LINC00312/YBX1 Axis Regulates Myofibroblast Activities in Oral Submucous Fibrosis



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
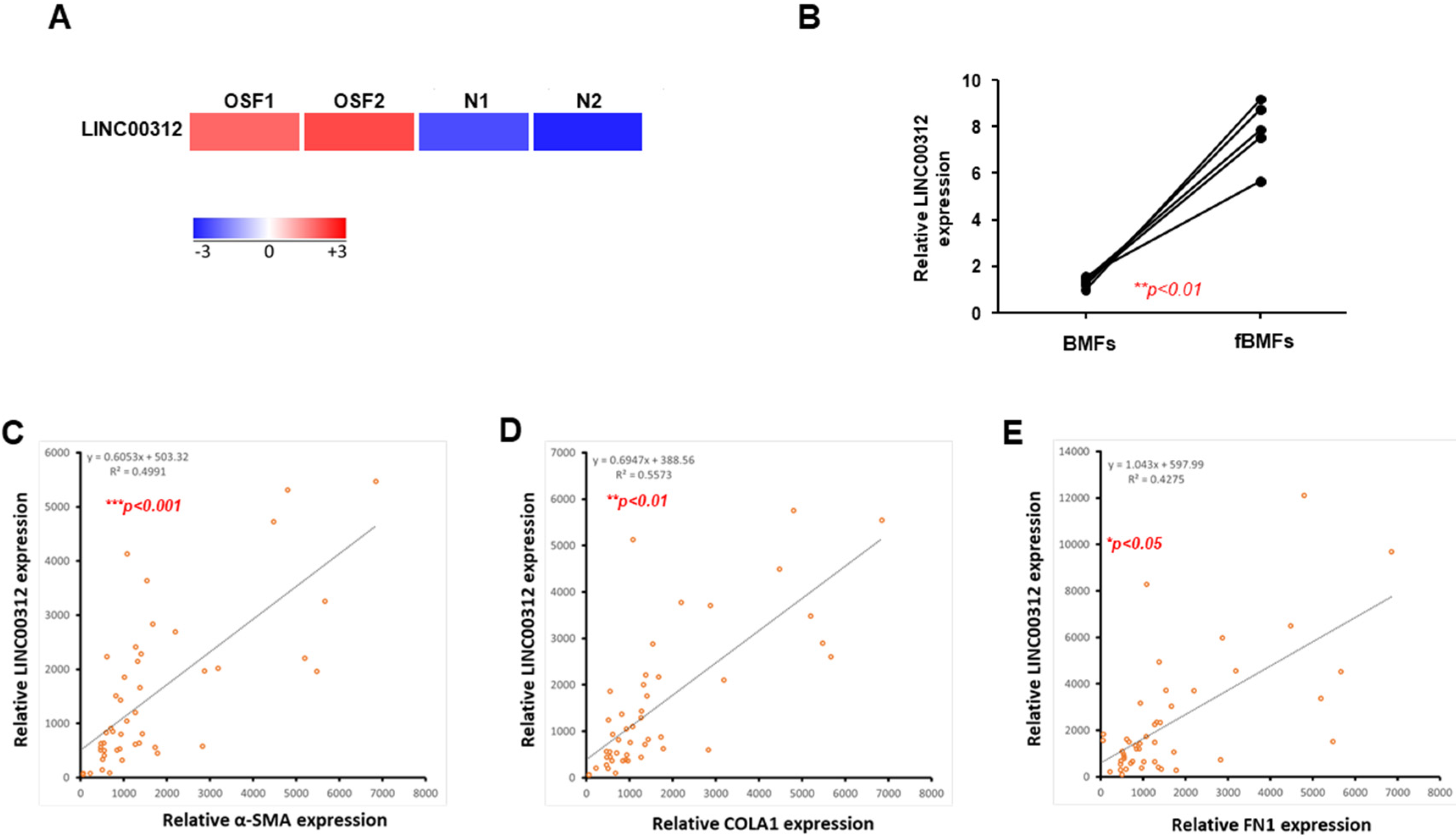
2.1. LINC00312 is Upregulated in OSF Tissues and Associated with Fibrosis Markers
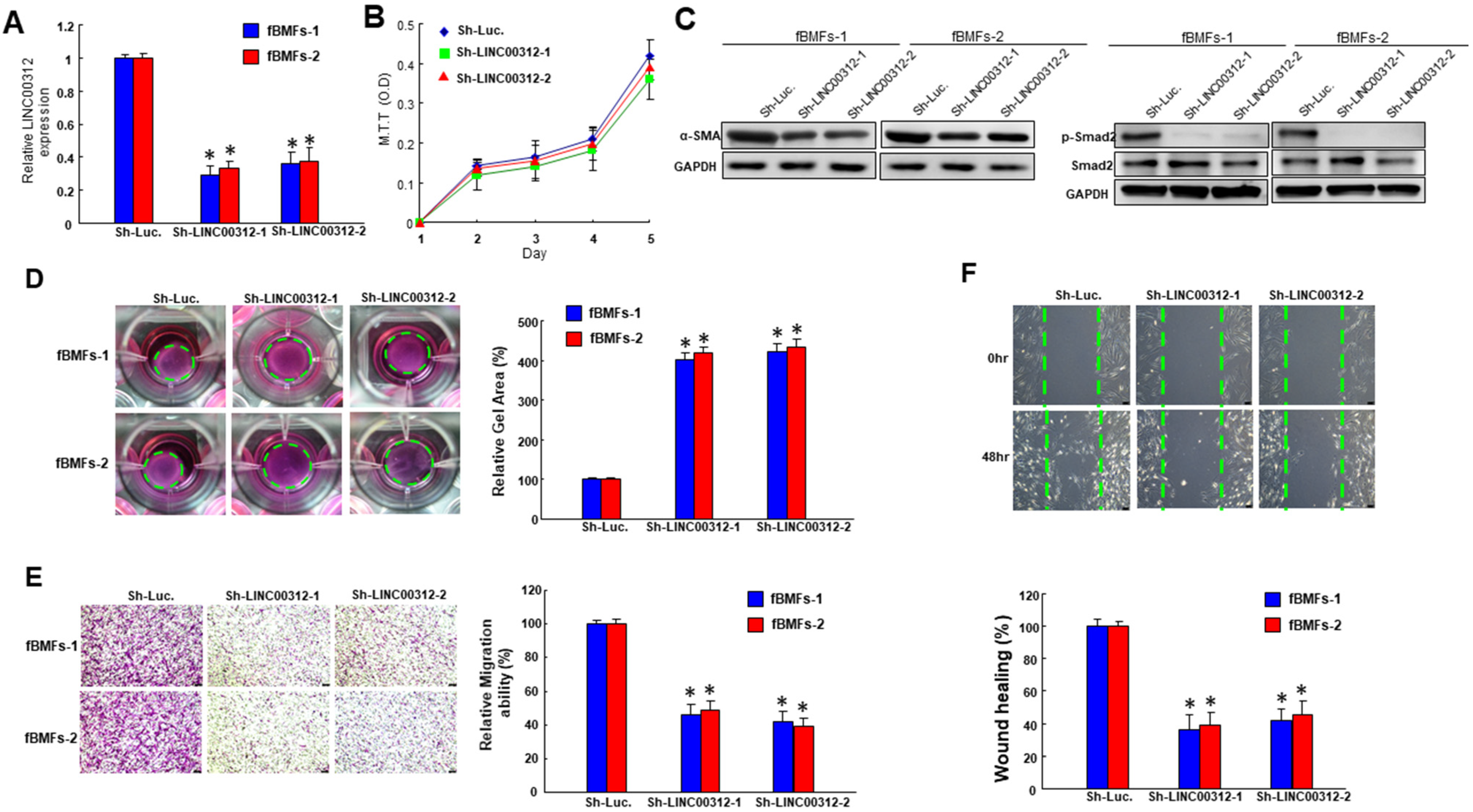
2.2. Suppression of LINC00312 in fBMFs Downregulates the Myofibroblast Features
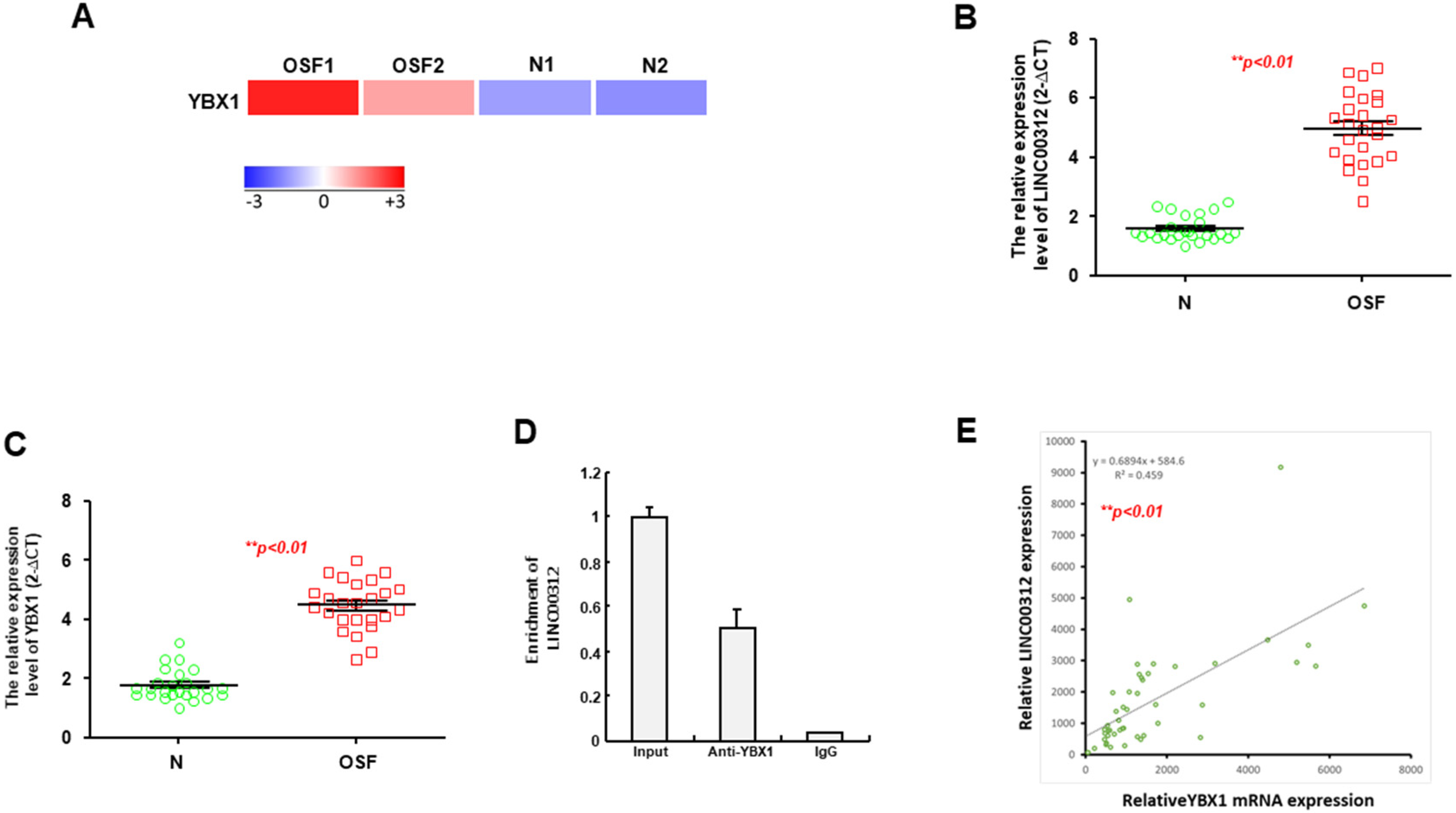
2.3. YBX1 is a Putative Target of LINC00312 and Increased in the OSF Specimens
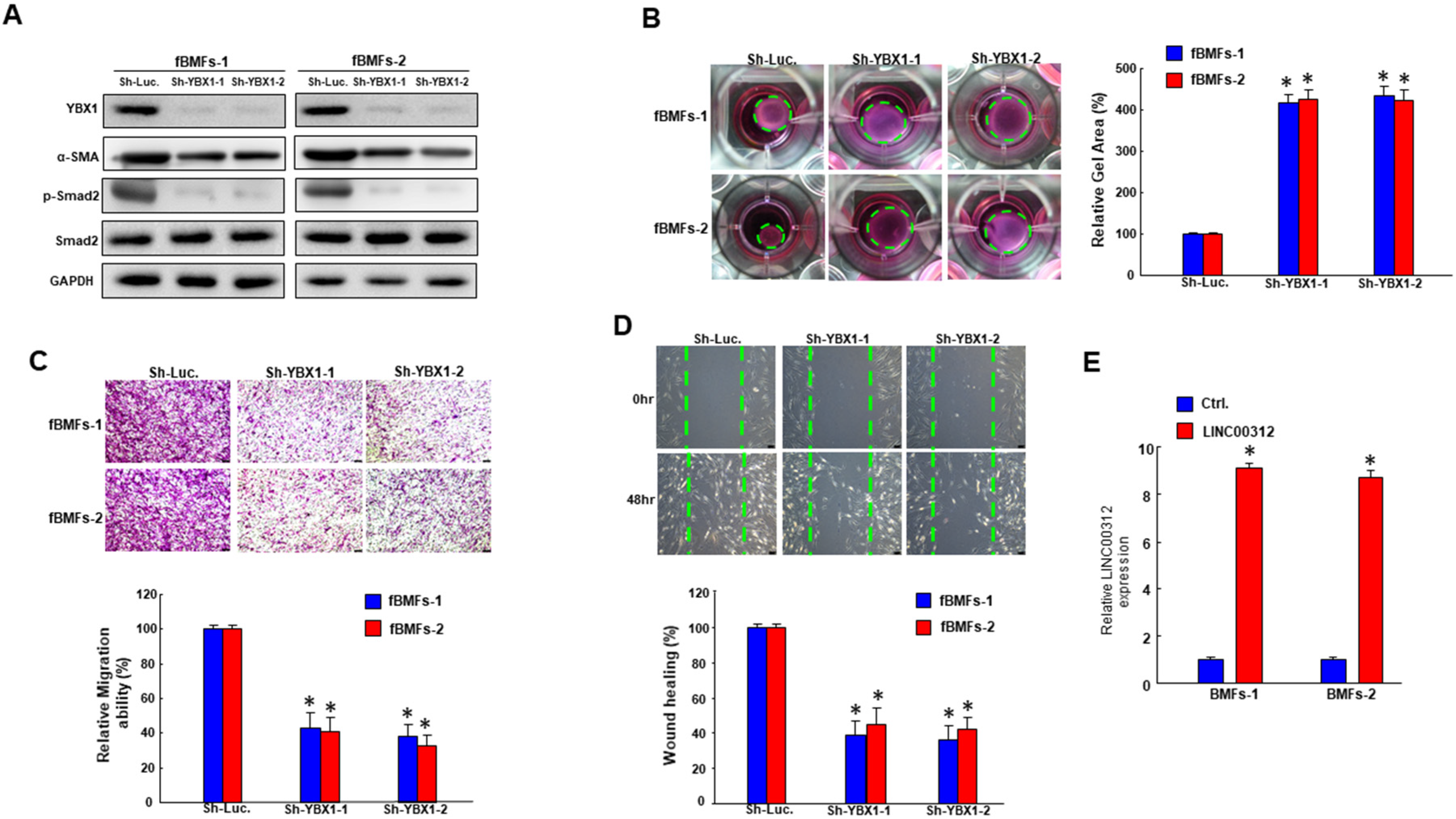
2.4. Inhibition of YBX1 ameliorates the characteristics of fBMFs
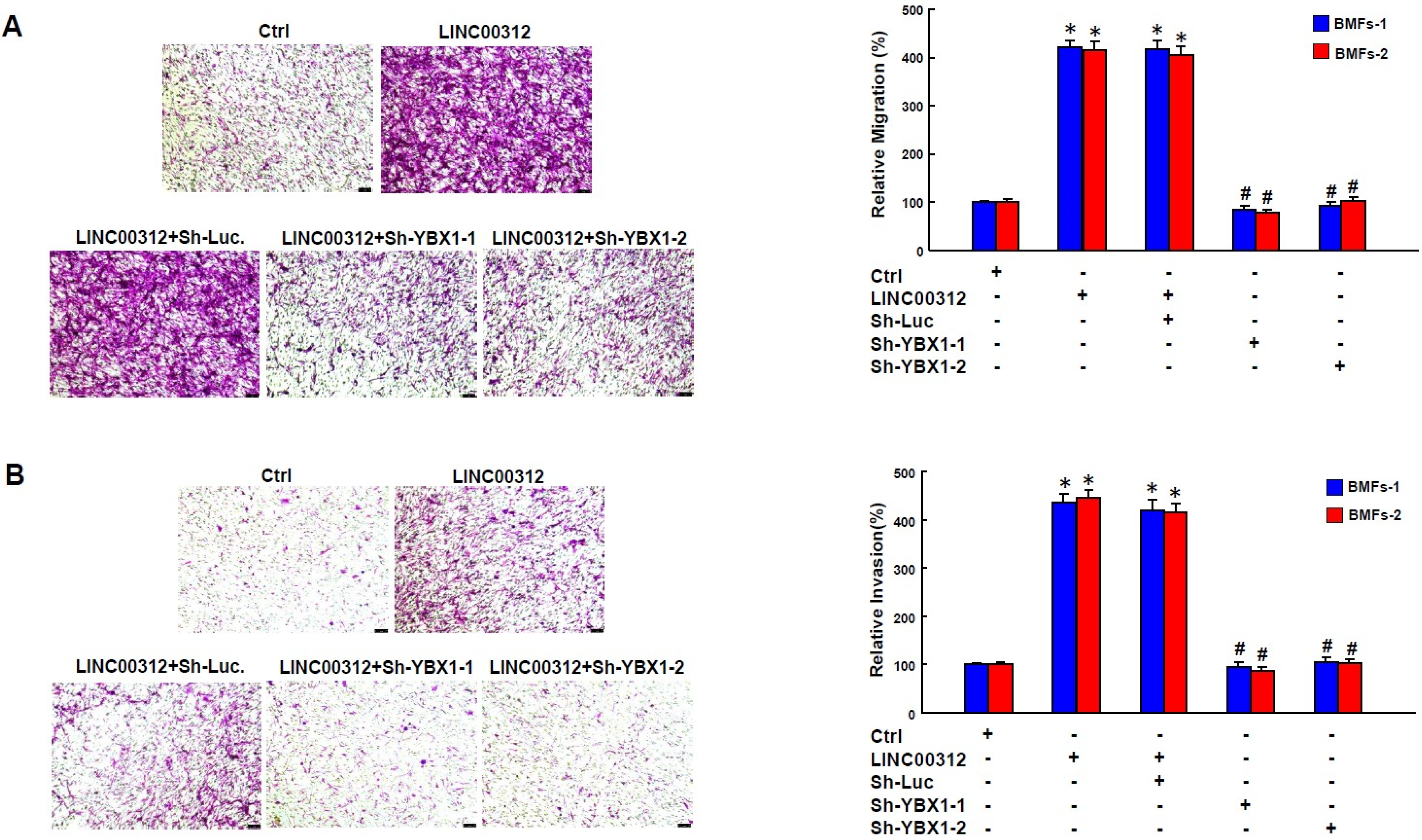
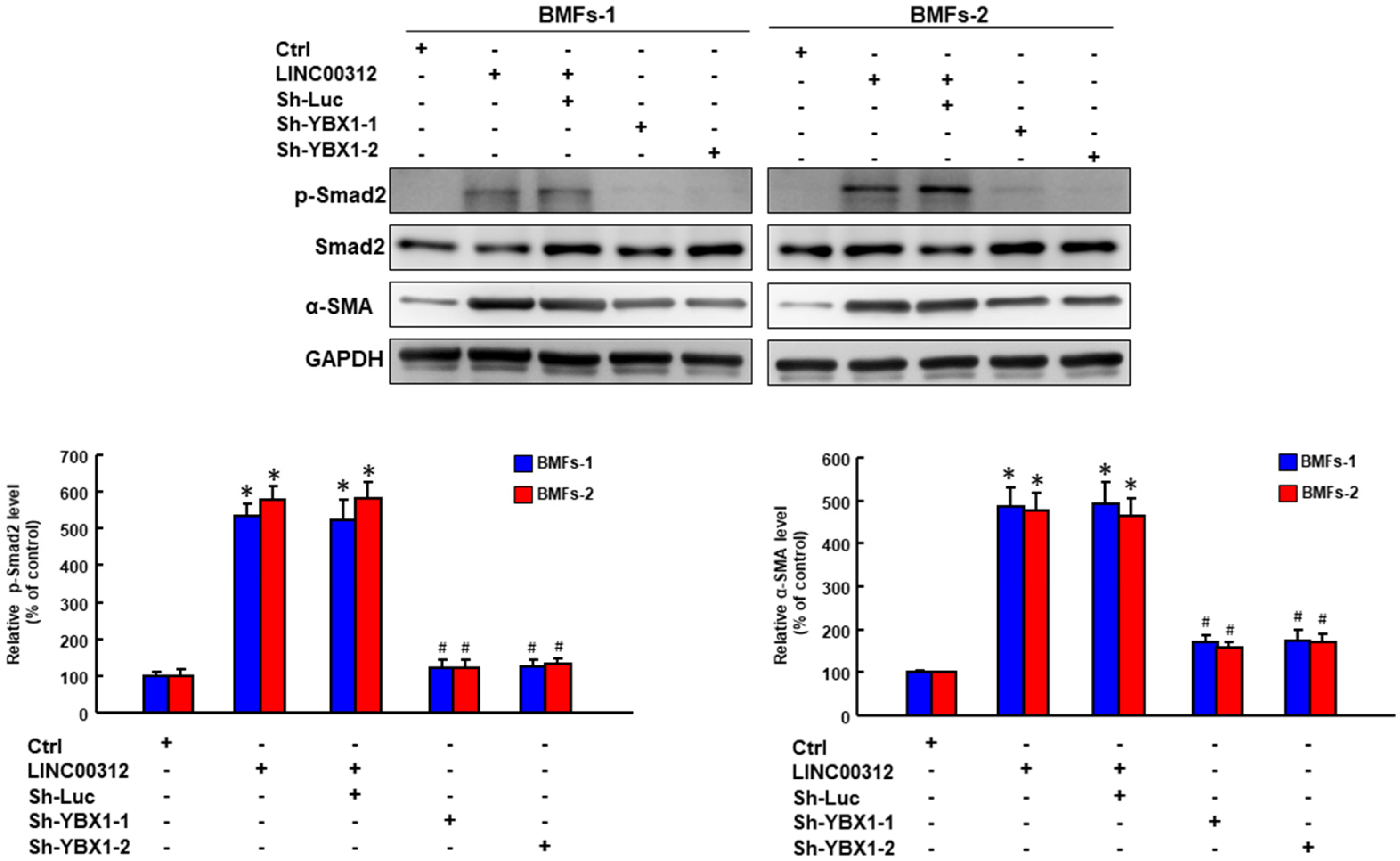
2.5. LINC00312 Enhances the Myofibroblast Activities via YBX1
3. Discussion
4. Materials and Methods
4.1. Cell Culture for Buccal Mucosa Fibroblasts (BMFs) and Fibrotic Buccal Mucosa Fibroblasts (fBMFs)
4.2. Tissues Acquirements
4.3. Quantitative Real-Time PCR (qRT-PCR)
4.4. RNA Sequencing
4.5. Lentiviral-Mediated Silencing of LINC00312 and YBX1
4.6. Collagen Gel Contraction Assay
4.7. Western Blot Analysis
4.8. Transwell Migration and Invasion Assay
4.9. Wound Healing Assay
4.10. Overexpression of LINC00312
