The Optogenetic Revolution in Cerebellar Investigations


Abstract
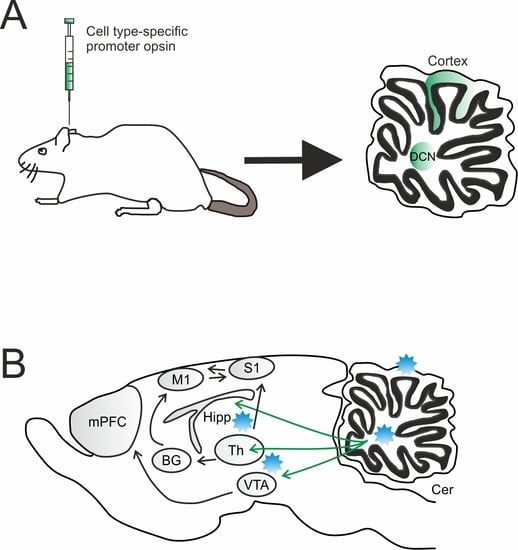
1. Introduction
2. Brief Overview of the Cerebellar Anatomy and Microcircuits Organization
3. Pros and Cons of Optogenetics
4. Sensorimotor Functions
4.1. Sensorimotor Integration and Voluntary Movement
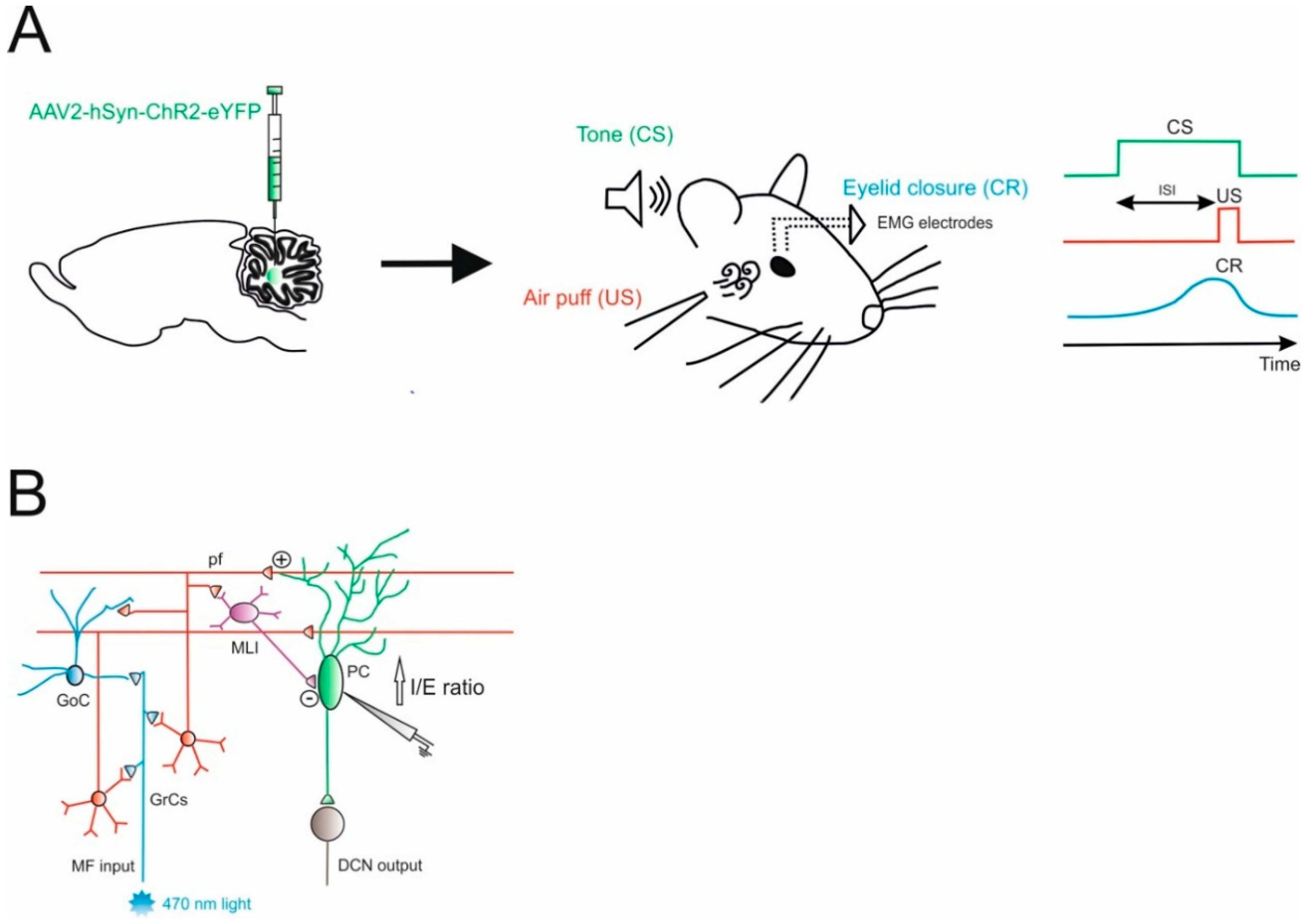
4.2. Associative Learning (Eyeblink Conditioning)
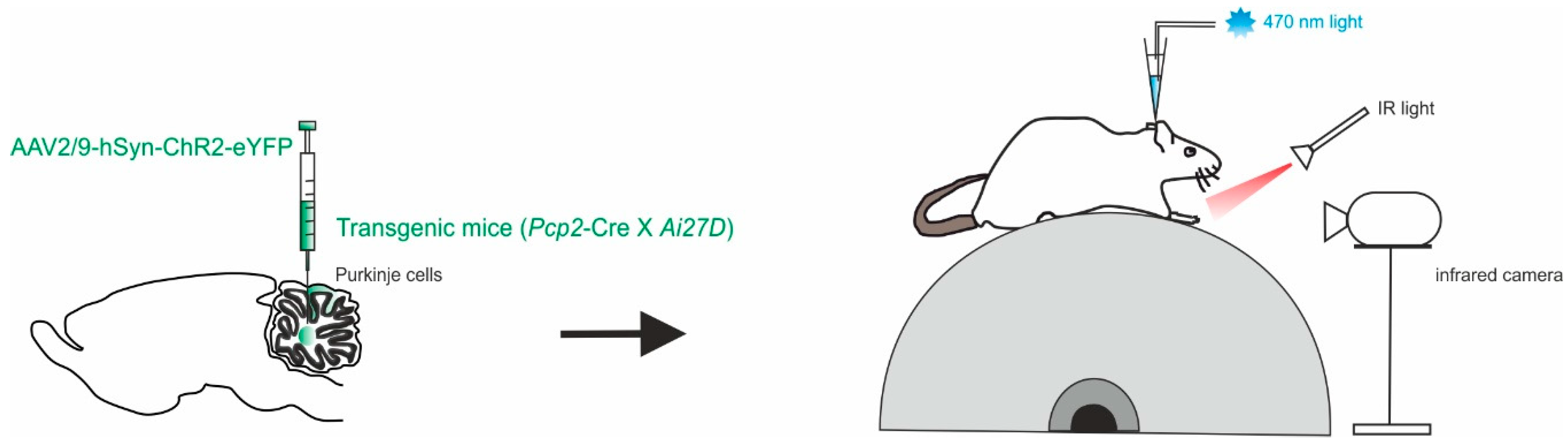
4.3. Eye Movements in Monkeys
4.4. Movement Kinematics
4.5. Movement Disorders
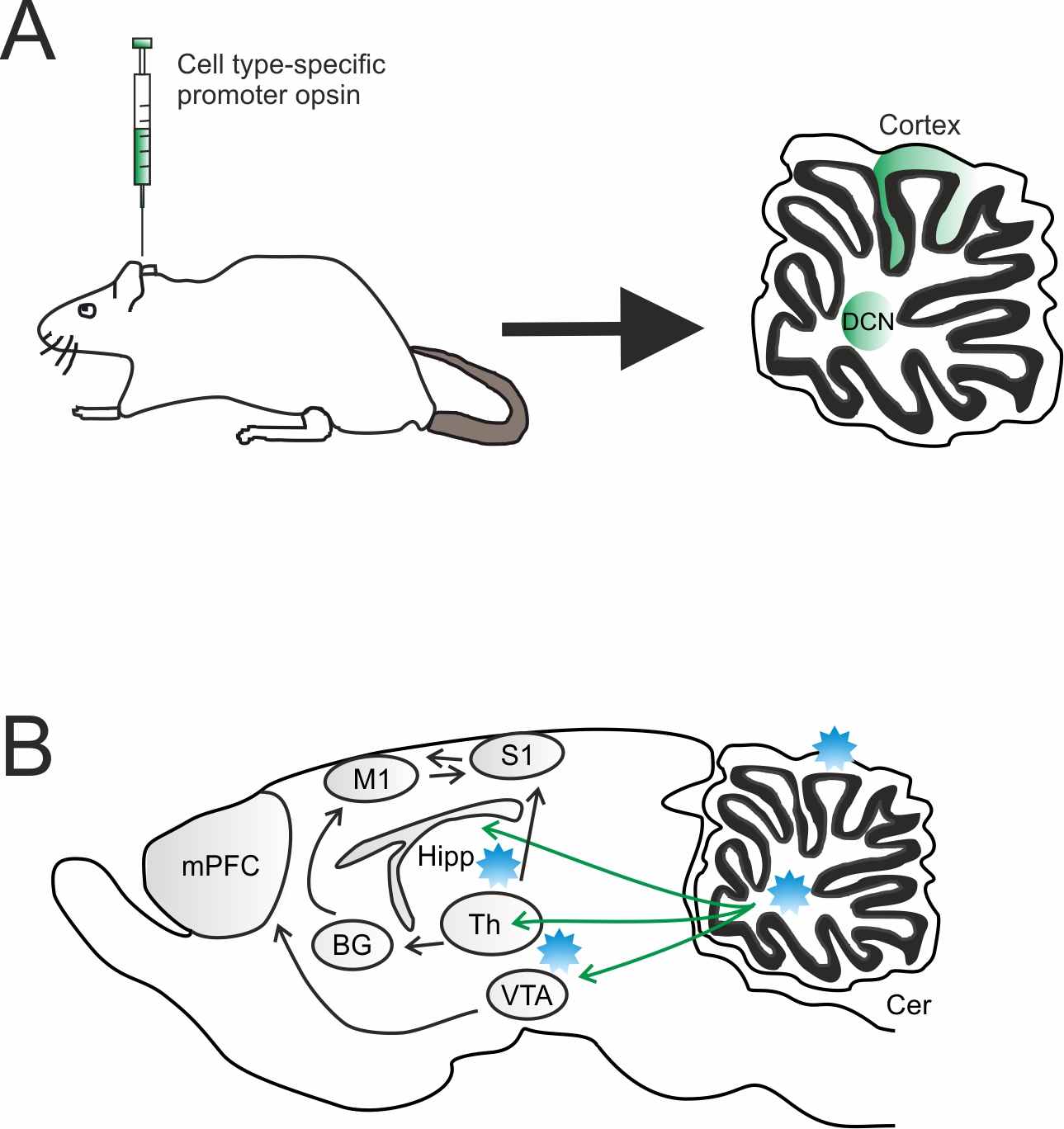
5. Non-Sensorimotor Functions
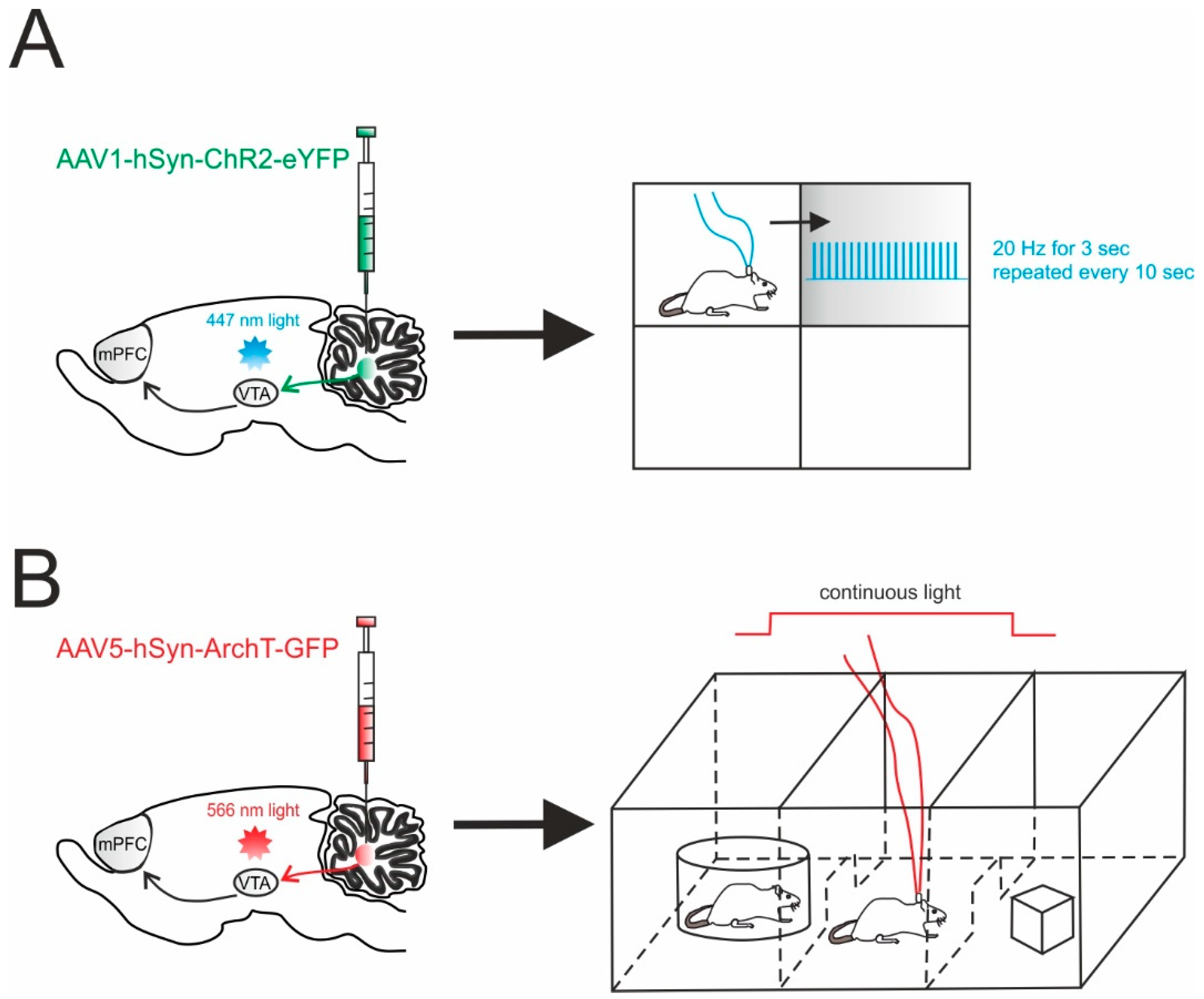
5.1. Reward and Social Behavior
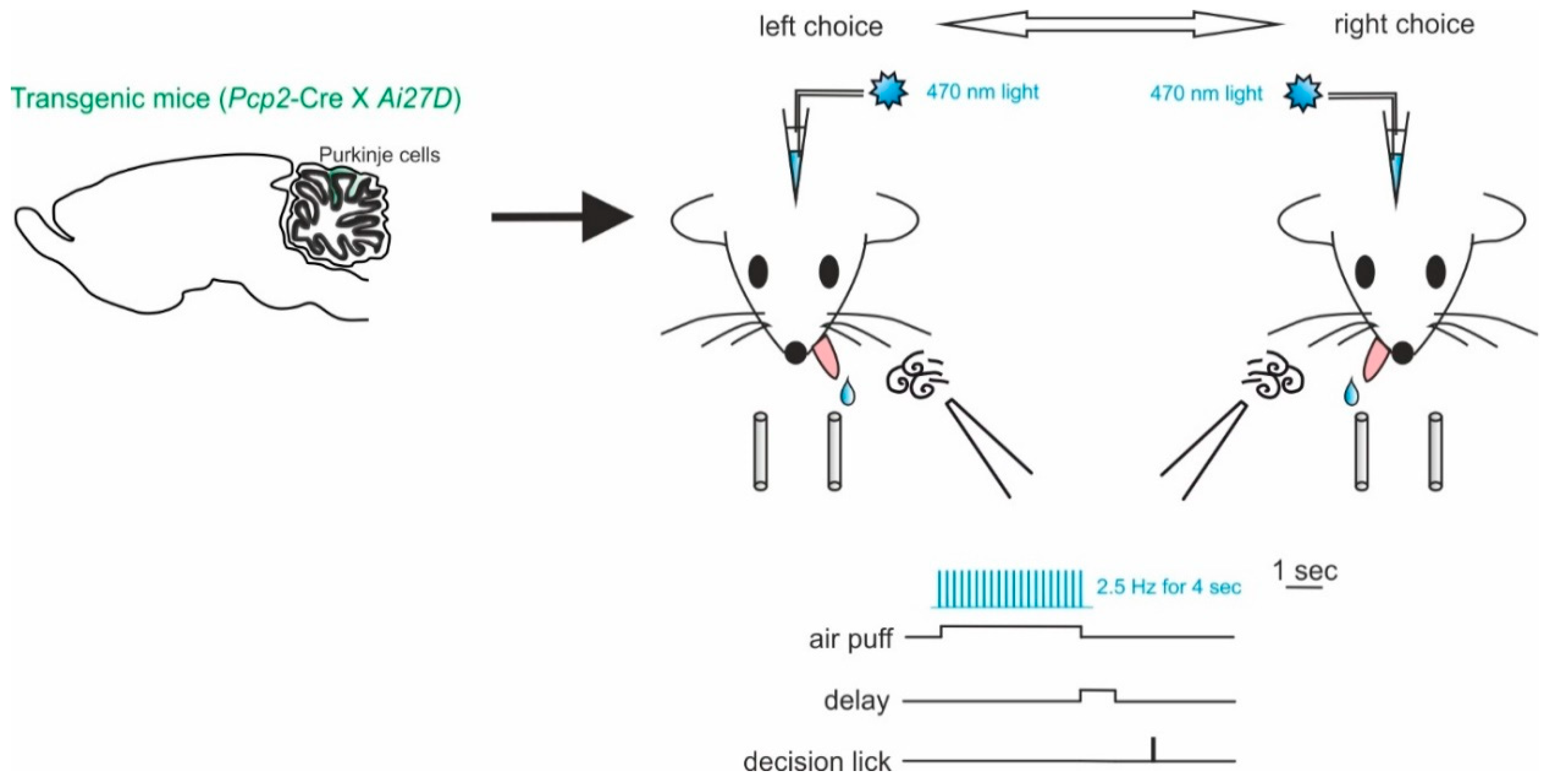
5.2. Working Memory and Decision-Making
5.3. Schizophrenia and Cognition
5.4. Temporal Lobe Epilepsy and Absence Seizure
5.5. Control of Blood Pressure
6. Clinical Aspects
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AAV | Adeno-associated viruses |
| Arch | Archaerhodopsin |
| ASD | Autism Spectrum Disorders |
| ChR2 | Channelrhodopsin-2 |
| CS | Conditioned stimulus |
| DCN | Deep Cerebellar Nuclei |
| EBC | Eyeblink conditioning |
| eNPHR3.0 | Enhanced Natronomonas halorhodopsin |
| GSWDs | Generalized spike-and-wave discharges |
| NPHR | Halorhodopsin |
| PC | Purkinje cell |
| TLE | Temporal lobe epilepsy |
| TMS | Transcranial magnetic resonance |
| US | Unconditioned stimulus |
| VTA | Ventral Tegmental Area |
| YFP | Yellow Fluorescent Protein |
References
- Malacarne, V. Nuova Esposizione della Vera Struttura del Cervelletto Umano; Briolo, Ed.; Appresso Giammichele Briolo nella contrada de’guardinfanti: Torino, Italy, 1776; p. 129. [Google Scholar]
- Flourens, P. Recherchers Experimentales sur le Proprietes et les Functions du Systeme Nerveux dans les Animaux Vertebres; Crevot: Paris, France, 1824. [Google Scholar]
- Schmahmann, J.D.; Sherman, J.C. The cerebellar cognitive affective syndrome. Brain 1998, 121 Pt 4, 561–579. [Google Scholar] [CrossRef]
- Schmahmann, J.D. Dysmetria of thought: Clinical consequences of cerebellar dysfunction on cognition and affect. Trends Cogn. Sci. 1998, 2, 362–371. [Google Scholar] [CrossRef]
- Timmann, D. Contribution of the cerebellum to cognition. Fortschr. Neurol. Psychiatr. 2012, 80, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Baillieux, H.; De Smet, H.J.; Paquier, P.F.; De Deyn, P.P.; Mariën, P. Cerebellar neurocognition: Insights into the bottom of the brain. Clin. Neurol. Neurosurg. 2008, 110, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Gottwald, B.; Wilde, B.; Mihajlovic, Z.; Mehdorn, H.M. Evidence for distinct cognitive deficits after focal cerebellar lesions. J. Neurol. Neurosurg. Psychiatry 2004, 75, 1524–1531. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D. Disorders of the cerebellum: Ataxia, dysmetria of thought, and the cerebellar cognitive affective syndrome. J. Neuropsychiatry Clin. Neurosci. 2004, 16, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Manto, M. Cerebellar motor syndrome from children to the elderly. Handb. Clin. Neurol. 2018, 154, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Prestori, F.; Mapelli, L.; D’Angelo, E. Diverse Neuron Properties and Complex Network Dynamics in the Cerebellar Cortical Inhibitory Circuit. Front. Mol. Neurosci. 2019, 12, 267. [Google Scholar] [CrossRef]
- D’Angelo, E.; Solinas, S.; Mapelli, J.; Gandolfi, D.; Mapelli, L.; Prestori, F. The cerebellar Golgi cell and spatiotemporal organization of granular layer activity. Front. Neural Circuits 2013, 7, 93. [Google Scholar] [CrossRef]
- Mapelli, L.; Solinas, S.; D’Angelo, E. Integration and regulation of glomerular inhibition in the cerebellar granular layer circuit. Front. Cell. Neurosci. 2014, 8, 55. [Google Scholar] [CrossRef]
- Apps, R.; Hawkes, R.; Aoki, S.; Bengtsson, F.; Brown, A.M.; Chen, G.; Ebner, T.J.; Isope, P.; Jörntell, H.; Lackey, E.P.; et al. Cerebellar Modules and Their Role as Operational Cerebellar Processing Units: A Consensus paper [corrected]. Cerebellum 2018, 17, 654–682. [Google Scholar] [CrossRef]
- Leto, K.; Arancillo, M.; Becker, E.B.; Buffo, A.; Chiang, C.; Ding, B.; Dobyns, W.B.; Dusart, I.; Haldipur, P.; Hatten, M.E.; et al. Consensus Paper: Cerebellar Development. Cerebellum 2016, 15, 789–828. [Google Scholar] [CrossRef] [PubMed]
- Marzban, H.; Del Bigio, M.R.; Alizadeh, J.; Ghavami, S.; Zachariah, R.M.; Rastegar, M. Cellular commitment in the developing cerebellum. Front. Cell. Neurosci. 2014, 8, 450. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.K.; Adhikari, A.; Deisseroth, K. Integration of optogenetics with complementary methodologies in systems neuroscience. Nat. Rev. Neurosci. 2017, 18, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Proville, R.D.; Spolidoro, M.; Guyon, N.; Dugue, G.P.; Selimi, F.; Isope, P.; Popa, D.; Lena, C. Cerebellum involvement in cortical sensorimotor circuits for the control of voluntary movements. Nat. Neurosci. 2014, 17, 1233–1239. [Google Scholar] [CrossRef]
