Mimicking Sampson’s Retrograde Menstrual Theory in Rats: A New Rat Model for Ongoing Endometriosis-Associated Pain

 ,
,
Abstract
1. Introduction
2. Results
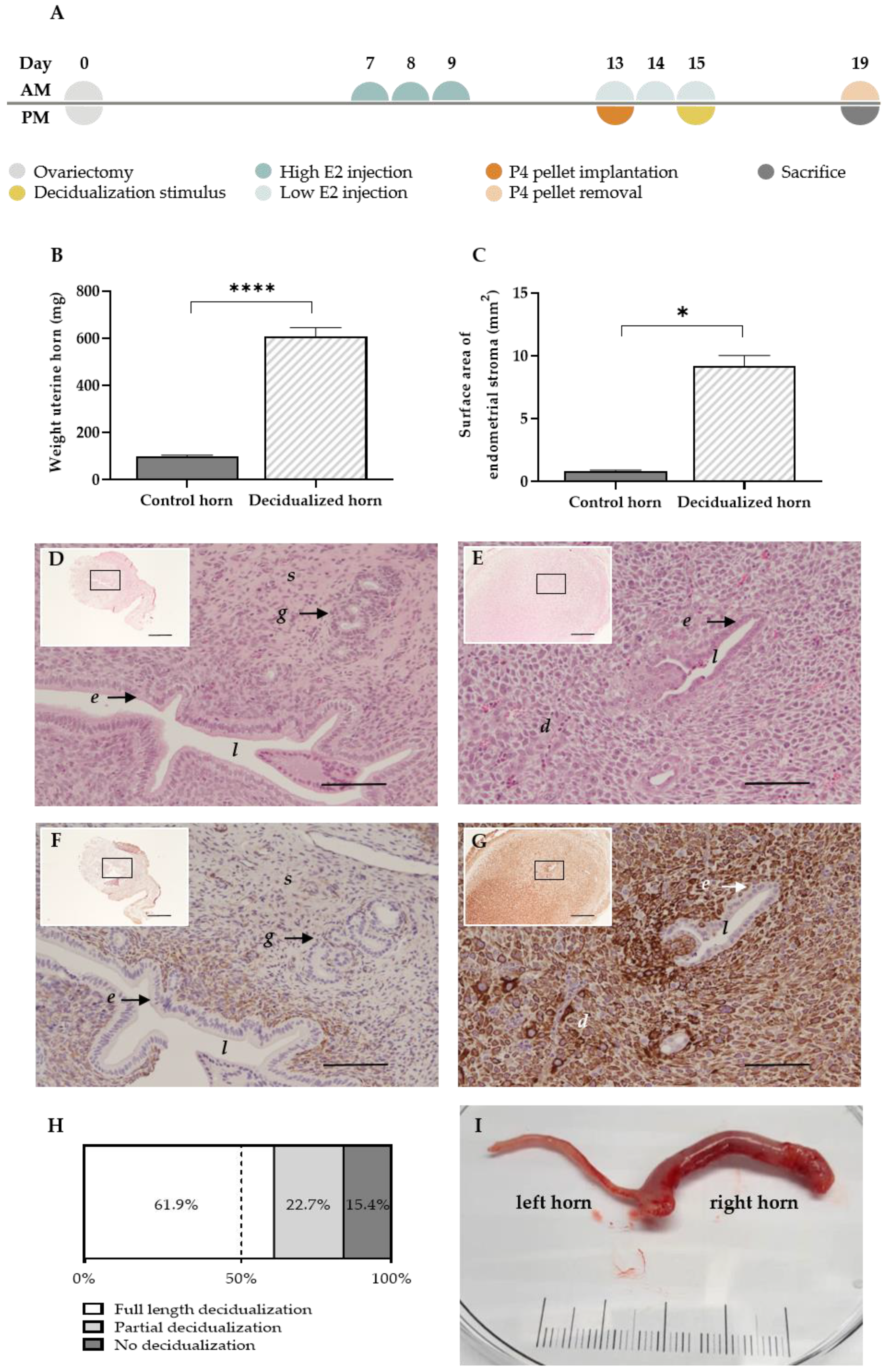
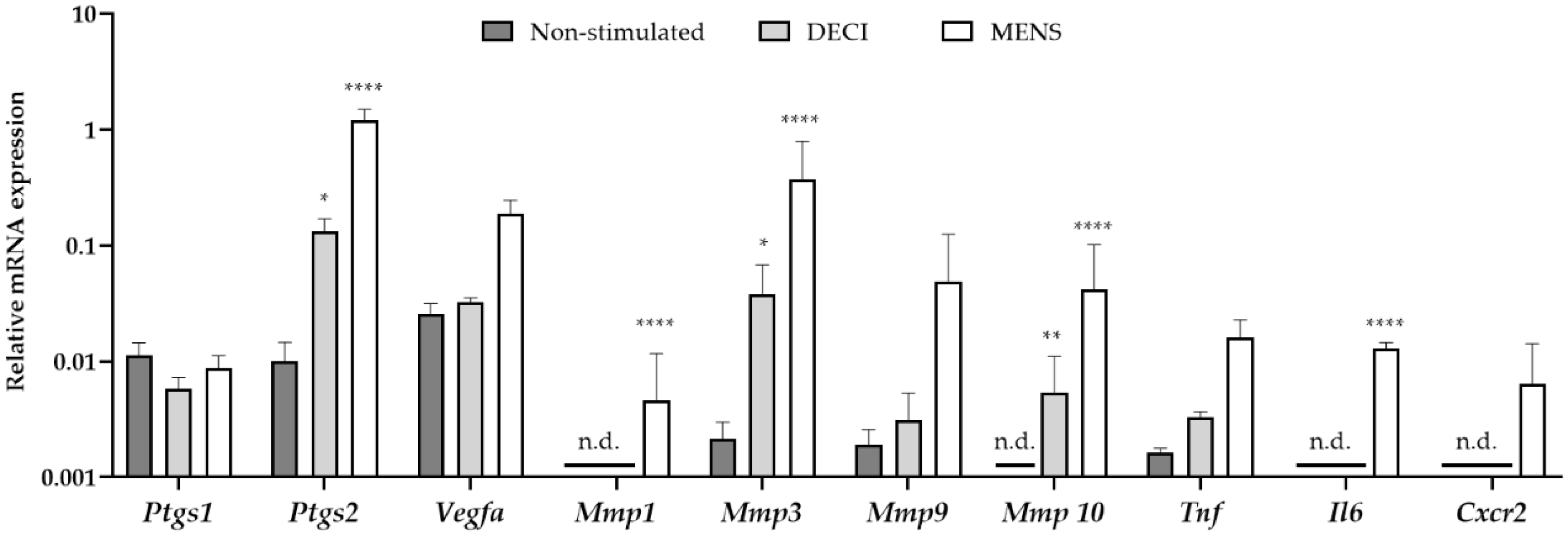
2.1. Generation of Menstrual Endometrial Tissue in Donor Rats
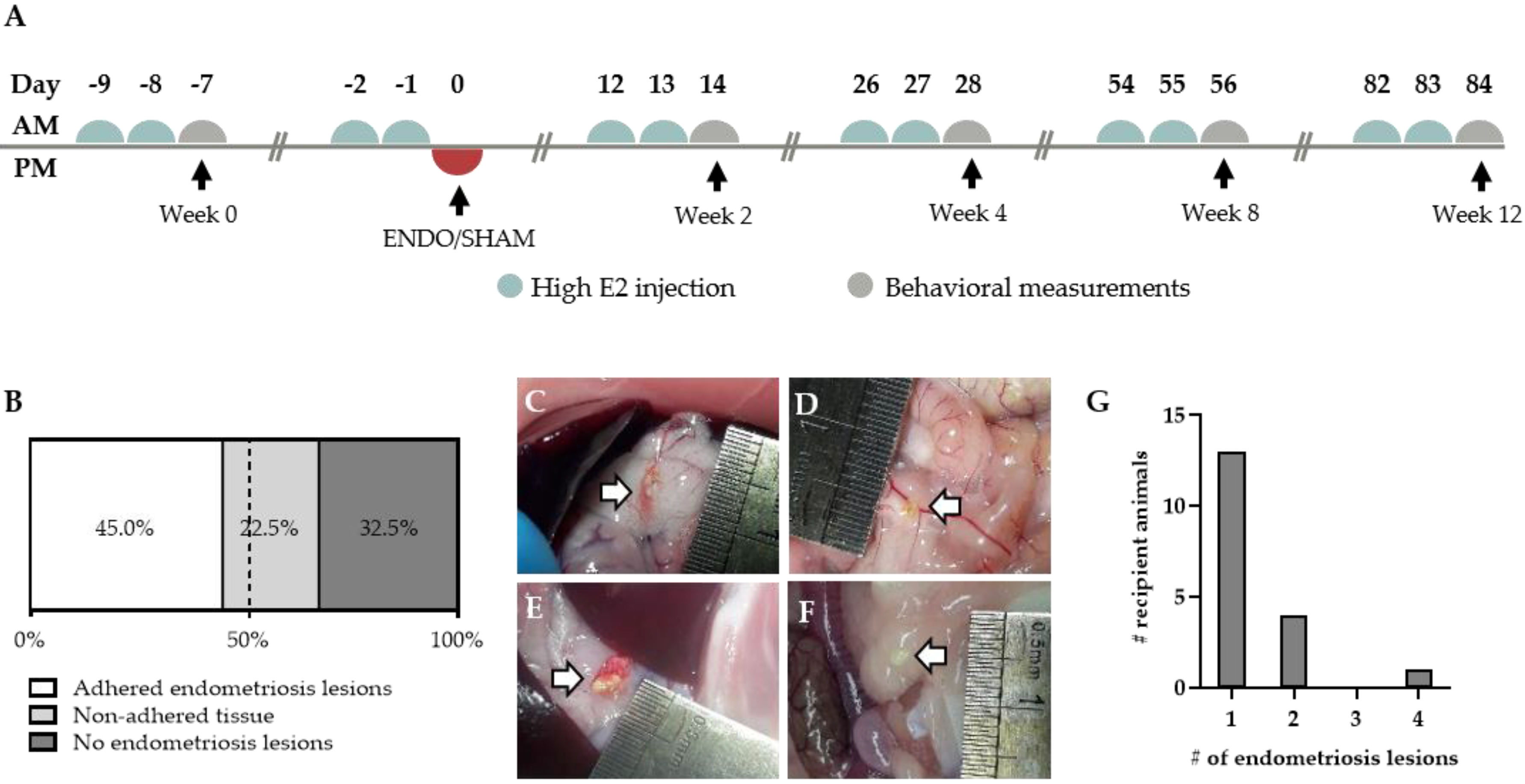
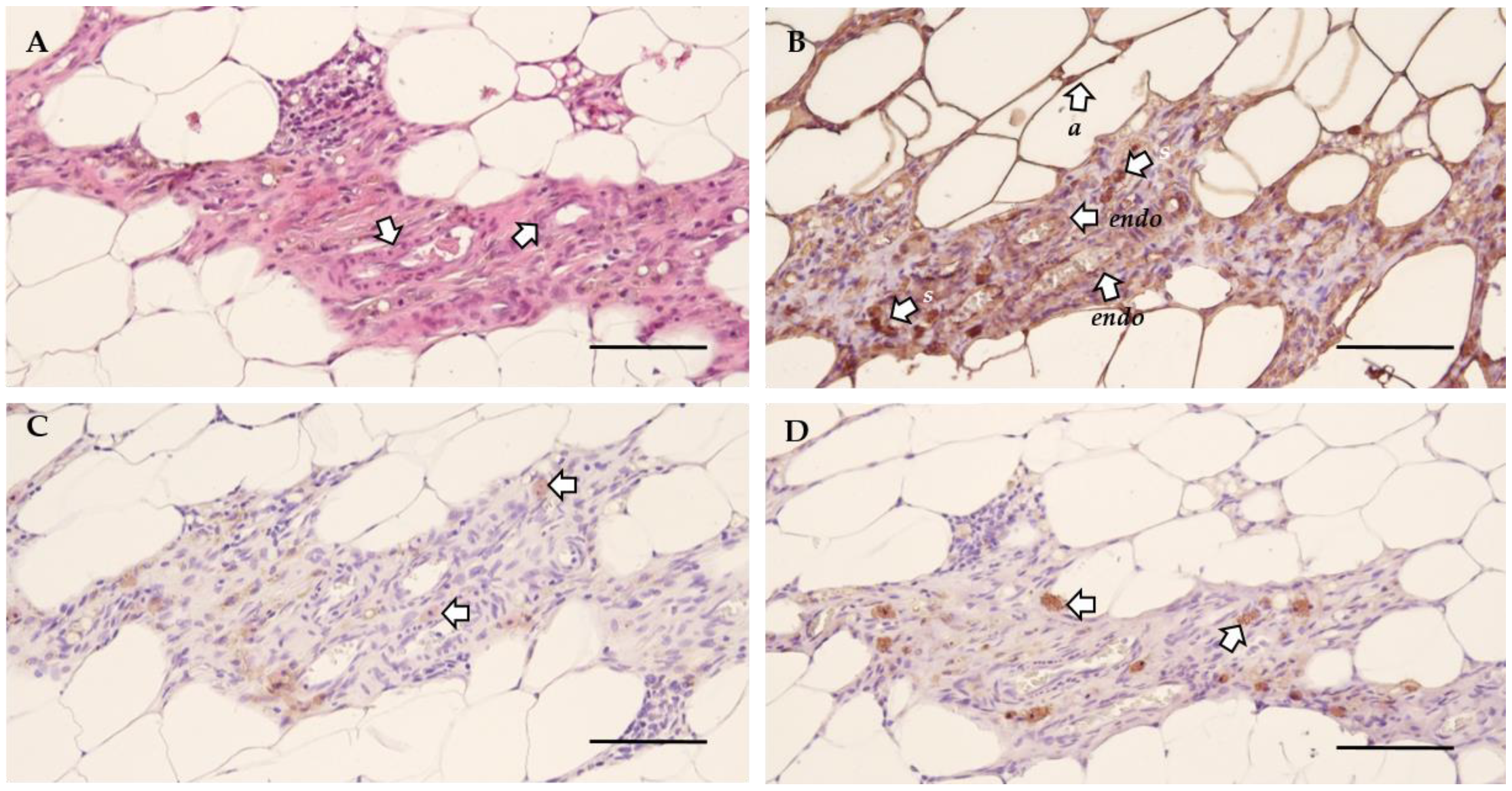
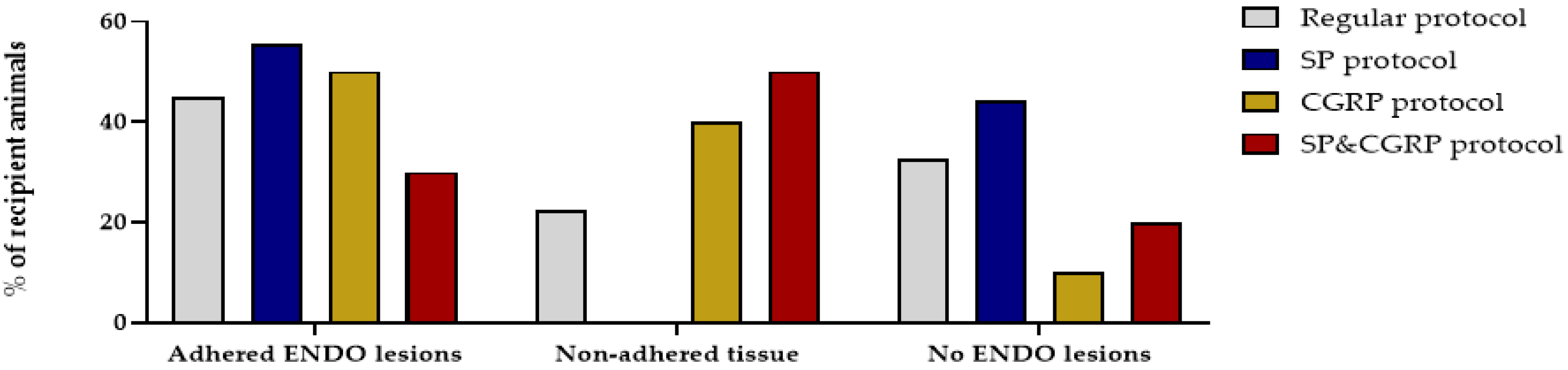
2.2. Endometriosis Lesions can be Retrieved from Recipient Rats
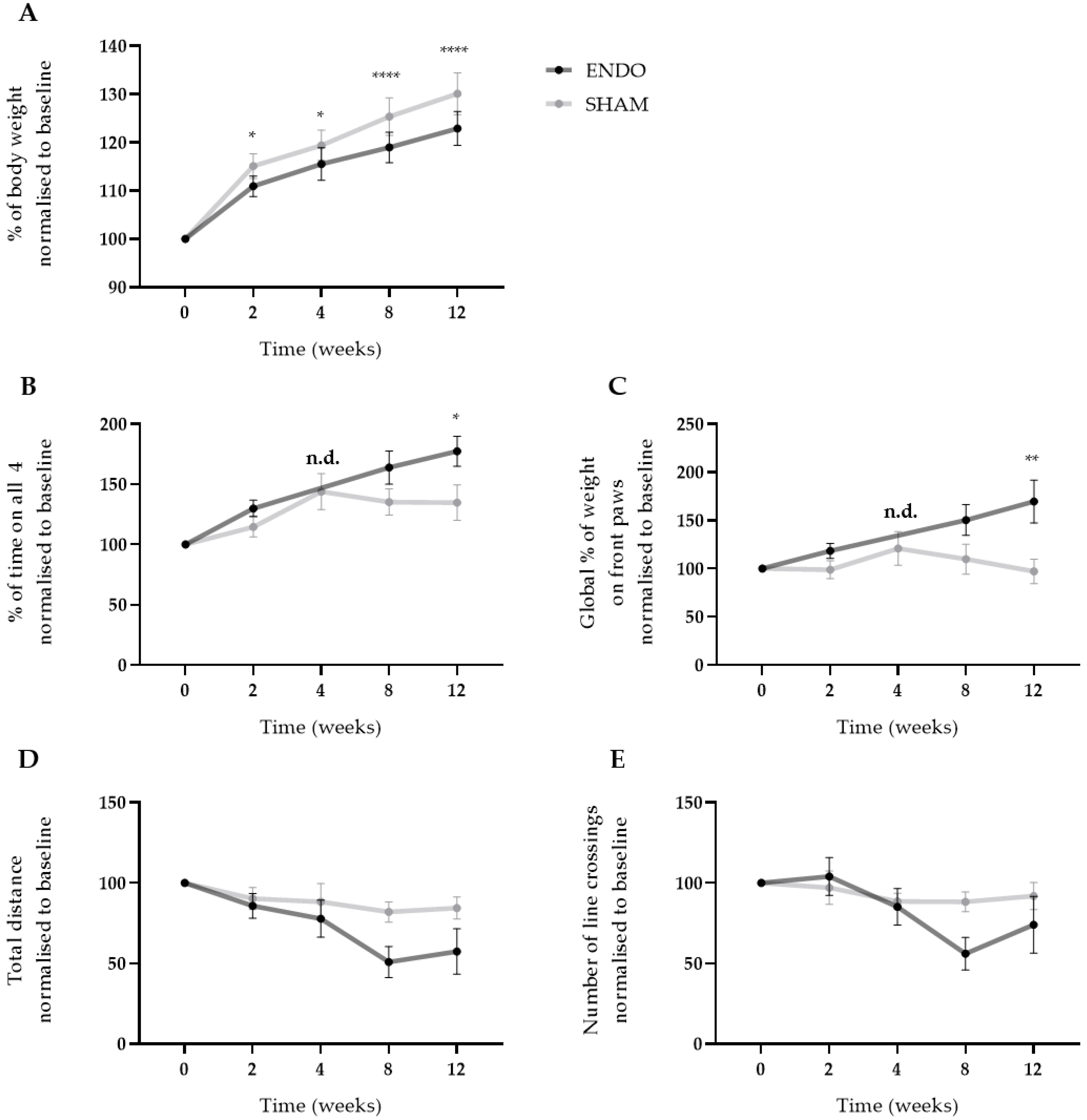
2.3. Ongoing Endometriosis-Associated Pain in Recipient Rats
2.4. Supplementation of Neuropeptides has no Effect on Endometriosis-Associated Pain
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Menstruating Rat Model
4.3. Vaginal Smear Examination
4.4. RT–qPCR Experiments
4.5. Endometriosis Induction
4.6. Immunohistochemistry
4.6.1. Hematoxylin & Eosin
4.6.2. Cytokeratin, Vimentin and GAP43
4.7. Behavioral Tests
4.7.1. Advanced Dynamic Weight Bearing Assessment
4.7.2. Open Field Assay
4.8. SP and CGRP Supplementation
4.8.1. Neuropeptide Pellets and Plasma Collection
4.8.2. SP & CGRP Plasma Concentration
4.9. Data Analysis and Display
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Asante, A.; Taylor, R.N. Endometriosis: The role of neuroangiogenesis. Annu. Rev. Physiol. 2011, 73, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Sampson, J.A. Peritoneal endometriosis due to the menstrual dissemination of endometrial tissue into the peritoneal cavity. Am. J. Obstet. Gynecol. 1927, 14, 422–469. [Google Scholar] [CrossRef]
