Conventional and Molecular Techniques from Simple Breeding to Speed Breeding in Crop Plants: Recent Advances and Future Outlook


 , ,
, , 

Abstract
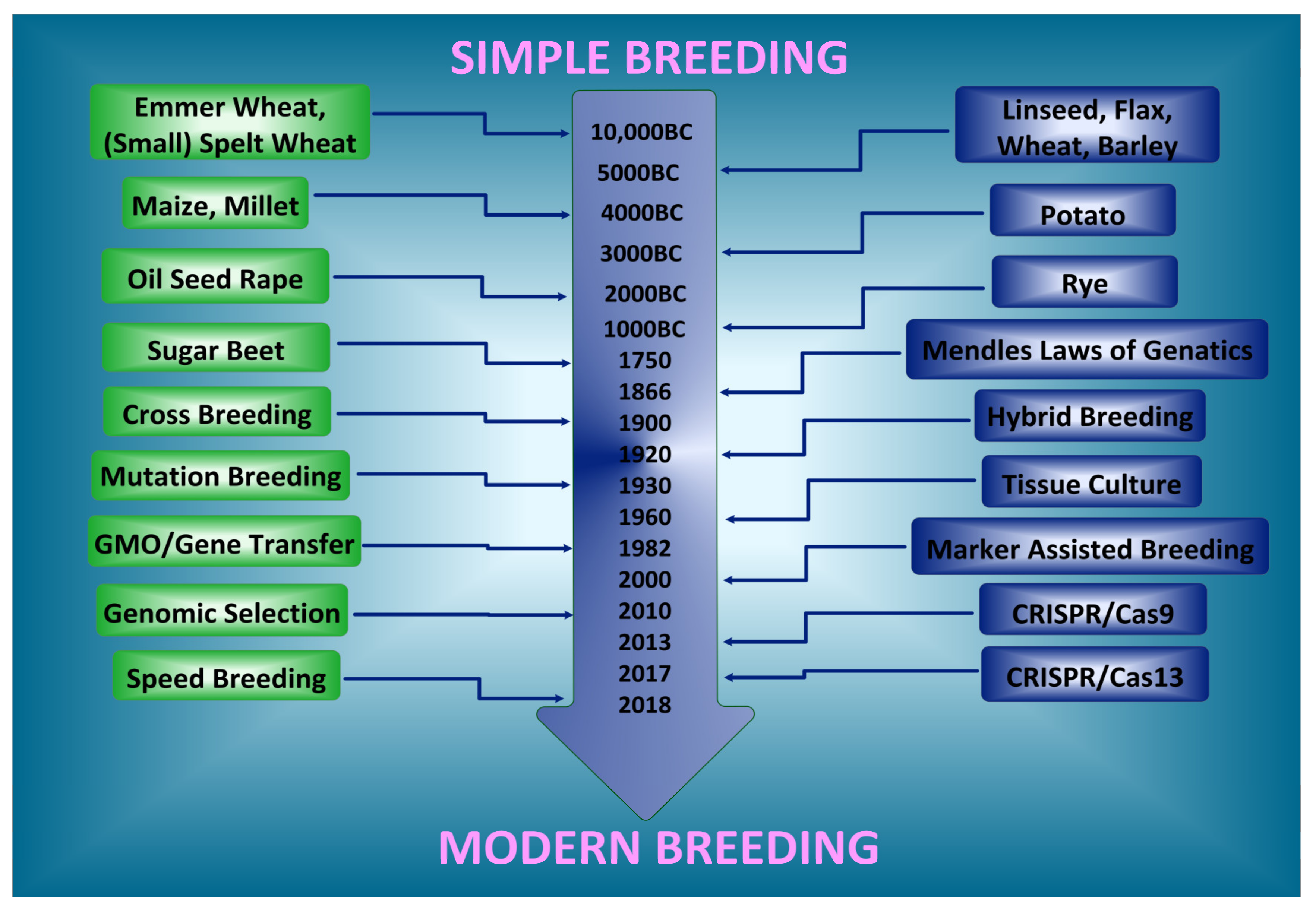
1. Introduction
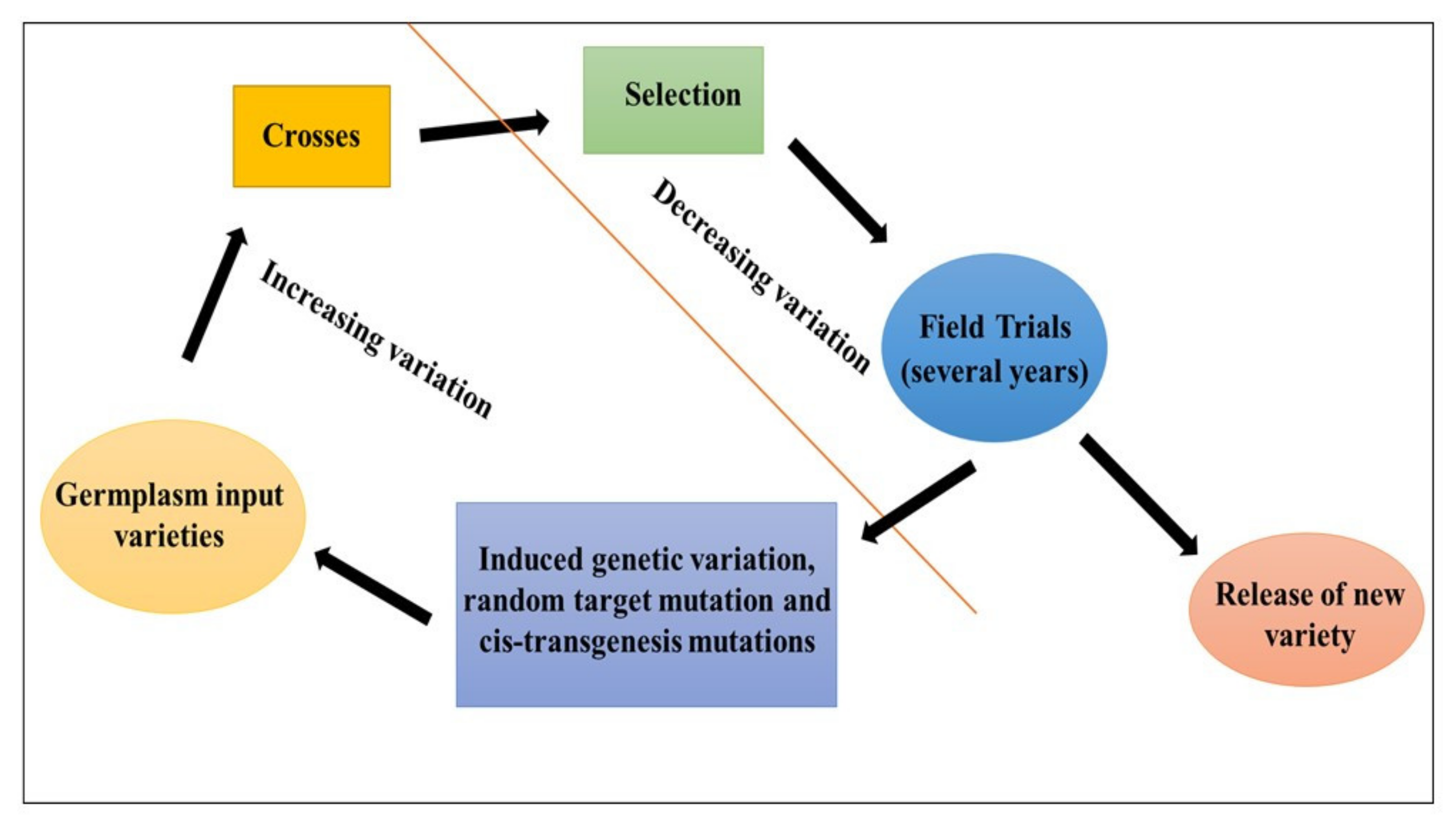
2. Mutation through Traditional or Conventional Breeding
3. Mutagens for Molecular Breeding
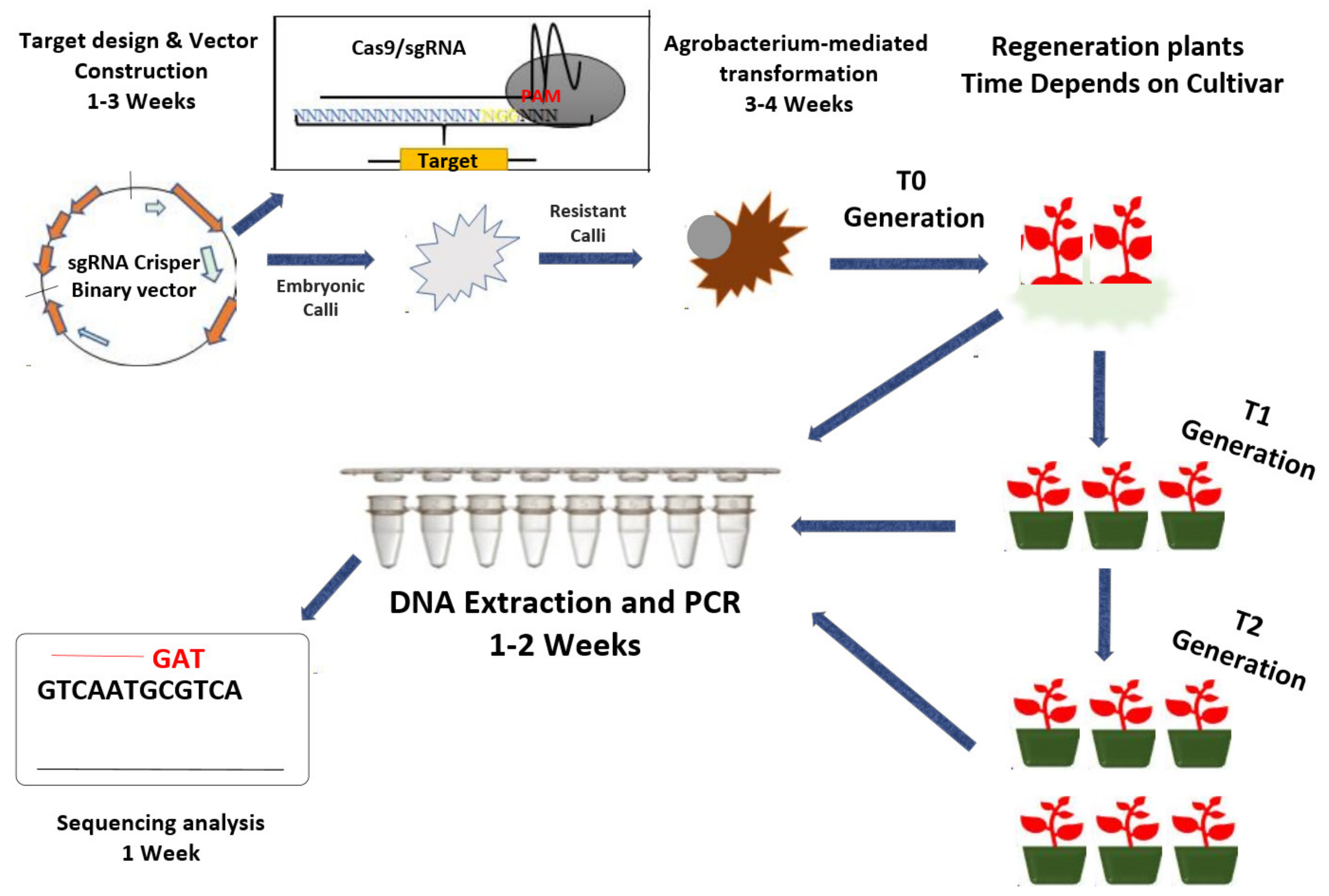
4. CRISPR/Cas9 and CRISPR/Cpf1 as Genetic Dissection Tools
5. Speed Breeding (Time-Saving Tools) for Accelerating Plant Breeding
6. Contribution of Plant Breeding to Crop Improvement
7. Future Outlook
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR); |
| Cas9 | CRISPR-associated Proteins; |
| Cpf1 | CRISPR-associated endonuclease in Prevotella and Francisella; |
| DSB | Double Strand Breaks; |
| HTP | High-Throughput Phenotyping; |
| NASA | National Aeronautics and Space Administration; |
| USU | Utah State University; |
| GMO | Genetically Modified Organism; |
| EU | European Union; |
| Non-GMO | Non-Genetically modified Organism; |
| SSD | Single Seed Descent; |
| SB | Speed Breeding; |
| RGA | Rapid Generation Advance; |
| UQ | University of Queensland; |
| DH | Double Haploid; |
| ICRISAT | International Crop Research Institute for the Semi-Arid Tropics. |
References
- Tester, M.; Langridge, P. Breeding technologies to increase crop production in a changing world. Science (80-. ). 2010, 327, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Shiferaw, B.; Smale, M.; Braun, H.-J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef]
- Von Braun, J.; Rosegrant, M.W.; Pandya-Lorch, R.; Cohen, M.J.; Cline, S.A.; Brown, M.A.; Bos, M.S. New Risks and Opportunities for Food Security Scenario Analyses for 2015 and 2050; IFPRI: Washington, DC, USA, 2005. [Google Scholar]
- Ristaino, J.B. Tracking historic migrations of the Irish potato famine pathogen, Phytophthora infestans. Microbes Infect. 2002, 4, 1369–1377. [Google Scholar] [CrossRef]
- Tatum, L.A. The Southern Corn Leaf Blight Epidemic. Science 1971, 171, 1113–1116. [Google Scholar] [CrossRef]
- UN World Population Projected to Reach 9.8 Billion in 2050, and 11.2 Billion in 2100. Available online: https://www.un.org/development/desa/en/news/population/world-population-prospects-2017.html.
- FAO How to Feed the World in 2050. In Insights from an Expert Meet; FAO: Roma, Italy, 2009; Volume 2050, pp. 1–35.
- Voss-Fels, K.P.; Stahl, A.; Hickey, L.T. Q&A: Modern crop breeding for future food security. BMC Biol. 2019, 17, 18. [Google Scholar]
- Cheema, K.S. K. Plant Breeding its Applications and Future Prospects. Int. J. Eng. Technol. Sci. Res. 2018, 5, 88–94. [Google Scholar]
- Moose, S.P.; Mumm, R.H. Molecular Plant Breeding as the Foundation for 21st Century Crop Improvement. Plant Physiol. 2008, 147, 969–977. [Google Scholar] [CrossRef]
- Varshney, R.K.; Hoisington, D.A.; Tyagi, A.K. Advances in cereal genomics and applications in crop breeding. Trends Biotechnol. 2006, 24, 490–499. [Google Scholar] [CrossRef]
- Li, H.; Rasheed, A.; Hickey, L.T.; He, Z. Fast-forwarding genetic gain. Trends Plant Sci. 2018, 23, 184–186. [Google Scholar] [CrossRef] [PubMed]
- Collins, F.S.; Green, E.D.; Guttmacher, A.E.; Guyer, M.S. A vision for the future of genomics research. Nature 2003, 431, 835–847. [Google Scholar]
- Majid, A.; Parray, G.A.; Wani, S.H.; Kordostami, M.; Sofi, N.R.; Waza, S.A.; Shikari, A.B.; Gulzar, S. Genome Editing and its Necessity in Agriculture. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 5435–5443. [Google Scholar] [CrossRef]
- Araus, J.L.; Kefauver, S.C.; Zaman-Allah, M.; Olsen, M.S.; Cairns, J.E. Translating High-Throughput Phenotyping into Genetic Gain. Trends Plant Sci. 2018, 23, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Asyraf Md Hatta, M.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wen, Y.; Guo, X. CRISPR/Cas9 for genome editing: Progress, implications and challenges. Hum. Mol. Genet. 2014, 23, R40–R46. [Google Scholar] [CrossRef] [PubMed]
- Murovec, J.; Pirc, Ž.; Yang, B. New variants of CRISPR RNA-guided genome editing enzymes. Plant Biotechnol. J. 2017, 15, 917–926. [Google Scholar] [CrossRef]
- Acquaah, G. Polyploidy in Plant Breeding. In Principles of Plant Genetics and Breeding; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp. 452–469. [Google Scholar]
- Muth, J.; Hartje, S.; Twyman, R.M.; Hofferbert, H.R.; Tacke, E.; Prüfer, D. Precision breeding for novel starch variants in potato. Plant Biotechnol. J. 2008, 6, 576–584. [Google Scholar] [CrossRef]
- Mujjassim, N.E.; Mallik, M.; Rathod, N.K.K.; Nitesh, S.D. Cisgenesis and intragenesis a new tool for conventional plant breeding: A review. J. Pharmacogn. Phytochem. 2019, 8, 2485–2489. [Google Scholar]
- Dreher, K.; Morris, M.; Khairallah, M.; Ribaut, J.M.; Shivaji, P.; Ganesan, S. Is marker-assisted selection cost-effective compared with conventional plant breeding methods? The case of quality protein Maize. Econ. Soc. Issues Agric. Biotechnol. 2009, 203–236. [Google Scholar]
- Abreu, G.B.; Ramalho, M.A.P.; Toledo, F.H.R.B.; De Souza, J.C. Strategies to improve mass selection in maize. Maydica 2010, 55, 219–225. [Google Scholar]
- Kandemir, N.; Saygili, İ. Apomixis: New horizons in plant breeding. Turkish J. Agric. For. 2015, 39, 549–556. [Google Scholar] [CrossRef]
- Leifert, C.; Tamm, L.; Lammerts van Bueren, E.T.; Jones, S.S.; Murphy, K.M.; Myers, J.R.; Messmer, M.M. The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: A review. NJAS - Wageningen J. Life Sci. 2011, 58, 193–205. [Google Scholar]
- Doust, A.; Diao, X. Plant Genetics and Genomics: Crops and Models Volume 19. Genet. Genom. Setaria Ser. 2017, 19, 377. [Google Scholar]
- Stetter, M.G.; Zeitler, L.; Steinhaus, A.; Kroener, K.; Biljecki, M.; Schmid, K.J. Crossing Methods and Cultivation Conditions for Rapid Production of Segregating Populations in Three Grain Amaranth Species. Front. Plant Sci. 2016, 7, 816. [Google Scholar] [CrossRef] [PubMed]