- Kelly, R.M.; Strick, P.L. Cerebellar loops with motor cortex and prefrontal cortex of a nonhuman primate. J. Neurosci. 2003, 23, 8432–8444. [Google Scholar] [CrossRef]
- Medina, J.F.; Nores, W.L.; Ohyama, T.; Mauk, M.D. Mechanisms of cerebellar learning suggested by eyelid conditioning. Curr. Opin. Neurobiol. 2000, 10, 717–724. [Google Scholar] [CrossRef]
- Kim, J.J.; Thompson, R.F. Cerebellar circuits and synaptic mechanisms involved in classical eyeblink conditioning. Trends Neurosci. 1997, 20, 177–181. [Google Scholar] [CrossRef]
- De Zeeuw, C.I.; Yeo, C.H. Time and tide in cerebellar memory formation. Curr. Opin. Neurobiol. 2005, 15, 667–674. [Google Scholar] [CrossRef]
- Carey, M.R. Synaptic mechanisms of sensorimotor learning in the cerebellum. Curr. Opin. Neurobiol. 2011, 21, 609–615. [Google Scholar] [CrossRef]
- Aiba, A.; Kano, M.; Chen, C.; Stanton, M.E.; Fox, G.D.; Herrup, K.; Zwingman, T.A.; Tonegawa, S. Deficient cerebellar long-term depression and impaired motor learning in mGluR1 mutant mice. Cell 1994, 79, 377–388. [Google Scholar] [PubMed]
- Ito, M.; Yamaguchi, K.; Nagao, S.; Yamazaki, T. Long-term depression as a model of cerebellar plasticity. Prog. Brain Res. 2014, 210, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Schonewille, M.; Belmeguenai, A.; Koekkoek, S.K.; Houtman, S.H.; Boele, H.J.; van Beugen, B.J.; Gao, Z.; Badura, A.; Ohtsuki, G.; Amerika, W.E.; et al. Purkinje cell-specific knockout of the protein phosphatase PP2B impairs potentiation and cerebellar motor learning. Neuron 2010, 67, 618–628. [Google Scholar] [CrossRef] [PubMed]
- Schonewille, M.; Gao, Z.; Boele, H.J.; Veloz, M.F.V.; Amerika, W.E.; Simek, A.A.; De Jeu, M.T.; Steinberg, J.P.; Takamiya, K.; Hoebeek, F.E.; et al. Reevaluating the Role of LTD in Cerebellar Motor Learning. Neuron 2011, 70, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.P.; Yamaguchi, H.; Zeng, X.H.; Kojo, M.; Nakada, Y.; Takagi, A.; Sugimori, M.; Llinas, R.R. Normal motor learning during pharmacological prevention of Purkinje cell long-term depression. Proc. Natl. Acad. Sci. USA 2005, 102, 17166–17171. [Google Scholar] [CrossRef] [PubMed]
- Grasselli, G.; Boele, H.J.; Titley, H.K.; Bradford, N.; van Beers, L.; Jay, L.; Beekhof, G.C.; Busch, S.E.; De Zeeuw, C.I.; Schonewille, M.; et al. SK2 channels in cerebellar Purkinje cells contribute to excitability modulation in motor-learning-specific memory traces. PLoS Biol. 2020, 18, e3000596. [Google Scholar] [CrossRef]
- Johansson, F.; Jirenhed, D.A.; Rasmussen, A.; Zucca, R.; Hesslow, G. Memory trace and timing mechanism localized to cerebellar Purkinje cells. Proc. Natl. Acad. Sci. USA 2014, 111, 14930–14934. [Google Scholar] [CrossRef]
- Gao, Z.; van Beugen, B.J.; De Zeeuw, C.I. Distributed synergistic plasticity and cerebellar learning. Nat. Rev. Neurosci. 2012, 13, 619–635. [Google Scholar] [CrossRef]
- ten Brinke, M.; Boele, H.J.; Spanke, J.; Potters, J.W.; Kornysheva, K.; Wulff, P.; Anna, I.J.; Koekkoek, S.; De Zeeuw, C. Evolving Models of Pavlovian Conditioning: Cerebellar Cortical Dynamics in Awake Behaving Mice. Cell Rep. 2015, 13, 1977–1988. [Google Scholar] [CrossRef]
- Steinmetz, J.E.; Rosen, D.J.; Chapman, P.F.; Lavond, D.G.; Thompson, R.F. Classical conditioning of the rabbit eyelid response with a mossy-fiber stimulation CS: I. Pontine nuclei and middle cerebellar peduncle stimulation. Behav. Neurosci. 1986, 100, 878–887. [Google Scholar] [CrossRef]
- Albergaria, C.; Silva, N.T.; Pritchett, D.L.; Carey, M.R. Locomotor activity modulates associative learning in mouse cerebellum. Nat. Neurosci. 2018, 21, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Proietti-Onori, M.; Lin, Z.; Ten Brinke, M.M.; Boele, H.J.; Potters, J.W.; Ruigrok, T.J.; Hoebeek, F.E.; De Zeeuw, C.I. Excitatory Cerebellar Nucleocortical Circuit Provides Internal Amplification during Associative Conditioning. Neuron 2016, 89, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Casellato, C.; Antonietti, A.; Garrido, J.A.; Ferrigno, G.; D’Angelo, E.; Pedrocchi, A. Distributed cerebellar plasticity implements generalized multiple-scale memory components in real-robot sensorimotor tasks. Front. Comput. Neurosci. 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, D.B.; Yao, J.; Sun, L.; Wu, B.; Li, X.; Liu, S.L.; Hou, J.M.; Liu, H.L.; Sui, J.F.; Wu, G.Y. Reevaluating the ability of cerebellum in associative motor learning. Sci. Rep. 2019, 9, 6029. [Google Scholar] [CrossRef]
- El-Shamayleh, Y.; Kojima, Y.; Soetedjo, R.; Horwitz, G.D. Selective Optogenetic Control of Purkinje Cells in Monkey Cerebellum. Neuron 2017, 95, 51–62.e4. [Google Scholar] [CrossRef]
- Noda, H.; Fujikado, T. Involvement of Purkinje cells in evoking saccadic eye movements by microstimulation of the posterior cerebellar vermis of monkeys. J. Neurophysiol. 1987, 57, 1247–1261. [Google Scholar] [CrossRef]
- McElligott, J.G.; Keller, E.L. Cerebellar vermis involvement in monkey saccadic eye movements: Microstimulation. Exp. Neurol. 1984, 86, 543–558. [Google Scholar] [CrossRef]
