiR-wt | pRADZ3-derived plasmid carrying the wildtype osiR gene, Cmr, Ampr | This study |
| pkatE2-wt | pRADZ3-derived plasmid carrying the wildtype katE2 gene, Cmr, Ampr | This study |
| posiR-mut | pRADZ3-derived plasmid carrying the mutant osiR gene, Cmr, Ampr | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, L.; Chen, X.; Tian, Y.; Yan, Y.; Zhan, Y.; Zhou, Z.; Zhang, W.; Lin, M.; Chen, M. The Novel ncRNA OsiR Positively Regulates Expression of katE2 and is Required for Oxidative Stress Tolerance in Deinococcus radiodurans. Int. J. Mol. Sci. 2020, 21, 3200. https://doi.org/10.3390/ijms21093200
Gao L, Chen X, Tian Y, Yan Y, Zhan Y, Zhou Z, Zhang W, Lin M, Chen M. The Novel ncRNA OsiR Positively Regulates Expression of katE2 and is Required for Oxidative Stress Tolerance in Deinococcus radiodurans. International Journal of Molecular Sciences. 2020; 21(9):3200. https://doi.org/10.3390/ijms21093200
Chicago/Turabian StyleGao, Lihua, Xiaonan Chen, Ye Tian, Yongliang Yan, Yuhua Zhan, Zhengfu Zhou, Wei Zhang, Min Lin, and Ming Chen. 2020. "The Novel ncRNA OsiR Positively Regulates Expression of katE2 and is Required for Oxidative Stress Tolerance in Deinococcus radiodurans" International Journal of Molecular Sciences 21, no. 9: 3200. https://doi.org/10.3390/ijms21093200
APA StyleGao, L., Chen, X., Tian, Y., Yan, Y., Zhan, Y., Zhou, Z., Zhang, W., Lin, M., & Chen, M. (2020). The Novel ncRNA OsiR Positively Regulates Expression of katE2 and is Required for Oxidative Stress Tolerance in Deinococcus radiodurans. International Journal of Molecular Sciences, 21(9), 3200. https://doi.org/10.3390/ijms21093200