- Bugbee, B.; Koerner, G. Yield comparisons and unique characteristics of the dwarf wheat cultivar “USU-Apogee”. Adv. Sp. Res. 1997, 20, 1891–1894. [Google Scholar] [CrossRef]
- Bula, R.J.; Morrow, R.C.; Tibbitts, T.W.; Barta, D.J.; Ignatius, R.W.; Martin, T.S. Light-emitting diodes as a radiation source for plants. HortScience 1991, 26, 203–205. [Google Scholar] [CrossRef]
- Ghosh, S.; Watson, A.; Gonzalez-Navarro, O.E.; Ramirez-Gonzalez, R.H.; Yanes, L.; Mendoza-Suárez, M.; Simmonds, J.; Wells, R.; Rayner, T.; Green, P.; et al. Speed breeding in growth chambers and glasshouses for crop breeding and model plant research. Nat. Protoc. 2018, 13, 2944–2963. [Google Scholar] [CrossRef]
- Hickey, L.T.; Germa, S.E.; Diaz, J.E.; Ziems, L.A.; Fowler, R.A.; Platz, G.J.; Franckowiak, J.D.; Dieters, M.J. Speed Breeding for Multiple Disease Resistance in Barley; Springer: New York, NY, USA, 2017. [Google Scholar]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu. Rev. Plant Biol. 2019, 70, annurev. [Google Scholar] [CrossRef]
- Godwin, I.D.; Rutkoski, J.; Varshney, R.K.; Hickey, L.T. Technological perspectives for plant breeding. Theor. Appl. Genet. 2019, 132, 555–557. [Google Scholar] [CrossRef]
- Lee, J.; Chung, J.H.; Kim, H.M.; Kim, D.W.; Kim, H. Designed nucleases for targeted genome editing. Plant Biotechnol. J. 2016, 14, 448–462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, J.; Lang, Z.; Botella, J.R.; Zhu, J.K. Genome Editing—Principles and Applications for Functional Genomics Research and Crop Improvement. Crit. Rev. Plant Sci. 2017, 36, 291–309. [Google Scholar] [CrossRef]
- Hsu, P.D.; Scott, D.A.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.; Fine, E.J.; Wu, X.; Shalem, O.; et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2013, 3, 827. [Google Scholar] [CrossRef] [PubMed]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; Van Der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015. [Google Scholar] [CrossRef]
- Basey, A.C.; Fant, J.B.; Kramer, A.T. Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Nativ. Plants J. 2015, 16, 37–53. [Google Scholar] [CrossRef]
- Krimsky, S. Traditional Plant Breeding. In GMOs Decoded; MIT Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Shepard, J.F.; Bidney, D.; Barsby, T.; Kemble, R. Fusion of Protoplasts. Biotechnol. Biol. Front. 2019. [Google Scholar]
- Marthe, F. Tissue culture approaches in relation to medicinal plant improvement. In Biotechnologies of Crop Improvement; Research Gate: Berlin, Germany, 2018; Volume 1, pp. 487–497. ISBN 9783319782836. [Google Scholar]
- Germana, M.A. Anther culture for haploid and doubled haploid production. Plant Cell Tissue Organ Cult. 2011, 104, 283–300. [Google Scholar] [CrossRef]
- Hajjar, R.; Hodgkin, T. The use of wild relatives in crop improvement: A survey of developments over the last 20 years. Euphytica 2007, 156, 1–13. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Guimaraes, E.P.; Weltzien, E. Plant breeding and farmer participation; NHBS: Devon, UK, 2009; ISBN 9789251063828. [Google Scholar]
- Cellini, F.; Chesson, A.; Colquhoun, I.; Constable, A.; Davies, H.V.; Engel, K.H.; Gatehouse, A.M.R.; Kärenlampi, S.; Kok, E.J.; Leguay, J.-J. Unintended effects and their detection in genetically modified crops. Food Chem. Toxicol. 2004, 42, 1089–1125. [Google Scholar] [CrossRef]
- Dolferus, R.; Ji, X.; Richards, R.A. Abiotic stress and control of grain number in cereals. Plant Sci. 2011, 181, 331–341. [Google Scholar] [CrossRef]
- Caligari, P.D.S.; Brown, J. Plant Breeding, Practice. In Encyclopedia of Applied Plant Sciences; Academic Press: Cambridge, MA, USA, 2016; Volume 2, pp. 229–235. ISBN 9780123948083. [Google Scholar]
- Samineni, S.; Sen, M.; Sajja, S.B.; Gaur, P.M. Rapid generation advance (RGA) in chickpea to produce up to seven generations per year and enable speed breeding. Crop J. 2019. [Google Scholar] [CrossRef]
- Ossowski, S.; Schneeberger, K.; Lucas-Lledó, J.I.; Warthmann, N.; Clark, R.M.; Shaw, R.G.; Weigel, D.; Lynch, M. The rate and molecular spectrum of spontaneous mutations in Arabidopsis thaliana. Science (80-. ). 2010, 327, 92–94. [Google Scholar] [CrossRef] [PubMed]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; Hussin, G.; Ramli, A.; Rahim, H.A.; Miah, G.; Usman, M. Principle and application of plant mutagenesis in crop improvement: A review. Biotechnol. Biotechnol. Equip. 2016, 30, 1–16. [Google Scholar] [CrossRef]
- Roychowdhury, R.; Tah, J. Mutagenesis—A potential approach for crop improvement. In Crop Improvement; Springer: New York, NY, USA, 2013; pp. 149–187. [Google Scholar]
- Forster, B.P.; Shu, Q.Y.; Nakagawa, H. Plant mutagenesis in crop improvement: Basic terms and applications. Plant Mutat. Breed. Biotechnol. 2012, 9–20. [Google Scholar]
- Van Harten, A.M. Mutation Breeding: Theory and Practical Applications; Cambridge University Press: Cambridge, MA, USA, 1998; ISBN 0521470749. [Google Scholar]
- Kharkwal, M.C. A brief history of plant mutagenesis. Plant Mutat. Breed. Biotechnol. 2012, 21–30. [Google Scholar]
- Mba, C.; Afza, R.; Bado, S.; Jain, S.M. Induced Mutagenesis in Plants. Plant Cell Cult. Essent. Methods 2010, 111–130. [Google Scholar]
- Mba, C. Induced Mutations Unleash the Potentials of Plant Genetic Resources for Food and Agriculture. Agronomy 2013, 3, 200–231. [Google Scholar] [CrossRef]
- Menda, N.; Semel, Y.; Peled, D.; Eshed, Y.; Zamir, D. In silico screening of a saturated mutation library of tomato. Plant J. 2004, 38, 861–872. [Google Scholar] [CrossRef]
- Watanabe, S.; Mizoguchi, T.; Aoki, K.; Kubo, Y.; Mori, H.; Imanishi, S.; Yamazaki, Y.; Shibata, D.; Ezura, H. Ethylmethanesulfonate (EMS) mutagenesis of Solanum lycopersicum cv. Micro-Tom for large-scale mutant screens. Plant Biotechnol. 2007, 24, 33–38. [Google Scholar] [CrossRef]