- Sampson, J.A. Metastatic or Embolic Endometriosis, due to the Menstrual Dissemination of Endometrial Tissue into the Venous Circulation. Am. J. Pathol. 1927, 3, 93–110. [Google Scholar] [PubMed]
- Koninckx, P.R. Is mild endometriosis a condition occurring intermittently in all women? Hum. Reprod. 1994, 9, 2202–2205. [Google Scholar] [CrossRef] [PubMed]
- Nap, A.W.; Groothuis, P.G.; Demir, A.Y.; Evers, J.L.; Dunselman, G.A. Pathogenesis of endometriosis. Best Pract. Res. Clin. Obstet. Gynaecol. 2004, 18, 233–244. [Google Scholar] [CrossRef]
- Bajaj, P.; Bajaj, P.; Madsen, H.; Arendt-Nielsen, L. Endometriosis is associated with central sensitization: A psychophysical controlled study. J. Pain 2003, 4, 372–380. [Google Scholar] [CrossRef]
- Wozniak, S. Chronic pelvic pain. Ann. Agric. Environ. Med. 2016, 23, 223–226. [Google Scholar] [CrossRef]
- Rogers, P.A.; D’Hooghe, T.M.; Fazleabas, A.; Giudice, L.C.; Montgomery, G.W.; Petraglia, F.; Taylor, R.N. Defining future directions for endometriosis research: Workshop report from the 2011 World Congress of Endometriosis In Montpellier, France. Reprod. Sci. 2013, 20, 483–499. [Google Scholar] [CrossRef]
- Greaves, E.; Cousins, F.L.; Murray, A.; Esnal-Zufiaurre, A.; Fassbender, A.; Horne, A.W.; Saunders, P.T. A novel mouse model of endometriosis mimics human phenotype and reveals insights into the inflammatory contribution of shed endometrium. Am. J. Pathol. 2014, 184, 1930–1939. [Google Scholar] [CrossRef]
