MiR-144: A New Possible Therapeutic Target and Diagnostic/Prognostic Tool in Cancers

 , ,
, ,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
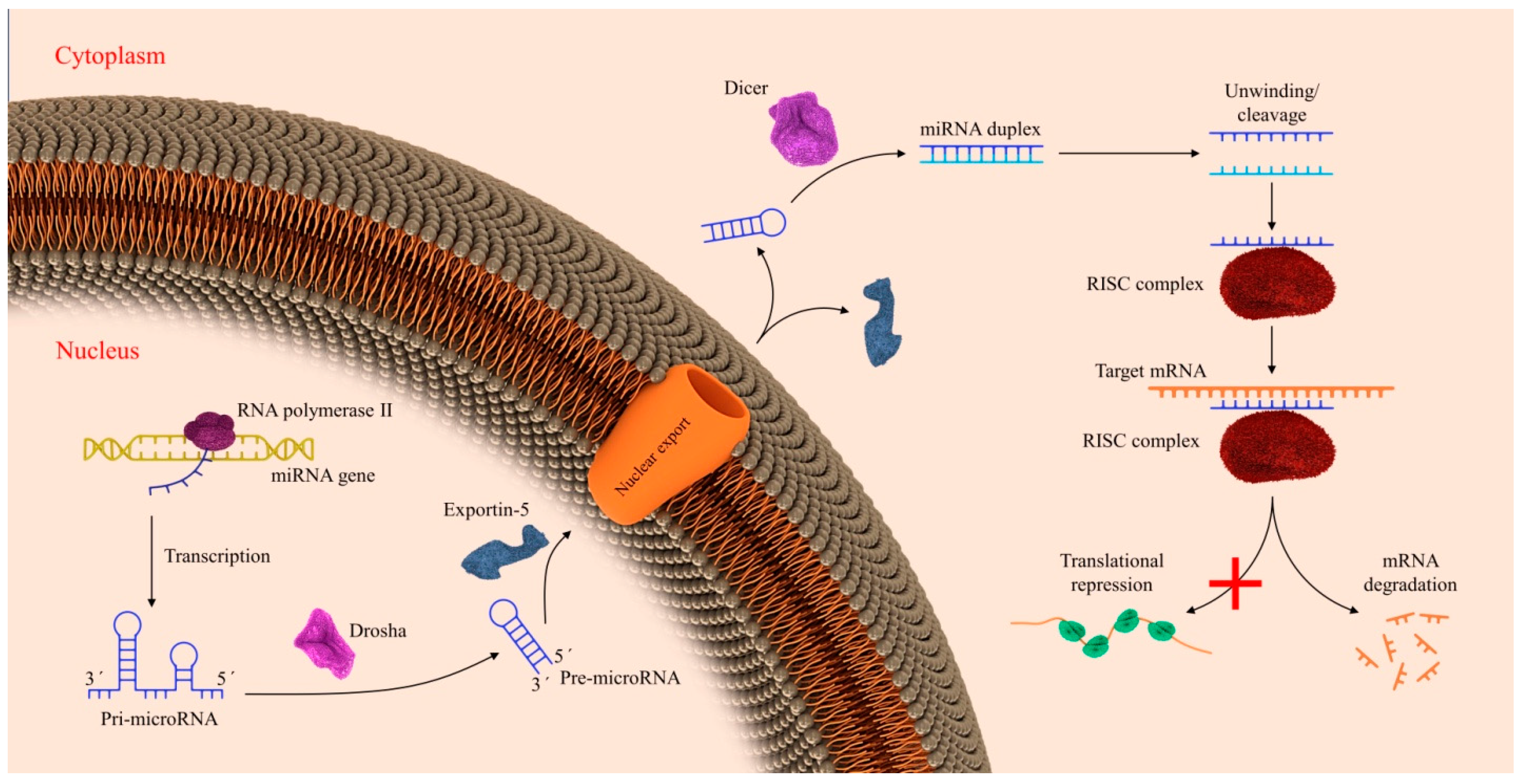
2. Biogenesis of miRNAs
3. MiR-144 in Lymphomas
3.1. MiR-144 in Non-Hodgkin’s Lymphomas
3.1.1. MiR-144 in Diffuse B Cell Lymphoma
3.1.2. MiR-144 in Follicular Lymphoma
3.1.3. MiR-144 in Splenic Marginal Zone Lymphoma
3.1.4. MiR-144 in the Lymphoma of the Primary Central Nervous System
4. MiR-144 in Leukemia
4.1. MiR-144 in Acute Myeloid Leukemia
4.2. MiR-144 in Acute Lymphoblastic Leukemia
4.3. MiR-144 in Chronic Myeloid Leukemia
4.4. MiR-144 in Chronic Lymphocytic Leukemia
5. MiR-144 in Gastrointestinal Cancers
5.1. MiR-144 in Gastric Cancer
5.2. MiR-144 in Colorectal Cancer
6. MiR-144 in Pancreatic Cancer
7. MiR-144 in Hepatocellular Carcinoma
8. MiR-144 in Cholangiocarcinoma
9. MiR-144 in Esophageal Cancer
10. MiR-144 in Genitourinary System
10.1. MiR-144 in Cervical Cancer
10.2. MiR-144 in Ovarian Cancer
10.3. MiR-144 in Prostate Cancer
10.4. MiR-144 in Renal Cell Carcinoma
10.5. MiR-144 in Bladder Cancer
11. MiR-144 in Lung Cancer
12. MiR-144 in Mesothelioma
13. MiR-144 in Breast Cancer
14. MiR-144 in Head and Neck Squamous Cell Carcinoma
14.1. MiR-144 in Nasopharyngeal Carcinoma
14.2. MiR-144 in Thyroid Cancer
14.3. MiR-144 in Glioblastoma
14.4. MiR-144 in Melanoma
14.5. MiR-144 in Osteosarcoma
15. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Pfeffer, S.R.; Yang, C.H.; Pfeffer, L.M. The role of mir-21 in cancer. Drug Dev. Res. 2015, 76, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Qin, C. General hallmarks of micrornas in brain evolution and development. RNA Biol. 2015, 12, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Shahabi, A.; Naghili, B.; Ansarin, K.; Zarghami, N. The relationship between micrornas and rab family gtpases in human cancers. J. Cell. Physiol. 2019, 234, 12341–12352. [Google Scholar] [CrossRef]
- Mollaei, H.; Safaralizadeh, R.; Rostami, Z. Microrna replacement therapy in cancer. J. Cell. Physiol. 2019, 234, 12369–12384. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Shirjang, S.; Baradaran, B. Micrornas in the diagnosis and treatment of cancer. Immunol. Investig. 2017, 46, 880–897. [Google Scholar] [CrossRef]
- Gilam, A.; Conde, J.; Weissglas-Volkov, D.; Oliva, N.; Friedman, E.; Artzi, N.; Shomron, N. Local microrna delivery targets palladin and prevents metastatic breast cancer. Nat. Commun 2016, 7, 12868. [Google Scholar] [CrossRef]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. Oncomir or tumor suppressor? The duplicity of micrornas in cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Shirjang, S.; Baradaran, B. Hmgi-c suppressing induces p53/caspase9 axis to regulate apoptosis in breast adenocarcinoma cells. Cell Cycle (Georget. Tex.) 2016, 15, 2585–2592. [Google Scholar] [CrossRef]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Radmark, O.; Kim, S.; et al. The nuclear rnase iii drosha initiates microrna processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Koscianska, E.; Starega-Roslan, J.; Krzyzosiak, W.J. The role of dicer protein partners in the processing of microrna precursors. PLoS ONE 2011, 6, e28548. [Google Scholar] [CrossRef]
- Grishok, A.; Pasquinelli, A.E.; Conte, D.; Li, N.; Parrish, S.; Ha, I.; Baillie, D.L.; Fire, A.; Ruvkun, G.; Mello, C.C. Genes and mechanisms related to rna interference regulate expression of the small temporal rnas that control C. elegans developmental timing. Cell 2001, 106, 23–34. [Google Scholar] [CrossRef]
- Khvorova, A.; Reynolds, A.; Jayasena, S.D. Functional sirnas and mirnas exhibit strand bias. Cell 2003, 115, 209–216. [Google Scholar] [CrossRef]
- Graves, P.; Zeng, Y. Biogenesis of mammalian micrornas: A global view. Genom. Proteom. Bioinform. 2012, 10, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Shi, R.; Zhao, S.; Li, X.; Lu, S.; Bu, H.; Ma, X.; Su, C. E2f8, a direct target of mir-144, promotes papillary thyroid cancer progression via regulating cell cycle. J. Exp. Clin. Cancer Res. 2017, 36, 40. [Google Scholar] [CrossRef]
- Cullen, B.R. Transcription and processing of human microrna precursors. Mol. Cell 2004, 16, 861–865. [Google Scholar] [CrossRef]
- Liu, Y.; Barta, S.K. Diffuse large b-cell lymphoma: 2019 update on diagnosis, risk stratification, and treatment. Am. J. Hematol. 2019, 94, 604–616. [Google Scholar] [CrossRef]
- Friedberg, J.W.; Fisher, R.I. Diffuse large b-cell lymphoma. Hematol. Oncol. Clin. N. Am. 2008, 22, 941–ix. [Google Scholar] [CrossRef]
- Musilova, K.; Mraz, M. Micrornas in b-cell lymphomas: How a complex biology gets more complex. Leukemia 2015, 29, 1004–1017. [Google Scholar] [CrossRef]
- Wang, H.; Wang, A.; Hu, Z.; Xu, X.; Liu, Z.; Wang, Z. A critical role of mir-144 in diffuse large b-cell lymphoma proliferation and invasion. Cancer Immunol. Res. 2016, 4, 337–344. [Google Scholar] [CrossRef]
- Wang, T.; Wu, F.; Yu, D. Mir-144/451 in hematopoiesis and beyond. ExRNA 2019, 1, 16. [Google Scholar] [CrossRef]
- Montes-Moreno, S.; Martinez, N.; Sanchez-Espiridión, B.; Díaz Uriarte, R.; Rodriguez, M.E.; Saez, A.; Montalbán, C.; Gomez, G.; Pisano, D.G.; García, J.F.; et al. Mirna expression in diffuse large b-cell lymphoma treated with chemoimmunotherapy. Blood 2011, 118, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Luminari, S.; Bellei, M.; Biasoli, I.; Federico, M. Follicular lymphoma - treatment and prognostic factors. Rev. Bras. Hematol. Hemoter. 2012, 34, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Hershkovitz-Rokah, O.; Geva, P.; Salmon-Divon, M.; Shpilberg, O.; Liberman-Aronov, S. Network analysis of micrornas, genes and their regulation in diffuse and follicular b-cell lymphomas. Oncotarget 2018, 9, 7928–7941. [Google Scholar] [CrossRef] [PubMed]
