Deeping in the Role of the MAP-Kinases Interacting Kinases (MNKs) in Cancer


 and
and
Abstract
1. Introduction
2. MNKs Substrates
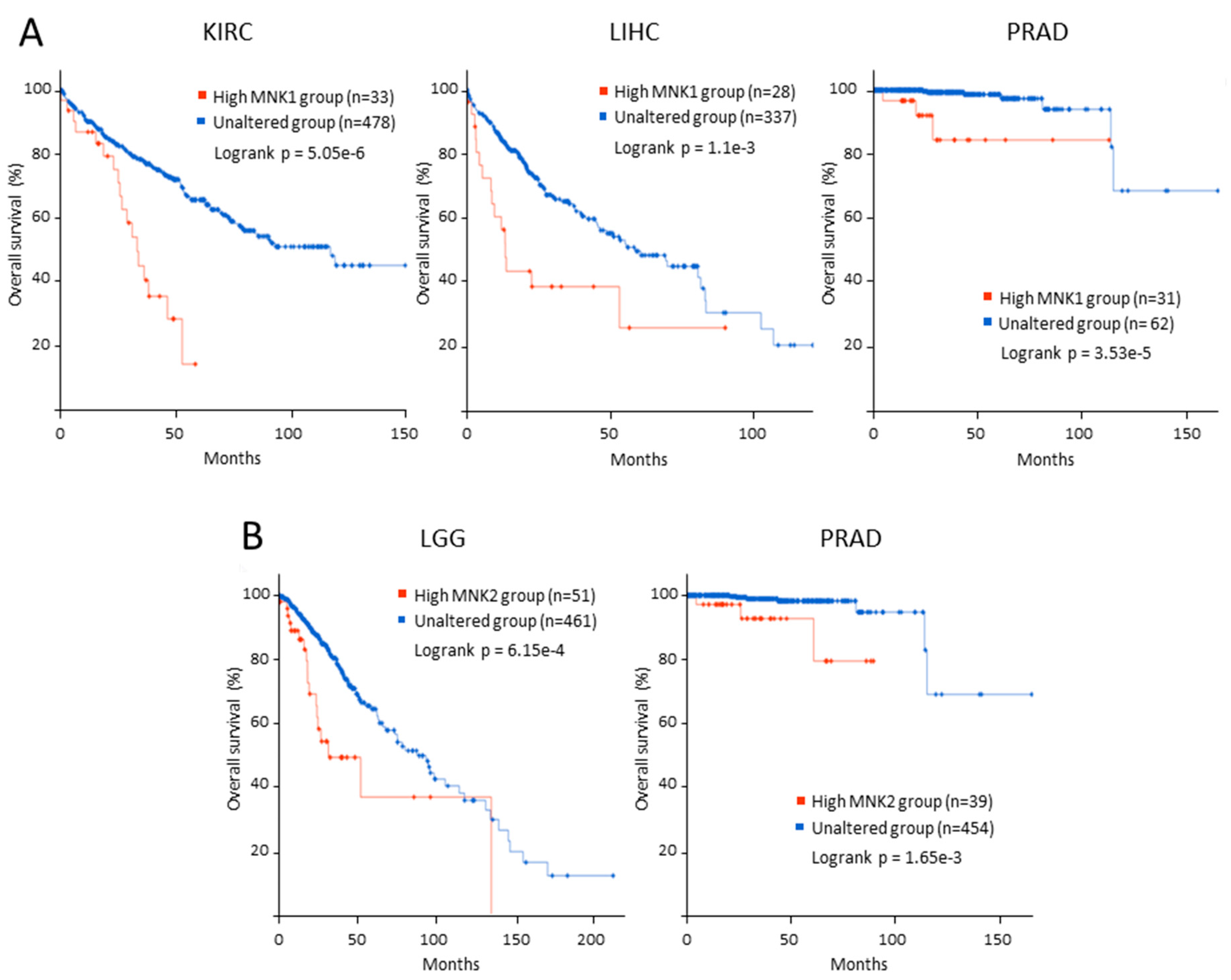
3. MNK and Cancer
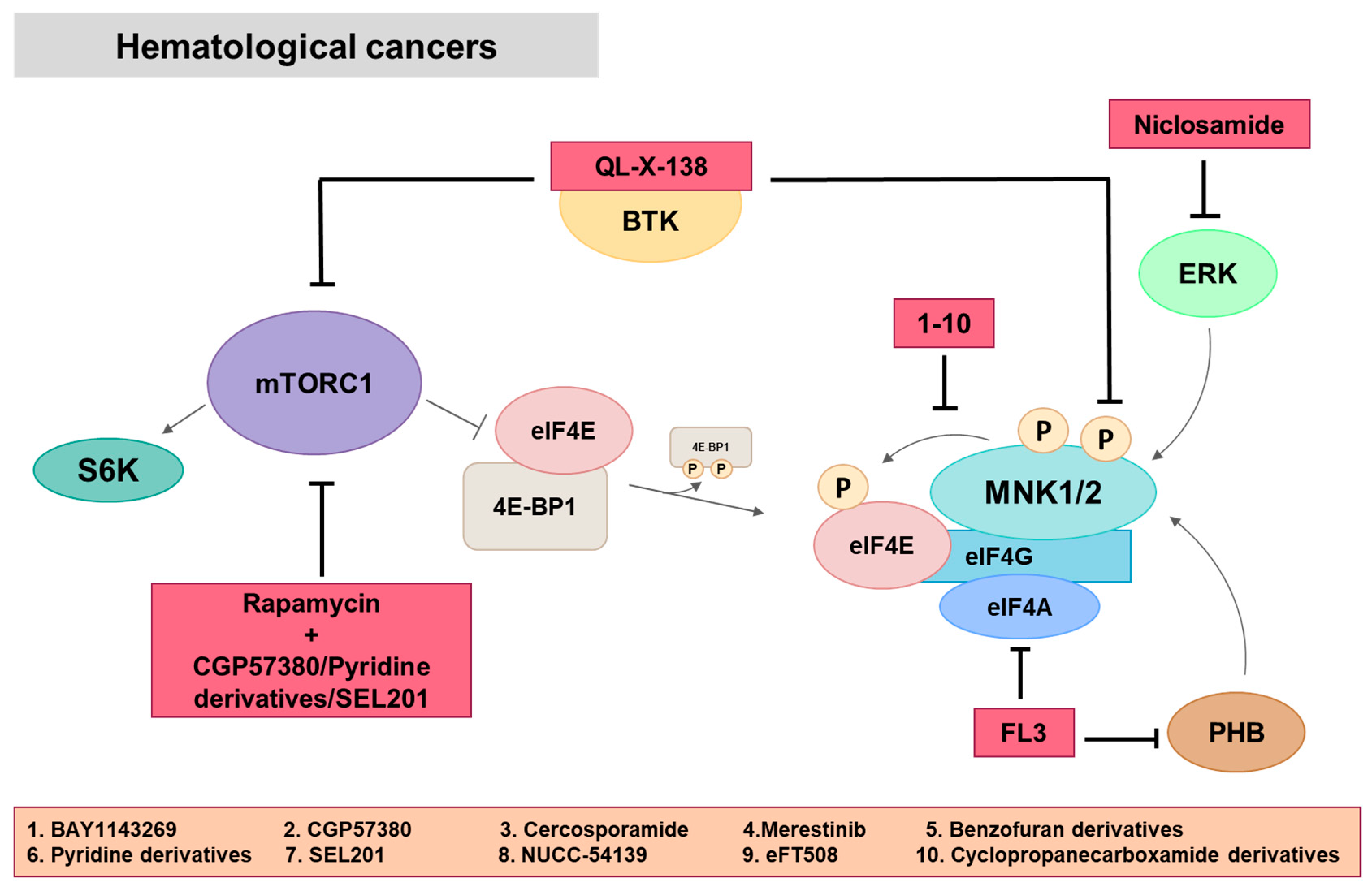
4. MNK in Hematological Cancers
5. MNK in Solid Tumors
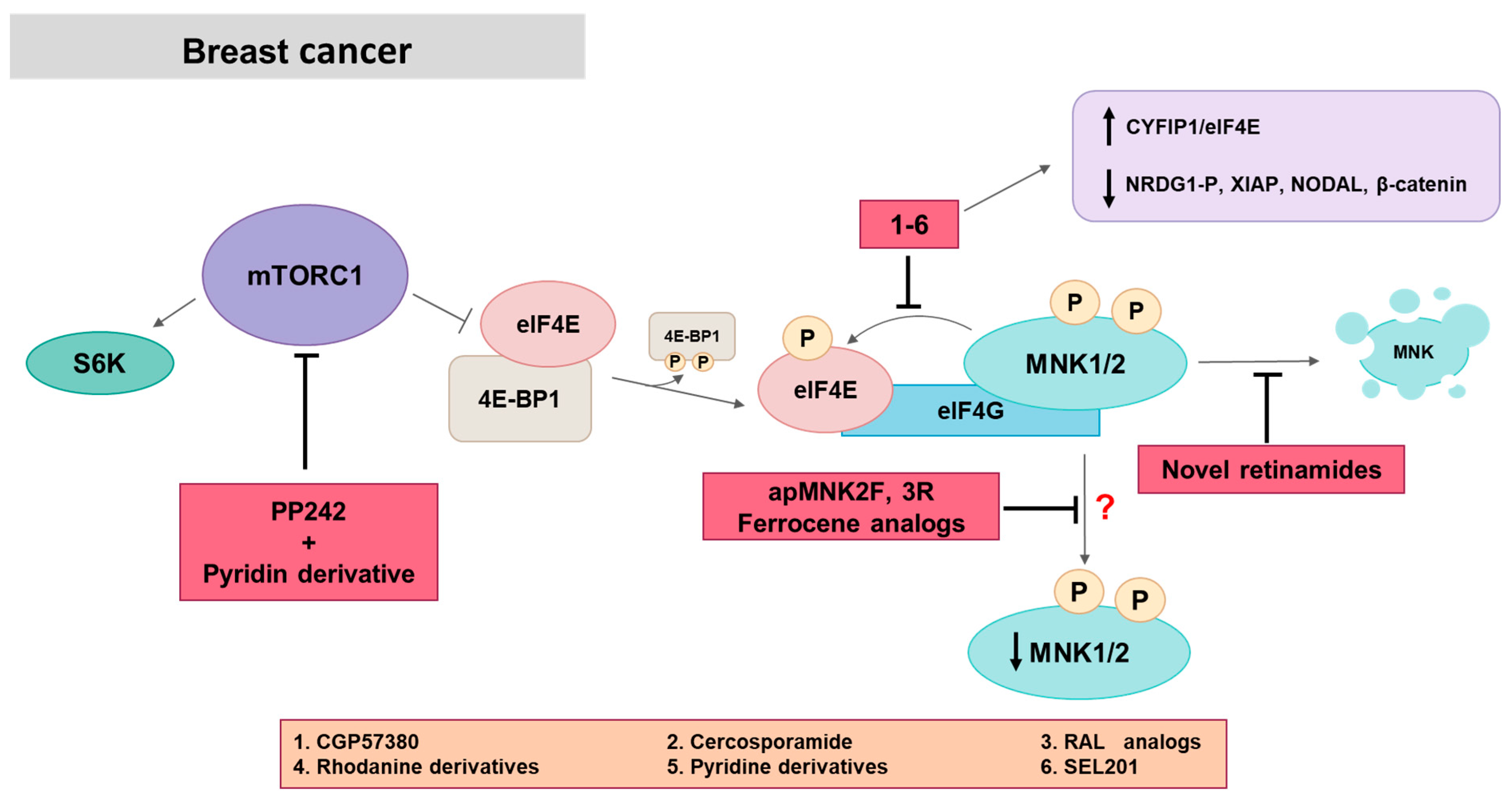
5.1. MNK in Breast Cancer
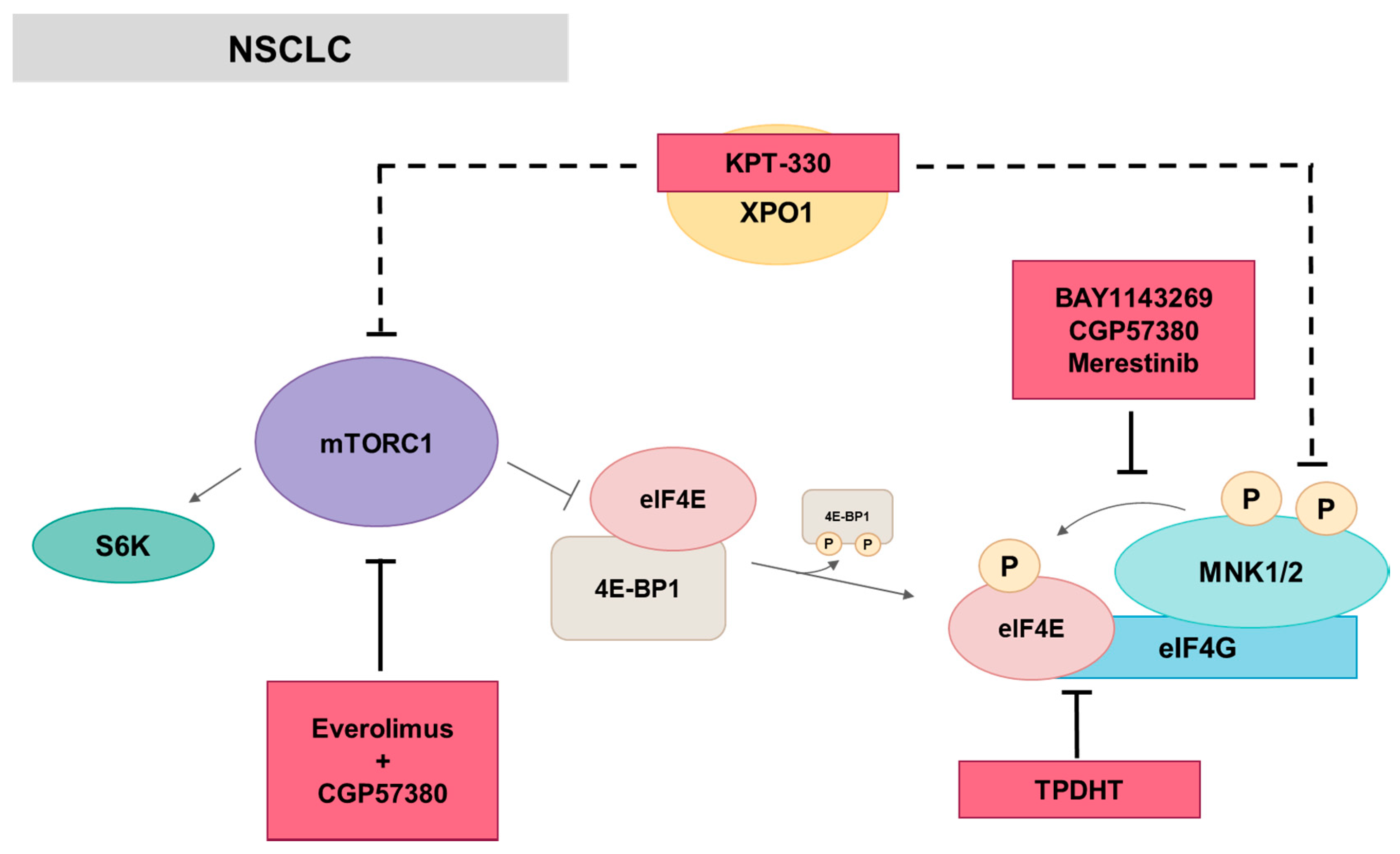
5.2. MNK in Lung Cancer
5.3. MNK in Prostate Cancer
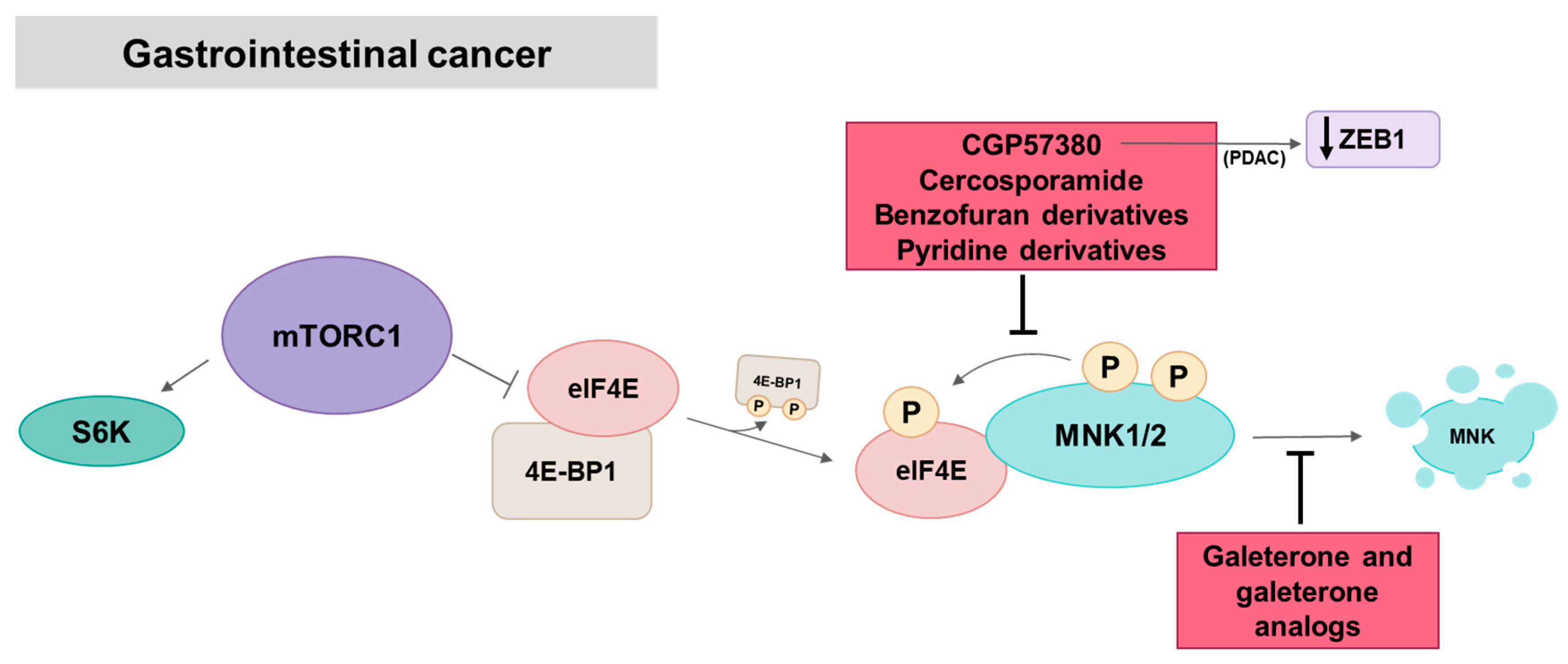
5.4. MNK in Gastrointestinal Cancer
5.5. MNK in Brain and CNS Tumors
5.6. MNK in Other Solid Tumors
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 5’-UTR | untranslated region |
| AKT | also known as protein kinase B |
| ALL | Acute lymphocytic leukemia |
| AML | Acute myeloid leukemia |
| AR | androgen receptor |
| ARE | 3’UTR regions of mRNA rich in residues A and U |
| ATO | arsenic trioxide |
| CML | chronic myeloid leukemia |
| CPLA2 | cytoplasmic phospholipase A2 |
| CRC | colorectal Cancer |
| CRPC | castration-resistant prostate cancer |
| DLBCL | Diffuse large B cell lymphoma |
| eIF | eukaryotic initiation factor |
| EMT | epithelial-mesenchymal transition |
| ERK | extracellular signal-regulated kinase |
| FGF | fibroblast growth factor |
| GBM | glioblastoma multiforme |
| HCC | hepatocellular carcinoma |
| HDM2 | human double minute 2 homolog |
| hnRNP A1 | heterogeneous nuclear ribonucleoprotein A1 |
| IBC | inflammatory breast cancer |
| IL | interleukin |
| IRES | internal ribosomal entry sites |
| MAPK | mitogen-activated protein kinase |
| MCL-1 | myeloid cell leukemia 1 |
| MMP | metalloproteases |
| MNK | MAPK interacting kinase |
| MPNST | malignant peripheral nerve sheath tumor |
| mTOR | mammalian target kinase protein of rapamycin |
| NES | nuclear export signal |
| NLS | nuclear localization signal |
| NPC | nasopharyngeal carcinoma |
| NSCLC | non-small cell lung cancer |
| ODC | ornithine decarboxylase (ODC) |
| PCa | Prostate cancer |
| PCNSL | Primary central nervous system lymphoma |
| PDAC | Pancreatic ductal adenocarcinoma |
| PDX | patient-derived xenograft |
| PI3K | phosphatidylinositol-3 kinase |
| PSF | polypyrimidine tract-binding protein-associated splicing factor |
| PTEN | phosphatase and tensin homolog |
| RCC | renal cell carcinoma |
| SCLC | small-cell lung cancer |
| Spry2 | sprouty 2 |
| TCGA | the cancer Genome Atlas |
| TGFβ | Transforming growth factor β |
| TMZ | Temozolamide |
| TNBC | triple-negative breast cancer tumors |
| TNF | tumor necrosis factor |
| TOP | terminal oligopyrimidine |
| VEGF | vascular endothelial growth factor |
| WHO | World Health Organization |
References
- Waskiewicz, A.J.; Flynn, A.; Proud, C.G.; Cooper, J.A. Mitogen-activated protein kinases activate the serine/threonine kinases Mnk1 and Mnk2. EMBO J. 1997, 16, 1909–1920. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, R.; Hunter, T. MNK1, a new MAP kinase-activated protein kinase, isolated by a novel expression screening method for identifying protein kinase substrates. EMBO J. 1997, 16, 1921–1933. [Google Scholar] [CrossRef] [PubMed]
- Slentz-Kesler, K.; Moore, J.T.; Lombard, M.; Zhang, J.; Hollingsworth, R.; Weiner, M.P. Identification of the human Mnk2 gene (MKNK2) through protein interaction with estrogen receptor beta. Genomics 2000, 69, 63–71. [Google Scholar] [CrossRef] [PubMed]
- O’Loghlen, A.; Gonzalez, V.M.; Pineiro, D.; Perez-Morgado, M.I.; Salinas, M.; Martin, M.E. Identification and molecular characterization of Mnk1b, a splice variant of human MAP kinase-interacting kinase Mnk1. Exp. Cell Res. 2004, 299, 343–355. [Google Scholar] [CrossRef]
- Buxade, M.; Parra, J.L.; Rousseau, S.; Shpiro, N.; Marquez, R.; Morrice, N.; Bain, J.; Espel, E.; Proud, C.G. The Mnks are novel components in the control of TNF alpha biosynthesis and phosphorylate and regulate hnRNP A1. Immunity 2005, 23, 177–189. [Google Scholar] [CrossRef]
- Scheper, G.C.; Parra, J.L.; Wilson, M.; Van Kollenburg, B.; Vertegaal, A.C.; Han, Z.G.; Proud, C.G. The N and C termini of the splice variants of the human mitogen-activated protein kinase-interacting kinase Mnk2 determine activity and localization. Mol. Cell. Biol. 2003, 23, 5692–5705. [Google Scholar] [CrossRef]
- Pyronnet, S.; Imataka, H.; Gingras, A.C.; Fukunaga, R.; Hunter, T.; Sonenberg, N. Human eukaryotic translation initiation factor 4G (eIF4G) recruits mnk1 to phosphorylate eIF4E. EMBO J. 1999, 18, 270–279. [Google Scholar] [CrossRef]
- Scheper, G.C.; Morrice, N.A.; Kleijn, M.; Proud, C.G. The mitogen-activated protein kinase signal-integrating kinase Mnk2 is a eukaryotic initiation factor 4E kinase with high levels of basal activity in mammalian cells. Mol. Cell. Biol. 2001, 21, 743–754. [Google Scholar] [CrossRef]
- Goto, S.; Yao, Z.; Proud, C.G. The C-terminal domain of Mnk1a plays a dual role in tightly regulating its activity. Biochem. J. 2009, 423, 279–290. [Google Scholar] [CrossRef]
- Jauch, R.; Jakel, S.; Netter, C.; Schreiter, K.; Aicher, B.; Jackle, H.; Wahl, M.C. Crystal structures of the Mnk2 kinase domain reveal an inhibitory conformation and a zinc binding site. Structure 2005, 13, 1559–1568. [Google Scholar] [CrossRef]
- Jauch, R.; Cho, M.K.; Jakel, S.; Netter, C.; Schreiter, K.; Aicher, B.; Zweckstetter, M.; Jackle, H.; Wahl, M.C. Mitogen-activated protein kinases interacting kinases are autoinhibited by a reprogrammed activation segment. EMBO J. 2006, 25, 4020–4032. [Google Scholar] [CrossRef] [PubMed]
- Waskiewicz, A.J.; Johnson, J.C.; Penn, B.; Mahalingam, M.; Kimball, S.R.; Cooper, J.A. Phosphorylation of the cap-binding protein eukaryotic translation initiation factor 4E by protein kinase Mnk1 in vivo. Mol. Cell. Biol. 1999, 19, 1871–1880. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Watanabe-Fukunaga, R.; Fukuyama, H.; Nagata, S.; Fukunaga, R. Mnk2 and Mnk1 are essential for constitutive and inducible phosphorylation of eukaryotic initiation factor 4E but not for cell growth or development. Mol. Cell. Biol. 2004, 24, 6539–6549. [Google Scholar] [CrossRef] [PubMed]
- Karaki, S.; Andrieu, C.; Ziouziou, H.; Rocchi, P. The Eukaryotic Translation Initiation Factor 4E (eIF4E) as a Therapeutic Target for Cancer. Adv. Protein Chem. Struct. Biol. 2015, 101, 1–26. [Google Scholar] [PubMed]
- Siddiqui, N.; Sonenberg, N. Signalling to eIF4E in cancer. Biochem. Soc. Trans. 2015, 43, 763–772. [Google Scholar] [CrossRef] [PubMed]
- O’Loghlen, A.; Gonzalez, V.M.; Salinas, M.; Martin, M.E. Suppression of human Mnk1 by small interfering RNA increases the eukaryotic initiation factor 4F activity in HEK293T cells. FEBS Lett. 2004, 578, 31–35. [Google Scholar] [CrossRef][Green Version]