- Greaves, E.; Horne, A.W.; Jerina, H.; Mikolajczak, M.; Hilferty, L.; Mitchell, R.; Fleetwood-Walker, S.N.; Saunders, P.T.K. EP2 receptor antagonism reduces peripheral and central hyperalgesia in a preclinical mouse model of endometriosis. Sci. Rep. 2017, 7, 44169. [Google Scholar] [CrossRef]
- Cousins, F.L.; Murray, A.; Esnal, A.; Gibson, D.A.; Critchley, H.O.; Saunders, P.T. Evidence from a mouse model that epithelial cell migration and mesenchymal-epithelial transition contribute to rapid restoration of uterine tissue integrity during menstruation. PLoS ONE 2014, 9, e86378. [Google Scholar] [CrossRef] [PubMed]
- Menning, A.; Walter, A.; Rudolph, M.; Gashaw, I.; Fritzemeier, K.H.; Roese, L. Granulocytes and vascularization regulate uterine bleeding and tissue remodeling in a mouse menstruation model. PLoS ONE 2012, 7, e41800. [Google Scholar] [CrossRef]
- Maeda, T.; Byers, M.R. Different localizations of growth-associated protein (GAP-43) in mechanoreceptors and free nerve endings of adult rat periodontal ligament, dental pulp and skin. Arch. Histol. Cytol. 1996, 59, 291–304. [Google Scholar]
- Morgan, M.A.; Pfaff, D.W. Effects of estrogen on activity and fear-related behaviors in mice. Horm. Behav. 2001, 40, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Liu, X.; Guo, S.W. The establishment of a mouse model of deep endometriosis. Hum. Reprod. 2019, 34, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, M.L.; Hengartner, M.P.; Kohl-Schwartz, A.; Geraedts, K.; Rauchfuss, M.; Woelfler, M.M.; Haeberlin, F.; Orelli, S.; Eberhard, M.; Maurer, F.; et al. Does endometriosis affect professional life? A matched case-control study in Switzerland, Germany and Austria. BMJ Open 2019, 9, e019570. [Google Scholar] [CrossRef] [PubMed]