- Ron, S.; Robinson, D.A. Eye movements evoked by cerebellar stimulation in the alert monkey. J. Neurophysiol. 1973, 36, 1004–1022. [Google Scholar] [CrossRef]
- Fujikado, T.; Noda, H. Saccadic eye movements evoked by microstimulation of lobule VII of the cerebellar vermis of macaque monkeys. J. Physiol. 1987, 394, 573–594. [Google Scholar] [CrossRef]
- Cavanaugh, J.; Monosov, I.; McAlonan, K.; Berman, R.A.; Smith, M.K.; Cao, V.; Wang, K.H.; Boyden, E.S.; Wurtz, R.H. Optogenetic Inactivation Modifies Monkey Visuomotor Behavior. Neuron 2012, 76, 901–907. [Google Scholar] [CrossRef]
- Diester, I.; Kaufman, M.T.; Mogri, M.; Pashaie, R.; Goo, W.; Yizhar, O.; Ramakrishnan, C.; Deisseroth, K.; Shenoy, K.V. An optogenetic toolbox designed for primates. Nat. Neurosci. 2011, 14, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Han, X. Optogenetics in the nonhuman primate. Prog. Brain Res. 2012, 196, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Qian, X.; Bernstein, J.G.; Zhou, H.; Franzesi, G.T.; Stern, P.; Bronson, R.T.; Graybiel, A.M.; Desimone, R.; Boyden, E.S. Millisecond-Timescale Optical Control of Neural Dynamics in the Nonhuman Primate Brain. Neuron 2009, 62, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Ohayon, S.; Grimaldi, P.; Schweers, N.; Tsao, D.Y. Saccade Modulation by Optical and Electrical Stimulation in the Macaque Frontal Eye Field. J. Neurosci. 2013, 33, 16684–16697. [Google Scholar] [CrossRef]
- Histed, M.H.; Bonin, V.; Reid, R.C. Direct activation of sparse, distributed populations of cortical neurons by electrical microstimulation. Neuron 2009, 63, 508–522. [Google Scholar] [CrossRef]
- Hass, C.A.; Glickfeld, L.L. High-fidelity optical excitation of cortico-cortical projections at physiological frequencies. J. Neurophysiol. 2016, 116, 2056–2066. [Google Scholar] [CrossRef]
- Jackman, S.L.; Beneduce, B.M.; Drew, I.R.; Regehr, W.G. Achieving High-Frequency Optical Control of Synaptic Transmission. J. Neurosci. 2014, 34, 7704–7714. [Google Scholar] [CrossRef] [PubMed]
- Payne, H.L.; French, R.L.; Guo, C.C.; Nguyen-Vu, T.B.; Manninen, T.; Raymond, J.L. Cerebellar Purkinje cells control eye movements with a rapid rate code that is invariant to spike irregularity. eLife 2019, 8. [Google Scholar] [CrossRef]
- Heiney, S.A.; Kim, J.; Augustine, G.J.; Medina, J.F. Precise Control of Movement Kinematics by Optogenetic Inhibition of Purkinje Cell Activity. J. Neurosci. 2014, 34, 2321–2330. [Google Scholar] [CrossRef]
- Tsuda, S.; Kee, M.Z.; Cunha, C.; Kim, J.; Yan, P.; Loew, L.M.; Augustine, G.J. Probing the function of neuronal populations: Combining micromirror-based optogenetic photostimulation with voltage-sensitive dye imaging. Neurosci. Res. 2013, 75, 76–81. [Google Scholar] [CrossRef]
- Perrett, S.; Ruiz, B.; Mauk, M. Cerebellar cortex lesions disrupt learning-dependent timing of conditioned eyelid responses. J. Neurosci. 1993, 13, 1708–1718. [Google Scholar] [CrossRef] [PubMed]
- Koekkoek, S.K.; Hulscher, H.C.; Dortland, B.R.; Hensbroek, R.A.; Elgersma, Y.; Ruigrok, T.J.; De Zeeuw, C.I. Cerebellar LTD and learning-dependent timing of conditioned eyelid responses. Science 2003, 301, 1736–1739. [Google Scholar] [CrossRef] [PubMed]
- Jelitai, M.; Puggioni, P.; Ishikawa, T.; Rinaldi, A.; Duguid, I. Dendritic excitation-inhibition balance shapes cerebellar output during motor behaviour. Nat. Commun. 2016, 7, 13722. [Google Scholar] [CrossRef] [PubMed]
- Hoogland, T.; De Gruijl, J.; Witter, L.; Canto, C.; De Zeeuw, C. Role of Synchronous Activation of Cerebellar Purkinje Cell Ensembles in Multi-joint Movement Control. Curr. Biol. 2015, 25, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Sarnaik, R.; Raman, I.M. Control of voluntary and optogenetically perturbed locomotion by spike rate and timing of neurons of the mouse cerebellar nuclei. eLife 2018, 7, e29546. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Yoshida, M.; Obata, K. Monosynaptic inhibition of the intracerebellar nuclei induced rom the cerebellar cortex. Experientia 1964, 20, 575–576. [Google Scholar] [CrossRef]
- Gilbert, P.F.; Thach, W.T. Purkinje cell activity during motor learning. Brain Res. 1977, 128, 309–328. [Google Scholar] [CrossRef]
- Marr, D. A theory of cerebellar cortex. J. Physiol. 1969, 202, 437–470. [Google Scholar] [CrossRef]
- Medina, J.F.; Mauk, M.D. Simulations of Cerebellar Motor Learning: Computational Analysis of Plasticity at the Mossy Fiber to Deep Nucleus Synapse. J. Neurosci. 1999, 19, 7140–7151. [Google Scholar] [CrossRef]
- Lee, K.H.; Mathews, P.J.; Reeves, A.M.; Choe, K.Y.; Jami, S.A.; Serrano, R.E.; Otis, T.S. Circuit Mechanisms Underlying Motor Memory Formation in the Cerebellum. Neuron 2015, 86, 529–540. [Google Scholar] [CrossRef]
- Witter, L.; Canto, C.B.; Hoogland, T.M.; de Gruijl, J.R.; De Zeeuw, C.I. Strength and timing of motor responses mediated by rebound firing in the cerebellar nuclei after Purkinje cell activation. Front. Neural Circuits 2013, 7, 133. [Google Scholar] [CrossRef] [PubMed]