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mello, F.W.; Miguel, A.F.P.; Dutra, K.L.; Porporatti, A.L.; Warnakulasuriya, S.; Guerra, E.N.S.; Rivero, E.R.C. Prevalence of oral potentially malignant disorders: A systematic review and meta-analysis. J. Oral Pathol. Med. Off. Publ. Int. Assoc. Oral Pathol. Am. Acad. Oral Pathol. 2018, 47, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Iocca, O.; Sollecito, T.P.; Alawi, F.; Weinstein, G.S.; Newman, J.G.; De Virgilio, A.; Di Maio, P.; Spriano, G.; Pardinas Lopez, S.; Shanti, R.M. Potentially malignant disorders of the oral cavity and oral dysplasia: A systematic review and meta-analysis of malignant transformation rate by subtype. Head Neck 2019. [Google Scholar] [CrossRef]
- Mascitti, M.; Orsini, G.; Tosco, V.; Monterubbianesi, R.; Balercia, A.; Putignano, A.; Procaccini, M.; Santarelli, A. An Overview on Current Non-invasive Diagnostic Devices in Oral Oncology. Front. Physiol. 2018, 9, 1510. [Google Scholar] [CrossRef]
- Vermorken, J.B.; Mesia, R.; Rivera, F.; Remenar, E.; Kawecki, A.; Rottey, S.; Erfan, J.; Zabolotnyy, D.; Kienzer, H.R.; Cupissol, D.; et al. Platinum-based chemotherapy plus cetuximab in head and neck cancer. N. Engl. J. Med. 2008, 359, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Ko, Y.C.; Huang, H.L.; Chao, Y.Y.; Tsai, C.C.; Shieh, T.Y.; Lin, L.M. The precancer risk of betel quid chewing, tobacco use and alcohol consumption in oral