- Malpeli, G.; Barbi, S.; Greco, C.; Zupo, S.; Bertolaso, A.; Scupoli, M.T.; Krampera, M.; Kamga, P.T.; Croce, C.M.; Scarpa, A.; et al. Microrna signatures and foxp3(+) cell count correlate with relapse occurrence in follicular lymphoma. Oncotarget 2018, 9, 19961–19979. [Google Scholar] [CrossRef][Green Version]
- Takei, Y.; Ohnishi, N.; Kisaka, M.; Mihara, K. Determination of abnormally expressed micrornas in bone marrow smears from patients with follicular lymphomas. SpringerPlus 2014, 3, 288. [Google Scholar] [CrossRef]
- Santos, T.S.D.; Tavares, R.S.; Farias, D.L.C.D. Splenic marginal zone lymphoma: A literature review of diagnostic and therapeutic challenges. Rev. Bras. Hematol. Hemoter. 2017, 39, 146–154. [Google Scholar] [CrossRef]
- Di Lisio, L.; Sánchez-Beato, M.; Gómez-López, G.; Rodríguez, M.E.; Montes-Moreno, S.; Mollejo, M.; Menárguez, J.; Martínez, M.A.; Alves, F.J.; Pisano, D.G.; et al. Microrna signatures in b-cell lymphomas. Blood Cancer J. 2012, 2, e57. [Google Scholar] [CrossRef]
- Bouteloup, M.; Verney, A.; Rachinel, N.; Callet-Bauchu, E.; Ffrench, M.; Coiffier, B.; Magaud, J.P.; Berger, F.; Salles, G.A.; Traverse-Glehen, A.J.B.j.o.h. Microrna expression profile in splenic marginal zone lymphoma. Br. J. Haematol. 2012, 156, 279–281. [Google Scholar] [CrossRef]
- Peveling-Oberhag, J.; Crisman, G.; Schmidt, A.; Döring, C.; Lucioni, M.; Arcaini, L.; Rattotti, S.; Hartmann, S.; Piiper, A.; Hofmann, W.J.L. Dysregulation of global microrna expression in splenic marginal zone lymphoma and influence of chronic hepatitis c virus infection. Leukemia 2012, 26, 1654. [Google Scholar] [CrossRef]
- Grommes, C.; DeAngelis, L.M. Primary cns lymphoma. J. Clin. Oncol. 2017, 35, 2410–2418. [Google Scholar] [CrossRef]
- Löw, S.; Han, C.H.; Batchelor, T.T. Primary central nervous system lymphoma. Adv. Neurol. Disord. 2018, 11, 1756286418793562. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Zhang, X.; Yin, K.-J. Micrornas in central nervous system diseases: A prospective role in regulating blood-brain barrier integrity. Exp. Neurol. 2020, 323, 113094. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Xue, Y.; Wang, P.; Wang, Z.; Li, Z.; Hu, Y.; Li, Z.; Shang, X.; Liu, Y. The long noncoding rna tug1 regulates blood-tumor barrier permeability by targeting mir-144. Oncotarget 2015, 6, 19759–19779. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Suolitiken, D.; Xu, X.; Chen, B.; Zhang, L.; Kang, H.; Lin, Z.; Wang, Q. Circulating micrornas is a potential prognostic biomarker in primary central nervous system lymphoma. J. Abbr. 2018, 8, 1839–1846. [Google Scholar]
- Saultz, J.N.; Garzon, R. Acute myeloid leukemia: A concise review. J. Clin. Med. 2016, 5, 33. [Google Scholar] [CrossRef]
- Villela, L.; Bolaños-Meade, J. Acute myeloid leukaemia: Optimal management and recent developments. Drugs 2011, 71, 1537–1550. [Google Scholar] [CrossRef]
- Arber, D.A. The 2016 who classification of acute myeloid leukemia: What the practicing clinician needs to know. Semin. Hematol. 2019, 56, 90–95. [Google Scholar] [CrossRef]
- Hasserjian, R.P. Acute myeloid leukemia: Advances in diagnosis and classification. Int. J. Lab. Hematol. 2013, 35, 358–366. [Google Scholar] [CrossRef]
- Sun, X.; Liu, D.; Xue, Y.; Hu, X. Enforced mir-144-3p expression as a non-invasive biomarker for the acute myeloid leukemia patients mainly by targeting nrf2. Clin. Lab. 2017, 63, 679–687. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, J.; Chen, S.; Shen, K.; Ai, G.; Dai, X.; Xie, B.; Shi, Y.; Jiang, S.; Feng, J.; et al. Decreased mir-144 expression as a non-invasive biomarker for acute myeloid leukemia patients. Die Pharm. 2017, 72, 232–235. [Google Scholar]
- Daver, N.; Schlenk, R.F.; Russell, N.H.; Levis, M.J. Targeting flt3 mutations in aml: Review of current knowledge and evidence. Leukemia 2019, 33, 299–312. [Google Scholar] [CrossRef]
- Wu, M.; Li, C.; Zhu, X. Flt3 inhibitors in acute myeloid leukemia. J. Hematol. Oncol. 2018, 11, 133. [Google Scholar] [CrossRef] [PubMed]
- Whitman, S.P.; Maharry, K.; Radmacher, M.D.; Becker, H.; Mrózek, K.; Margeson, D.; Holland, K.B.; Wu, Y.-Z.; Schwind, S.; Metzeler, K.H.; et al. Flt3 internal tandem duplication associates with adverse outcome and gene- and microrna-expression signatures in patients 60 years of age or older with primary cytogenetically normal acute myeloid leukemia: A cancer and leukemia group b study. Blood 2010, 116, 3622–3626. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.M. Letting micrornas overcome resistance to chemotherapy in acute myeloid leukemia. Leuk. Lymphoma 2014, 55, 1449–1450. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, R.; Tawa, A.; Hanada, R.; Horibe, K.; Tsuchida, M.; Tsukimoto, I. Extramedullary infiltration at diagnosis and prognosis in children with acute myelogenous leukemia. Pediatric Blood Cancer 2007, 48, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Meng, W.; Yu, G.; Yin, C.; Wang, Z.; Liao, L.; Meng, F. Microrna-144 targets app to regulate aml1/eto(+) leukemia cell migration via the p-erk/c-myc/mmp-2 pathway. Oncol. Lett. 2019, 18, 2034–2042. [Google Scholar] [CrossRef]