- ten Brinke, M.M.; Heiney, S.A.; Wang, X.; Proietti-Onori, M.; Boele, H.J.; Bakermans, J.; Medina, J.F.; Gao, Z.; De Zeeuw, C.I. Dynamic modulation of activity in cerebellar nuclei neurons during pavlovian eyeblink conditioning in mice. eLife 2017, 6, e28132. [Google Scholar] [CrossRef] [PubMed]
- Goodkin, H.P.; Thach, W.T. Cerebellar control of constrained and unconstrained movements. I. Nuclear inactivation. J. Neurophysiol. 2003, 89, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Mason, C.R.; Miller, L.E.; Baker, J.F.; Houk, J.C. Organization of reaching and grasping movements in the primate cerebellar nuclei as revealed by focal muscimol inactivations. J. Neurophysiol. 1998, 79, 537–554. [Google Scholar] [CrossRef]
- Glickstein, M. Cerebellar agenesis. Brain 1994, 117 Pt 5, 1209–1212. [Google Scholar] [CrossRef]
- Mark, M.D.; Krause, M.; Boele, H.J.; Kruse, W.; Pollok, S.; Kuner, T.; Dalkara, D.; Koekkoek, S.; De Zeeuw, C.I.; Herlitze, S. Spinocerebellar Ataxia Type 6 Protein Aggregates Cause Deficits in Motor Learning and Cerebellar Plasticity. J. Neurosci. 2015, 35, 8882–8895. [Google Scholar] [CrossRef]
- Vinueza Veloz, M.F.; Zhou, K.; Bosman, L.W.J.; Potters, J.W.; Negrello, M.; Seepers, R.M.; Strydis, C.; Koekkoek, S.K.E.; De Zeeuw, C.I. Cerebellar control of gait and interlimb coordination. Brain Struct. Funct. 2015, 220, 3513–3536. [Google Scholar] [CrossRef]
- Yu, F.; Jiang, Q.; Sun, X.; Zhang, R. A new case of complete primary cerebellar agenesis: Clinical and imaging findings in a living patient. Brain 2015, 138, e353. [Google Scholar] [CrossRef]
- Chen, C.H.; Fremont, R.; Arteaga-Bracho, E.E.; Khodakhah, K. Short latency cerebellar modulation of the basal ganglia. Nat. Neurosci. 2014, 17, 1767–1775. [Google Scholar] [CrossRef]
- Doya, K. What are the computations of the cerebellum, the basal ganglia and the cerebral cortex? Neural Netw. 1999, 12, 961–974. [Google Scholar] [CrossRef]
- Doya, K. Complementary roles of basal ganglia and cerebellum in learning and motor control. Curr. Opin. Neurobiol. 2000, 10, 732–739. [Google Scholar] [CrossRef]
- Albin, R.L.; Young, A.B.; Penney, J.B. The functional anatomy of basal ganglia disorders. Trends Neurosci. 1989, 12, 366–375. [Google Scholar] [CrossRef]
- Menardy, F.; Varani, A.P.; Combes, A.; Lena, C.; Popa, D. Functional Alteration of Cerebello-Cerebral Coupling in an Experimental Mouse Model of Parkinson’s Disease. Cereb. Cortex 2019, 29, 1752–1766. [Google Scholar] [CrossRef] [PubMed]
- Molnar, G.F.; Sailer, A.; Gunraj, C.A.; Lang, A.E.; Lozano, A.M.; Chen, R. Thalamic deep brain stimulation activates the cerebellothalamocortical pathway. Neurology 2004, 63, 907–909. [Google Scholar] [CrossRef]
- Ni, Z.; Pinto, A.D.; Lang, A.E.; Chen, R. Involvement of the cerebellothalamocortical pathway in Parkinson disease. Ann. Neurol. 2010, 68, 816–824. [Google Scholar] [CrossRef]
- Carrillo, F.; Palomar, F.J.; Conde, V.; Diaz-Corrales, F.J.; Porcacchia, P.; Fernandez-Del-Olmo, M.; Koch, G.; Mir, P. Study of cerebello-thalamocortical pathway by transcranial magnetic stimulation in Parkinson’s disease. Brain Stimul. 2013, 6, 582–589. [Google Scholar] [CrossRef]
- Timmermann, L.; Gross, J.; Dirks, M.; Volkmann, J.; Freund, H.J.; Schnitzler, A. The cerebral oscillatory network of parkinsonian resting tremor. Brain 2003, 126, 199–212. [Google Scholar] [CrossRef]
- Helmich, R.C.; Hallett, M.; Deuschl, G.; Toni, I.; Bloem, B.R. Cerebral causes and consequences of parkinsonian resting tremor: A tale of two circuits? Brain 2012, 135, 3206–3226. [Google Scholar] [CrossRef]
- Llinas, R.; Sasaki, K. The Functional Organization of the Olivo-Cerebellar System as Examined by Multiple Purkinje Cell Recordings. Eur. J. Neurosci. 1989, 1, 587–602. [Google Scholar] [CrossRef]
- Welsh, J.P.; Lang, E.J.; Suglhara, I.; Llinas, R. Dynamic organization of motor control within the olivocerebellar system. Nature 1995, 374, 453–457. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, J.; Shi, Q.; Li, M.; Wang, C.; Wang, X.; Wang, W.; Wu, Y. The Neuronal Activation of Deep Cerebellar Nuclei Is Essential for Environmental Enrichment-Induced Post-Stroke Motor Recovery. Aging Dis. 2019, 10, 530–543. [Google Scholar] [CrossRef]
- Elias, G.J.B.; Namasivayam, A.A.; Lozano, A.M. Deep brain stimulation for stroke: Current uses and future directions. Brain Stimul. 2018, 11, 3–28. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Wu, Y.; Zhang, Q.; Xie, H.; Liu, G.; Guo, Z.; Li, F.; Jia, J.; Kuang, S.; Hu, R. Enriched environment induces angiogenesis and improves neural function outcomes in rat stroke model. J. Neurol. Sci. 2014, 347, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.; Baker, K.B. Upside down Crossed Cerebellar Diaschisis: Proposing Chronic Stimulation of the Dentatothalamocortical Pathway for Post-Stroke Motor Recovery. Front. Integr. Neurosci. 2012, 6. [Google Scholar] [CrossRef] [PubMed]