- Scheper, G.C.; van Kollenburg, B.; Hu, J.; Luo, Y.; Goss, D.J.; Proud, C.G. Phosphorylation of eukaryotic initiation factor 4E markedly reduces its affinity for capped mRNA. J. Biol. Chem. 2002, 277, 3303–3309. [Google Scholar] [CrossRef]
- Slepenkov, S.V.; Darzynkiewicz, E.; Rhoads, R.E. Stopped-flow kinetic analysis of eIF4E and phosphorylated eIF4E binding to cap analogs and capped oligoribonucleotides: Evidence for a one-step binding mechanism. J. Biol. Chem. 2006, 281, 14927–14938. [Google Scholar] [CrossRef]
- Rousseau, D.; Kaspar, R.; Rosenwald, I.; Gehrke, L.; Sonenberg, N. Translation initiation of ornithine decarboxylase and nucleocytoplasmic transport of cyclin D1 mRNA are increased in cells overexpressing eukaryotic initiation factor 4E. Proc. Natl. Acad. Sci. USA 1996, 93, 1065–1070. [Google Scholar] [CrossRef]
- Topisirovic, I.; Ruiz-Gutierrez, M.; Borden, K.L. Phosphorylation of the eukaryotic translation initiation factor eIF4E contributes to its transformation and mRNA transport activities. Cancer Res. 2004, 64, 8639–8642. [Google Scholar] [CrossRef]
- Kjellerup, R.B.; Kragballe, K.; Iversen, L.; Johansen, C. Pro-inflammatory cytokine release in keratinocytes is mediated through the MAPK signal-integrating kinases. Exp. Dermatol. 2008, 17, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Rowlett, R.M.; Chrestensen, C.A.; Nyce, M.; Harp, M.G.; Pelo, J.W.; Cominelli, F.; Ernst, P.B.; Pizarro, T.T.; Sturgill, T.W.; Worthington, M.T. MNK kinases regulate multiple TLR pathways and innate proinflammatory cytokines in macrophages. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G452–G459. [Google Scholar] [CrossRef] [PubMed]
- Buxade, M.; Parra-Palau, J.L.; Proud, C.G. The Mnks: MAP kinase-interacting kinases (MAP kinase signal-integrating kinases). Front. Biosci. A. J. Virtual Libr. 2008, 13, 5359–5373. [Google Scholar] [CrossRef] [PubMed]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. Mmbr. 2011, 75, 50–83. [Google Scholar] [CrossRef]
- Shav-Tal, Y.; Zipori, D. PSF and p54 (nrb)/NonO--multi-functional nuclear proteins. FEBS Lett. 2002, 531, 109–114. [Google Scholar] [CrossRef]
- Cobbold, L.C.; Spriggs, K.A.; Haines, S.J.; Dobbyn, H.C.; Hayes, C.; de Moor, C.H.; Lilley, K.S.; Bushell, M.; Willis, A.E. Identification of internal ribosome entry segment (IRES)-trans-acting factors for the Myc family of IRESs. Mol. Cell. Biol. 2008, 28, 40–49. [Google Scholar] [CrossRef]
- Buxade, M.; Morrice, N.; Krebs, D.L.; Proud, C.G. The PSF.p54nrb complex is a novel Mnk substrate that binds the mRNA for tumor necrosis factor alpha. J. Biol. Chem. 2008, 283, 57–65. [Google Scholar] [CrossRef]
- Pashenkov, M.V.; Balyasova, L.S.; Dagil, Y.A.; Pinegin, B.V. The Role of the p38-MNK-eIF4E Signaling Axis in TNF Production Downstream of the NOD1 Receptor. J. Immunol. 2017, 198, 1638–1648. [Google Scholar] [CrossRef]
- Hefner, Y.; Borsch-Haubold, A.G.; Murakami, M.; Wilde, J.I.; Pasquet, S.; Schieltz, D.; Ghomashchi, F.; Yates, J.R., 3rd; Armstrong, C.G.; Paterson, A.; et al. Serine 727 phosphorylation and activation of cytosolic phospholipase A2 by MNK1-related protein kinases. J. Biol. Chem. 2000, 275, 37542–37551. [Google Scholar] [CrossRef]
- DaSilva, J.; Xu, L.; Kim, H.J.; Miller, W.T.; Bar-Sagi, D. Regulation of sprouty stability by Mnk1-dependent phosphorylation. Mol. Cell. Biol. 2006, 26, 1898–1907. [Google Scholar] [CrossRef]
- Lazaris-Karatzas, A.; Montine, K.S.; Sonenberg, N. Malignant transformation by a eukaryotic initiation factor subunit that binds to mRNA 5’ cap. Nature 1990, 345, 544–547. [Google Scholar] [CrossRef] [PubMed]
- De Benedetti, A.; Graff, J.R. eIF-4E expression and its role in malignancies and metastases. Oncogene 2004, 23, 3189–3199. [Google Scholar] [CrossRef]
- Bilanges, B.; Stokoe, D. Mechanisms of translational deregulation in human tumors and therapeutic intervention strategies. Oncogene 2007, 26, 5973–5990. [Google Scholar] [CrossRef] [PubMed]
- Culjkovic, B.; Topisirovic, I.; Borden, K.L. Controlling gene expression through RNA regulons: the role of the eukaryotic translation initiation factor eIF4E. Cell Cycle 2007, 6, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.; Blaydes, J.P. MNK1 and EIF4E are downstream effectors of MEKs in the regulation of the nuclear export of HDM2 mRNA. Oncogene 2008, 27, 1645–1649. [Google Scholar] [CrossRef] [PubMed]
- Wendel, H.G.; Silva, R.L.; Malina, A.; Mills, J.R.; Zhu, H.; Ueda, T.; Watanabe-Fukunaga, R.; Fukunaga, R.; Teruya-Feldstein, J.; Pelletier, J.; et al. Dissecting eIF4E action in tumorigenesis. Genes Dev. 2007, 21, 3232–3237. [Google Scholar] [CrossRef]
- Fan, S.; Ramalingam, S.S.; Kauh, J.; Xu, Z.; Khuri, F.R.; Sun, S.Y. Phosphorylated eukaryotic translation initiation factor 4 (eIF4E) is elevated in human cancer tissues. Cancer Biol. Ther. 2009, 8, 1463–1469. [Google Scholar] [CrossRef]
- Adesso, L.; Calabretta, S.; Barbagallo, F.; Capurso, G.; Pilozzi, E.; Geremia, R.; Delle Fave, G.; Sette, C. Gemcitabine triggers a pro-survival response in pancreatic cancer cells through activation of the MNK2/eIF4E pathway. Oncogene 2013, 32, 2848–2857. [Google Scholar] [CrossRef] [PubMed]
- Chrestensen, C.A.; Shuman, J.K.; Eschenroeder, A.; Worthington, M.; Gram, H.; Sturgill, T.W. MNK1 and MNK2 regulation in HER2-overexpressing breast cancer lines. J. Biol. Chem. 2007, 282, 4243–4252. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Sasaki, M.; Elia, A.J.; Chio, I.I.C.; Hamada, K.; Fukunaga, R.; Mak, T.W. Combined deficiency for MAP kinase-interacting kinase 1 and 2 (Mnk1 and Mnk2) delays tumor development. Proc. Natl. Acad. Sci. USA 2010, 107, 13984–13990. [Google Scholar] [CrossRef] [PubMed]
- Raught, B.; Gingras, A.-C. Signaling to Translation Initiation. In Translational Control in Biology and Medicine; Mathews, M.B., Sonenberg, N., Hershey, J.W.B., Eds.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2007; pp. 369–400. [Google Scholar]
- Lineham, E.; Spencer, J.; Morley, S.J. Dual abrogation of MNK and mTOR: A novel therapeutic approach for the treatment of aggressive cancers. Future Med. Chem. 2017, 9, 1539–1555. [Google Scholar] [CrossRef]
- Chappell, W.H.; Steelman, L.S.; Long, J.M.; Kempf, R.C.; Abrams, S.L.; Franklin, R.A.; Basecke, J.; Stivala, F.; Donia, M.; Fagone, P.; et al. Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR inhibitors: rationale and importance to inhibiting these pathways in human health. Oncotarget 2011, 2, 135–164. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Steelman, L.S.; Kempf, C.R.; Chappell, W.H.; Abrams, S.L.; Stivala, F.; Malaponte, G.; Nicoletti, F.; Libra, M.; Basecke, J.; et al. Therapeutic resistance resulting from mutations in Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR signaling pathways. J. Cell. Physiol. 2011, 226, 2762–2781. [Google Scholar] [CrossRef] [PubMed]