- Wani, M.R.; Kozgar, M.I.; Tomlekova, N.; Khan, S.; Kazi, A.G.; Sheikh, S.A.; Ahmad, P. Mutation breeding: A novel technique for genetic improvement of pulse crops particularly Chickpea (Cicer arietinum L.). In Improvement of Crops in the Era of Climatic Changes; Springer: New York, NY, USA, 2014; pp. 217–248. [Google Scholar]
- Mba, C.; Afza, R.; Shu, Q.Y.; Forster, B.P.; Nakagawa, H. Mutagenic radiations: X-rays, ionizing particles and ultraviolet. Plant Mutat. Breed. Biotechnol. 2012, 83–90. [Google Scholar]
- Acreche, M.M.; Briceño-Félix, G.; Sánchez, J.A.M.; Slafer, G.A. Physiological bases of genetic gains in Mediterranean bread wheat yield in Spain. Eur. J. Agron. 2008, 28, 162–170. [Google Scholar] [CrossRef]
- Sadras, V.O.; Lawson, C. Genetic gain in yield and associated changes in phenotype, trait plasticity and competitive ability of South Australian wheat varieties released between 1958 and 2007. Crop Pasture Sci. 2011, 62, 533–549. [Google Scholar] [CrossRef]
- Araus, J.L.; Cairns, J.E. Field high-throughput phenotyping: The new crop breeding frontier. Trends Plant Sci. 2014, 19, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, F.; Cabrera-Bosquet, L.; Pridmore, T.; Bennett, M. Plant Phenomics, From Sensors to Knowledge. Curr. Biol. 2017, 27, R770–R783. [Google Scholar] [CrossRef] [PubMed]
- Crossa, J.; Pérez, P.; de los Campos, G.; Mahuku, G.; Dreisigacker, S.; Magorokosho, C. Genomic selection and prediction in plant breeding. J. Crop Improv. 2011, 25, 239–261. [Google Scholar] [CrossRef]
- Blum, A. Genomics for drought resistance-getting down to earth. In Functional Plant Biology; CSIRO Publishing: Melbourne, Australia, 2014; Volume 41, pp. 1191–1198. [Google Scholar]
- Deery, D.; Jimenez-Berni, J.; Jones, H.; Sirault, X.; Furbank, R. Proximal remote sensing buggies and potential applications for field-based phenotyping. Agronomy 2014, 4, 349–379. [Google Scholar] [CrossRef]
- White, J.W.; Conley, M.M. A flexible, low-cost cart for proximal sensing. Crop Sci. 2013, 53, 1646–1649. [Google Scholar] [CrossRef]
- Saxena, K.; Saxena, R.K.; Varshney, R.K. Use of immature seed germination and single seed descent for rapid genetic gains in pigeonpea. Plant Breed. 2017, 136, 954–957. [Google Scholar] [CrossRef]
- Shakoor, N.; Lee, S.; Mockler, T.C. High throughput phenotyping to accelerate crop breeding and monitoring of diseases in the field. Curr. Opin. Plant Biol. 2017, 38, 184–192. [Google Scholar] [CrossRef]
- Janila, P.; Variath, M.T.; Pandey, M.K.; Desmae, H.; Motagi, B.N.; Okori, P.; Manohar, S.S.; Rathnakumar, A.L.; Radhakrishnan, T.; Liao, B.; et al. Genomic tools in groundnut breeding program: Status and perspectives. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Sarutayophat, T.; Nualsri, C. The efficiency of pedigree and single seed descent selections for yield improvement at generation 4 (F4) of two yardlong bean populations. Kasetsart J. Nat. Sci. 2010, 44, 343–352. [Google Scholar]
- Holbrook, C.C.; Timper, P.; Culbreath, A.K.; Kvien, C.K. Registration of “Tifguard” Peanut. J. Plant Regist. 2008, 2, 92. [Google Scholar] [CrossRef]
- Huang, X. From Genetic Mapping to Molecular Breeding: Genomics Have Paved the Highway. Mol. Plant 2016. [Google Scholar] [CrossRef] [PubMed]
- Jung, C. Chapter 3 Molecular Tools for Plant Breeding; Springer: New York, NY, USA, 2000; pp. 25–37. [Google Scholar]
- Schaart, J.G.; van de Wiel, C.C.M.; Lotz, L.A.P.; Smulders, M.J.M. Opportunities for Products of New Plant Breeding Techniques. Trends Plant Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, S.; Cañizares, J.; Pascual, L.; Blanca, J.M.; Díez, M.J.; Prohens, J.; Picó, B. Application of Genomic Tools in Plant Breeding. Curr. Genomics 2012, 13, 179–195. [Google Scholar]
- Wendler, N.; Mascher, M.; Nöh, C.; Himmelbach, A.; Scholz, U.; Ruge-Wehling, B.; Stein, N. Unlocking the secondary gene-pool of barley with next-generation sequencing. Plant Biotechnol. J. 2014, 12, 1122–1131. [Google Scholar] [CrossRef]
- Metzker, M.L. Sequencing technologies—The next generation. Nat. Rev. Genet. 2010, 11, 31. [Google Scholar] [CrossRef]
- Dwivedi, S.; Perotti, E.; Ortiz, R. Towards molecular breeding of reproductive traits in cereal crops. Plant Biotechnol. J. 2008, 6, 529–559. [Google Scholar] [CrossRef]
- Wang, T.L.; Uauy, C.; Robson, F.; Till, B. TILLING in extremis. Plant Biotechnol. J. 2012, 10, 761–772. [Google Scholar] [CrossRef]
- Zou, C.; Wang, P.; Xu, Y. Bulked sample analysis in genetics, genomics and crop improvement. Plant Biotechnol. J. 2016, 14, 1941–1955. [Google Scholar] [CrossRef]
- Bolger, M.E.; Weisshaar, B.; Scholz, U.; Stein, N.; Usadel, B.; Mayer, K.F.X. Plant genome sequencing—Applications for crop improvement. Curr. Opin. Biotechnol. 2014, 26, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.; Batley, J. Plant genome sequencing: Applications for crop improvement. Plant Biotechnol. J. 2010, 8, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Dhingani, R.M.; Umrania, V.V.; Tomar, R.S.; Parakhia, M.V.; Golakiya, B. Introduction to QTL mapping in plants. Ann. Plant Sci 2015, 4, 1072–1079. [Google Scholar]
- McCallum, C.M.; Comai, L.; Greene, E.A.; Henikoff, S. Targeted screening for induced mutations. Nat. Biotechnol. 2000, 18, 455. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoğlu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N.; et al. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2018, 32, 261–285. [Google Scholar] [CrossRef]
- Lloyd, A.; Plaisier, C.L.; Carroll, D.; Drews, G.N. Targeted mutagenesis using zinc-finger nucleases in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005. [Google Scholar] [CrossRef]
- Symington, L.S.; Gautier, J. Double-strand break end resection and repair pathway choice. Annu. Rev. Genet. 2011, 45, 247–271. [Google Scholar] [CrossRef]