- Terwilliger, T.; Abdul-Hay, M. Acute lymphoblastic leukemia: A comprehensive review and 2017 update. Blood Cancer J. 2017, 7, e577. [Google Scholar] [CrossRef]
- Inaba, H.; Greaves, M.; Mullighan, C.G. Acute lymphoblastic leukaemia. Lancet 2013, 381, 1943–1955. [Google Scholar] [CrossRef]
- Ultimo, S.; Martelli, A.M.; Zauli, G.; Vitale, M.; Calin, G.A.; Neri, L.M. Roles and clinical implications of micrornas in acute lymphoblastic leukemia. J. Cell. Physiol. 2018, 233, 5642–5654. [Google Scholar] [CrossRef]
- Jin, J.; Wang, Y.; Xu, Y.; Zhou, X.; Liu, Y.; Li, X.; Wang, J. Microrna-144 regulates cancer cell proliferation and cell-cycle transition in acute lymphoblastic leukemia through the interaction of fmn2. J. Gene Med. 2017, 19, e2898. [Google Scholar] [CrossRef]
- Mavrakis, K.J.; Van Der Meulen, J.; Wolfe, A.L.; Liu, X.; Mets, E.; Taghon, T.; Khan, A.A.; Setty, M.; Rondou, P.; Vandenberghe, P.; et al. A cooperative microrna-tumor suppressor gene network in acute t-cell lymphoblastic leukemia (t-all). Nat. Genet. 2011, 43, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Granatowicz, A.; Piatek, C.I.; Moschiano, E.; El-Hemaidi, I.; Armitage, J.D.; Akhtari, M. An overview and update of chronic myeloid leukemia for primary care physicians. Korean J. Fam. Med. 2015, 36, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, E.; Kantarjian, H. Chronic myeloid leukemia: 2018 update on diagnosis, therapy and monitoring. Am. J. Hematol. 2018, 93, 442–459. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, M.; Croce, C.M.; Calin, G.A. Micrornas in the ontogeny of leukemias and lymphomas. Leuk Lymphoma 2009, 50, 160–170. [Google Scholar] [CrossRef]
- Liu, L.; Wang, S.; Chen, R.; Wu, Y.; Zhang, B.; Huang, S.; Zhang, J.; Xiao, F.; Wang, M.; Liang, Y. Myc induced mir-144/451 contributes to the acquired imatinib resistance in chronic myelogenous leukemia cell k562. Biochem. Biophys. Res. Commun. 2012, 425, 368–373. [Google Scholar] [CrossRef]
- Hehlmann, R. How i treat cml blast crisis. Blood 2012, 120, 737–747. [Google Scholar] [CrossRef]
- Machova Polakova, K.; Koblihova, J.; Stopka, T. Role of epigenetics in chronic myeloid leukemia. Curr. Hematol. Malig. Rep. 2013, 8, 28–36. [Google Scholar] [CrossRef]
- Kipps, T.J.; Stevenson, F.K.; Wu, C.J.; Croce, C.M.; Packham, G.; Wierda, W.G.; O’Brien, S.; Gribben, J.; Rai, K. Chronic lymphocytic leukaemia. Nat. Rev. Dis Primers 2017, 3, 16096. [Google Scholar] [CrossRef]
- Hallek, M. Chronic lymphocytic leukemia: 2017 update on diagnosis, risk stratification, and treatment. Am. J. Hematol. 2017, 92, 946–965. [Google Scholar] [CrossRef]
- Van Roosbroeck, K.; Calin, G.A. Micrornas in chronic lymphocytic leukemia: Miracle or mirage for prognosis and targeted therapies? Semin. Oncol. 2016, 43, 209–214. [Google Scholar] [CrossRef]
- Ruiz-Lafuente, N.; Alcaraz-García, M.-J.; Sebastián-Ruiz, S.; García-Serna, A.-M.; Gómez-Espuch, J.; Moraleda, J.-M.; Minguela, A.; García-Alonso, A.-M.; Parrado, A. Il-4 up-regulates mir-21 and the mirnas hosted in the clcn5 gene in chronic lymphocytic leukemia. PLoS ONE 2015, 10, e0124936. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Zhou, C.; Zhuang, J.; Liu, L.; Wei, J.; Liu, C.; Li, H.; Sun, C. Identification of key candidate genes and mirna-mrna target pairs in chronic lymphocytic leukemia by integrated bioinformatics analysis. Mol. Med. Rep. 2019, 19, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Sitarz, R.; Skierucha, M.; Mielko, J.; Offerhaus, G.J.A.; Maciejewski, R.; Polkowski, W.P. Gastric cancer: Epidemiology, prevention, classification, and treatment. Cancer Manag Res. 2018, 10, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yao, G.; Zhai, J.; Hu, D.; Fan, Y. Lncrna ftx promotes proliferation and invasion of gastric cancer via mir-144/zfx axis. Onco Targets 2019, 12, 11701–11713. [Google Scholar] [CrossRef] [PubMed]
- Akiyoshi, S.; Fukagawa, T.; Ueo, H.; Ishibashi, M.; Takahashi, Y.; Fabbri, M.; Sasako, M.; Maehara, Y.; Mimori, K.; Mori, M. Clinical significance of mir-144-zfx axis in disseminated tumour cells in bone marrow in gastric cancer cases. Br. J. Cancer 2012, 107, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, F.; Zhang, J.; Li, J. Mir-144 suppresses cell proliferation and invasion in gastric cancer through downregulation of activating enhancer-binding protein 4. Oncol. Lett. 2019, 17, 5686–5692. [Google Scholar] [CrossRef]
- Liu, J.; Xue, H.; Zhang, J.; Suo, T.; Xiang, Y.; Zhang, W.; Ma, J.; Cai, D.; Gu, X. Microrna-144 inhibits the metastasis of gastric cancer by targeting met expression. J. Exp. Clin. Cancer Res. 2015, 34, 35. [Google Scholar] [CrossRef]
- Ren, K.; Liu, Q.Q.; An, Z.F.; Zhang, D.P.; Chen, X.H. Mir-144 functions as tumor suppressor by targeting pim1 in gastric cancer. Eur. Rev. Med. Pharm. Sci. 2017, 21, 3028–3037. [Google Scholar]
- Ji, T.T.; Huang, X.; Jin, J.; Pan, S.H.; Zhuge, X.J. Inhibition of long non-coding rna tug1 on gastric cancer cell transference and invasion through regulating and controlling the expression of mir-144/c-met axis. Asian Pac. J. Trop. Med. 2016, 9, 508–512. [Google Scholar] [CrossRef]
- Lario, S.; Brunet-Vega, A.; Quílez, M.E.; Ramírez-Lázaro, M.J.; Lozano, J.J.; García-Martínez, L.; Pericay, C.; Miquel, M.; Junquera, F.; Campo, R.; et al. Expression profile of circulating micrornas in the correa pathway of progression to gastric cancer. United Eur. Gastroenterol. J. 2018, 6, 691–701. [Google Scholar] [CrossRef]