leukoplakia and oral submucous fibrosis in southern Taiwan. Br. J. Cancer 2003, 88, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Tsai, C.H.; Lai, Y.L.; Yu, C.C.; Chi, W.Y.; Li, J.J.; Chang, W.W. Arecoline-induced myofibroblast transdifferentiation from human buccal mucosal fibroblasts is mediated by ZEB1. J. Cell. Mol. Med. 2014, 18, 698–708. [Google Scholar] [CrossRef]
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef]
- Fang, C.Y.; Yu, C.C.; Liao, Y.W.; Hsieh, P.L.; Lu, M.Y.; Lin, K.C.; Wu, C.Z.; Tsai, L.L. LncRNA LINC00974 activates TGF-beta/Smad signaling to promote oral fibrogenesis. J. Oral Pathol. Med. Off. Publ. Int. Assoc. Oral Pathol. Am. Acad. Oral Pathol. 2019, 48, 151–158. [Google Scholar] [CrossRef]
- Lin, C.Y.; Liao, Y.W.; Hsieh, P.L.; Lu, M.Y.; Peng, C.Y.; Chu, P.M.; Yang, H.W.; Huang, Y.F.; Yu, C.C.; Yu, C.H. LncRNA GAS5-AS1 inhibits myofibroblasts activities in oral submucous fibrosis. J. Formos. Med Assoc. Taiwan Yi Zhi 2018, 117, 727–733. [Google Scholar] [CrossRef]
- Zhou, S.; Zhu, Y.; He, Z.; Zhang, D.; Guo, F.; Jian, X.; Zhang, C. Long Non-Coding RNA Expression Profile Associated with Malignant Progression of Oral Submucous Fibrosis. J. Oncol. 2019, 2019, 6835176. [Google Scholar] [CrossRef]