- Shahbazian, D.; Roux, P.P.; Mieulet, V.; Cohen, M.S.; Raught, B.; Taunton, J.; Hershey, J.W.; Blenis, J.; Pende, M.; Sonenberg, N. The mTOR/PI3K and MAPK pathways converge on eIF4B to control its phosphorylation and activity. EMBO J. 2006, 25, 2781–2791. [Google Scholar] [CrossRef] [PubMed]
- Grzmil, M.; Morin, P., Jr.; Lino, M.M.; Merlo, A.; Frank, S.; Wang, Y.; Moncayo, G.; Hemmings, B.A. MAP kinase-interacting kinase 1 regulates SMAD2-dependent TGF-beta signaling pathway in human glioblastoma. Cancer Res. 2011, 71, 2392–2402. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, A.; Loiarro, M.; Bielli, P.; Busa, R.; Paronetto, M.P.; Loreni, F.; Geremia, R.; Sette, C. Phosphorylation of eIF4E by MNKs supports protein synthesis, cell cycle progression and proliferation in prostate cancer cells. Carcinogenesis 2008, 29, 2279–2288. [Google Scholar] [CrossRef]
- Sun, S.Y.; Rosenberg, L.M.; Wang, X.; Zhou, Z.; Yue, P.; Fu, H.; Khuri, F.R. Activation of Akt and eIF4E survival pathways by rapamycin-mediated mammalian target of rapamycin inhibition. Cancer Res. 2005, 65, 7052–7058. [Google Scholar] [CrossRef]
- Wang, X.; Yue, P.; Chan, C.B.; Ye, K.; Ueda, T.; Watanabe-Fukunaga, R.; Fukunaga, R.; Fu, H.; Khuri, F.R.; Sun, S.Y. Inhibition of mammalian target of rapamycin induces phosphatidylinositol 3-kinase-dependent and Mnk-mediated eukaryotic translation initiation factor 4E phosphorylation. Mol. Cell. Biol. 2007, 27, 7405–7413. [Google Scholar] [CrossRef]
- Shi, Y.; Frost, P.; Hoang, B.; Yang, Y.; Fukunaga, R.; Gera, J.; Lichtenstein, A. MNK kinases facilitate c-myc IRES activity in rapamycin-treated multiple myeloma cells. Oncogene 2013, 32, 190–197. [Google Scholar] [CrossRef]
- Shi, Y.; Yang, Y.; Hoang, B.; Bardeleben, C.; Holmes, B.; Gera, J.; Lichtenstein, A. Therapeutic potential of targeting IRES-dependent c-myc translation in multiple myeloma cells during ER stress. Oncogene 2016, 35, 1015–1024. [Google Scholar] [CrossRef]
- Brown, M.C.; Bryant, J.D.; Dobrikova, E.Y.; Shveygert, M.; Bradrick, S.S.; Chandramohan, V.; Bigner, D.D.; Gromeier, M. Induction of viral, 7-methyl-guanosine cap-independent translation and oncolysis by mitogen-activated protein kinase-interacting kinase-mediated effects on the serine/arginine-rich protein kinase. J. Virol. 2014, 88, 13135–13148. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.C.; Dobrikov, M.I.; Gromeier, M. Mitogen-activated protein kinase-interacting kinase regulates mTOR/AKT signaling and controls the serine/arginine-rich protein kinase-responsive type 1 internal ribosome entry site-mediated translation and viral oncolysis. J. Virol. 2014, 88, 13149–13160. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.C.; Gromeier, M. MNK Controls mTORC1: Substrate Association through Regulation of TELO2 Binding with mTORC1. Cell Rep. 2017, 18, 1444–1457. [Google Scholar] [CrossRef] [PubMed]
- Wheater, M.J.; Johnson, P.W.; Blaydes, J.P. The role of MNK proteins and eIF4E phosphorylation in breast cancer cell proliferation and survival. Cancer Biol. Ther. 2010, 10, 728–735. [Google Scholar] [CrossRef]
- Guo, Z.; Peng, G.; Li, E.; Xi, S.; Zhang, Y.; Li, Y.; Lin, X.; Li, G.; Wu, Q.; He, J. MAP kinase-interacting serine/threonine kinase 2 promotes proliferation, metastasis, and predicts poor prognosis in non-small cell lung cancer. Sci. Rep. 2017, 7, 10612. [Google Scholar] [CrossRef]
- Hou, S.; Du, P.; Wang, P.; Wang, C.; Liu, P.; Liu, H. Significance of MNK1 in prognostic prediction and chemotherapy development of epithelial ovarian cancer. Clin. Transl. Oncol. 2017, 19, 1107–1116. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Zhang, Q.; Zhuang, H.; Chen, B. MAP Kinase-Interacting Kinase 1 Promotes Proliferation and Invasion of Hepatocellular Carcinoma and Is an Unfavorable Prognostic Biomarker. Med. Sci. Monit. 2018, 24, 1759–1767. [Google Scholar] [CrossRef]
- Pinto-Diez, C.; Garcia-Recio, E.M.; Perez-Morgado, M.I.; Garcia-Hernandez, M.; Sanz-Criado, L.; Sacristan, S.; Toledo-Lobo, M.V.; Perez-Mies, B.; Esteban-Rodriguez, I.; Pascual, A.; et al. Increased expression of MNK1b, the spliced isoform of MNK1, predicts poor prognosis and is associated with triple-negative breast cancer. Oncotarget 2018, 9, 13501–13516. [Google Scholar] [CrossRef]
- Maimon, A.; Mogilevsky, M.; Shilo, A.; Golan-Gerstl, R.; Obiedat, A.; Ben-Hur, V.; Lebenthal-Loinger, I.; Stein, I.; Reich, R.; Beenstock, J.; et al. Mnk2 alternative splicing modulates the p38-MAPK pathway and impacts Ras-induced transformation. Cell Rep. 2014, 7, 501–513. [Google Scholar] [CrossRef]
- Santag, S.; Siegel, F.; Wengner, A.M.; Lange, C.; Bomer, U.; Eis, K.; Puhler, F.; Lienau, P.; Bergemann, L.; Michels, M.; et al. BAY 1143269, a novel MNK1 inhibitor, targets oncogenic protein expression and shows potent anti-tumor activity. Cancer Lett. 2017, 390, 21–29. [Google Scholar] [CrossRef]
- Kosciuczuk, E.M.; Saleiro, D.; Kroczynska, B.; Beauchamp, E.M.; Eckerdt, F.; Blyth, G.T.; Abedin, S.M.; Giles, F.J.; Altman, J.K.; Platanias, L.C. Merestinib blocks Mnk kinase activity in acute myeloid leukemia progenitors and exhibits antileukemic effects in vitro and in vivo. Blood 2016, 128, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-B.; Yang, C.-M.; Han, Q.-M.; Ye, X.-J.; Lei, W.; Qian, W.-B. MNK1 inhibitor CGP57380 overcomes mTOR inhibitor-induced activation of eIF4E: the mechanism of synergic killing of human T-ALL cells. Acta Pharmacol. Sin. 2018, 39, 1894–1901. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Wang, X.; Proud, C.G. Oncogenic MNK signalling regulates the metastasis suppressor NDRG1. Oncotarget 2017, 8, 46121–46135. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Sun, Y.; Qu, M.; Wan, H.; Cai, F.; Zhang, P. Inhibiting the MNK-eIF4E-beta-catenin axis increases the responsiveness of aggressive breast cancer cells to chemotherapy. Oncotarget 2017, 8, 2906–2915. [Google Scholar] [PubMed]
- Wen, Q.; Wang, W.; Luo, J.; Chu, S.; Chen, L.; Xu, L.; Zang, H.; Alnemah, M.M.; Ma, J.; Fan, S. CGP57380 enhances efficacy of RAD001 in non-small cell lung cancer through abrogating mTOR inhibition-induced phosphorylation of eIF4E and activating mitochondrial apoptotic pathway. Oncotarget 2016, 7, 27787–27801. [Google Scholar] [CrossRef]
- Knauf, U.; Tschopp, C.; Gram, H. Negative regulation of protein translation by mitogen-activated protein kinase-interacting kinases 1 and 2. Mol. Cell. Biol. 2001, 21, 5500–5511. [Google Scholar] [CrossRef]