- Liu, S.; Suo, J.; Wang, C.; Sun, X.; Wang, D.; He, L.; Zhang, Y.; Li, W. Prognostic significance of low mir-144 expression in gastric cancer. Cancer Biomark. Sect. A Dis. Markers 2017, 20, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Gu, A.; Wang, Z.; Xue, Y. Microrna-144 functions as a tumor suppressor in gastric cancer by targeting cyclooxygenase-2. Exp. Ther. Med. 2018, 15, 3088–3095. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.G.; Tian, R.C.; Liu, Y.; Niu, A.Y.; Zhang, J.; Gao, W.F. The role of mir-144/gspt1 axis in gastric cancer. Eur. Rev. Med. Pharm. Sci. 2018, 22, 4138–4145. [Google Scholar]
- Vatandoust, S.; Price, T.J.; Karapetis, C.S. Colorectal cancer: Metastases to a single organ. World J. Gastroenterol. 2015, 21, 11767–11776. [Google Scholar] [CrossRef]
- Jiang, Y.; Cai, Y.; Shao, W.; Li, F.; Guan, Z.; Zhou, Y.; Tang, C.; Feng, S. Microrna144 suppresses aggressive phenotypes of tumor cells by targeting ano1 in colorectal cancer. Oncol. Rep. 2019, 41, 2361–2370. [Google Scholar]
- Han, B.; Feng, D.; Yu, X.; Liu, Y.; Yang, M.; Luo, F.; Zhou, L.; Liu, F. Microrna-144 mediates chronic inflammation and tumorigenesis in colorectal cancer progression via regulating c-x-c motif chemokine ligand 11. Exp. Ther. Med. 2018, 16, 1935–1943. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. Mtor signaling in growth, metabolism, and disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Iwaya, T.; Yokobori, T.; Nishida, N.; Kogo, R.; Sudo, T.; Tanaka, F.; Shibata, K.; Sawada, G.; Takahashi, Y.; Ishibashi, M.; et al. Downregulation of mir-144 is associated with colorectal cancer progression via activation of mtor signaling pathway. Carcinogenesis 2012, 33, 2391–2397. [Google Scholar] [CrossRef]
- Koga, Y.; Yasunaga, M.; Takahashi, A.; Kuroda, J.; Moriya, Y.; Akasu, T.; Fujita, S.; Yamamoto, S.; Baba, H.; Matsumura, Y. Microrna expression profiling of exfoliated colonocytes isolated from feces for colorectal cancer screening. Cancer Prev. Res. (Phila. Pa.) 2010, 3, 1435–1442. [Google Scholar] [CrossRef]
- Kalimutho, M.; Del Vecchio Blanco, G.; Di Cecilia, S.; Sileri, P.; Cretella, M.; Pallone, F.; Federici, G.; Bernardini, S. Differential expression of mir-144* as a novel fecal-based diagnostic marker for colorectal cancer. J. Gastroenterol. 2011, 46, 1391–1402. [Google Scholar] [CrossRef]
- Choi, H.H.; Cho, Y.-S.; Choi, J.H.; Kim, H.-K.; Kim, S.S.; Chae, H.-S. Stool-based mir-92a and mir-144* as noninvasive biomarkers for colorectal cancer screening. Oncology 2019, 97, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M. Pancreatic cancer. N. Engl. J. Med. 2010, 362, 1605–1617. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, O.; Russo, A.; Scarpa, A.; Santini, D.; Reni, M.; Bittoni, A.; Azzariti, A.; Aprile, G.; Delcuratolo, S.; Signorile, M.; et al. Microrna in pancreatic adenocarcinoma: Predictive/prognostic biomarkers or therapeutic targets? Oncotarget 2015, 6, 23323–23341. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sun, P.; Yue, Z.; Zhang, D.; You, K.; Wang, J. Mir-144-3p induces cell cycle arrest and apoptosis in pancreatic cancer cells by targeting proline-rich protein 11 expression via the mitogen-activated protein kinase signaling pathway. DNA Cell Biol. 2017, 36, 619–626. [Google Scholar] [CrossRef]
- Liu, S.; Luan, J.; Ding, Y. Mir-144-3p targets fosb proto-oncogene, ap-1 transcription factor subunit (fosb) to suppress proliferation, migration, and invasion of panc-1 pancreatic cancer cells. Oncol. Res. 2018, 26, 683–690. [Google Scholar] [CrossRef]
- Balogh, J.; Victor, D., 3rd; Asham, E.H.; Burroughs, S.G.; Boktour, M.; Saharia, A.; Li, X.; Ghobrial, R.M.; Monsour, H.P., Jr. Hepatocellular carcinoma: A review. J. Hepatocell. Carcinoma 2016, 3, 41–53. [Google Scholar] [CrossRef]
- He, Q.; Wang, F.; Honda, T.; Lindquist, D.M.; Dillman, J.R.; Timchenko, N.A.; Redington, A.N. Intravenous mir-144 inhibits tumor growth in diethylnitrosamine-induced hepatocellular carcinoma in mice. Tumour Biol. J. Int. Soc. Oncodevelopmental. Biol. Med. 2017, 39, 1010428317737729. [Google Scholar] [CrossRef]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharm. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef]
- Zhou, S.; Ye, W.; Zhang, Y.; Yu, D.; Shao, Q.; Liang, J.; Zhang, M. Mir-144 reverses chemoresistance of hepatocellular carcinoma cell lines by targeting nrf2-dependent antioxidant pathway. Am. J. Transl. Res. 2016, 8, 2992–3002. [Google Scholar]
- Gao, Y.; Feng, B.; Lu, L.; Han, S.; Chu, X.; Chen, L.; Wang, R. Mirnas and e2f3: A complex network of reciprocal regulations in human cancers. Oncotarget 2017, 8, 60624–60639. [Google Scholar] [CrossRef]
- Cao, T.; Li, H.; Hu, Y.; Ma, D.; Cai, X. Mir-144 suppresses the proliferation and metastasis of hepatocellular carcinoma by targeting e2f3. Tumour Biol. J. Int. Soc. Oncodevelopmental. Biol. Med. 2014, 35, 10759–10764. [Google Scholar] [CrossRef] [PubMed]
- Pu, C.; Huang, H.; Wang, Z.; Zou, W.; Lv, Y.; Zhou, Z.; Zhang, Q.; Qiao, L.; Wu, F.; Shao, S. Extracellular vesicle-associated mir-21 and mir-144 are markedly elevated in serum of patients with hepatocellular carcinoma. Front. Physiol. 2018, 9, 930. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Huang, C.; Huang, X.; Liang, R.; Feng, Y.; Luo, X. Microrna-144-3p suppresses tumor growth and angiogenesis by targeting sgk3 in hepatocellular carcinoma. Oncol. Rep. 2017, 38, 2173–2181. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Lin, L.; Du, Y.; Hao, X.; Zhao, Y.; Liu, X. Microrna-588 suppresses tumor cell migration and invasion by targeting grn in lung squamous cell carcinoma. Mol. Med. Rep. 2016, 14, 3021–3028. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Lin, Y.; Zhou, Y.; Jin, H.; Hou, B.; Wu, Z.; Li, Z.; Jian, Z.; Sun, J. Mir-144 suppresses cell proliferation, migration, and invasion in hepatocellular carcinoma by targeting smad4. Onco Targets 2016, 9, 4705–4714. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Kong, Y.; Gao, Z.; Liu, Y.; Zhu, P.; Yu, Z. Lncrna tug1 interacting with mir-144 contributes to proliferation, migration and tumorigenesis through activating the jak2/stat3 pathway in hepatocellular carcinoma. Int. J. Biochem. Cell Biol. 2018, 101, 19–28. [Google Scholar] [CrossRef]