- Cooperrider, J.; Furmaga, H.; Plow, E.; Park, H.J.; Chen, Z.; Kidd, G.; Baker, K.B.; Gale, J.T.; Machado, A.G. Chronic Deep Cerebellar Stimulation Promotes Long-Term Potentiation, Microstructural Plasticity, and Reorganization of Perilesional Cortical Representation in a Rodent Model. J. Neurosci. 2014, 34, 9040–9050. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.M.; Ishizaka, S.; Cheng, M.Y.; Wang, E.H.; Bautista, A.R.; Levy, S.; Smerin, D.; Sun, G.; Steinberg, G.K. Optogenetic neuronal stimulation of the lateral cerebellar nucleus promotes persistent functional recovery after stroke. Sci. Rep. 2017, 7, 46612. [Google Scholar] [CrossRef]
- Baker, K.B.; Schuster, D.; Cooperrider, J.; Machado, A.G. Deep Brain Stimulation of the Lateral Cerebellar Nucleus Produces Frequency-Specific Alterations in Motor Evoked Potentials in the Rat in Vivo. Exp. Neurol. 2010, 226, 259–264. [Google Scholar] [CrossRef]
- Van Overwalle, F.; D’Aes, T.; Marien, P. Social cognition and the cerebellum: A meta-analytic connectivity analysis. Hum. Brain Mapp. 2015, 36, 5137–5154. [Google Scholar] [CrossRef]
- Moulton, E.A.; Elman, I.; Becerra, L.R.; Goldstein, R.Z.; Borsook, D. The cerebellum and addiction: Insights gained from neuroimaging research. Addict. Biol. 2014, 19, 317–331. [Google Scholar] [CrossRef]
- Schmahmann, J.D.; Caplan, D. Cognition, emotion and the cerebellum. Brain 2006, 129, 290–292. [Google Scholar] [CrossRef]
- D’Angelo, E. The cerebellum gets social. Science 2019, 363, 229. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, E. Brainstem, cerebellar and limbic neuroanatomical abnormalities in autism. Curr. Opin. Neurobiol. 1997, 7, 269–278. [Google Scholar] [CrossRef]
- Soda, T.; Mapelli, L.; Locatelli, F.; Botta, L.; Goldfarb, M.; Prestori, F.; D’Angelo, E. Hyperexcitability and Hyperplasticity Disrupt Cerebellar Signal Transfer in the IB2 KO Mouse Model of Autism. J. Neurosci. 2019, 39, 2383–2397. [Google Scholar] [CrossRef] [PubMed]
- Badura, A.; Verpeut, J.L.; Metzger, J.W.; Pereira, T.D.; Pisano, T.J.; Deverett, B.; Bakshinskaya, D.E.; Wang, S.S. Normal cognitive and social development require posterior cerebellar activity. eLife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Kloth, A.D.; Badura, A. The cerebellum, sensitive periods, and autism. Neuron 2014, 83, 518–532. [Google Scholar] [CrossRef] [PubMed]
- Andreasen, N.C.; Pierson, R. The role of the cerebellum in schizophrenia. Biol. Psychiatry 2008, 64, 81–88. [Google Scholar] [CrossRef]
- Picard, H.; Amado, I.; Mouchet-Mages, S.; Olie, J.P.; Krebs, M.O. The role of the cerebellum in schizophrenia: An update of clinical, cognitive, and functional evidences. Schizophr. Bull. 2008, 34, 155–172. [Google Scholar] [CrossRef]
- Giza, J.; Urbanski, M.J.; Prestori, F.; Bandyopadhyay, B.; Yam, A.; Friedrich, V.; Kelley, K.; D’Angelo, E.; Goldfarb, M. Behavioral and cerebellar transmission deficits in mice lacking the autism-linked gene islet brain-2. J. Neurosci. 2010, 30, 14805–14816. [Google Scholar] [CrossRef]
- Whyatt, C.; Craig, C. Sensory-motor problems in Autism. Front. Integr. Neurosci. 2013, 7, 51. [Google Scholar] [CrossRef]
- Schmahmann, J.D.; Guell, X.; Stoodley, C.J.; Halko, M.A. The Theory and Neuroscience of Cerebellar Cognition. Annu. Rev. Neurosci. 2019, 42, 337–364. [Google Scholar] [CrossRef]
- Strick, P.L.; Dum, R.P.; Fiez, J.A. Cerebellum and nonmotor function. Annu. Rev. Neurosci. 2009, 32, 413–434. [Google Scholar] [CrossRef]
- Dum, R.P.; Strick, P.L. An unfolded map of the cerebellar dentate nucleus and its projections to the cerebral cortex. J. Neurophysiol. 2003, 89, 634–639. [Google Scholar] [CrossRef]
- Steele, C.J.; Anwander, A.; Bazin, P.L.; Trampel, R.; Schaefer, A.; Turner, R.; Ramnani, N.; Villringer, A. Human Cerebellar Sub-millimeter Diffusion Imaging Reveals the Motor and Non-motor Topography of the Dentate Nucleus. Cereb. Cortex 2017, 27, 4537–4548. [Google Scholar] [CrossRef] [PubMed]
- Bernard, J.A.; Peltier, S.J.; Benson, B.L.; Wiggins, J.L.; Jaeggi, S.M.; Buschkuehl, M.; Jonides, J.; Monk, C.S.; Seidler, R.D. Dissociable Functional Networks of the Human Dentate Nucleus. Cereb. Cortex 2014, 24, 2151–2159. [Google Scholar] [CrossRef] [PubMed]
- Magnotta, V.A.; Adix, M.L.; Caprahan, A.; Lim, K.; Gollub, R.; Andreasen, N.C. Investigating connectivity between the cerebellum and thalamus in schizophrenia using diffusion tensor tractography: A pilot study. Psychiatry Res. 2008, 163, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Gornati, S.V.; Schafer, C.B.; Eelkman Rooda, O.H.J.; Nigg, A.L.; De Zeeuw, C.I.; Hoebeek, F.E. Differentiating Cerebellar Impact on Thalamic Nuclei. Cell Rep. 2018, 23, 2690–2704. [Google Scholar] [CrossRef]
- Parnaudeau, S.; O’Neill, P.K.; Bolkan, S.S.; Ward, R.D.; Abbas, A.I.; Roth, B.L.; Balsam, P.D.; Gordon, J.A.; Kellendonk, C. Inhibition of mediodorsal thalamus disrupts thalamofrontal connectivity and cognition. Neuron 2013, 77, 1151–1162. [Google Scholar] [CrossRef]