- Wen, Q.; Zhan, Y.; Zheng, H.; Zang, H.; Luo, J.; Zhang, Y.; Wang, W.; Feng, J.; Lu, J.; Chen, L.; et al. Elevated expression of mcl-1 inhibits apoptosis and predicts poor prognosis in patients with surgically resected non-small cell lung cancer. Diagn. Pathol. 2019, 14, 108. [Google Scholar] [CrossRef]
- Cendrowski, J.; Lobo, V.J.; Sendler, M.; Salas, A.; Kuhn, J.P.; Molero, X.; Fukunaga, R.; Mayerle, J.; Lerch, M.M.; Real, F.X. Mnk1 is a novel acinar cell-specific kinase required for exocrine pancreatic secretion and response to pancreatitis in mice. Gut 2015, 64, 937–947. [Google Scholar] [CrossRef]
- Kumar, K.; Chow, C.R.; Ebine, K.; Arslan, A.D.; Kwok, B.; Bentrem, D.J.; Eckerdt, F.D.; Platanias, L.C.; Munshi, H.G. Differential Regulation of ZEB1 and EMT by MAPK-Interacting Protein Kinases (MNK) and eIF4E in Pancreatic Cancer. Mol. Cancer Res. Mcr. 2016, 14, 216–227. [Google Scholar] [CrossRef]
- Muta, D.; Makino, K.; Nakamura, H.; Yano, S.; Kudo, M.; Kuratsu, J. Inhibition of eIF4E phosphorylation reduces cell growth and proliferation in primary central nervous system lymphoma cells. J. Neuro-Oncol. 2011, 101, 33–39. [Google Scholar] [CrossRef]
- Grzmil, M.; Huber, R.M.; Hess, D.; Frank, S.; Hynx, D.; Moncayo, G.; Klein, D.; Merlo, A.; Hemmings, B.A. MNK1 pathway activity maintains protein synthesis in rapalog-treated gliomas. J. Clin. Investig. 2014, 124, 742–754. [Google Scholar] [CrossRef] [PubMed]
- Eckerdt, F.; Beauchamp, E.; Bell, J.; Iqbal, A.; Su, B.; Fukunaga, R.; Lulla, R.R.; Goldman, S.; Platanias, L.C. Regulatory effects of a Mnk2-eIF4E feedback loop during mTORC1 targeting of human medulloblastoma cells. Oncotarget 2014, 5, 8442–8451. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wen, Q.; Luo, J.; Chu, S.; Chen, L.; Xu, L.; Zang, H.; Alnemah, M.M.; Li, J.; Zhou, J.; et al. Suppression Of beta-catenin Nuclear Translocation By CGP57380 Decelerates Poor Progression And Potentiates Radiation-Induced Apoptosis in Nasopharyngeal Carcinoma. Theranostics 2017, 7, 2134–2149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Jiang, L.; Tao, J.; Pan, Z.; He, M.; Su, D.; He, G.; Jiang, Q. Design, synthesis and biological evaluation of 4-aniline-thieno[2,3-d]pyrimidine derivatives as MNK1 inhibitors against renal cell carcinoma and nasopharyngeal carcinoma. Bioorganic Med. Chem. 2019, 27, 2268–2279. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Y.; Lv, C.; Wang, L.; Song, H. Anthelmintic drug niclosamide enhances the sensitivity of chronic myeloid leukemia cells to dasatinib through inhibiting Erk/Mnk1/eIF4E pathway. Biochem. Biophys. Res. Commun. 2016, 478, 893–899. [Google Scholar] [CrossRef]
- Konicek, B.W.; Stephens, J.R.; McNulty, A.M.; Robichaud, N.; Peery, R.B.; Dumstorf, C.A.; Dowless, M.S.; Iversen, P.W.; Parsons, S.; Ellis, K.E.; et al. Therapeutic inhibition of MAP kinase interacting kinase blocks eukaryotic initiation factor 4E phosphorylation and suppresses outgrowth of experimental lung metastases. Cancer Res. 2011, 71, 1849–1857. [Google Scholar] [CrossRef]
- Yan, S.B.; Peek, V.L.; Ajamie, R.; Buchanan, S.G.; Graff, J.R.; Heidler, S.A.; Hui, Y.H.; Huss, K.L.; Konicek, B.W.; Manro, J.R.; et al. LY2801653 is an orally bioavailable multi-kinase inhibitor with potent activity against MET, MST1R, and other oncoproteins, and displays anti-tumor activities in mouse xenograft models. Investig. New Drugs 2013, 31, 833–844. [Google Scholar] [CrossRef]
- Kawada, I.; Hasina, R.; Arif, Q.; Mueller, J.; Smithberger, E.; Husain, A.N.; Vokes, E.E.; Salgia, R. Dramatic antitumor effects of the dual MET/RON small-molecule inhibitor LY2801653 in non-small cell lung cancer. Cancer Res. 2014, 74, 884–895. [Google Scholar] [CrossRef]
- Wu, W.; Bi, C.; Credille, K.M.; Manro, J.R.; Peek, V.L.; Donoho, G.P.; Yan, L.; Wijsman, J.A.; Yan, S.B.; Walgren, R.A. Inhibition of tumor growth and metastasis in non-small cell lung cancer by LY2801653, an inhibitor of several oncokinases, including MET. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 5699–5710. [Google Scholar] [CrossRef]
- Bell, J.B.; Eckerdt, F.D.; Alley, K.; Magnusson, L.P.; Hussain, H.; Bi, Y.; Arslan, A.D.; Clymer, J.; Alvarez, A.A.; Goldman, S.; et al. MNK Inhibition Disrupts Mesenchymal Glioma Stem Cells and Prolongs Survival in a Mouse Model of Glioblastoma. Mol. Cancer Res. Mcr. 2016, 14, 984–993. [Google Scholar] [CrossRef]
- Wang, S.; Li, B.; Liu, B.; Huang, M.; Li, D.; Guan, L.; Zang, J.; Liu, D.; Zhao, L. Design and synthesis of novel 6-hydroxy-4-methoxy-3-methylbenzofuran-7-carboxamide derivatives as potent Mnks inhibitors by fragment-based drug design. Bioorganic Med. Chem. 2018, 26, 4602–4614. [Google Scholar] [CrossRef]
- Yuan, X.; Wu, H.; Bu, H.; Zheng, P.; Zhou, J.; Zhang, H. Design, synthesis and biological evaluation of pyridone-aminal derivatives as MNK1/2 inhibitors. Bioorganic Med. Chem. 2019, 27, 1211–1225. [Google Scholar] [CrossRef]
- Ramalingam, S.; Gediya, L.; Kwegyir-Afful, A.K.; Ramamurthy, V.P.; Purushottamachar, P.; Mbatia, H.; Njar, V.C. First MNKs degrading agents b5phosphorylation of eIF4E, induce apoptosis, inhibit cell growth, migration and invasion in triple negative and Her2-overexpressing breast cancer cell lines. Oncotarget 2014, 5, 530–543. [Google Scholar] [CrossRef] [PubMed]
- Mbatia, H.W.; Ramalingam, S.; Ramamurthy, V.P.; Martin, M.S.; Kwegyir-Afful, A.K.; Njar, V.C. Novel C-4 heteroaryl 13-cis-retinamide Mnk/AR degrading agents inhibit cell proliferation and migration and induce apoptosis in human breast and prostate cancer cells and suppress growth of MDA-MB-231 human breast and CWR22Rv1 human prostate tumor xenografts in mice. J. Med. Chem. 2015, 58, 1900–1914. [Google Scholar] [PubMed]
- Ramalingam, S.; Ramamurthy, V.P.; Gediya, L.K.; Murigi, F.N.; Purushottamachar, P.; Huang, W.; Choi, E.Y.; Zhang, Y.; Vasaitis, T.S.; Kane, M.A.; et al. The Novel Mnk1/2 Degrader and Apoptosis Inducer VNLG-152 Potently Inhibits TNBC Tumor Growth and Metastasis. Cancers (Basel) 2019, 11, 299. [Google Scholar] [CrossRef] [PubMed]
- Ramamurthy, V.P.; Ramalingam, S.; Gediya, L.; Kwegyir-Afful, A.K.; Njar, V.C. Simultaneous targeting of androgen receptor (AR) and MAPK-interacting kinases (MNKs) by novel retinamides inhibits growth of human prostate cancer cell lines. Oncotarget 2015, 6, 3195–3210. [Google Scholar] [CrossRef] [PubMed]
- Ramamurthy, V.P.; Ramalingam, S.; Gediya, L.K.; Njar, V.C.O. The retinamide VNLG-152 inhibits f-AR/AR-V7 and MNK-eIF4E signaling pathways to suppress EMT and castration-resistant prostate cancer xenograft growth. FEBS J. 2018, 285, 1051–1063. [Google Scholar] [CrossRef]
- Kwegyir-Afful, A.K.; Bruno, R.D.; Purushottamachar, P.; Murigi, F.N.; Njar, V.C. Galeterone and VNPT55 disrupt Mnk-eIF4E to inhibit prostate cancer cell migration and invasion. FEBS J. 2016, 283, 3898–3918. [Google Scholar] [CrossRef]