- Blechacz, B. Cholangiocarcinoma: Current knowledge and new developments. Gut Liver 2017, 11, 13–26. [Google Scholar] [CrossRef]
- Alsaleh, M.; Leftley, Z.; Barbera, T.A.; Sithithaworn, P.; Khuntikeo, N.; Loilome, W.; Yongvanit, P.; Cox, I.J.; Chamodol, N.; Syms, R.R.; et al. Cholangiocarcinoma: A guide for the nonspecialist. Int J. Gen. Med. 2018, 12, 13–23. [Google Scholar] [CrossRef]
- Ghouri, Y.A.; Mian, I.; Blechacz, B. Cancer review: Cholangiocarcinoma. J. Carcinog. 2015, 14. [Google Scholar] [CrossRef]
- Yang, R.; Chen, Y.; Tang, C.; Li, H.; Wang, B.; Yan, Q.; Hu, J.; Zou, S. Microrna-144 suppresses cholangiocarcinoma cell proliferation and invasion through targeting platelet activating factor acetylhydrolase isoform 1b. BMC Cancer 2014, 14, 917. [Google Scholar] [CrossRef]
- He, Y.; Liu, C.; Song, P.; Pang, Z.; Mo, Z.; Huang, C.; Yan, T.; Sun, M.; Fa, X. Investigation of mirna- and lncrna-mediated competing endogenous rna network in cholangiocarcinoma. Oncol. Lett. 2019, 18, 5283–5293. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. Epidemiology of esophageal cancer. World J. Gastroenterol. 2013, 19, 5598–5606. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Liu, R.; Liao, J.; Yang, M.; Pan, E.; Yin, L.; Pu, Y. Possible tumor suppressive role of the mir-144/451 cluster in esophageal carcinoma as determined by principal component regression analysis. Mol. Med. Rep. 2016, 14, 3805–3813. [Google Scholar] [CrossRef]
- Gao, Z.; Zhang, P.; Xie, M.; Gao, H.; Yin, L.; Liu, R. Mir-144/451 cluster plays an oncogenic role in esophageal cancer by inhibiting cell invasion. Cancer Cell Int. 2018, 18, 184. [Google Scholar] [CrossRef]
- Sharma, P.; Sharma, R. Mir-144 functions as an oncomir in kyse-410 human esophageal carcinoma cell line in vitro and targets pura. Neoplasma 2018, 65, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Hou, W.; Wu, Z.; Wang, Y.; Yi, Y.; Lin, W. [mirna-144 in the saliva is a genetic marker for early diagnosis of esophageal cancer]. Nan Fang Yi Ke Da Xue Xue Bao = J. South. Med. Univ. 2013, 33, 1783–1786. [Google Scholar]
- Sharma, P.; Saraya, A.; Sharma, R. Potential diagnostic implications of mir-144 overexpression in human oesophageal cancer. Indian J. Med. Res. 2016, 143, S91–s103. [Google Scholar] [CrossRef]
- Tsikouras, P.; Zervoudis, S.; Manav, B.; Tomara, E.; Iatrakis, G.; Romanidis, C.; Bothou, A.; Galazios, G. Cervical cancer: Screening, diagnosis and staging. J. B.U.On. Off. J. Balk. Union Oncol. 2016, 21, 320–325. [Google Scholar]
- Tao, P.; Wen, H.; Yang, B.; Zhang, A.; Wu, X.; Li, Q. Mir-144 inhibits growth and metastasis of cervical cancer cells by targeting vegfa and vegfc. Exp. Ther. Med. 2018, 15, 562–568. [Google Scholar] [CrossRef]
- Shi, F.; Su, J.; Liu, Z.; Wang, J.; Wang, T. Mir-144 reverses cisplatin resistance in cervical cancer via targeting lhx2. J. Cell. Biochem. 2019, 120, 15018–15026. [Google Scholar] [CrossRef]
- Wu, J.; Zhao, Y.; Li, F.; Qiao, B. Mir-144-3p: A novel tumor suppressor targeting mapk6 in cervical cancer. J. Physiol. Biochem. 2019, 75, 143–152. [Google Scholar] [CrossRef]
- Reid, B.M.; Permuth, J.B.; Sellers, T.A. Epidemiology of ovarian cancer: A review. Cancer Biol Med. 2017, 14, 9–32. [Google Scholar] [PubMed]
- Thériault, B.L.; Basavarajappa, H.D.; Lim, H.; Pajovic, S.; Gallie, B.L.; Corson, T.W. Transcriptional and epigenetic regulation of kif14 overexpression in ovarian cancer. PLoS ONE 2014, 9, e91540. [Google Scholar] [CrossRef] [PubMed]
- Palma Flores, C.; Garcia-Vazquez, R.; Gallardo Rincon, D.; Ruiz-Garcia, E.; Astudillo de la Vega, H.; Marchat, L.A.; Salinas Vera, Y.M.; Lopez-Camarillo, C. Micrornas driving invasion and metastasis in ovarian cancer: Opportunities for translational medicine (review). Int. J. Oncol. 2017, 50, 1461–1476. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.; Woo, H.-H.; Varghese, F.; Zhou, M.; Chambers, S.K. Circulating mirna profiling of women at high risk for ovarian cancer. Transl. Oncol. 2019, 12, 714–725. [Google Scholar] [CrossRef]
- Pogge von Strandmann, E.; Reinartz, S.; Wager, U.; Muller, R. Tumor-host cell interactions in ovarian cancer: Pathways to therapy failure. Trends Cancer 2017, 3, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Penyige, A.; Márton, É.; Soltész, B.; Szilágyi-Bónizs, M.; Póka, R.; Lukács, J.; Széles, L.; Nagy, B. Circulating mirna profiling in plasma samples of ovarian cancer patients. Int. J. Mol. Sci. 2019, 20, 4533. [Google Scholar] [CrossRef]
- Walter, B.A.; Valera, V.A.; Pinto, P.A.; Merino, M.J. Comprehensive microrna profiling of prostate cancer. J. Cancer 2013, 4, 350–357. [Google Scholar] [CrossRef]
- Litwin, M.S.; Tan, H.J. The diagnosis and treatment of prostate cancer: A review. Jama 2017, 317, 2532–2542. [Google Scholar] [CrossRef]