- Wood, A.J.; Lo, T.W.; Zeitler, B.; Pickle, C.S.; Ralston, E.J.; Lee, A.H.; Amora, R.; Miller, J.C.; Leung, E.; Meng, X.; et al. Targeted genome editing across species using ZFNs and TALENs. Science 2011, 333, 307. [Google Scholar] [CrossRef]
- Sprink, T.; Metje, J.; Hartung, F. Plant genome editing by novel tools: TALEN and other sequence specific nucleases. Curr. Opin. Biotechnol. 2015. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, H.; Xu, N.; Zhang, B.; Gou, F.; Zhu, J.K. Application of the CRISPR-Cas system for efficient genome engineering in plants. Mol. Plant 2013. [Google Scholar] [CrossRef]
- Schneider, K.; Schiermeyer, A.; Dolls, A.; Koch, N.; Herwartz, D.; Kirchhoff, J.; Fischer, R.; Russell, S.M.; Cao, Z.; Corbin, D.R. Targeted gene exchange in plant cells mediated by a zinc finger nuclease double cut. Plant Biotechnol. J. 2016, 14, 1151–1160. [Google Scholar] [CrossRef] [PubMed]
- De Pater, S.; Pinas, J.E.; Hooykaas, P.J.J.; van der Zaal, B.J. ZFN-mediated gene targeting of the Arabidopsis protoporphyrinogen oxidase gene through Agrobacterium-mediated floral dip transformation. Plant Biotechnol. J. 2013, 11, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Huang, S.; Jiang, W.Z.; Wright, D.; Spalding, M.H.; Weeks, D.P.; Yang, B. TAL nucleases (TALNs): Hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain. Nucleic Acids Res. 2010, 39, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Char, S.N.; Unger-Wallace, E.; Frame, B.; Briggs, S.A.; Main, M.; Spalding, M.H.; Vollbrecht, E.; Wang, K.; Yang, B. Heritable site-specific mutagenesis using TALENs in maize. Plant Biotechnol. J. 2015, 13, 1002–1010. [Google Scholar] [CrossRef]
- Mahfouz, M.M.; Piatek, A.; Stewart, C.N. Genome engineering via TALENs and CRISPR/Cas9 systems: Challenges and perspectives. Plant Biotechnol. J. 2014, 12, 1006–1014. [Google Scholar] [CrossRef]
- Zhang, Y.; Xie, X.; Liu, Y.G.; Zhang, Y.; Xie, X.; Liu, Y.G.; Ma, X. CRISPR/Cas9-Based Genome Editing in Plants, 1st ed; Elsevier Inc.: Amsterdam, The Netherlands, 2017; Volume 149. [Google Scholar]
- Zaidi, S.S.-e.-A.; Mahfouz, M.M.; Mansoor, S. CRISPR-Cpf1: A New Tool for Plant Genome Editing. Trends Plant Sci. 2017, 22, 550–553. [Google Scholar] [CrossRef]
- Li, S.; Zhang, X.; Wang, W.; Guo, X.; Wu, Z.; Du, W.; Zhao, Y.; Xia, L. Expanding the scope of CRISPR/Cpf1-mediated genome editing in rice. Mol. Plant 2018, 11, 995–998. [Google Scholar] [CrossRef]
- Qin, L.; Li, J.; Wang, Q.; Xu, Z.; Sun, L.; Alariqi, M.; Manghwar, H.; Wang, G.; Li, B.; Ding, X.; et al. High Efficient and Precise Base Editing of C•G to T•A in the Allotetraploid Cotton (Gossypium hirsutum) Genome Using a Modified CRISPR /Cas9 System. Plant Biotechnol. J. 2020, 18, 45–56. [Google Scholar] [CrossRef]
- Bortesi, L.; Zhu, C.; Zischewski, J.; Perez, L.; Bassié, L.; Nadi, R.; Forni, G.; Lade, S.B.; Soto, E.; Jin, X.; et al. Patterns of CRISPR/Cas9 activity in plants, animals and microbes. Plant Biotechnol. J. 2016, 14, 2203–2216. [Google Scholar] [CrossRef]
- Lee, K.; Zhang, Y.; Kleinstiver, B.P.; Guo, J.A.; Aryee, M.J.; Miller, J.; Malzahn, A.; Zarecor, S.; Lawrence-Dill, C.J.; Joung, J.K.; et al. Activities and specificities of CRISPR/Cas9 and Cas12a nucleases for targeted mutagenesis in maize. Plant Biotechnol. J. 2019, 17, 362–372. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, Z.; Zong, Y.; Wang, Y.; Liu, J.; Chen, K.; Qiu, J.L.; Gao, C. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 2016. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science (80-. ). 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Wang, X.; Tu, M.; Wang, D.; Liu, J.; Li, Y.; Li, Z.; Wang, Y.; Wang, X. CRISPR/Cas9-mediated efficient targeted mutagenesis in grape in the first generation. Plant Biotechnol. J. 2018, 16, 844–855. [Google Scholar] [CrossRef] [PubMed]
- Murugan, K.; Babu, K.; Sundaresan, R.; Rajan, R.; Sashital, D.G. The Revolution Continues: Newly Discovered Systems Expand the CRISPR-Cas Toolkit. Mol. Cell 2017, 68, 15–25. [Google Scholar] [CrossRef]
- Ma, X.; Chen, X.; Jin, Y.; Ge, W.; Wang, W.; Kong, L.; Ji, J.; Guo, X.; Huang, J.; Feng, X.H.; et al. Small molecules promote CRISPR-Cpf1-mediated genome editing in human pluripotent stem cells. Nat. Commun. 2018. [Google Scholar] [CrossRef]
- Riesenberg, S.; Maricic, T. Targeting repair pathways with small molecules increases precise genome editing in pluripotent stem cells. Nat. Commun. 2018. [Google Scholar] [CrossRef]
- Yang, F.; Li, Y. The new generation tool for CRISPR genome editing: CRISPR/Cpf1. Sheng wu gong cheng xue bao= Chinese J. Biotechnol. 2017, 33, 361–371. [Google Scholar]
- Maruyama, T.; Dougan, S.K.; Truttmann, M.C.; Bilate, A.M.; Ingram, J.R.; Ploegh, H.L. Corrigendum: Increasing the efficiency of precise genome editing with CRISPR-Cas9 by inhibition of nonhomologous end joining. Nat. Biotechnol. 2016, 34, 210. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Araki, M. A future scenario of the global regulatory landscape regarding genome-edited crops. GM Crop. Food 2017, 8, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Wolt, J.D.; Wang, K.; Yang, B. The Regulatory Status of Genome-edited Crops. Plant Biotechnol. J. 2016, 14, 510–518. [Google Scholar] [CrossRef]
- Callaway, E. CRISPR plants now subject to tough GM laws in European Union. Nature 2018, 560, 16. [Google Scholar] [CrossRef] [PubMed]
- Sprink, T.; Eriksson, D.; Schiemann, J.; Hartung, F. Regulatory hurdles for genome editing: Process- vs. product-based approaches in different regulatory contexts. Plant Cell Rep. 2016, 35, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Slama-Ayed, O.; Bouhaouel, I.; Ayed, S.; De Buyser, J.; Picard, E.; Amara, H.S. Efficiency of three haplomethods in durum wheat (Triticum turgidum subsp. durum Desf.): Isolated microspore culture, gynogenesis and wheat× maize crosses. Czech J. Genet. Plant Breed. 2019, 55, 101–109. [Google Scholar] [CrossRef]