- Xie, Y.; Bin, L.; Yang, J.; Li, Z.; Yu, Y.; Zhang, X.; Cao, L.; Li, G. Molecular cloning and characterization of NAG-7: A novel gene downregulated in human nasopharyngeal carcinoma. Chin. Med J. 2001, 114, 530–534. [Google Scholar] [PubMed]
- Huang, C.; Wu, M.; Tang, Y.; Li, X.; Ouyang, J.; Xiao, L.; Li, D.; Li, G. NAG7 promotes human nasopharyngeal carcinoma invasion through inhibition of estrogen receptor alpha and up-regulation of JNK2/AP-1/MMP1 pathways. J. Cell. Physiol. 2009, 221, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Wu, Z.Y.; Wang, G.C.; Liu, K.; Niu, X.B.; Gu, S.; Meng, J.S. LINC00312 inhibits the migration and invasion of bladder cancer cells by targeting miR-197-3p. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2016, 37, 14553–14563. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Huang, W.; Yan, D.Q.; Luo, Q.; Min, X. Overexpression of long intergenic noncoding RNA LINC00312 inhibits the invasion and migration of thyroid cancer cells by down-regulating microRNA-197-3p. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef]
- Min, X.; Liu, K.; Zhu, H.; Zhang, J. Long Noncoding RNA LINC003121 Inhibits Proliferation and Invasion of Thyroid Cancer Cells by Suppression of the Phosphatidylinositol-3-Kinase (PI3K)/Akt Signaling Pathway. Med Sci. Monit. Int. Med J. Exp. Clin. Res. 2018, 24, 4592–4601. [Google Scholar] [CrossRef]
