Origins of Alterations to Rankl Null Mutant Mouse Dental Root Development

 , ,
, ,  ,
,  and
and
Abstract
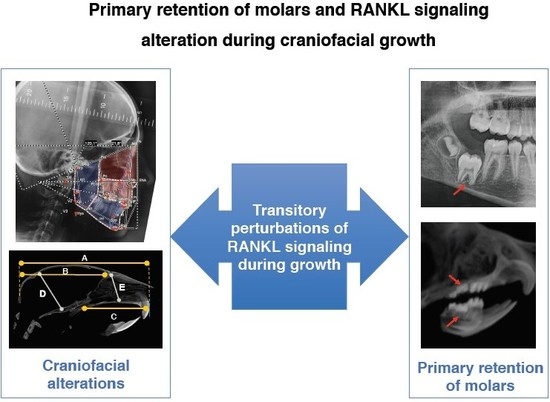
1. Introduction
2. Results
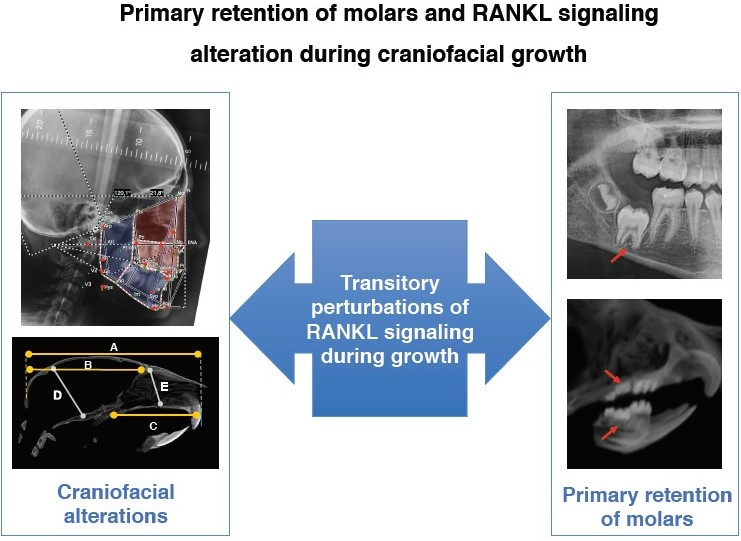
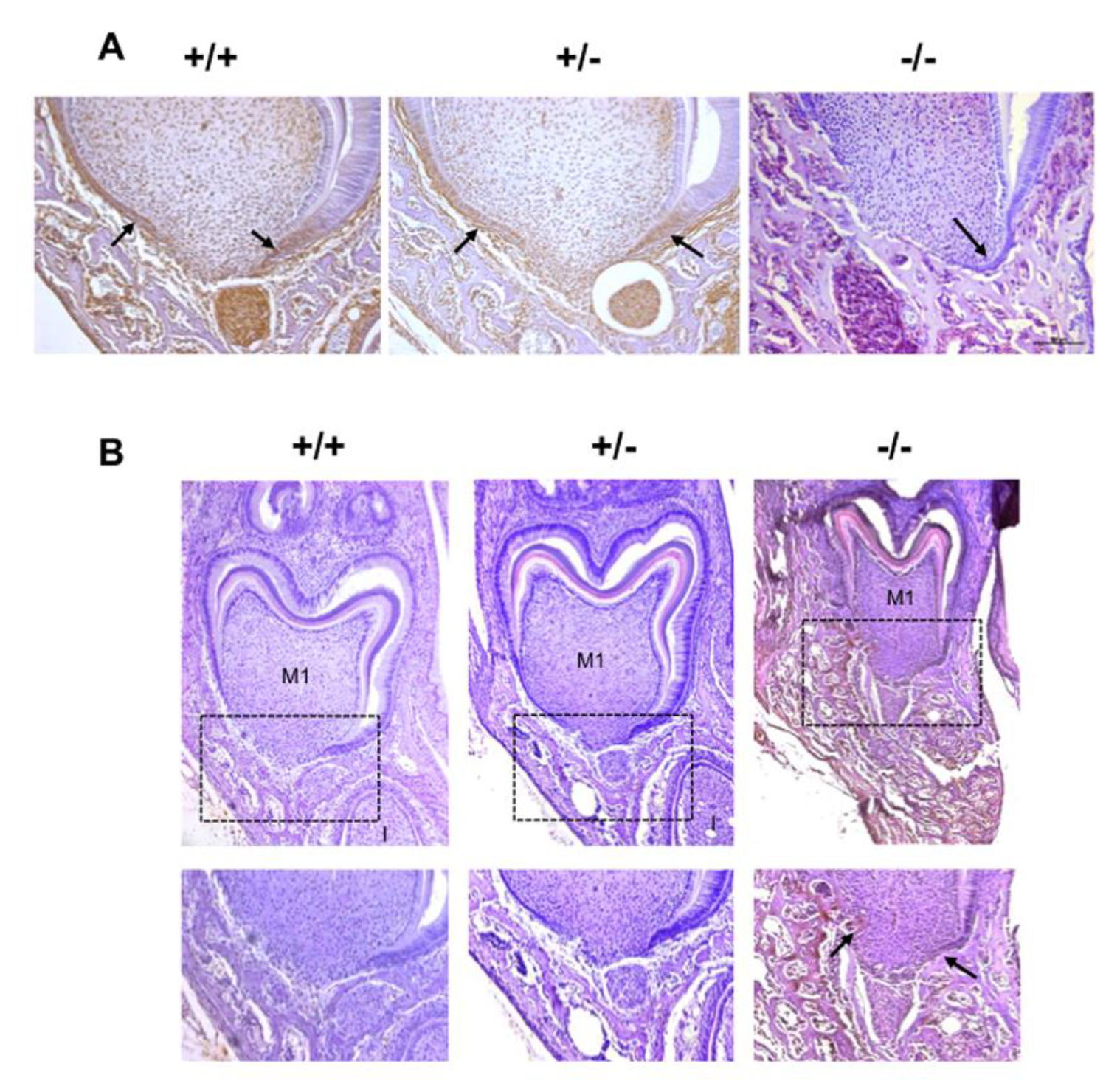
2.1. Phenotypic Alterations in the Dento-Alveolar Bone Complex Associated with the Different Rankl Genotypes