- Cuzick, J.; Thorat, M.A.; Andriole, G.; Brawley, O.W.; Brown, P.H.; Culig, Z.; Eeles, R.A.; Ford, L.G.; Hamdy, F.C.; Holmberg, L.; et al. Prevention and early detection of prostate cancer. Lancet Oncol. 2014, 15, e484–e492. [Google Scholar] [CrossRef]
- Liu, F.; Wang, J.; Fu, Q.; Zhang, X.; Wang, Y.; Liu, J.; Huang, J.; Lv, X. Vegf-activated mir-144 regulates autophagic survival of prostate cancer cells against cisplatin. Tumour Biol. J. Int. Soc. Oncodevelopmental. Biol. Med. 2015, 37, 15627–15633. [Google Scholar] [CrossRef]
- Gu, H.; Liu, M.; Ding, C.; Wang, X.; Wang, R.; Wu, X.; Fan, R. Hypoxia-responsive mir-124 and mir-144 reduce hypoxia-induced autophagy and enhance radiosensitivity of prostate cancer cells via suppressing pim1. Cancer Med. 2016, 5, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Guo, Z.; Zheng, X.; Cheng, W.; Huang, X. Microrna-144-3p inhibits cell proliferation and induces cell apoptosis in prostate cancer by targeting cep55. Am. J. Transl. Res. 2018, 10, 2457–2468. [Google Scholar] [PubMed]
- Hsieh, J.J.; Purdue, M.P.; Signoretti, S.; Swanton, C.; Albiges, L.; Schmidinger, M.; Heng, D.Y.; Larkin, J.; Ficarra, V. Renal cell carcinoma. Nat. Rev. Dis Primers 2017, 3, 17009. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Arai, T.; Kojima, S.; Sugawara, S.; Kato, M.; Okato, A.; Yamazaki, K.; Naya, Y.; Ichikawa, T.; Seki, N. Regulation of antitumor mir-144-5p targets oncogenes: Direct regulation of syndecan-3 and its clinical significance. Cancer Sci 2018, 109, 2919–2936. [Google Scholar] [CrossRef] [PubMed]
- Xiang, C.; Cui, S.P.; Ke, Y. Mir-144 inhibits cell proliferation of renal cell carcinoma by targeting mtor. J. Huazhong Univ. Sci. Technol. Med. Sci. = Hua zhong ke ji da xue xue bao. Yi xue Ying De wen ban = Huazhong keji daxue xuebao. Yixue Yingdewen ban 2016, 36, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Chen, N.; Xiao, R.; Wang, W.; Pan, Z. Mir-144-3p serves as a tumor suppressor for renal cell carcinoma and inhibits its invasion and metastasis by targeting map3k8. Biochem. Biophys. Res. Commun. 2016, 480, 87–93. [Google Scholar] [CrossRef]
- Xiao, W.; Lou, N.; Ruan, H.; Bao, L.; Xiong, Z.; Yuan, C.; Tong, J.; Xu, G.; Zhou, Y.; Qu, Y.; et al. Mir-144-3p promotes cell proliferation, metastasis, sunitinib resistance in clear cell renal cell carcinoma by downregulating arid1a. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 43, 2420–2433. [Google Scholar] [CrossRef]
- Lou, N.; Ruan, A.M.; Qiu, B.; Bao, L.; Xu, Y.C.; Zhao, Y.; Sun, R.L.; Zhang, S.T.; Xu, G.H.; Ruan, H.L.; et al. Mir-144-3p as a novel plasma diagnostic biomarker for clear cell renal cell carcinoma. Urol. Oncol. 2017, 35, 36.e7–36.e14. [Google Scholar] [CrossRef]
- Boormans, J.L.; Zwarthoff, E.C. Limited funds for bladder cancer research and what can we do about it. Bladder Cancer 2016, 2, 49–51. [Google Scholar] [CrossRef]
- DeGeorge, K.C.; Holt, H.R.; Hodges, S.C. Bladder cancer: Diagnosis and treatment. Am. Fam. Physician 2017, 96, 507–514. [Google Scholar] [PubMed]
- Guo, Y.; Ying, L.; Tian, Y.; Yang, P.; Zhu, Y.; Wang, Z.; Qiu, F.; Lin, J. Mir-144 downregulation increases bladder cancer cell proliferation by targeting ezh2 and regulating wnt signaling. FEBS J. 2013, 280, 4531–4538. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, R.; Seki, N.; Chiyomaru, T.; Inoguchi, S.; Ishihara, T.; Goto, Y.; Nishikawa, R.; Mataki, H.; Tatarano, S.; Itesako, T.; et al. Tumour-suppressive microrna-144-5p directly targets ccne1/2 as potential prognostic markers in bladder cancer. Br. J. Cancer 2015, 113, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Tölle, A.; Jung, M.; Rabenhorst, S.; Kilic, E.; Jung, K.; Weikert, S. Identification of micrornas in blood and urine as tumour markers for the detection of urinary bladder cancer. Oncol. Rep. 2013, 30, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Kavanaugh, J.; Hugo, G.; Robinson, C.G.; Roach, M.C. Anatomical adaptation-early clinical evidence of benefit and future needs in lung cancer. Semin. Radiat. Oncol. 2019, 29, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Uchida, A.; Seki, N.; Mizuno, K.; Misono, S.; Yamada, Y.; Kikkawa, N.; Sanada, H.; Kumamoto, T.; Suetsugu, T.; Inoue, H. Involvement of dual-strand of the mir-144 duplex and their targets in the pathogenesis of lung squamous cell carcinoma. Cancer Sci. 2019, 110, 420–432. [Google Scholar] [CrossRef]
- Yang, Q.S.; Li, B.; Xu, G.; Yang, S.Q.; Wang, P.; Tang, H.H.; Liu, Y.Y. Long noncoding rna linc00483/microrna-144 regulates radiosensitivity and epithelial-mesenchymal transition in lung adenocarcinoma by interacting with hoxa10. J. Cell. Physiol. 2019, 234, 11805–11821. [Google Scholar] [CrossRef]
- Chen, S.; Li, P.; Li, J.; Wang, Y.; Du, Y.; Chen, X.; Zang, W.; Wang, H.; Chu, H.; Zhao, G.; et al. Mir-144 inhibits proliferation and induces apoptosis and autophagy in lung cancer cells by targeting tigar. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2015, 35, 997–1007. [Google Scholar] [CrossRef]
- Wu, K.L.; Tsai, Y.M.; Lien, C.T.; Kuo, P.L.; Hung, A.J. The roles of microrna in lung cancer. Int. J. Mol. Sci. 2019, 20, 1611. [Google Scholar] [CrossRef]
- Song, L.; Peng, L.; Hua, S.; Li, X.; Ma, L.; Jie, J.; Chen, D.; Wang, Y.; Li, D. Mir-144-5p enhances the radiosensitivity of non-small-cell lung cancer cells via targeting atf2. Biomed. Res. Int. 2018, 2018, 5109497. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Guo, Y.-N.; Shi, K.; Huang, H.-M.; Huang, S.-P.; Xu, W.-Q.; Li, Z.-Y.; Wei, K.-L.; Gan, T.-Q.; Chen, G. Down-regulation of microrna-144-3p and its clinical value in non-small cell lung cancer: A comprehensive analysis based on microarray, mirna-sequencing, and quantitative real-time pcr data. Respir. Res. 2019, 20, 48. [Google Scholar] [CrossRef] [PubMed]