- Ramathal, C.Y.; Bagchi, I.C.; Taylor, R.N.; Bagchi, M.K. Endometrial decidualization: Of mice and men. Semin. Reprod. Med. 2010, 28, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Peterse, D.; Clercq, K.; Goossens, C.; Binda, M.M.; Dorien, F.O.; Saunders, P.; Vriens, J.; Fassbender, A.; D’Hooghe, T.M. Optimization of Endometrial Decidualization in the Menstruating Mouse Model for Preclinical Endometriosis Research. Reprod. Sci. 2018, 25, 1577–1588. [Google Scholar] [CrossRef]
- Finn, C.A.; Pope, M. Vascular and cellular changes in the decidualized endometrium of the ovariectomized mouse following cessation of hormone treatment: A possible model for menstruation. J. Endocrinol. 1984, 100, 295–300. [Google Scholar] [CrossRef]
- Glasser, S.R.; Lampelo, S.; Munir, M.I.; Julian, J. Expression of Desmin, Laminin and Fibronectin during Insitu Differentiation (Decidualization) of Rat Uterine Stromal Cells. Differentiation 1987, 35, 132–142. [Google Scholar] [CrossRef]
- Brasted, M.; White, C.A.; Kennedy, T.G.; Salamonsen, L.A. Mimicking the events of menstruation in the murine uterus. Biol. Reprod. 2003, 69, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, H.N.; Kelly, R.W.; Fraser, H.M.; Critchley, H.O. Endocrine regulation of menstruation. Endocr. Rev. 2006, 27, 17–46. [Google Scholar] [CrossRef]