- Chiurugwi, T.; Kemp, S.; Powell, W.; Hickey, L.T.; Powell, W. Speed breeding orphan crops. Theor. Appl. Genet. 2018. [Google Scholar] [CrossRef] [PubMed]
- Weller, J.L.; Beauchamp, N.; Kerckhoffs, L.H.J.; Platten, J.D.; Reid, J.B. Interaction of phytochromes A and B in the control of de-etiolation and flowering in pea. Plant J. 2001, 26, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Giliberto, L.; Perrotta, G.; Pallara, P.; Weller, J.L.; Fraser, P.D.; Bramley, P.M.; Fiore, A.; Tavazza, M.; Giuliano, G. Manipulation of the blue light photoreceptor cryptochrome 2 in tomato affects vegetative development, flowering time, and fruit antioxidant content. Plant Physiol. 2005, 137, 199–208. [Google Scholar] [CrossRef]
- Croser, J.S.; Pazos-Navarro, M.; Bennett, R.G.; Tschirren, S.; Edwards, K.; Erskine, W.; Creasy, R.; Ribalta, F.M. Time to flowering of temperate pulses in vivo and generation turnover in vivo–in vitro of narrow-leaf lupin accelerated by low red to far-red ratio and high intensity in the far-red region. Plant Cell. Tissue Organ Cult. 2016, 127, 591–599. [Google Scholar] [CrossRef]
- Ribalta, F.M.; Pazos-Navarro, M.; Nelson, K.; Edwards, K.; Ross, J.J.; Bennett, R.G.; Munday, C.; Erskine, W.; Ochatt, S.J.; Croser, J.S. Precocious floral initiation and identification of exact timing of embryo physiological maturity facilitate germination of immature seeds to truncate the lifecycle of pea. Plant Growth Regul. 2017, 81, 345–353. [Google Scholar] [CrossRef]
- Moe, R.; Heins, R. Control Of plant morphogenesis and flowering by light quality and temperature. Acta Hortic. 1990, 81–90. [Google Scholar] [CrossRef]
- Ausín, I.; Alonso-Blanco, C.; Martínez-Zapater, J.M. Environmental regulation of flowering. Int. J. Dev. Biol. 2005, 49, 689–705. [Google Scholar] [CrossRef]
- Ochatt, S.J.; Sangwan, R.S.; Marget, P.; Assoumou Ndong, Y.; Rancillac, M.; Perney, P. New approaches towards the shortening of generation cycles for faster breeding of protein legumes. Plant Breed. 2002, 121, 436–440. [Google Scholar] [CrossRef]
- Ochatt, S.J.; Sangwan, R.S. In vitro shortening of generation time in Arabidopsis thaliana. Plant Cell. Tissue Organ Cult. 2008, 93, 133–137. [Google Scholar] [CrossRef]
- Heuschele, D.J.; Case, A.; Smith, K.P. Evaluation of Fast Generation Cycling in Oat (Avena sativa). Cereal Res. Commun. 2019, 47, 626–635. [Google Scholar] [CrossRef]
- Yao, Y.; Zhang, P.; Liu, H.; Lu, Z.; Yan, G. A fully in vitro protocol towards large scale production of recombinant inbred lines in wheat (Triticum aestivum L.). Plant Cell. Tissue Organ Cult. 2017, 128, 655–661. [Google Scholar] [CrossRef]
- Sysoeva, M.I.; Markovskaya, E.F.; Shibaeva, T.G. Plants under continuous light: A review. Plant Stress 2010, 4, 5–17. [Google Scholar]
- Achigan-Dako, E.G.; Sogbohossou, O.E.; Maundu, P. Current knowledge on Amaranthus spp.: research avenues for improved nutritional value and yield in leafy amaranths in sub-Saharan Africa. Euphytica 2014, 197, 303–317. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Beredo, J.C.; Lenaerts, B.; Mendoza, R.; Santelices, R.; Lopena, V.; Verdeprado, H.; Raghavan, C.; Gregorio, G.B.; Vial, L.; et al. Revisiting rice breeding methods–evaluating the use of rapid generation advance (RGA) for routine rice breeding. Plant Prod. Sci. 2017, 20, 337–352. [Google Scholar] [CrossRef]
- Wolter, F.; Schindele, P.; Puchta, H. Plant breeding at the speed of light: The power of CRISPR/Cas to generate directed genetic diversity at multiple sites. BMC Plant Biol. 2019, 19, 1–8. [Google Scholar] [CrossRef]
- Bartley, G. Wheat (Triticum aestivum) residue management before growing soybean (Glycine max) in Manitoba. Master’s Thesis, Department of Plant Science, University of Manitoba, Winnipeg, Manitoba, 2019. [Google Scholar]
- Wilton, M. A Broad-Scale Characterization of Corn (Zea mays)-Soybean (Glycine max) Intercropping as a Sustainable-Intensive Cropping Practice. Ph.D. Thesis, University of Waterloo, Ontario, Canada, 2019. [Google Scholar]
- Borlaug, N.E. Sixty-two years of fighting hunger: Personal recollections. Euphytica 2007, 157, 287–297. [Google Scholar] [CrossRef]
- Ferrie, A.M.R.; Möllers, C. Haploids and doubled haploids in Brassica spp. for genetic and genomic research. Plant Cell Tissue Organ. Cult. 2011, 104, 375–386. [Google Scholar] [CrossRef]
- Lübberstedt, T.; Frei, U.K. Application of doubled haploids for target gene fixation in backcross programmes of maize. Plant Breed. 2012, 131, 449–452. [Google Scholar] [CrossRef]
- Dirks, R.; Van Dun, K.; De Snoo, C.B.; Van Den Berg, M.; Lelivelt, C.L.C.; Voermans, W.; Woudenberg, L.; De Wit, J.P.C.; Reinink, K.; Schut, J.W.; et al. Reverse breeding: A novel breeding approach based on engineered meiosis. Plant Biotechnol. J. 2009, 7, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Kebede, A.Z.; Dhillon, B.S.; Schipprack, W.; Araus, J.L.; Bänziger, M.; Semagn, K.; Alvarado, G.; Melchinger, A.E. Effect of source germplasm and season on the in vivo haploid induction rate in tropical maize. Euphytica 2011, 180, 219–226. [Google Scholar] [CrossRef]
- Castillo, A.M.; Cistué, L.; Vallés, M.P.; Soriano, M. Chromosome Doubling in Monocots. In Advances in Haploid Production in Higher Plants; Springer: New York, NY, USA, 2009; pp. 329–338. [Google Scholar]
- Prigge, V.; Melchinger, A.E. Production of haploids and doubled haploids in maize. In Plant Cell Culture Protocols; Springer: New York, NY, USA, 2012; pp. 161–172. [Google Scholar]
- Raju, S.K.K.; Shao, M.R.; Sanchez, R.; Xu, Y.Z.; Sandhu, A.; Graef, G.; Mackenzie, S. An epigenetic breeding system in soybean for increased yield and stability. Plant Biotechnol. J. 2018, 16, 1836–1847. [Google Scholar] [CrossRef]
- Halpin, C. Gene stacking in transgenic plants—The challenge for 21st century plant biotechnology. Plant Biotechnol. J. 2005, 3, 141–155. [Google Scholar] [CrossRef]