- Kwegyir-Afful, A.K.; Ramalingam, S.; Ramamurthy, V.P.; Purushottamachar, P.; Murigi, F.N.; Vasaitis, T.S.; Huang, W.; Kane, M.A.; Zhang, Y.; Ambulos, N.; et al. Galeterone and The Next Generation Galeterone Analogs, VNPP414 and VNPP433-3beta Exert Potent Therapeutic Effects in Castration-/Drug-Resistant Prostate Cancer Preclinical Models In Vitro and In Vivo. Cancers (Basel) 2019, 11, 1637. [Google Scholar] [CrossRef]
- Kwegyir-Afful, A.K.; Murigi, F.N.; Purushottamachar, P.; Ramamurthy, V.P.; Martin, M.S.; Njar, V.C.O. Galeterone and its analogs inhibit Mnk-eIF4E axis, synergize with gemcitabine, impede pancreatic cancer cell migration, invasion and proliferation and inhibit tumor growth in mice. Oncotarget 2017, 8, 52381–52402. [Google Scholar] [CrossRef]
- Diab, S.; Teo, T.; Kumarasiri, M.; Li, P.; Yu, M.; Lam, F.; Basnet, S.K.; Sykes, M.J.; Albrecht, H.; Milne, R.; et al. Discovery of 5-(2-(phenylamino)pyrimidin-4-yl)thiazol-2(3H)-one derivatives as potent Mnk2 inhibitors: synthesis, SAR analysis and biological evaluation. ChemMedChem 2014, 9, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Teo, T.; Yang, Y.; Yu, M.; Basnet, S.K.; Gillam, T.; Hou, J.; Schmid, R.M.; Kumarasiri, M.; Diab, S.; Albrecht, H.; et al. An integrated approach for discovery of highly potent and selective Mnk inhibitors: Screening, synthesis and SAR analysis. Eur. J. Med. Chem. 2015, 103, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Li, P.; Basnet, S.K.; Kumarasiri, M.; Diab, S.; Teo, T.; Albrecht, H.; Wang, S. Discovery of 4-(dihydropyridinon-3-yl)amino-5-methylthieno[2,3-d]pyrimidine derivatives as potent Mnk inhibitors: Synthesis, structure-activity relationship analysis and biological evaluation. Eur. J. Med. Chem. 2015, 95, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, B.A.; Kaplan, J.B.; Altman, J.K.; Giles, F.J.; Platanias, L.C. Targeting mTOR signaling pathways and related negative feedback loops for the treatment of acute myeloid leukemia. Cancer Biol. Ther. 2015, 16, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Teo, T.; Yu, M.; Yang, Y.; Gillam, T.; Lam, F.; Sykes, M.J.; Wang, S. Pharmacologic co-inhibition of Mnks and mTORC1 synergistically suppresses proliferation and perturbs cell cycle progression in blast crisis-chronic myeloid leukemia cells. Cancer Lett. 2015, 357, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, S.; Sharma, B.; Joshi, S.; Kroczynska, B.; Majchrzak, B.; Stein, B.L.; McMahon, B.; Altman, J.K.; Licht, J.D.; Baker, D.P.; et al. Essential role for the Mnk pathway in the inhibitory effects of type I interferons on myeloproliferative neoplasm (MPN) precursors. J. Biol. Chem. 2013, 288, 23814–23822. [Google Scholar] [CrossRef]
- Abdelaziz, A.M.; Basnet, S.K.C.; Islam, S.; Li, M.; Tadesse, S.; Albrecht, H.; Gerber, C.; Yu, M.; Wang, S. Synthesis and evaluation of 2’H-spiro[cyclohexane-1,3’-imidazo[1,5-a]pyridine]-1’,5’-dione derivatives as Mnk inhibitors. Bioorg. Med. Chem. Lett. 2019, 29, 2650–2654. [Google Scholar] [CrossRef]
- Jin, X.; Merrett, J.; Tong, S.; Flower, B.; Xie, J.; Yu, R.; Tian, S.; Gao, L.; Zhao, J.; Wang, X.; et al. Design, synthesis and activity of Mnk1 and Mnk2 selective inhibitors containing thieno[2,3-d]pyrimidine scaffold. Eur. J. Med. Chem. 2019, 162, 735–751. [Google Scholar] [CrossRef]
- Beggs, J.E.; Tian, S.; Jones, G.G.; Xie, J.; Iadevaia, V.; Jenei, V.; Thomas, G.; Proud, C.G. The MAP kinase-interacting kinases regulate cell migration, vimentin expression and eIF4E/CYFIP1 binding. Biochem. J. 2015, 467, 63–76. [Google Scholar] [CrossRef]
- Zhang, W.; Su, X.; Li, S.; Wang, Y.; Wang, Q.; Zeng, H. Inhibiting MNK Selectively Targets Cervical Cancer via Suppressing eIF4E-Mediated beta-Catenin Activation. Am. J. Med. Sci. 2019, 358, 227–234. [Google Scholar] [CrossRef]
- Kosciuczuk, E.M.; Kar, A.K.; Blyth, G.T.; Fischietti, M.; Abedin, S.; Mina, A.A.; Siliezar, R.; Rzymski, T.; Brzozka, K.; Eklund, E.A.; et al. Inhibitory effects of SEL201 in acute myeloid leukemia. Oncotarget 2019, 10, 7112–7121. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Guo, Q.; Li, V.Z.; Nichol, J.N.; Huang, F.; Yang, W.; Preston, S.E.J.; Talat, Z.; Lefrere, H.; Yu, H.; Zhang, G.; et al. MNK1/NODAL signaling promotes invasive progression of breast ductal carcinoma in situ. Cancer Res. 2019, 79, 1646–1657. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Guo, J.; Yang, W.; Goncalves, C.; Rzymski, T.; Dreas, A.; Zylkiewicz, E.; Mikulski, M.; Brzozka, K.; Golas, A.; et al. MNK1/2 inhibition limits oncogenicity and metastasis of KIT-mutant melanoma. J. Clin. Investig. 2017, 127, 4179–4192. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Khoury, E.; Guo, Q.; Prabhu, S.A.; Emond, A.; Huang, F.; Goncalves, C.; Zhan, Y.; Plourde, D.; Nichol, J.N.; et al. MNK1 signaling induces an ANGPTL4-mediated gene signature to drive melanoma progression. Oncogene 2020. [Google Scholar] [CrossRef]
- Mishra, R.K.; Clutter, M.R.; Blyth, G.T.; Kosciuczuk, E.M.; Blackburn, A.Z.; Beauchamp, E.M.; Schiltz, G.E.; Platanias, L.C. Discovery of novel Mnk inhibitors using mutation-based induced-fit virtual high-throughput screening. Chem. Biol. Drug Des. 2019, 94, 1813–1823. [Google Scholar] [CrossRef]
- Reich, S.H.; Sprengeler, P.A.; Chiang, G.G.; Appleman, J.R.; Chen, J.; Clarine, J.; Eam, B.; Ernst, J.T.; Han, Q.; Goel, V.K.; et al. Structure-based Design of Pyridone-Aminal eFT508 Targeting Dysregulated Translation by Selective Mitogen-activated Protein Kinase Interacting Kinases 1 and 2 (MNK1/2) Inhibition. J. Med. Chem. 2018, 61, 3516–3540. [Google Scholar] [CrossRef]
- Wu, H.; Hu, C.; Wang, A.; Weisberg, E.L.; Chen, Y.; Yun, C.H.; Wang, W.; Liu, Y.; Liu, X.; Tian, B.; et al. Discovery of a BTK/MNK dual inhibitor for lymphoma and leukemia. Leukemia 2016, 30, 173–181. [Google Scholar] [CrossRef]
- Bentayeb, H.; Aitamer, M.; Petit, B.; Dubanet, L.; Elderwish, S.; Desaubry, L.; de Gramont, A.; Raymond, E.; Olivrie, A.; Abraham, J.; et al. Prohibitin (PHB) expression is associated with aggressiveness in DLBCL and flavagline-mediated inhibition of cytoplasmic PHB functions induces anti-tumor effects. J. Exp. Clin. Cancer Res. 2019, 38, 450. [Google Scholar] [CrossRef]
- Cherian, J.; Nacro, K.; Poh, Z.Y.; Guo, S.; Jeyaraj, D.A.; Wong, Y.X.; Ho, M.; Yang, H.Y.; Joy, J.K.; Kwek, Z.P.; et al. Structure-Activity Relationship Studies of Mitogen Activated Protein Kinase Interacting Kinase (MNK) 1 and 2 and BCR-ABL1 Inhibitors Targeting Chronic Myeloid Leukemic Cells. J. Med. Chem. 2016, 59, 3063–3078. [Google Scholar] [CrossRef]