- Smith, W.L.; DeWitt, D.L.; Garavito, R.M. Cyclooxygenases: Structural, cellular, and molecular biology. Annu. Rev. Biochem. 2000, 69, 145–182. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Park, H.J.; Seol, J.W.; Jang, J.Y.; Cho, Y.S.; Kim, K.R.; Choi, Y.; Lydon, J.P.; DeMayo, F.J.; Shibuya, M.; et al. VEGF-A regulated by progesterone governs uterine angiogenesis and vascular remodelling during pregnancy. EMBO Mol. Med. 2013, 5, 1415–1430. [Google Scholar] [CrossRef] [PubMed]
- Salamonsen, L.A.; Woolley, D.E. Matrix metalloproteinases in normal menstruation. Hum. Reprod. 1996, 11, 124–133. [Google Scholar] [CrossRef] [PubMed]
- von Wolff, M.; Thaler, C.J.; Strowitzki, T.; Broome, J.; Stolz, W.; Tabibzadeh, S. Regulated expression of cytokines in human endometrium throughout the menstrual cycle: Dysregulation in habitual abortion. Mol. Hum. Reprod. 2000, 6, 627–634. [Google Scholar] [CrossRef]
- Vigano, P.; Candiani, M.; Monno, A.; Giacomini, E.; Vercellini, P.; Somigliana, E. Time to redefine endometriosis including its pro-fibrotic nature. Hum. Reprod. 2018, 33, 347–352. [Google Scholar] [CrossRef]
- Dodds, K.N.; Beckett, E.A.H.; Evans, S.F.; Hutchinson, M.R. Lesion development is modulated by the natural estrous cycle and mouse strain in a minimally invasive model of endometriosis. Biol. Reprod. 2017, 97, 810–821. [Google Scholar] [CrossRef]
- Noble, L.S.; Simpson, E.R.; Johns, A.; Bulun, S.E. Aromatase expression in endometriosis. J. Clin. Endocrinol. Metab. 1996, 81, 174–179. [Google Scholar]