- Belhaj, K.; Chaparro-Garcia, A.; Kamoun, S.; Nekrasov, V. Plant genome editing made easy: Targeted mutagenesis in model and crop plants using the CRISPR/Cas system. Plant Methods 2013. [Google Scholar] [CrossRef]
- Low, J.W.; Mwanga, R.O.M.; Andrade, M.; Carey, E.; Ball, A.-M. Tackling vitamin A deficiency with biofortified sweetpotato in sub-Saharan Africa. Glob. Food Sec. 2017, 14, 23–30. [Google Scholar] [CrossRef]
- Møller, I.S.; Gilliham, M.; Jha, D.; Mayo, G.M.; Roy, S.J.; Coates, J.C.; Haseloff, J.; Tester, M. Shoot Na+ exclusion and increased salinity tolerance engineered by cell type–specific alteration of Na+ transport in Arabidopsis. Plant Cell 2009, 21, 2163–2178. [Google Scholar] [CrossRef]
- Singh, N.K.; Gupta, D.K.; Jayaswal, P.K.; Mahato, A.K.; Dutta, S.; Singh, S.; Bhutani, S.; Dogra, V.; Singh, B.P.; Kumawat, G.; et al. The first draft of the pigeonpea genome sequence. J. Plant Biochem. Biotechnol. 2012, 21, 98–112. [Google Scholar] [CrossRef]
- Jackson, S.A. Rice: The First Crop Genome. Rice 2016, 9. [Google Scholar] [CrossRef]
- Egan, A.N.; Schlueter, J.; Spooner, D.M. Applications of next-generation sequencing in plant biology. Am. J. Bot. 2012, 99, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Bernier, G.; Périlleux, C. A physiological overview of the genetics of flowering time control. Plant Biotechnol. J. 2005, 3, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Kondić-špika, A.; Kobiljski, B. Biotechnology in Modern Breeding and Agriculture. In Proceedings of the International Conference on BioScience: Biotechnology and Biodiversity-Step in the Future. The Fourth Joint UNS-PSU Conference, Novi Sad, Serbia, 18–20 June 2012. [Google Scholar]
- Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P. The genes of the Green Revolution. TRENDS Genet. 2003, 19, 5–9. [Google Scholar] [CrossRef]
- Knoch, D.; Abbadi, A.; Grandke, F.; Meyer, R.C.; Samans, B.; Werner, C.R.; Snowdon, R.J.; Altmann, T. Strong temporal dynamics of QTL action on plant growth progression revealed through high-throughput phenotyping in canola. Plant Biotechnol. J. 2020, 18, 68–82. [Google Scholar] [CrossRef]
- Yin, K.; Gao, C.; Qiu, J.L. Progress and prospects in plant genome editing. Nat. Plants 2017, 3, 1–6. [Google Scholar] [CrossRef]
- Waltz, E. CRISPR-Edited Crops Free to Enter Market, Skip Regulation; Nature Publishing Group: Berlin, Germany, 2016. [Google Scholar]
- Eş, I.; Gavahian, M.; Marti-Quijal, F.J.; Lorenzo, J.M.; Mousavi Khaneghah, A.; Tsatsanis, C.; Kampranis, S.C.; Barba, F.J. The application of the CRISPR-Cas9 genome editing machinery in food and agricultural science: Current status, future perspectives, and associated challenges. Biotechnol. Adv. 2019. [Google Scholar] [CrossRef]
- Xu, R.; Yang, Y.; Qin, R.; Li, H.; Qiu, C.; Li, L.; Wei, P.; Yang, J. Rapid improvement of grain weight via highly efficient CRISPR/Cas9-mediated multiplex genome editing in rice. J. Genet. Genomics 2016, 43, 529–532. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.; Botella, J.R.; Zhu, J. Generation of new glutinous rice by CRISPR/Cas9-targeted mutagenesis of the Waxy gene in elite rice varieties. J. Integr. Plant Biol. 2018, 60, 369–375. [Google Scholar] [CrossRef]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion. Sci. Rep. 2017. [Google Scholar] [CrossRef]
- Wang, F.; Wang, C.; Liu, P.; Lei, C.; Hao, W.; Gao, Y.; Liu, Y.G.; Zhao, K. Enhanced rice blast resistance by CRISPR/ Cas9-Targeted mutagenesis of the ERF transcription factor gene OsERF922. PLoS ONE 2016. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Aslam, M.; Manisha, Y.; Manoj, N. An Update on Genetic Modification of Chickpea for Increased Yield and Stress Tolerance. Mol. Biotechnol. 2018, 60, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Domoney, C.; Knox, M.; Moreau, C.; Ambrose, M.; Palmer, S.; Smith, P.; Christodoulou, V.; Isaac, P.G.; Hegarty, M.; Blackmore, T.; et al. Exploiting a fast neutron mutant genetic resource in Pisum sativum (pea) for functional genomics. Funct. Plant Biol. 2013, 40, 1261. [Google Scholar] [CrossRef]
- Raman, H.; Raman, R.; Kilian, A.; Detering, F.; Carling, J.; Coombes, N.; Diffey, S.; Kadkol, G.; Edwards, D.; Mccully, M.; et al. Genome-Wide Delineation of Natural Variation for Pod Shatter Resistance in Brassica napus. PLoS ONE 2014, 9, e101673. [Google Scholar] [CrossRef] [PubMed]
- Panigrahi, R.; Kariali, E.; Panda, B.; Lafarge, T.; Mohapatra, P.K. Controlling the trade-off between spikelet number and grain filling; the hierarchy of starch synthesis in spikelets of rice panicle in relation to hormone dynamics. Funct. Plant Biol. 2019, 46, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Witcombe, J.R.; Gyawali, S.; Subedi, M.; Virk, D.S.; Joshi, K.D. Plant breeding can be made more efficient by having fewer, better crosses. BMC Plant Biol. 2013, 13, 22. [Google Scholar] [CrossRef]
- Basnet, B.R.; Crossa, J.; Dreisigacker, S.; Perez-Rodriguez, P.; Manes, Y.; Singh, R.P.; Rosyara, U.R.; Camarillo-Castillo, F.; Murua, M. Hybrid Wheat Prediction Using Genomic, Pedigree, and Environmental Covariables Interaction Models. Plant Genome 2019, 12. [Google Scholar] [CrossRef]
- Xu, Y.; Babu, R.; Skinner, D.J.; Vivek, B.S.; Crouch, J.H. Maize Mutant opaque2 and the Improvement of Protein Quality through Conventional and Molecular Approaches. In Proceedings of the International Symposium on Induced Mutation in Plants, Vienna, Austria, 2–15 August 2008. [Google Scholar]
- Shuwen, S.; Lianghong, L.; Jiangsheng, W.; Yongming, Z. In vitro screening stem rot resistant ( tolerant ) materials in Brassica napus L. Chin. J. Oil Crop Sci. 2003, 25, 5–8. [Google Scholar]
- Pathirana, R. Plant mutation breeding in agriculture. Plant Sci. Rev. 2011, 107–126. [Google Scholar] [CrossRef]
- International Atomic Energy Agency. Proceedings of the International Symposium on Plant Mutation Breeding and Biotechnology, Vienna, Austria, 27–31 August 2018.