- Mott, F.E. Mesothelioma: A review. Ochsner J. 2012, 12, 70–79. [Google Scholar] [PubMed]
- Yang, H.; Testa, J.R.; Carbone, M. Mesothelioma epidemiology, carcinogenesis, and pathogenesis. Curr. Treat. Options Oncol. 2008, 9, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Rivera, V.; Negrete-García, M.C.; Ávila-Moreno, F.; Ortiz-Quintero, B. Secreted and tissue mirnas as diagnosis biomarkers of malignant pleural mesothelioma. Int. J. Mol. Sci. 2018, 19, 595. [Google Scholar] [CrossRef] [PubMed]
- Guled, M.; Lahti, L.; Lindholm, P.M.; Salmenkivi, K.; Bagwan, I.; Nicholson, A.G.; Knuutila, S.J.G. Cancer. Cdkn2a, nf2, and jun are dysregulated among other genes by mirnas in malignant mesothelioma—A mirna microarray analysis. Genes Chromosomes Cancer 2009, 48, 615–623. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, J.; Fu, H.; Shen, L. Mir-144 functions as a tumor suppressor in breast cancer through inhibiting zeb1/2-mediated epithelial mesenchymal transition process. Onco Targets 2016, 9, 6247–6255. [Google Scholar] [CrossRef]
- Yin, Y.; Cai, J.; Meng, F.; Sui, C.; Jiang, Y. Mir-144 suppresses proliferation, invasion, and migration of breast cancer cells through inhibiting cep55. Cancer Biol. Ther. 2018, 19, 306–315. [Google Scholar] [CrossRef]
- Yu, L.; Yang, Y.; Hou, J.; Zhai, C.; Song, Y.; Zhang, Z.; Qiu, L.; Jia, X. Microrna-144 affects radiotherapy sensitivity by promoting proliferation, migration and invasion of breast cancer cells. Oncol. Rep. 2015, 34, 1845–1852. [Google Scholar] [CrossRef]
- Kahraman, M.; Röske, A.; Laufer, T.; Fehlmann, T.; Backes, C.; Kern, F.; Kohlhaas, J.; Schrörs, H.; Saiz, A.; Zabler, C.; et al. Microrna in diagnosis and therapy monitoring of early-stage triple-negative breast cancer. Sci. Rep. 2018, 8, 11584. [Google Scholar] [CrossRef]
- Madhavan, D.; Peng, C.; Wallwiener, M.; Zucknick, M.; Nees, J.; Schott, S.; Rudolph, A.; Riethdorf, S.; Trumpp, A.; Pantel, K.; et al. Circulating mirnas with prognostic value in metastatic breast cancer and for early detection of metastasis. Carcinogenesis 2016, 37, 461–470. [Google Scholar] [CrossRef]
- Troiano, G.; Caponio, V.C.A.; Adipietro, I.; Tepedino, M.; Santoro, R.; Laino, L.; Lo Russo, L.; Cirillo, N.; Lo Muzio, L. Prognostic significance of cd68+ and cd163+ tumor associated macrophages in head and neck squamous cell carcinoma: A systematic review and meta-analysis. Oral Oncol. 2019, 93, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Lu, J.; Peng, X.; Wang, J.; Liu, X.; Chen, X.; Jiang, Y.; Li, X.; Zhang, B. Integrated analysis of microrna regulatory network in nasopharyngeal carcinoma with deep sequencing. J. Exp. Clin. Cancer Res. 2016, 35, 17. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cui, C.L.; Chen, W.L.; Fu, Z.Y.; Cui, X.Y.; Gong, X. Mir-144 suppresses the growth and metastasis of laryngeal squamous cell carcinoma by targeting irs1. Am. J. Transl. Res. 2016, 8, 1–11. [Google Scholar] [PubMed]
- Salazar-Ruales, C.; Arguello, J.V.; López-Cortés, A.; Cabrera-Andrade, A.; García-Cárdenas, J.M.; Guevara-Ramírez, P.; Peralta, P.; Leone, P.E.; Paz, Y.M.C. Salivary micrornas for early detection of head and neck squamous cell carcinoma: A case-control study in the high altitude mestizo ecuadorian population. Biomed. Res. Int. 2018, 2018, 9792730. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shen, C.; Lu, X.; Hu, C. The incidence and prognosis of nasopharyngeal carcinoma patients with family history. Oncotarget 2017, 8, 97323–97330. [Google Scholar]
- Zhang, L.Y.; Ho-Fun Lee, V.; Wong, A.M.; Kwong, D.L.; Zhu, Y.H.; Dong, S.S.; Kong, K.L.; Chen, J.; Tsao, S.W.; Guan, X.Y.; et al. Microrna-144 promotes cell proliferation, migration and invasion in nasopharyngeal carcinoma through repression of pten. Carcinogenesis 2013, 34, 454–463. [Google Scholar] [CrossRef]
- Song, L.; Chen, L.; Luan, Q.; Kong, Q. Mir-144-3p facilitates nasopharyngeal carcinoma via crosstalk with pten. J. Cell. Physiol. 2019, 234, 17912–17924. [Google Scholar] [CrossRef]
- Wu, C.W.; Wang, S.G.; Lin, M.L.; Chen, S.S. Downregulation of mir-144 by triptolide enhanced p85alpha-pten complex formation causing s phase arrest of human nasopharyngeal carcinoma cells. Eur. J. Pharmacol. 2019, 855, 137–148. [Google Scholar] [CrossRef]
- Nguyen, Q.T.; Lee, E.J.; Huang, M.G.; Park, Y.I.; Khullar, A.; Plodkowski, R.A. Diagnosis and treatment of patients with thyroid cancer. Am. Health Drug Benefits 2015, 8, 30–40. [Google Scholar]
- Rossing, M.; Borup, R.; Henao, R.; Winther, O.; Vikesaa, J.; Niazi, O.; Godballe, C.; Krogdahl, A.; Glud, M.; Hjort-Sorensen, C.; et al. Down-regulation of micrornas controlling tumourigenic factors in follicular thyroid carcinoma. J. Mol. Endocrinol. 2012, 48, 11–23. [Google Scholar] [CrossRef]
- Guan, H.; Liang, W.; Xie, Z.; Li, H.; Liu, J.; Liu, L.; Xiu, L.; Li, Y. Down-regulation of mir-144 promotes thyroid cancer cell invasion by targeting zeb1 and zeb2. Endocrine 2015, 48, 566–574. [Google Scholar] [CrossRef]
- Sun, W.; Lan, X.; Wang, Z.; Dong, W.; He, L.; Zhang, T.; Zhang, P.; Zhang, H. Microrna-144 inhibits proliferation by targeting ww domain-containing transcription regulator protein 1 in papillary thyroid cancer. Oncol. Lett. 2018, 15, 1007–1013. [Google Scholar] [CrossRef]