- Cullinan-Bove, K.; Koos, R.D. Vascular endothelial growth factor/vascular permeability factor expression in the rat uterus: Rapid stimulation by estrogen correlates with estrogen-induced increases in uterine capillary permeability and growth. Endocrinology 1993, 133, 829–837. [Google Scholar] [CrossRef]
- Sondell, M.; Lundborg, G.; Kanje, M. Vascular endothelial growth factor has neurotrophic activity and stimulates axonal outgrowth, enhancing cell survival and Schwann cell proliferation in the peripheral nervous system. J. Neurosci. 1999, 19, 5731–5740. [Google Scholar] [CrossRef] [PubMed]
- Dall, G.; Vieusseux, J.; Unsworth, A.; Anderson, R.; Britt, K. Low Dose, Low Cost Estradiol Pellets Can Support MCF-7 Tumour Growth in Nude Mice without Bladder Symptoms. J. Cancer 2015, 6, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Burns, K.A.; Rodriguez, K.F.; Hewitt, S.C.; Janardhan, K.S.; Young, S.L.; Korach, K.S. Role of estrogen receptor signaling required for endometriosis-like lesion establishment in a mouse model. Endocrinology 2012, 153, 3960–3971. [Google Scholar] [CrossRef] [PubMed]
- Mogil, J.S. The Measurement of Pain in the Laboratory Rodent; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Laux-Biehlmann, A.; Boyken, J.; Dahllöf, H.; Schmidt, N.; Zollner, T.M.; Nagel, J. Dynamic weight bearing as a non-reflexive method for the measurement of abdominal pain in mice. Eur. J. Pain 2016, 20, 742–752. [Google Scholar] [CrossRef]