- Ferguson, B.R.; Gao, W.J. Thalamic Control of Cognition and Social Behavior Via Regulation of Gamma-Aminobutyric Acidergic Signaling and Excitation/Inhibition Balance in the Medial Prefrontal Cortex. Biol. Psychiatry 2018, 83, 657–669. [Google Scholar] [CrossRef]
- Sieveritz, B.; Garcia-Munoz, M.; Arbuthnott, G.W. Thalamic afferents to prefrontal cortices from ventral motor nuclei in decision-making. Eur. J. Neurosci. 2019, 49, 646–657. [Google Scholar] [CrossRef]
- Collins, D.P.; Anastasiades, P.G.; Marlin, J.J.; Carter, A.G. Reciprocal Circuits Linking the Prefrontal Cortex with Dorsal and Ventral Thalamic Nuclei. Neuron 2018, 98, 366–379.e4. [Google Scholar] [CrossRef]
- Rogers, T.D.; Dickson, P.E.; McKimm, E.; Heck, D.H.; Goldowitz, D.; Blaha, C.D.; Mittleman, G. Reorganization of circuits underlying cerebellar modulation of prefrontal cortical dopamine in mouse models of autism spectrum disorder. Cerebellum 2013, 12, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Thierry, A.M.; Tassin, J.P.; Blanc, G.; Glowinski, J. Selective activation of mesocortical DA system by stress. Nature 1976, 263, 242–244. [Google Scholar] [CrossRef] [PubMed]
- Brozoski, T.J.; Brown, R.M.; Rosvold, H.E.; Goldman, P.S. Cognitive deficit caused by regional depletion of dopamine in prefrontal cortex of rhesus monkey. Science 1979, 205, 929–932. [Google Scholar] [CrossRef] [PubMed]
- Chudasama, Y.; Robbins, T.W. Dopaminergic modulation of visual attention and working memory in the rodent prefrontal cortex. Neuropsychopharmacology 2004, 29, 1628–1636. [Google Scholar] [CrossRef] [PubMed]
- Floresco, S.B. Prefrontal dopamine and behavioral flexibility: Shifting from an “inverted-U” toward a family of functions. Front. Neurosci. 2013, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Braver, T.S.; Krug, M.K.; Chiew, K.S.; Kool, W.; Westbrook, J.A.; Clement, N.J.; Adcock, R.A.; Barch, D.M.; Botvinick, M.M.; Carver, C.S.; et al. Mechanisms of motivation-cognition interaction: Challenges and opportunities. Cogn. Affect. Behav. Neurosci. 2014, 14, 443–472. [Google Scholar] [CrossRef]
- Westbrook, A.; Braver, T.S. Dopamine Does Double Duty in Motivating Cognitive Effort. Neuron 2016, 91, 708. [Google Scholar] [CrossRef]
- Howes, O.D.; Kapur, S. The dopamine hypothesis of schizophrenia: Version III—The final common pathway. Schizophr. Bull. 2009, 35, 549–562. [Google Scholar] [CrossRef]
- Ernst, M.; Zametkin, A.J.; Matochik, J.A.; Pascualvaca, D.; Cohen, R.M. Low medial prefrontal dopaminergic activity in autistic children. Lancet 1997, 350, 638. [Google Scholar] [CrossRef]
- Nakamura, K.; Sekine, Y.; Ouchi, Y.; Tsujii, M.; Yoshikawa, E.; Futatsubashi, M.; Tsuchiya, K.J.; Sugihara, G.; Iwata, Y.; Suzuki, K.; et al. Brain serotonin and dopamine transporter bindings in adults with high-functioning autism. Arch. Gen. Psychiatry 2010, 67, 59–68. [Google Scholar] [CrossRef]
- Mittleman, G.; Goldowitz, D.; Heck, D.; Blaha, C. Cerebellar modulation of frontal cortex dopamine efflux in mice: Relevance to autism and schizophrenia. Synapse 2008, 62, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Rogers, T.D.; Dickson, P.E.; Heck, D.H.; Goldowitz, D.; Mittleman, G.; Blaha, C.D. Connecting the dots of the cerebro-cerebellar role in cognitive function: Neuronal pathways for cerebellar modulation of dopamine release in the prefrontal cortex. Synapse 2011, 65, 1204–1212. [Google Scholar] [CrossRef] [PubMed]
- Carta, I.; Chen, C.H.; Schott, A.L.; Dorizan, S.; Khodakhah, K. Cerebellar modulation of the reward circuitry and social behavior. Science 2019, 363. [Google Scholar] [CrossRef] [PubMed]
- Krook-Magnuson, E.; Szabo, G.G.; Armstrong, C.; Oijala, M.; Soltesz, I. Cerebellar Directed Optogenetic Intervention Inhibits Spontaneous Hippocampal Seizures in a Mouse Model of Temporal Lobe Epilepsy. eNeuro 2014, 1. [Google Scholar] [CrossRef] [PubMed]
- Streng, M.L.; Krook-Magnuson, E. Excitation, but not inhibition, of the fastigial nucleus provides powerful control over temporal lobe seizures. J. Physiol. 2020, 598, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Kros, L.; Eelkman Rooda, O.H.; Spanke, J.K.; Alva, P.; van Dongen, M.N.; Karapatis, A.; Tolner, E.A.; Strydis, C.; Davey, N.; Winkelman, B.H.; et al. Cerebellar output controls generalized spike-and-wave discharge occurrence. Ann. Neurol. 2015, 77, 1027–1049. [Google Scholar] [CrossRef]
- Deverett, B.; Kislin, M.; Tank, D.W.; Wang, S.S. Cerebellar disruption impairs working memory during evidence accumulation. Nat. Commun. 2019, 10, 3128. [Google Scholar] [CrossRef]
- Tsubota, T.; Ohashi, Y.; Tamura, K.; Miyashita, Y. Optogenetic inhibition of Purkinje cell activity reveals cerebellar control of blood pressure during postural alterations in anesthetized rats. Neuroscience 2012, 210, 137–144. [Google Scholar] [CrossRef]
- Parker, K.L.; Kim, Y.C.; Kelley, R.M.; Nessler, A.J.; Chen, K.H.; Muller-Ewald, V.A.; Andreasen, N.C.; Narayanan, N.S. Delta-frequency stimulation of cerebellar projections can compensate for schizophrenia-related medial frontal dysfunction. Mol. Psychiatry 2017, 22, 647–655. [Google Scholar] [CrossRef]