- Li, G.; Wang, C.; Wang, Y.; Xu, B.; Zhang, W. LINC00312 represses proliferation and metastasis of colorectal cancer cells by regulation of miR-21. J. Cell. Mol. Med. 2018, 22, 5565–5572. [Google Scholar] [CrossRef]
- Wu, J.; Zhou, X.; Fan, Y.; Cheng, X.; Lu, B.; Chen, Z. Long non-coding RNA 00312 downregulates cyclin B1 and inhibits hepatocellular carcinoma cell proliferation in vitro and in vivo. Biochem. Biophys. Res. Commun. 2018, 497, 173–180. [Google Scholar] [CrossRef]
- Zhu, Q.; Lv, T.; Wu, Y.; Shi, X.; Liu, H.; Song, Y. Long non-coding RNA 00312 regulated by HOXA5 inhibits tumour proliferation and promotes apoptosis in Non-small cell lung cancer. J. Cell. Mol. Med. 2017, 21, 2184–2198. [Google Scholar] [CrossRef]
- Hinz, B.; Celetta, G.; Tomasek, J.J.; Gabbiani, G.; Chaponnier, C. Alpha-smooth muscle actin expression upregulates fibroblast contractile activity. Molecular biology Cell 2001, 12, 2730–2741. [Google Scholar] [CrossRef]
- Kuo, M.Y.; Chen, H.M.; Hahn, L.J.; Hsieh, C.C.; Chiang, C.P. Collagen biosynthesis in human oral submucous fibrosis fibroblast cultures. J. Dent. Res. 1995, 74, 1783–1788. [Google Scholar] [CrossRef]