- Liu, C.; Su, C.; Chen, Y.; Li, G. Mir-144-3p promotes the tumor growth and metastasis of papillary thyroid carcinoma by targeting paired box gene 8. Cancer Cell Int. 2018, 18, 54. [Google Scholar] [CrossRef] [PubMed]
- Wirsching, H.G.; Galanis, E.; Weller, M. Glioblastoma. Handb. Clin. Neurol. 2016, 134, 381–397. [Google Scholar] [PubMed]
- Areeb, Z.; Stylli, S.; Koldej, R.; Ritchie, D.; Siegal, T.; Morokoff, A.; Kaye, A.; Luwor, R. Microrna as potential biomarkers in glioblastoma. J. Neuro-Oncol. 2015, 125, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Lan, F.; Yu, H.; Hu, M.; Xia, T.; Yue, X. Mir-144-3p exerts anti-tumor effects in glioblastoma by targeting c-met. J. Neurochem. 2015, 135, 274–286. [Google Scholar] [CrossRef]
- Song, J.; Ma, Q.; Hu, M.; Qian, D.; Wang, B.; He, N. The inhibition of mir-144-3p on cell proliferation and metastasis by targeting top2a in hcmv-positive glioblastoma cells. Molecules 2018, 23, 3259. [Google Scholar] [CrossRef]
- Cheng, Z.X.; Song, Y.X.; Wang, Z.Y.; Wang, Y.; Dong, Y. Mir-144-3p serves as a tumor suppressor by targeting fzd7 and predicts the prognosis of human glioblastoma. Eur. Rev. Med. Pharm. Sci. 2017, 21, 4079–4086. [Google Scholar]
- Liu, N.; Tu, Y. Systematic review of micrornas and its therapeutic potential in glioma. Cancer Transl. Med. 2015, 1, 50–66. [Google Scholar]
- Cardoso, A.M.S.; Sousa, M.; Morais, C.M.; Oancea-Castillo, L.R.; Regnier-Vigouroux, A.; Rebelo, O.; Tao, H.; Barbosa, M.; Pedroso de Lima, M.C.; Jurado, A.S. Mir-144 overexpression as a promising therapeutic strategy to overcome glioblastoma cell invasiveness and resistance to chemotherapy. Hum. Mol. Genet. 2019. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Ren, J.J.; Zhao, J.L.; Zang, J.; Long, Q.F.; Du, J.J.; Jia, X.T.; Gu, N.B.; Di, Z.L.; Qian, Y.H.; et al. Microrna-144 represses gliomas progression and elevates susceptibility to temozolomide by targeting cav2 and fgf7. Sci. Rep. 2020, 10, 4155. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowska, D.; Kluska, P.; Reich, A. Beyond pd-1 immunotherapy in malignant melanoma. Dermatol. Ther. 2019, 9, 243–257. [Google Scholar] [CrossRef]
- Peng, Y.-G.; Zhang, L. Baohuoside-i suppresses cell proliferation and migration by up-regulating mir-144 in melanoma. Pharm Biol. 2017, 56, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Bian, G.; Meng, Z.; Dang, G.; Shi, D.; Mi, S. Mir-144 inhibits uveal melanoma cell proliferation and invasion by regulating c-met expression. PLoS ONE 2015, 10, e0124428. [Google Scholar] [CrossRef] [PubMed]
- Misaghi, A.; Goldin, A.; Awad, M.; Kulidjian, A.A. Osteosarcoma: A comprehensive review. Sicot J. 2018, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Namlos, H.M.; Meza-Zepeda, L.A.; Baroy, T.; Ostensen, I.H.; Kresse, S.H.; Kuijjer, M.L.; Serra, M.; Burger, H.; Cleton-Jansen, A.M.; Myklebost, O. Modulation of the osteosarcoma expression phenotype by micrornas. PLoS ONE 2012, 7, e48086. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Huang, J.; Gui, K.; Xiong, M.; Cai, G.; Xu, J.; Wang, K.; Liu, D.; Zhang, X.; Yin, W. The downregulation of mir-144 is associated with the growth and invasion of osteosarcoma cells through the regulation of tagln expression. Int. J. Mol. Med. 2014, 34, 1565–1572. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, X.; Wei, M. Microrna-144 suppresses osteosarcoma growth and metastasis by targeting rock1 and rock2. Oncotarget 2015, 6, 10297–10308. [Google Scholar] [CrossRef]
- Yu, H.; Sun, H.; Bai, Y.; Han, J.; Liu, G.; Liu, Y.; Zhang, N. Mef2d overexpression contributes to the progression of osteosarcoma. Gene 2015, 563, 130–135. [Google Scholar] [CrossRef]
- Cui, S.Q.; Wang, H. Microrna-144 inhibits the proliferation, apoptosis, invasion, and migration of osteosarcoma cell line f5m2. Tumour Biol. J. Int. Soc. Oncodevelopmental. Biol. Med. 2015, 36, 6949–6958. [Google Scholar] [CrossRef]
- Ren, Y.F.; Zhang, T.H.; Zhong, S.; Zhao, Y.T.; Lv, Y.N. Mir-144 suppresses proliferation and induces apoptosis of osteosarcoma cells via direct regulation of mtor expression. Oncol. Lett. 2018, 15, 1163–1169. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kooshkaki, O.; Rezaei, Z.; Rahmati, M.; Vahedi, P.; Derakhshani, A.; Brunetti, O.; Baghbanzadeh, A.; Mansoori, B.; Silvestris, N.; Baradaran, B. MiR-144: A New Possible Therapeutic Target and Diagnostic/Prognostic Tool in Cancers. Int. J. Mol. Sci. 2020, 21, 2578. https://doi.org/10.3390/ijms21072578
Kooshkaki O, Rezaei Z, Rahmati M, Vahedi P, Derakhshani A, Brunetti O, Baghbanzadeh A, Mansoori B, Silvestris N, Baradaran B. MiR-144: A New Possible Therapeutic Target and Diagnostic/Prognostic Tool in Cancers. International Journal of Molecular Sciences. 2020; 21(7):2578. https://doi.org/10.3390/ijms21072578
Chicago/Turabian StyleKooshkaki, Omid, Zohre Rezaei, Meysam Rahmati, Parviz Vahedi, Afshin Derakhshani, Oronzo Brunetti, Amir Baghbanzadeh, Behzad Mansoori, Nicola Silvestris, and Behzad Baradaran. 2020. "MiR-144: A New Possible Therapeutic Target and Diagnostic/Prognostic Tool in Cancers" International Journal of Molecular Sciences 21, no. 7: 2578. https://doi.org/10.3390/ijms21072578
APA StyleKooshkaki, O., Rezaei, Z., Rahmati, M., Vahedi, P., Derakhshani, A., Brunetti, O., Baghbanzadeh, A., Mansoori, B., Silvestris, N., & Baradaran, B. (2020). MiR-144: A New Possible Therapeutic Target and Diagnostic/Prognostic Tool in Cancers. International Journal of Molecular Sciences, 21(7), 2578. https://doi.org/10.3390/ijms21072578