- Bicks, L.K.; Koike, H.; Akbarian, S.; Morishita, H. Prefrontal Cortex and Social Cognition in Mouse and Man. Front. Psychol. 2015, 6, 1805. [Google Scholar] [CrossRef]
- Murray, J.D.; Jaramillo, J.; Wang, X.J. Working Memory and Decision-Making in a Frontoparietal Circuit Model. J. Neurosci. 2017, 37, 12167–12186. [Google Scholar] [CrossRef]
- Ferrari, C.; Cattaneo, Z.; Oldrati, V.; Casiraghi, L.; Castelli, F.; D’Angelo, E.; Vecchi, T. TMS Over the Cerebellum Interferes with Short-term Memory of Visual Sequences. Sci. Rep. 2018, 8, 6722. [Google Scholar] [CrossRef]
- Ravizza, S.M.; McCormick, C.A.; Schlerf, J.E.; Justus, T.; Ivry, R.B.; Fiez, J.A. Cerebellar damage produces selective deficits in verbal working memory. Brain 2006, 129, 306–320. [Google Scholar] [CrossRef]
- Kansal, K.; Yang, Z.; Fishman, A.M.; Sair, H.I.; Ying, S.H.; Jedynak, B.M.; Prince, J.L.; Onyike, C.U. Structural cerebellar correlates of cognitive and motor dysfunctions in cerebellar degeneration. Brain 2017, 140, 707–720. [Google Scholar] [CrossRef]
- Reeber, S.L.; Otis, T.S.; Sillitoe, R.V. New roles for the cerebellum in health and disease. Front. Syst. Neurosci. 2013, 7, 83. [Google Scholar] [CrossRef]
- Blackwood, N.; Ffytche, D.; Simmons, A.; Bentall, R.; Murray, R.; Howard, R. The cerebellum and decision making under uncertainty. Brain Res. Cogn. Brain Res. 2004, 20, 46–53. [Google Scholar] [CrossRef]
- Brody, C.D.; Hanks, T.D. Neural underpinnings of the evidence accumulator. Curr. Opin. Neurobiol. 2016, 37, 149–157. [Google Scholar] [CrossRef]
- Gold, J.I.; Shadlen, M.N. The neural basis of decision making. Annu. Rev. Neurosci. 2007, 30, 535–574. [Google Scholar] [CrossRef]
- Demirtas-Tatlidede, A.; Freitas, C.; Cromer, J.R.; Safar, L.; Ongur, D.; Stone, W.S.; Seidman, L.J.; Schmahmann, J.D.; Pascual-Leone, A. Safety and proof of principle study of cerebellar vermal theta burst stimulation in refractory schizophrenia. Schizophr. Res. 2010, 124, 91–100. [Google Scholar] [CrossRef]
- Schutter, D.J.; van Honk, J.; d’Alfonso, A.A.; Peper, J.S.; Panksepp, J. High frequency repetitive transcranial magnetic over the medial cerebellum induces a shift in the prefrontal electroencephalography gamma spectrum: A pilot study in humans. Neurosci. Lett. 2003, 336, 73–76. [Google Scholar] [CrossRef]
- Garg, S.; Sinha, V.K.; Tikka, S.K.; Mishra, P.; Goyal, N. The efficacy of cerebellar vermal deep high frequency (theta range) repetitive transcranial magnetic stimulation (rTMS) in schizophrenia: A randomized rater blind-sham controlled study. Psychiatry Res. 2016, 243, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.L.; Narayanan, N.S.; Andreasen, N.C. The therapeutic potential of the cerebellum in schizophrenia. Front. Syst. Neurosci. 2014, 8, 163. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.L. Timing Tasks Synchronize Cerebellar and Frontal Ramping Activity and Theta Oscillations: Implications for Cerebellar Stimulation in Diseases of Impaired Cognition. Front. Psychiatry 2015, 6, 190. [Google Scholar] [CrossRef] [PubMed]
- Okubo, Y.; Suhara, T.; Suzuki, K.; Kobayashi, K.; Inoue, O.; Terasaki, O.; Someya, Y.; Sassa, T.; Sudo, Y.; Matsushima, E.; et al. Decreased prefrontal dopamine D1 receptors in schizophrenia revealed by PET. Nature 1997, 385, 634–636. [Google Scholar] [CrossRef] [PubMed]
- Goldman-Rakic, P.S.; Castner, S.A.; Svensson, T.H.; Siever, L.J.; Williams, G.V. Targeting the dopamine D1 receptor in schizophrenia: Insights for cognitive dysfunction. Psychopharmacology 2004, 174, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Abi-Dargham, A.; Mawlawi, O.; Lombardo, I.; Gil, R.; Martinez, D.; Huang, Y.; Hwang, D.R.; Keilp, J.; Kochan, L.; Van Heertum, R.; et al. Prefrontal dopamine D1 receptors and working memory in schizophrenia. J. Neurosci. 2002, 22, 3708–3719. [Google Scholar] [CrossRef]
- D’Angelo, E.; De Zeeuw, C.I. Timing and plasticity in the cerebellum: Focus on the granular layer. Trends Neurosci. 2009, 32, 30–40. [Google Scholar] [CrossRef]
- Ivry, R.B.; Spencer, R.M.; Zelaznik, H.N.; Diedrichsen, J. The cerebellum and event timing. Ann. N. Y. Acad. Sci. 2002, 978, 302–317. [Google Scholar] [CrossRef]
- Ivry, R.B.; Spencer, R.M. The neural representation of time. Curr. Opin. Neurobiol. 2004, 14, 225–232. [Google Scholar] [CrossRef]
- Mesiwala, A.H.; Kuratani, J.D.; Avellino, A.M.; Roberts, T.S.; Sotero, M.A.; Ellenbogen, R.G. Focal motor seizures with secondary generalization arising in the cerebellum. Case report and review of the literature. J. Neurosurg. 2002, 97, 190–196. [Google Scholar] [CrossRef]
- Harvey, A.S.; Jayakar, P.; Duchowny, M.; Resnick, T.; Prats, A.; Altman, N.; Renfroe, J.B. Hemifacial seizures and cerebellar ganglioglioma: An epilepsy syndrome of infancy with seizures of cerebellar origin. Ann. Neurol. 1996, 40, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Kandel, A.; Buzsaki, G. Cerebellar neuronal activity correlates with spike and wave EEG patterns in the rat. Epilepsy Res. 1993, 16, 1–9. [Google Scholar] [CrossRef]