- Tsai, C.C.; Ma, R.H.; Shieh, T.Y. Deficiency in collagen and fibronectin phagocytosis by human buccal mucosa fibroblasts in vitro as a possible mechanism for oral submucous fibrosis. J. Oral Pathol. Med. Off. Publ. Int. Assoc. Oral Pathol. Am. Acad. Oral Pathol. 1999, 28, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Wang, J.; Shan, B.; Li, B.; Peng, W.; Dong, Y.; Shi, W.; Zhao, W.; He, D.; Duan, M.; et al. The long noncoding RNA LINC00312 induces lung adenocarcinoma migration and vasculogenic mimicry through directly binding YBX1. Mol. Cancer 2018, 17, 167. [Google Scholar] [CrossRef] [PubMed]
- Higashi, K.; Inagaki, Y.; Suzuki, N.; Mitsui, S.; Mauviel, A.; Kaneko, H.; Nakatsuka, I. Y-box-binding protein YB-1 mediates transcriptional repression of human alpha 2(I) collagen gene expression by interferon-gamma. J. Biol. Chem. 2003, 278, 5156–5162. [Google Scholar] [CrossRef] [PubMed]
- Higashi, K.; Inagaki, Y.; Fujimori, K.; Nakao, A.; Kaneko, H.; Nakatsuka, I. Interferon-gamma interferes with transforming growth factor-beta signaling through direct interaction of YB-1 with Smad3. J. Biol. Chem. 2003, 278, 43470–43479. [Google Scholar] [CrossRef]
- Norman, J.T.; Lindahl, G.E.; Shakib, K.; En-Nia, A.; Yilmaz, E.; Mertens, P.R. The Y-box binding protein YB-1 suppresses collagen alpha 1(I) gene transcription via an evolutionarily conserved regulatory element in the proximal promoter. J. Biol. Chem. 2001, 276, 29880–29890. [Google Scholar] [CrossRef]