- Ceballos, H.; Sanchez, T.; Denyer, K.; Tofino, A.P.; Rosero, E.A.; Dufour, D.; Smith, A.; Morante, N.; Perez, J.C.; Fahy, B. Induction and identification of a small-granule, high-amylose mutant in cassava (Manihot esculenta Crantz). J. Agric. Food Chem. 2008, 56, 7215–7222. [Google Scholar] [CrossRef]
- Hamid, M.A.; Azad, M.A.K.; Howelider, M.A.R. Development of Three Groundnut Varieties with Improved Quantitative and Qualitative Traits through Induced Mutation. Plant Mutat. reports 2006, 1, 14–16. [Google Scholar]
- Newell-McGloughlin, M. Nutritionally improved agricultural crops. Plant Physiol. 2008, 147, 939–953. [Google Scholar] [CrossRef] [PubMed]
- Dunwell, J.M. Transgenic approaches to crop improvement. J. Exp. Bot. 2000, 51, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Toojinda, T.; Baird, E.; Booth, A.; Broers, L.; Hayes, P.; Powell, W.; Thomas, W.; Vivar, H.; Young, G. Introgression of quantitative trait loci (QTLs) determining stripe rust resistance in barley: An example of marker-assisted line development. Theor. Appl. Genet. 1998, 96, 123–131. [Google Scholar] [CrossRef]
- Gimode, W.; Clevenger, J.; McGregor, C. Fine-mapping of a major quantitative trait locus Qdff3-1 controlling flowering time in watermelon. Mol. Breed. 2020, 40, 1–12. [Google Scholar] [CrossRef]
- Joshi, S.G.; Schaart, J.G.; Groenwold, R.; Jacobsen, E.; Schouten, H.J.; Krens, F.A. Functional analysis and expression profiling of HcrVf1 and HcrVf2 for development of scab resistant cisgenic and intragenic apples. Plant Mol. Biol. 2011, 75, 579–591. [Google Scholar] [CrossRef]
- Würdig, J.; Flachowsky, H.; Saß, A.; Peil, A.; Hanke, M.V. Improving resistance of different apple cultivars using the Rvi6 scab resistance gene in a cisgenic approach based on the Flp/FRT recombinase system. Mol. Breed. 2015, 35. [Google Scholar] [CrossRef]
- Holme, I.B.; Wendt, T.; Holm, P.B. Intragenesis and cisgenesis as alternatives to transgenic crop development. Plant Biotechnol. J. 2013, 11, 395–407. [Google Scholar] [CrossRef]
- Gadaleta, A.; Giancaspro, A.; Blechl, A.E.; Blanco, A. A transgenic durum wheat line that is free of marker genes and expresses 1Dy10. J. Cereal Sci. 2008, 48, 439–445. [Google Scholar] [CrossRef]
- Cardi, T. Cisgenesis and genome editing: Combining concepts and efforts for a smarter use of genetic resources in crop breeding. Plant Breed. 2016, 135, 139–147. [Google Scholar] [CrossRef]
- Jo, K.-R.; Kim, C.-J.; Kim, S.-J.; Kim, T.-Y.; Bergervoet, M.; Jongsma, M.A.; Visser, R.G.F.; Jacobsen, E.; Vossen, J.H. Development of late blight resistant potatoes by cisgene stacking. BMC Biotechnol. 2014, 14, 50. [Google Scholar] [CrossRef] [PubMed]
- Schaart, J.G. Towards Consumer-Friendly Cisgenic Strawberries which are Less Susceptible to Botrytis Cinerea; Research Gate: Berlin, Germany, 2004; ISBN 908504104X. [Google Scholar]
- Sawai, S.; Ohyama, K.; Yasumoto, S.; Seki, H.; Sakuma, T.; Yamamoto, T.; Takebayashi, Y.; Kojima, M.; Sakakibara, H.; Aoki, T.; et al. Sterol side chain reductase 2 is a key enzyme in the biosynthesis of cholesterol, the common precursor of toxic steroidal glycoalkaloids in potato. Plant Cell 2014, 26, 3763–3774. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Li, H.; Chakraborty, S.; Morbitzer, R.; Rinaldo, A.; Upadhyaya, N.; Bhatt, D.; Louis, S.; Richardson, T.; Lahaye, T.; et al. Efficient TALEN mediated gene editing in wheat. Plant Biotechnol. J. 2019, 17, 2026–2028. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Peng, Z.; Long, J.; Sosso, D.; Liu, B.; Eom, J.-S.; Huang, S.; Liu, S.; Vera Cruz, C.; Frommer, W.B.; et al. Gene targeting by the TAL effector PthXo2 reveals cryptic resistance gene for bacterial blight of rice. Plant J. 2015, 82, 632–643. [Google Scholar] [CrossRef] [PubMed]
- Svitashev, S.; Young, J.K.; Schwartz, C.; Gao, H.; Falco, S.C.; Cigan, A.M. Targeted Mutagenesis, Precise Gene Editing, and Site-Specific Gene Insertion in Maize Using Cas9 and Guide RNA. Plant Physiol. 2015, 169, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Jacobs, T.B.; Xue, L.-J.; Harding, S.A.; Tsai, C.-J. Exploiting SNPs for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-coumarate:CoA ligase specificity and redundancy. New Phytol. 2015, 208, 298–301. [Google Scholar] [CrossRef] [PubMed]
- Brooks, C.; Nekrasov, V.; Lippman, Z.B.; Van Eck, J. Efficient gene editing in tomato in the first generation using the clustered regularly interspaced short palindromic repeats/CRISPR-associated9 system. Plant Physiol. 2014, 166, 1292–1297. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Z.-B.; Xing, A.; Moon, B.P.; Koellhoffer, J.P.; Huang, L.; Ward, R.T.; Clifton, E.; Falco, S.C.; Cigan, A.M. Cas9-Guide RNA Directed Genome Editing in Soybean. Plant Physiol. 2015, 169, 960–970. [Google Scholar] [CrossRef]
- Zhao, X.; Meng, Z.; Wang, Y.; Chen, W.; Sun, C.; Cui, B.; Cui, J.; Yu, M.; Zeng, Z.; Guo, S.; et al. Pollen magnetofection for genetic modification with magnetic nanoparticles as gene carriers. Nat. Plants 2017, 3, 956–964. [Google Scholar] [CrossRef]
- Han, J.; Guo, B.; Guo, Y.; Zhang, B.; Wang, X.; Qiu, L.-J. Creation of Early Flowering Germplasm of Soybean by CRISPR/Cas9 Technology. Front. Plant Sci. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Sánchez-León, S.; Gil-Humanes, J.; Ozuna, C.V.; Giménez, M.J.; Sousa, C.; Voytas, D.F.; Barro, F. Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 2018, 16, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Farhat, S.; Jain, N.; Singh, N.; Sreevathsa, R.; Das, P.K.; Rai, R.; Yadav, S.; Kumar, P.; Sarkar, A.; Jain, A. CRISPR-cas 9 directed genome engineering for enhancing salt stress tolerance in rice. In Proceedings of the Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Zaman, Q.U.; Chu, W.; Hao, M.; Shi, Y.; Sun, M.; Sang, S.-F.; Mei, D.; Cheng, H.; Liu, J.; Li, C. CRISPR/Cas9-Mediated Multiplex Genome Editing of JAGGED Gene in Brassica napus L. Biomolecules 2019, 9, 725. [Google Scholar] [CrossRef] [PubMed]
- Veillet, F.; Perrot, L.; Chauvin, L.; Kermarrec, M.-P.; Guyon-Debast, A.; Chauvin, J.-E.; Nogué, F.; Mazier, M. Transgene-free genome editing in tomato and potato plants using agrobacterium-mediated delivery of a CRISPR/Cas9 cytidine base editor. Int. J. Mol. Sci. 2019, 20, 402. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-T.; Cheng, Y.-J.; Yuan, Y.-H.; Hung, W.-F.; Cheng, Q.-W.; Wu, F.-H.; Lee, L.-Y.; Gelvin, S.B.; Lin, C.-S. Application of Cas12a and nCas9-activation-induced cytidine deaminase for genome editing and as a non-sexual strategy to generate homozygous/multiplex edited plants in the allotetraploid genome of tobacco. Plant Mol. Biol. 2019. [Google Scholar] [CrossRef]
- Yin, X.; Anand, A.; Quick, P.; Bandyopadhyay, A. Editing a Stomatal Developmental Gene in Rice with CRISPR/Cpf1. In Plant Genome Editing with CRISPR Systems; Springer: New York, NY, USA, 2019; pp. 257–268. [Google Scholar]
- Malzahn, A.A.; Tang, X.; Lee, K.; Ren, Q.; Sretenovic, S.; Zhang, Y.; Chen, H.; Kang, M.; Bao, Y.; Zheng, X.; et al. Application of CRISPR-Cas12a temperature sensitivity for improved genome editing in rice, maize, and Arabidopsis. BMC Biol. 2019, 17, 1–14. [Google Scholar] [CrossRef]
- Oconnor, D.; Wright, G.; George, D.; Hunter, M. Development and Application of Speed Breeding Technologies in a Commercial Peanut Breeding Program Development and Application of Speed Breeding Technologies in a Commercial Peanut Breeding Program. Peanut Sci. 2013, 40, 107–114. [Google Scholar] [CrossRef]
- Riaz, A. Unlocking new sources of adult plant resistance to wheat leaf rust. Ph.D. Thesis, The University of Queensland, Queensland, Australia, 2018; pp. 1–241. [Google Scholar]
- Wang, X.; Xuan, H.; Evers, B.; Shrestha, S.; Pless, R.; Poland, J. High-throughput phenotyping with deep learning gives insight into the genetic architecture of flowering time in wheat. bioRxiv 2019, 527911. [Google Scholar]
- Danzi, D.; Briglia, N.; Petrozza, A.; Summerer, S.; Povero, G.; Stivaletta, A.; Cellini, F.; Pignone, D.; de Paola, D.; Janni, M. Can high throughput phenotyping help food security in the mediterranean area? Front. Plant. Sci. 2019, 10, 15. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types | Mutagens | Characteristics (Sources and Description) | Hazards | References |
|---|---|---|---|---|