- Kottamasu, S.R. Brain imaging during seizure: Ictal brain SPECT. Indian J. Pediatr. 1997, 64, 575–580. [Google Scholar] [CrossRef]
- Blumenfeld, H.; Varghese, G.I.; Purcaro, M.J.; Motelow, J.E.; Enev, M.; McNally, K.A.; Levin, A.R.; Hirsch, L.J.; Tikofsky, R.; Zubal, I.G.; et al. Cortical and subcortical networks in human secondarily generalized tonic-clonic seizures. Brain 2009, 132, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Gartside, I.B. The activity of cerebellar neurones during epileptiform activity induced by penicillin in the cerebral cortex of the rat. Electroencephalogr. Clin. Neurophysiol. 1979, 46, 189–196. [Google Scholar] [CrossRef]
- Rochefort, C.; Arabo, A.; Andre, M.; Poucet, B.; Save, E.; Rondi-Reig, L. Cerebellum shapes hippocampal spatial code. Science 2011, 334, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Moscato, L.; Montagna, I.; De Propris, L.; Tritto, S.; Mapelli, L.; D’Angelo, E. Long-Lasting Response Changes in Deep Cerebellar Nuclei. Front. Cell. Neurosci. 2019, 13, 84. [Google Scholar] [CrossRef]
- Chaumont, J.; Guyon, N.; Valera, A.M.; Dugue, G.P.; Popa, D.; Marcaggi, P.; Gautheron, V.; Reibel-Foisset, S.; Dieudonne, S.; Stephan, A.; et al. Clusters of cerebellar Purkinje cells control their afferent climbing fiber discharge. Proc. Natl. Acad. Sci. USA 2013, 110, 16223–16228. [Google Scholar] [CrossRef]
- Brown, S.T.; Raman, I.M. Sensorimotor Integration and Amplification of Reflexive Whisking by Well-Timed Spiking in the Cerebellar Corticonuclear Circuit. Neuron 2018, 99, 564–575.e2. [Google Scholar] [CrossRef]
- Masoli, S.; D’Angelo, E. Synaptic Activation of a Detailed Purkinje Cell Model Predicts Voltage-Dependent Control of Burst-Pause Responses in Active Dendrites. Front. Cell. Neurosci. 2017, 11, 278. [Google Scholar] [CrossRef]
- La Noce, A.; Bradley, D.J.; Goring, M.A.; Spyer, K.M. The influence of lobule IX of the cerebellar posterior vermis on the baroreceptor reflex in the decerebrate rabbit. J. Auton. Nerv. Syst. 1991, 32, 31–36. [Google Scholar] [CrossRef]
- Holmes, M.J.; Cotter, L.A.; Arendt, H.E.; Cass, S.P.; Yates, B.J. Effects of lesions of the caudal cerebellar vermis on cardiovascular regulation in awake cats. Brain Res. 2002, 938, 62–72. [Google Scholar] [CrossRef]
- Miterko, L.N.; Baker, K.B.; Beckinghausen, J.; Bradnam, L.V.; Cheng, M.Y.; Cooperrider, J.; DeLong, M.R.; Gornati, S.V.; Hallett, M.; Heck, D.H.; et al. Consensus Paper: Experimental Neurostimulation of the Cerebellum. Cerebellum 2019, 18, 1064–1097. [Google Scholar] [CrossRef]
- Baker, C.K.; Flannery, J.G. Innovative Optogenetic Strategies for Vision Restoration. Front. Cell. Neurosci. 2018, 12, 316. [Google Scholar] [CrossRef] [PubMed]
- Delbeke, J.; Hoffman, L.; Mols, K.; Braeken, D.; Prodanov, D. And Then There Was Light: Perspectives of Optogenetics for Deep Brain Stimulation and Neuromodulation. Front. Neurosci. 2017, 11, 663. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serotype | Promoter | Expression (Cell type) | Opsin | Cerebellar Region | Behavior | Behavioral Outcomes |
|---|---|---|---|---|---|---|
| AAV1 | hSyn | Neuron-specific | ChR2 | DCN | Reward | Increased place preference [125] |
| AAV5 | CAG | All cells | ArchT | DCN | Social behavior | Altered social preference [125] |
| Pcp2-Cre * | Purkinje cells | ChR2 | Cortex (simplex) | Epilepsy | Reduction in hippocampal seizure duration and seizure-induced inhibition [126] | |
| AAV9 | VGluT-Cre * | Glutamatergic | ChR2 | DCN (fastigial) | Epilepsy | Reduction in hippocampal seizure duration [127] |
| AAV2 | hSyn | Neuron-specific | ChR2 | DCN (dentate) | Epilepsy | Reduction in thalamocortical oscillations [128] |
| AAV2/9 | Pcp2-Cre * | Purkinje cells | ChR2 | Cortex (Crus I) | Working memory | Reduction in performance accuracy [129] |
| Lentivirus | L7 | Purkinje cells | eNpHR3.0 | Cortex (uvula) | Postural alterations | Reduction in the extent of blood pressure recovery [130] |
| AAV | CamKII | Glutamatergic | ChR2 | DCN (dentate) | Schizophrenia | Increase in prefrontal activity [131] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prestori, F.; Montagna, I.; D’Angelo, E.; Mapelli, L. The Optogenetic Revolution in Cerebellar Investigations. Int. J. Mol. Sci. 2020, 21, 2494. https://doi.org/10.3390/ijms21072494
Prestori F, Montagna I, D’Angelo E, Mapelli L. The Optogenetic Revolution in Cerebellar Investigations. International Journal of Molecular Sciences. 2020; 21(7):2494. https://doi.org/10.3390/ijms21072494
Chicago/Turabian StylePrestori, Francesca, Ileana Montagna, Egidio D’Angelo, and Lisa Mapelli. 2020. "The Optogenetic Revolution in Cerebellar Investigations" International Journal of Molecular Sciences 21, no. 7: 2494. https://doi.org/10.3390/ijms21072494
APA StylePrestori, F., Montagna, I., D’Angelo, E., & Mapelli, L. (2020). The Optogenetic Revolution in Cerebellar Investigations. International Journal of Molecular Sciences, 21(7), 2494. https://doi.org/10.3390/ijms21072494