- Xiong, P.; Zhang, J.; Xu, D.; Zhu, J.; Li, W.; Liu, J.; Liu, F. Positive feedback loop of YB-1 interacting with Smad2 promotes liver fibrosis. Biochem. Biophys. Res. Commun. 2017, 484, 753–761. [Google Scholar] [CrossRef]
- Higashi, K.; Tomigahara, Y.; Shiraki, H.; Miyata, K.; Mikami, T.; Kimura, T.; Moro, T.; Inagaki, Y.; Kaneko, H. A novel small compound that promotes nuclear translocation of YB-1 ameliorates experimental hepatic fibrosis in mice. J. Biol. Chem. 2011, 286, 4485–4492. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, X.; Cogan, J.G.; Fuerst, M.D.; Polikandriotis, J.A.; Kelm, R.J., Jr.; Strauch, A.R. YB-1 coordinates vascular smooth muscle alpha-actin gene activation by transforming growth factor beta1 and thrombin during differentiation of human pulmonary myofibroblasts. Mol. Biol. Cell 2005, 16, 4931–4940. [Google Scholar] [CrossRef]
- Sumeth Perera, M.W.; Gunasinghe, D.; Perera, P.A.; Ranasinghe, A.; Amaratunga, P.; Warnakulasuriya, S.; Kaluarachchi, K. Development of an in vivo mouse model to study oral submucous fibrosis. J. Oral Pathol. Med. 2007, 36, 273–280. [Google Scholar] [CrossRef]
- Chiang, M.H.; Chen, P.H.; Chen, Y.K.; Chen, C.H.; Ho, M.L.; Wang, Y.H. Characterization of a Novel Dermal Fibrosis Model Induced by Areca Nut Extract that Mimics Oral Submucous Fibrosis. PLoS ONE 2016, 11, e0166454. [Google Scholar] [CrossRef]