- De Clercq, K.; Van den Eynde, C.; Hennes, A.; Van Bree, R.; Voets, T.; Vriens, J. The functional expression of transient receptor potential channels in the mouse endometrium. Hum. Reprod. 2017, 32, 615–630. [Google Scholar] [CrossRef] [PubMed]
- Caligioni, C.S. Assessing reproductive status/stages in mice. Curr. Protoc. Neurosci. 2009, 48, A-41. [Google Scholar] [CrossRef]
- Cook, M. The Anatomy of the Laboratory Mouse; Academic Press: London, UK, 1965. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vimentin | Cytokeratin | GAP43 | |
|---|---|---|---|
| 1st Antibody | Anti-vimentin | Anti-rat cytokeratin 8 | Anti-GAP43 |
| Company-ref | Abcam-ab92547 | Dako-Z0622 | Abcam-ab75810 |
| Marker | Endothelial and stromal cells | Epithelial cells | Nerve fibers |
| Type | Rabbit monoclonal | Rabbit polyclonal | Rabbit monoclonal |
| Blocking time | 15′ | 15′ | 15′ |
| Blocking | Regular | Regular + 1/30 NGS | Regular |
| Antigen retrieval | 1 h at 90 °C in Tris EDTA | 10′ at 37 °C in 0.04% pepsin in 0.01M HCl | 1 h at 90 °C in citrate; pH = 6 |
| Dilution 1st Ab | 1/1500 | 1/1500 | 1/250 |
| Incubation 1st Ab | 1 h; 37 °C | 2 h; RT | ON; 4 °C |
| 2nd Antibody | Goat anti-rabbit PO labelled | Goat anti-rabbit PO labelled | Swine anti rabbitHRP labelled |
| Dilution 2nd Ab | 1/100 | 1/100 | 1/400 |
| Incubation 2nd Ab | 30′ | 30′ | 30′ |
| Zone Detection Parameters | |
|---|---|
| Low weight threshold (g) | 1.00 |
| Weight threshold (g) | 2.00 |
| Surface threshold | 3 |
| Stable segment detection parameter | |
| Minimum neighbor image | 3 |
| Easy scoring | |