- Lock, R.; Ingraham, R.; Maertens, O.; Miller, A.L.; Weledji, N.; Legius, E.; Konicek, B.M.; Yan, S.C.; Graff, J.R.; Cichowski, K. Cotargeting MNK and MEK kinases induces the regression of NF1-mutant cancers. J. Clin. Investig. 2016, 126, 2181–2190. [Google Scholar] [CrossRef]
- Garcia-Recio, E.M.; Pinto-Diez, C.; Perez-Morgado, M.I.; Garcia-Hernandez, M.; Fernandez, G.; Martin, M.E.; Gonzalez, V.M. Characterization of MNK1b DNA Aptamers That Inhibit Proliferation in MDA-MB231 Breast Cancer Cells. Mol. Ther. Nucleic Acids 2016, 5, e275. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Tamamyan, G.; Kadia, T.; Ravandi, F.; Borthakur, G.; Cortes, J.; Jabbour, E.; Daver, N.; Ohanian, M.; Kantarjian, H.; Konopleva, M. Frontline treatment of acute myeloid leukemia in adults. Crit. Rev. Oncol. Hematol. 2017, 110, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Diab, S.; Li, P.; Basnet, S.K.; Lu, J.; Yu, M.; Albrecht, H.; Milne, R.W.; Wang, S. Unveiling new chemical scaffolds as Mnk inhibitors. Future Med. Chem. 2016, 8, 271–285. [Google Scholar] [CrossRef]
- Altman, J.K.; Szilard, A.; Konicek, B.W.; Iversen, P.W.; Kroczynska, B.; Glaser, H.; Sassano, A.; Vakana, E.; Graff, J.R.; Platanias, L.C. Inhibition of Mnk kinase activity by cercosporamide and suppressive effects on acute myeloid leukemia precursors. Blood 2013, 121, 3675–3681. [Google Scholar] [CrossRef]
- Li, P.; Diab, S.; Yu, M.; Adams, J.; Islam, S.; Basnet, S.K.; Albrecht, H.; Milne, R.; Wang, S. Inhibition of Mnk enhances apoptotic activity of cytarabine in acute myeloid leukemia cells. Oncotarget 2016, 7, 56811–56825. [Google Scholar] [CrossRef]
- Abdelaziz, A.M.; Diab, S.; Islam, S.; Basnet, S.K.C.; Noll, B.; Li, P.; Mekonnen, L.B.; Lu, J.; Albrecht, H.; Milne, R.W.; et al. Discovery of N-Phenyl-4-(1H-pyrrol-3-yl)pyrimidin-2-amine Derivatives as Potent Mnk2 Inhibitors: Design, Synthesis, SAR Analysis, and Evaluation of in vitro Anti-leukaemic Activity. Med. Chem. 2019, 15, 602–623. [Google Scholar] [CrossRef]
- Zhang, M.; Fu, W.; Prabhu, S.; Moore, J.C.; Ko, J.; Kim, J.W.; Druker, B.J.; Trapp, V.; Fruehauf, J.; Gram, H.; et al. Inhibition of polysome assembly enhances imatinib activity against chronic myelogenous leukemia and overcomes imatinib resistance. Mol. Cell. Biol. 2008, 28, 6496–6509. [Google Scholar] [CrossRef]
- Jain, P.; Kantarjian, H.M.; Ghorab, A.; Sasaki, K.; Jabbour, E.J.; Nogueras Gonzalez, G.; Kanagal-Shamanna, R.; Issa, G.C.; Garcia-Manero, G.; Kc, D.; et al. Prognostic factors and survival outcomes in patients with chronic myeloid leukemia in blast phase in the tyrosine kinase inhibitor era: Cohort study of 477 patients. Cancer 2017, 123, 4391–4402. [Google Scholar] [CrossRef]
- Marzec, M.; Liu, X.; Wysocka, M.; Rook, A.H.; Odum, N.; Wasik, M.A. Simultaneous inhibition of mTOR-containing complex 1 (mTORC1) and MNK induces apoptosis of cutaneous T-cell lymphoma (CTCL) cells. PLoS ONE 2011, 6, e24849. [Google Scholar] [CrossRef]
- Dubois, S.; Jardin, F. Novel molecular classifications of DLBCL. Nat. Rev. Clin. Oncol. 2018, 15, 474–476. [Google Scholar] [CrossRef] [PubMed]
- Gajek, A.; Poczta, A.; Lukawska, M.; Cecuda-Adamczewska, V.; Tobiasz, J.; Marczak, A. Chemical modification of melphalan as a key to improving treatment of haematological malignancies. Sci. Rep. 2020, 10, 4479. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Chen, A.; Joy, J.; Xavier, V.J.; Ong, E.H.; Hill, J.; Chai, C.L. Rational design of resorcylic acid lactone analogues as covalent MNK1/2 kinase inhibitors by tuning the reactivity of an enamide Michael acceptor. ChemMedChem 2013, 8, 1483–1494. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, V.; Ananda Rao, B.; Sharma, P.; Swarna, B.; Thummuri, D.; Srinivas, K.; Naidu, V.G.; Jayathirtha Rao, V. Synthesis and biological evaluation of new rhodanine analogues bearing 2-chloroquinoline and benzo[h]quinoline scaffolds as anticancer agents. Eur. J. Med. Chem. 2014, 83, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Lineham, E.; Tizzard, G.J.; Coles, S.J.; Spencer, J.; Morley, S.J. Synergistic effects of inhibiting the MNK-eIF4E and PI3K/AKT/ mTOR pathways on cell migration in MDA-MB-231 cells. Oncotarget 2018, 9, 14148–14159. [Google Scholar] [CrossRef] [PubMed]
- Sansook, S.; Lineham, E.; Hassell-Hart, S.; Tizzard, G.J.; Coles, S.J.; Spencer, J.; Morley, S.J. Probing the Anticancer Action of Novel Ferrocene Analogues of MNK Inhibitors. Molecules 2018, 23, 2126. [Google Scholar] [CrossRef]
- Astanehe, A.; Finkbeiner, M.R.; Krzywinski, M.; Fotovati, A.; Dhillon, J.; Berquin, I.M.; Mills, G.B.; Marra, M.A.; Dunn, S.E. MKNK1 is a YB-1 target gene responsible for imparting trastuzumab resistance and can be blocked by RSK inhibition. Oncogene 2012, 31, 4434–4446. [Google Scholar] [CrossRef]
- Evans, M.K.; Brown, M.C.; Geradts, J.; Bao, X.; Robinson, T.J.; Jolly, M.K.; Vermeulen, P.B.; Palmer, G.M.; Gromeier, M.; Levine, H.; et al. XIAP Regulation by MNK Links MAPK and NFkappaB Signaling to Determine an Aggressive Breast Cancer Phenotype. Cancer Res. 2018, 78, 1726–1738. [Google Scholar] [CrossRef]
- Travis, W.D. Update on small cell carcinoma and its differentiation from squamous cell carcinoma and other non-small cell carcinomas. Mod. Pathol. 2012, 25, S18–S30. [Google Scholar] [CrossRef]
- Lu, J.; Zang, H.; Zheng, H.; Zhan, Y.; Yang, Y.; Zhang, Y.; Liu, S.; Feng, J.; Wen, Q.; Long, M.; et al. Overexpression of p-Akt, p-mTOR and p-eIF4E proteins associates with metastasis and unfavorable prognosis in non-small cell lung cancer. PLoS ONE 2020, 15, e0227768. [Google Scholar] [CrossRef]
- Zhu, Z.C.; Liu, J.W.; Yang, C.; Zhao, M.; Xiong, Z.Q. XPO1 inhibitor KPT-330 synergizes with Bcl-xL inhibitor to induce cancer cell apoptosis by perturbing rRNA processing and Mcl-1 protein synthesis. Cell Death Dis. 2019, 10, 395. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Hou, Y.; Yu, X.; Zhang, F.; Wu, X.; Wu, X.; Wang, L. The ERK-MNK-eIF4F signaling pathway mediates TPDHT-induced A549 cell death in vitro and in vivo. Food Chem. Toxicol. 2020, 137, 111158. [Google Scholar] [CrossRef] [PubMed]
- Scrima, M.; De Marco, C.; Fabiani, F.; Franco, R.; Pirozzi, G.; Rocco, G.; Ravo, M.; Weisz, A.; Zoppoli, P.; Ceccarelli, M.; et al. Signaling networks associated with AKT activation in non-small cell lung cancer (NSCLC): New insights on the role of phosphatydil-inositol-3 kinase. PLoS ONE 2012, 7, e30427. [Google Scholar] [CrossRef] [PubMed]