- Yu, C.C.; Yu, C.H.; Chang, Y.C. Aberrant SSEA-4 upregulation mediates myofibroblast activity to promote pre-cancerous oral submucous fibrosis. Sci. Rep. 2016, 6, 37004. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, C.-H.; Fang, C.-Y.; Yu, C.-C.; Hsieh, P.-L.; Liao, Y.-W.; Tsai, L.-L.; Chu, P.-M. LINC00312/YBX1 Axis Regulates Myofibroblast Activities in Oral Submucous Fibrosis. Int. J. Mol. Sci. 2020, 21, 2979. https://doi.org/10.3390/ijms21082979
Yu C-H, Fang C-Y, Yu C-C, Hsieh P-L, Liao Y-W, Tsai L-L, Chu P-M. LINC00312/YBX1 Axis Regulates Myofibroblast Activities in Oral Submucous Fibrosis. International Journal of Molecular Sciences. 2020; 21(8):2979. https://doi.org/10.3390/ijms21082979
Chicago/Turabian StyleYu, Chuan-Hang, Chih-Yuan Fang, Cheng-Chia Yu, Pei-Ling Hsieh, Yi-Wen Liao, Lo-Lin Tsai, and Pei-Ming Chu. 2020. "LINC00312/YBX1 Axis Regulates Myofibroblast Activities in Oral Submucous Fibrosis" International Journal of Molecular Sciences 21, no. 8: 2979. https://doi.org/10.3390/ijms21082979
APA StyleYu, C.-H., Fang, C.-Y., Yu, C.-C., Hsieh, P.-L., Liao, Y.-W., Tsai, L.-L., & Chu, P.-M. (2020). LINC00312/YBX1 Axis Regulates Myofibroblast Activities in Oral Submucous Fibrosis. International Journal of Molecular Sciences, 21(8), 2979. https://doi.org/10.3390/ijms21082979