| Stable images | 15 |
| Movement threshold | 4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Persoons, E.; De Clercq, K.; Van den Eynde, C.; Pinto, S.J.P.c.; Luyten, K.; Van Bree, R.; Tomassetti, C.; Voets, T.; Vriens, J. Mimicking Sampson’s Retrograde Menstrual Theory in Rats: A New Rat Model for Ongoing Endometriosis-Associated Pain. Int. J. Mol. Sci. 2020, 21, 2326. https://doi.org/10.3390/ijms21072326
Persoons E, De Clercq K, Van den Eynde C, Pinto SJPc, Luyten K, Van Bree R, Tomassetti C, Voets T, Vriens J. Mimicking Sampson’s Retrograde Menstrual Theory in Rats: A New Rat Model for Ongoing Endometriosis-Associated Pain. International Journal of Molecular Sciences. 2020; 21(7):2326. https://doi.org/10.3390/ijms21072326
Chicago/Turabian StylePersoons, Eleonora, Katrien De Clercq, Charlotte Van den Eynde, Sílvia João Poseiro coutinho Pinto, Katrien Luyten, Rita Van Bree, Carla Tomassetti, Thomas Voets, and Joris Vriens. 2020. "Mimicking Sampson’s Retrograde Menstrual Theory in Rats: A New Rat Model for Ongoing Endometriosis-Associated Pain" International Journal of Molecular Sciences 21, no. 7: 2326. https://doi.org/10.3390/ijms21072326
APA StylePersoons, E., De Clercq, K., Van den Eynde, C., Pinto, S. J. P. c., Luyten, K., Van Bree, R., Tomassetti, C., Voets, T., & Vriens, J. (2020). Mimicking Sampson’s Retrograde Menstrual Theory in Rats: A New Rat Model for Ongoing Endometriosis-Associated Pain. International Journal of Molecular Sciences, 21(7), 2326. https://doi.org/10.3390/ijms21072326