- Solomon, B.; Pearson, R.B. Class IA phosphatidylinositol 3-kinase signaling in non-small cell lung cancer. J. Thorac. Oncol. Off. Publ. Int. Assoc. Study Lung Cancer 2009, 4, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Fumarola, C.; Bonelli, M.A.; Petronini, P.G.; Alfieri, R.R. Targeting PI3K/AKT/mTOR pathway in non small cell lung cancer. Biochem. Pharmacol. 2014, 90, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Furic, L.; Rong, L.; Larsson, O.; Koumakpayi, I.H.; Yoshida, K.; Brueschke, A.; Petroulakis, E.; Robichaud, N.; Pollak, M.; Gaboury, L.A.; et al. eIF4E phosphorylation promotes tumorigenesis and is associated with prostate cancer progression. Proc. Natl. Acad. Sci. USA 2010, 107, 14134–14139. [Google Scholar] [CrossRef]
- Graff, J.R.; Konicek, B.W.; Lynch, R.L.; Dumstorf, C.A.; Dowless, M.S.; McNulty, A.M.; Parsons, S.H.; Brail, L.H.; Colligan, B.M.; Koop, J.W.; et al. eIF4E activation is commonly elevated in advanced human prostate cancers and significantly related to reduced patient survival. Cancer Res. 2009, 69, 3866–3873. [Google Scholar] [CrossRef]
- D’Abronzo, L.S.; Bose, S.; Crapuchettes, M.E.; Beggs, R.E.; Vinall, R.L.; Tepper, C.G.; Siddiqui, S.; Mudryj, M.; Melgoza, F.U.; Durbin-Johnson, B.P.; et al. The androgen receptor is a negative regulator of eIF4E phosphorylation at S209: Implications for the use of mTOR inhibitors in advanced prostate cancer. Oncogene 2017, 36, 6359–6373. [Google Scholar] [CrossRef]
- ur Rasool, R.; Rah, B.; Amin, H.; Nayak, D.; Chakraborty, S.; Rawoof, A.; Mintoo, M.J.; Yousuf, K.; Mukherjee, D.; Kumar, L.D.; et al. Dual modulation of Ras-Mnk and PI3K-AKT-mTOR pathways: A Novel c-FLIP inhibitory mechanism of 3-AWA mediated translational attenuation through dephosphorylation of eIF4E. Sci. Rep. 2016, 6, 18800. [Google Scholar] [CrossRef]
- Stead, R.L.; Proud, C.G. Rapamycin enhances eIF4E phosphorylation by activating MAP kinase-interacting kinase 2a (Mnk2a). FEBS Lett. 2013, 587, 2623–2628. [Google Scholar] [CrossRef]
- Rahaman, M.H.; Lam, F.; Zhong, L.; Teo, T.; Adams, J.; Yu, M.; Milne, R.W.; Pepper, C.; Lokman, N.A.; Ricciardelli, C.; et al. Targeting CDK9 for treatment of colorectal cancer. Mol. Oncol. 2019, 13, 2178–2193. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Reineke, L.C.; Knutsen, E.; Chen, M.; Pichler, M.; Ling, H.; Calin, G.A. Metformin blocks MYC protein synthesis in colorectal cancer via mTOR-4EBP-eIF4E and MNK1-eIF4G-eIF4E signaling. Mol. Oncol. 2018, 12, 1856–1870. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sun, L.; Su, X.; Guo, S. Inhibition of eukaryotic initiation factor 4E phosphorylation by cercosporamide selectively suppresses angiogenesis, growth and survival of human hepatocellular carcinoma. Biomed. Pharmacother. 2016, 84, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Molinaro, A.M.; Taylor, J.W.; Wiencke, J.K.; Wrensch, M.R. Genetic and molecular epidemiology of adult diffuse glioma. Nat. Rev. Neurol. 2019, 15, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Wang, W.; Mao, X.; Chu, S.; Feng, J.; Xiao, D.; Zhou, J.; Fan, S. Elevated levels of p-Mnk1, p-eIF4E and p-p70S6K proteins are associated with tumor recurrence and poor prognosis in astrocytomas. J. Neuro-Oncol. 2017, 131, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.B.; Eckerdt, F.; Dhruv, H.D.; Finlay, D.; Peng, S.; Kim, S.; Kroczynska, B.; Beauchamp, E.M.; Alley, K.; Clymer, J.; et al. Differential Response of Glioma Stem Cells to Arsenic Trioxide Therapy Is Regulated by MNK1 and mRNA Translation. Mol. Cancer Res. Mcr. 2018, 16, 32–46. [Google Scholar] [CrossRef]
- Das, F.; Ghosh-Choudhury, N.; Bera, A.; Kasinath, B.S.; Choudhury, G.G. TGFbeta-induced PI 3 kinase-dependent Mnk-1 activation is necessary for Ser-209 phosphorylation of eIF4E and mesangial cell hypertrophy. J. Cell. Physiol. 2013, 228, 1617–1626. [Google Scholar] [CrossRef] [PubMed]
- Robichaud, N.; del Rincon, S.V.; Huor, B.; Alain, T.; Petruccelli, L.A.; Hearnden, J.; Goncalves, C.; Grotegut, S.; Spruck, C.H.; Furic, L.; et al. Phosphorylation of eIF4E promotes EMT and metastasis via translational control of SNAIL and MMP-3. Oncogene 2015, 34, 2032–2042. [Google Scholar] [CrossRef]
- Northcott, P.A.; Robinson, G.W.; Kratz, C.P.; Mabbott, D.J.; Pomeroy, S.L.; Clifford, S.C.; Rutkowski, S.; Ellison, D.W.; Malkin, D.; Taylor, M.D.; et al. Medulloblastoma. Nat. Rev. Dis. Primers 2019, 5, 11. [Google Scholar] [CrossRef]
- Grzmil, M.; Seebacher, J.; Hess, D.; Behe, M.; Schibli, R.; Moncayo, G.; Frank, S.; Hemmings, B.A. Inhibition of MNK pathways enhances cancer cell response to chemotherapy with temozolomide and targeted radionuclide therapy. Cell Signal. 2016, 28, 1412–1421. [Google Scholar] [CrossRef]
- Wahba, A.; Rath, B.H.; Bisht, K.; Camphausen, K.; Tofilon, P.J. Polysome Profiling Links Translational Control to the Radioresponse of Glioblastoma Stem-like Cells. Cancer Res. 2016, 76, 3078–3087. [Google Scholar] [CrossRef]
- Eckerdt, F.; Bell, J.B.; Beauchamp, E.M.; Clymer, J.; Blyth, G.T.; Kosciuczuk, E.M.; Ma, Q.; Chen, D.Z.; Horbinski, C.; Goldman, S.; et al. Potent Antineoplastic Effects of Combined PI3Kalpha-MNK Inhibition in Medulloblastoma. Mol. Cancer Res. Mcr. 2019, 17, 1305–1315. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Zhang, J.; Yu, M.; Xiong, C. Inhibition of Mnk-eIF4E pathway sensitizes the efficacy to chemotherapy in anaplastic thyroid cancer. Future Oncol. 2017, 13, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.N.D.; Kumar, K.; DeCant, B.T.; Shang, M.; Munshi, S.Z.; Matsangou, M.; Ebine, K.; Munshi, H.G. Induction of MNK Kinase-dependent eIF4E Phosphorylation by Inhibitors Targeting BET Proteins Limits Efficacy of BET Inhibitors. Mol. Cancer Ther. 2019, 18, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zha, J.; Lei, M. Inhibiting ERK/Mnk/eIF4E broadly sensitizes ovarian cancer response to chemotherapy. Clin. Transl. Oncol. 2018, 20, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Li, J.; Xu, L.; Xie, G.; Wen, Q.; Luo, J.; Li, D.; Huang, D.; Fan, S. Phosphorylated Mnk1 and eIF4E are associated with lymph node metastasis and poor prognosis of nasopharyngeal carcinoma. PLoS ONE 2014, 9, e89220. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Liu, Y.; Di, X.; Kang, H.; Zeng, H.; Zhao, Y.; Cai, K.; Pang, T.; Wang, S.; Yao, Y.; et al. EIF4E over-expresses and enhances cell proliferation and cell cycle progression in nasopharyngeal carcinoma. Med. Oncol. 2013, 30, 400. [Google Scholar] [CrossRef] [PubMed]
- Ichiyanagi, O.; Ito, H.; Naito, S.; Kabasawa, T.; Kanno, H.; Narisawa, T.; Ushijima, M.; Kurota, Y.; Ozawa, M.; Sakurai, T.; et al. Impact of eIF4E phosphorylation at Ser209 via MNK2a on tumour recurrence after curative surgery in localized clear cell renal cell carcinoma. Oncotarget 2019, 10, 4053–4068. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Comments | Indications | Reference |
|---|---|---|---|
| BAY1143269 | MNK inhibitor | NSCLC, leukemia | [61] |