| X-rays | Electromagnetic radiation; penetrates tissues from just a few millimeters to many centimeters. | Dangerous, penetrating | [59] | |
| Gamma rays | 60Co (Cobalt-60) and 137Cs (Caesium-137); electric magnet radiation generated with radiation isotope and nuclear reactors. | Dangerous, penetrating | [60,59] | |
| Physical Mutagens | Neutron | 235U; there are fast, slow, thermal types; formed in nuclear reactors; unloaded particles; penetrate tissues up to large numbers centimeter; | Very dangerous | [59,60] |
| Beta particles | 32P and 14C; reduced particle accelerators or radioisotopes; electrons; ionizing and penetrating tissues shallowly | Maybe dangerous | [60] | |
| Alpha particles | Sources originating from radiological isotopes; helium nucleus able to penetrate tissues heavily | Very dangerous | [59] | |
| Proton | Present in nuclear reactors and accelerators; derived from the nucleus of hydrogen; penetrate tissues up to several inches. | Very dangerous | [59,60] | |
| Ion beam | Positively charged ions are accelerated at a high speed and used to irradiate living materials, including plant seeds and tissue culture. | Dangerous | [60] | |
| Alkylating agents | The alkylated base can then degrade with bases to create a primary site which is mutagenic or recombinogenic or mispairs in DNA replication mutations, depending on the atom concerned. | Dangerous | [59] | |
| Azide | Just like alkylating agents. | Dangerous | [59] | |
| Hydroxylamine | Just like alkylating agents. | Dangerous | [59,56] | |
| Chemical Mutagens | Nitrous acid | Acts through deamination, replacing cytosine with uracil, which can pair with adenine and thus result in transitions via subsequent replication cycles. | Very Hazard | [56] |
| Acridines | Interspersing between the DNA bases, thus distorting the DNA double helix and the DNA polymerase, recognizes the new basis for this expanded (intercalated) molecule and inserts a frameshift in front of it. | Dangerous | [56] | |
| Base analog | Comprises the transformations (purine to purine and pyrimidine to pyrimidine) into DNA in place of the regular bases during DNA replication and tautomerizing (existent in two forms, which interconvert into one another such that guanine may be present in keto and enol forms). | Some may be dangerous | [56] |
| Sr.no. | Species | Method | Traits | References |
|---|---|---|---|---|
| 1 | Rice | Cross Breeding | Increased spikelet number per panicle | [164] |
| 2 | Rice | Cross Breeding | Yield Increases | [165] |
| 3 | Wheat | Cross Breeding | Increase Grain Yield | [166] |
| 4 | Tomato | Mutation Breeding | Resistance to bacterial wilt (Ralstonia solanacearum) | [167] |
| 5 | Rapeseed | Mutation Breeding | Resistance to stem rot (Sclerotinia sclerotiorum) | [168] |
| 6 | Cotton | Mutation Breeding | Resistance to bacterial blight, cotton leaf curl virus | [169] |
| 7 | Barley | Mutation Breeding | Salinity tolerance | [170] |
| 8 | Sunflower | Mutation Breeding | Semi-dwarf cultivar/dwarf | |
| 9 | Cassava | Mutation Breeding | High-amylose content preferred by diabetes patients because it lowers the insulin level, which prevents quick spikes in glucose contents. | [171] |
| 10 | Groundnut | Mutation Breeding | Dark green, obovate leaf pod; increased seed size, higher yield, moderately resistant to diseases, increased oil and protein content | [172] |
| 11 | Maize | Transgenic Breeding | increased vitamin content (vitamins C, E, or provitamin A) | [173] |
| 12 | Tomato | Transgenic Breeding | Dry Matter Increases | [174] |
| 13 | Soybean | Transgenic Breeding | Altered carbohydrates metabolism | [174] |
| 14 | Barley | Molecular Marker | Adult resistance to stripe rust | [175] |
| 15 | Maize | Molecular Marker | Development of quality protein maize | [22] |
| 16 | Watermelon | Marker-Assisted Selection | Early Flowering | [176] |
| 17 | Canola | QTL | Dynamic growth QTL | [153] |
| 18 | Alfalfa | Intragenesis | Lignin content | [129] |
| 19 | Apple | Cisgenesis, Intragenesis | Scab resistance | [177,178] |
| 20 | Barley | Cisgenesis | Grain phytase activity | [179] |
| 21 | Durum wheat | Cisgenesis | Baking quality | [180] |
| 22 | Perennial ryegrass | Intragenesis | Drought tolerance | [181] |
| 23 | Poplar | Cisgenesis | Plant growth and stature, wood properties | [181] |
| 24 | Potato | Cisgenesis | Late blight resistance | [182] |
| 25 | Strawberry | Intragenesis | Gray mold resistance | [183] |
| 26 | Tomato | Gene editing/ZFN | Reduction of cholesterol and steroidal glycoalkaloids, such as toxic α-solanine and α- chaconine | [184] |
| 27 | Wheat | Gene editing/TALEN | Heritable Modification | [185] |
| 28 | Rice | Gene knockout/ CRISPR/Cas9 | Fragrance | [186] |
| 29 | Bread Wheat and Maize | Gene knockout/ CRISPR/Cas9 | Leaf development; Male fertility, Herbicide resistance | [187] |
| 30 | Poplar | Gene knockout/ CRISPR/Cas9 | Lignin content; Condensed tannin content | [188] |
| 31 | Tomato | Gene editing/ CRISPR/Cas9 | Leaf development | [189] |
| 32 | Soybean | Gene replacement/ CRISPR/Cas9 | Herbicide resistance | [190] |
| 33 | Maize | Gene replacement/ CRISPR/Cas9 | Herbicide resistance | [187] |
| 34 | Cotton | Genome Editing/ CRISPR/Cas9 | Produce transgenic seeds without regeneration | [191] |
| 35 | Soybean | Genome Editing/ CRISPR/Cas9 | Early Flowering | [192] |
| 36 | Rice | Genome Editing/ CRISPR/Cas9 | Increased grain weight | [157] |
| 37 | Tomato | Genome Editing/ CRISPR/Cas9 | Resistance to powdery mildew | [159] |
| 38 | Wheat | Gene knockout/ CRISPR/Cas9 | low-gluten foodstuff | [193] |
| 39 | Rice | Gene knockout/ CRISPR/Cas9 | Generate mutant plants which is sensitive to salt stress | [194] |
| 40 | Rapeseed | Gene knockout/ CRISPR/Cas9 | Controlling pod shattering resistance in oilseed rape | [195] |
| 41 | Tomato, Potato | CRISPR/Cas9 Cytidine Base Editor | Transgene-free plants in the first generation in tomato and potato | [196] |
| 42 | Tobacco | Genome Editing /CRISPR/Cpf1 | Plants harboring | [197] |
| 43 | Rice | Genome Editing /CRISPR/Cpf1 | Regulate the stomatal density in leaf | [198] |
| 44 | Rice | Genome Editing /CRISPR/Cpf1 | Stable mRNA equal | [100,199] |
| 45 | Maize | Genome Editing /CRISPR/Cpf1 | Mutation frequencies doubled | [199] |
| 46 | Chickpea | Rapid generation advance (RGA) | Seven generations per year and enable speed breeding | [48] |
| 47 | Pea | Greenhouse strategy | 6 Generation/year | [124] |
| 48 | Chickpea | Speed Breeding | 4-6 Generation/year | [200] |
| 49 | Barley | Speed Breeding | Resistance to Leaf Rust | [16] |
| 50 | Spring wheat | Speed Breeding | Resistance to Stem Rust | [201] |
| 51 | Spring wheat | Speed Breeding | 4-6 Generation/year | [16] |
| 52 | Barley | Speed Breeding | 4-6 Generation/year | [16] |
| 53 | Peanut | Speed Breeding | 2-3 Generation/year | [200] |
| 54 | Canola | Speed Breeding | 4-6 Generation/year | [16] |
| 55 | Wheat | High-throughput phenotyping (HTP) | Development of improved, high-yielding crop varieties | [202] |
| 56 | Tomato | High-throughput phenotyping (HTP) | Using biostimulants to increase the plant capacity of using water | [203] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmar, S.; Gill, R.A.; Jung, K.-H.; Faheem, A.; Qasim, M.U.; Mubeen, M.; Zhou, W. Conventional and Molecular Techniques from Simple Breeding to Speed Breeding in Crop Plants: Recent Advances and Future Outlook. Int. J. Mol. Sci. 2020, 21, 2590. https://doi.org/10.3390/ijms21072590
Ahmar S, Gill RA, Jung K-H, Faheem A, Qasim MU, Mubeen M, Zhou W. Conventional and Molecular Techniques from Simple Breeding to Speed Breeding in Crop Plants: Recent Advances and Future Outlook. International Journal of Molecular Sciences. 2020; 21(7):2590. https://doi.org/10.3390/ijms21072590
Chicago/Turabian StyleAhmar, Sunny, Rafaqat Ali Gill, Ki-Hong Jung, Aroosha Faheem, Muhammad Uzair Qasim, Mustansar Mubeen, and Weijun Zhou. 2020. "Conventional and Molecular Techniques from Simple Breeding to Speed Breeding in Crop Plants: Recent Advances and Future Outlook" International Journal of Molecular Sciences 21, no. 7: 2590. https://doi.org/10.3390/ijms21072590
APA StyleAhmar, S., Gill, R. A., Jung, K.-H., Faheem, A., Qasim, M. U., Mubeen, M., & Zhou, W. (2020). Conventional and Molecular Techniques from Simple Breeding to Speed Breeding in Crop Plants: Recent Advances and Future Outlook. International Journal of Molecular Sciences, 21(7), 2590. https://doi.org/10.3390/ijms21072590