| CGP57380 | MNK inhibitor | NSCLC, breast cancer, leukemia, lymphoma, myeloma, glioma, PCNSL, medulloblastoma, nasopharyngeal carcinoma, PDAC, ovarian cancer | [40,46,50,51,55,62,63,64,65,66,67,68,69,70,71,72,73,74,75] |
| Cercosporamide | MNK inhibitor | NSCLC, colorectal cancer, liver cancer, breast cancer, leukemia, lymphoma | [40,62,65,76,77] |
| Merestinib | Multi-kinase inhibitor | NSCLC, leukemia, glioma | [78,79,80,81] |
| Benzofuran derivatives (5o,8k) | MNK inhibitors | Colorectal cancer, leukemia | [82,83] |
| Novel retinamides (VNLG-152R, VNHM-1-81, VNHM-1-66, VNHM-1-73) | MNK degraders | Prostate and breast cancer | [84,85,86,87,88] |
| Galeterone and galeterone analogs | MNK degraders | Prostate cancer, PDAC | [89,90,91] |
| Pyridine derivatives (4t, MNK7g, MNKI-1, MNKI-8e, MNKI-8i, MNKI-19, MNKI-85, MNKI-4, MNKI-57, 12j) | MNK inhibitors | Cervical, breast, colorectal, ovarian, pancreatic and prostate cancer, medulloblastoma, leukemia, myeloma, nasopharyngeal carcinoma | [64,75,92,93,94,95,96,97,98,99,100,101] |
| SEL201 = SLV-2436 | MNK inhibitor | Breast cancer, leukemia, melanoma | [102,103,104,105] |
| NUCC-54139 | MNK inhibitor | Leukemia | [106] |
| Niclosamide (anthelmintic drug) | Targets ERK/MNK1/eIF4E | Leukemia | [76] |
| eFT508 | MNK inhibitor | Lymphoma | [107] |
| QL-X-138 | BTK/MNK inhibitor | Leukemia, lymphoma | [108] |
| FL3 (synthetic flavagline) | Ligand of PHBs (Targets ERK/MNK/eIF4E) | Lymphoma | [109] |
| Cyclopropanecarboxamide derivatives (53,54) | MNK inhibitors | Leukemia | [110] |
| Cabozantinib | Multi-kinase inhibitor | MPNSTs | [111] |
| apMNK2F, apMNK3R (aptamer) | MNK inhibitor | Breast cancer | [112] |
| Compound | Phase | Name and Identifier | Status | Type of Cancer | Combination |
|---|---|---|---|---|---|
| BAY1143269 | I | Phase I Dose Escalation and Expansion of Oral BAY1143269 in Combination With Intravenous Docetaxel NCT02439346 | Terminated | Metastatic solid tumors | Docetaxel |
| eFT508 (Tomivosertib) | II | An Open-label Study Examining the Effect of Tomivosertib (eFT508) in Patients With Advanced Castrate-resistant Prostate Cancer (CRCP) NCT03690141 | Active, not recruiting | Castrate-resistant prostate cancer | |
| II | A Study to Evaluate eFT508 Alone and in Combination With Avelumab in Subljects With MSS Colorectal Cancer NCT03258398 | Completed | Refractory colorectal cancer | Avelumab | |
| II | A PD Study of Oral eFT508 in Subjects With Advanced TNBC and HCC NCT03318562 | Terminated | Triple negative breast cancer and hepatocellular carcinoma | ||
| I-II | A Dose Escalation and Cohort-Expansion Study of Oral eFT508 in Subjects With Advanced Solid Tumors NCT02605083 | Terminated | Solid tumors | ||
| I-II | A Phase 1–2 Dose-Escalation and Cohort-Expansion Study of Oral Tomivosertib (eFT508) in Subjects With Hematological Malignancies NCT02937675 | Terminated | Lymphoma | ||
| II | Tomivosertib (eFT508) in Combination With PD-1/PDL-1 Inhibitor Therapy NCT03616834 | Active, not recruiting | Solid tumors | PD-1/PD-L1 | |
| I | Safety, Pharmacodynamics, Pharmacokinetics, and Efficacy of Tomivosertib Combined With Paclitaxel in Advanced Breast Cancer NCT04261218 | Not yet recruiting | Advanced breast cancer | Paclitaxel | |
| Merestinib | I | Combination Merestinib and LY2874455 for Patients With Relapsed or Refractory Acute Myeloid Leukemia NCT03125239 | Recruiting | Relapsed and Refractory Adult Acute Myeloid Leukemia | LY2874455 |
| I | A Study of Merestinib (LY2801653) in Japanese Participants With Advanced or Metastatic Cancer NCT03027284 | Active, not recruiting | Advanced cancer, metastatic cancer, biliary tract carcinoma, cholangiocarcinoma, gall bladder carcinoma, solid tumor, non-Hodgkin’s lymphoma | Cisplatin and Gemcitabine | |
| I | Merestinib on Bone Metastases in Subjects With Breast Cancer NCT03292536 | Recruiting | Bone metastases, breast cancer | ||
| I | A Study of Merestinib (LY2801653) in Healthy Participants NCT02779738 | Completed | |||
| II | Merestinib in Non-Small Cell Lung Cancer and Solid Tumors NCT02920996 | Active, not recruiting | Non-small cell lung cancer, solid tumors | ||
| II | A Study of Ramucirumab (LY3009806) or Merestinib (LY2801653) in Advanced or Metastatic Biliary Tract Cancer NCT02711553 | Active, not recruiting | Biliary tract cancer, metastatic cancer, advanced cancer | Cisplatine and Gemcitabine | |
| I | A Study in Advanced Cancers Using Ramucirumab (LY3009806) and Other Targeted Agents NCT02745769 | Completed | Advanced cancer, colorectal cancer, mantle cell lymphoma | Ramucirumab | |
| I | A Study of Anti-PD-L1 Checkpoint Antibody (LY3300054) Alone and in Combination in Participants With Advanced Refractory Solid Tumors NCT02791334 | Recruiting | Solid tumor, microsatellite instability—high (MSI-H) solid tumors, cutaneous melanoma, pancreatic cancer, breast cancer (HR+HER2-) | LY3300054 | |
| Galeterone | II | 1911GCCC: Galeterone or Galeterone With Gemcitabine for Patients With Metastatic Pancreatic Adenocarcinoma NCT04098081 | Recruiting | Advanced Pancreatic Cancer | Gemcitabine |
| I | Single-Dose Study to Assess the Absorption, Metabolism, Excretion, and Mass Balance of Radiolabeled Galeterone NCT02729376 | Completed | Healthy | ||
| III | A Study of Galeterone Compared to Enzalutamide In Men Expressing Androgen Receptor Splice Variant-7 mRNA (AR-V7) Metastatic CRPC (ARMOR3-SV) NCT02438007 | Terminated | Prostate cancer | ||
| II | A 2 Part Phase 2 Trial of Galeterone in the Treatment of Castration Resistant Prostate Cancer (ARMOR2) NCT01709734 | Completed | Prostate cancer |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinto-Díez, C.; Ferreras-Martín, R.; Carrión-Marchante, R.; González, V.M.; Martín, M.E. Deeping in the Role of the MAP-Kinases Interacting Kinases (MNKs) in Cancer. Int. J. Mol. Sci. 2020, 21, 2967. https://doi.org/10.3390/ijms21082967
Pinto-Díez C, Ferreras-Martín R, Carrión-Marchante R, González VM, Martín ME. Deeping in the Role of the MAP-Kinases Interacting Kinases (MNKs) in Cancer. International Journal of Molecular Sciences. 2020; 21(8):2967. https://doi.org/10.3390/ijms21082967
Chicago/Turabian StylePinto-Díez, Celia, Raquel Ferreras-Martín, Rebeca Carrión-Marchante, Víctor M. González, and María Elena Martín. 2020. "Deeping in the Role of the MAP-Kinases Interacting Kinases (MNKs) in Cancer" International Journal of Molecular Sciences 21, no. 8: 2967. https://doi.org/10.3390/ijms21082967
APA StylePinto-Díez, C., Ferreras-Martín, R., Carrión-Marchante, R., González, V. M., & Martín, M. E. (2020). Deeping in the Role of the MAP-Kinases Interacting Kinases (MNKs) in Cancer. International Journal of Molecular Sciences, 21(8), 2967. https://doi.org/10.3390/ijms21082967