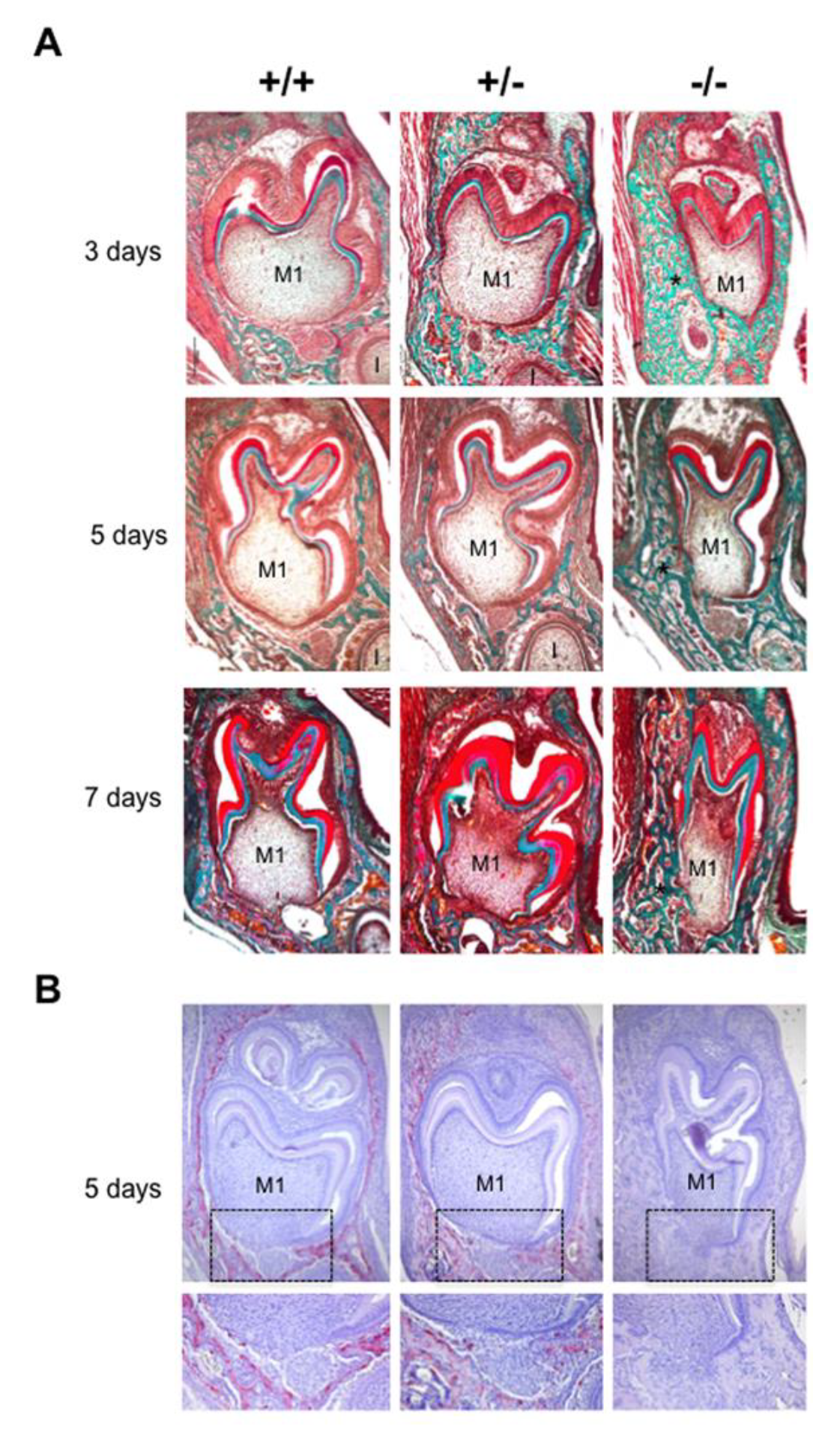
2.2. Alterations in Hertwig’s Epithelial Root Sheath Elongation Associated with the Different Rankl Genotypes
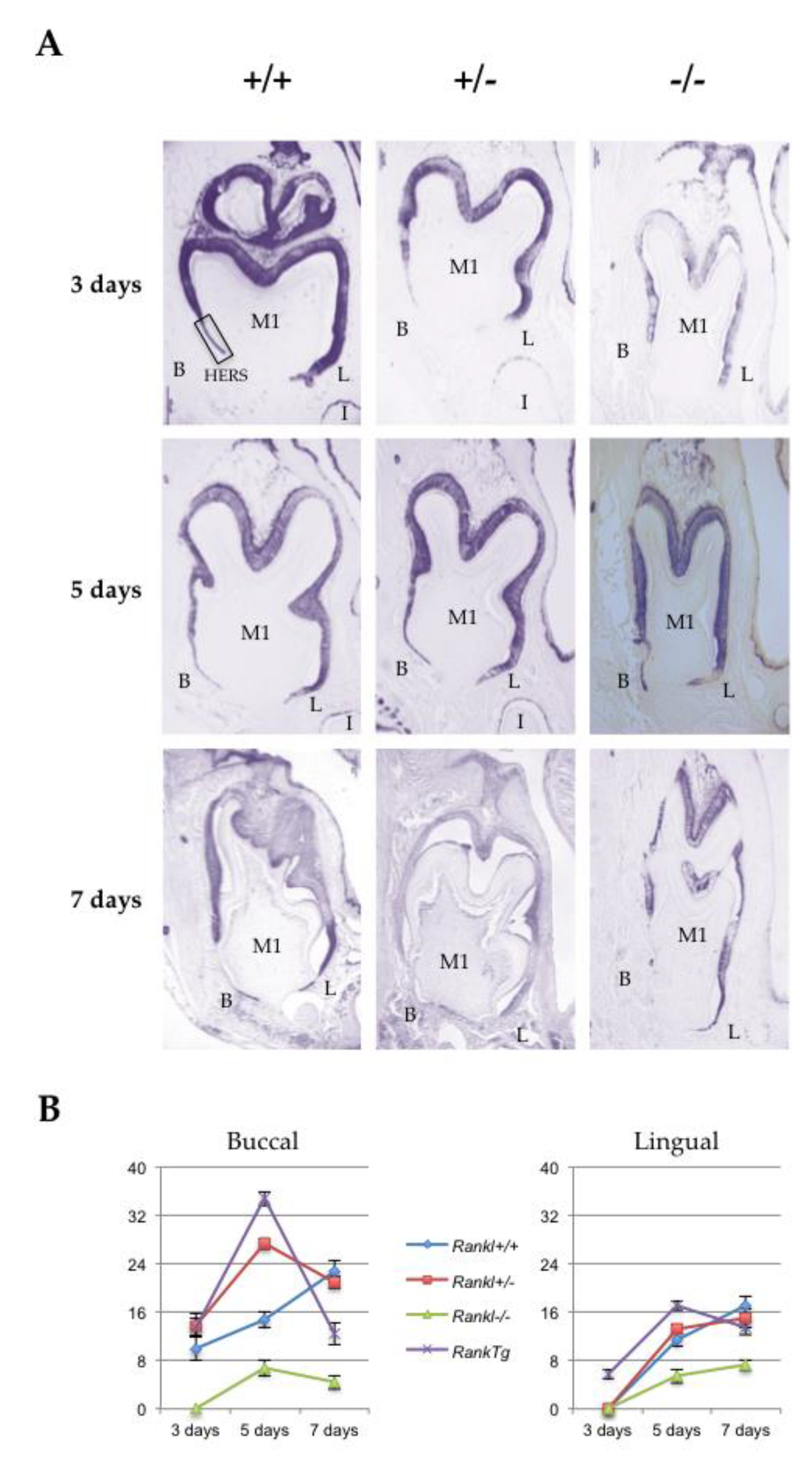
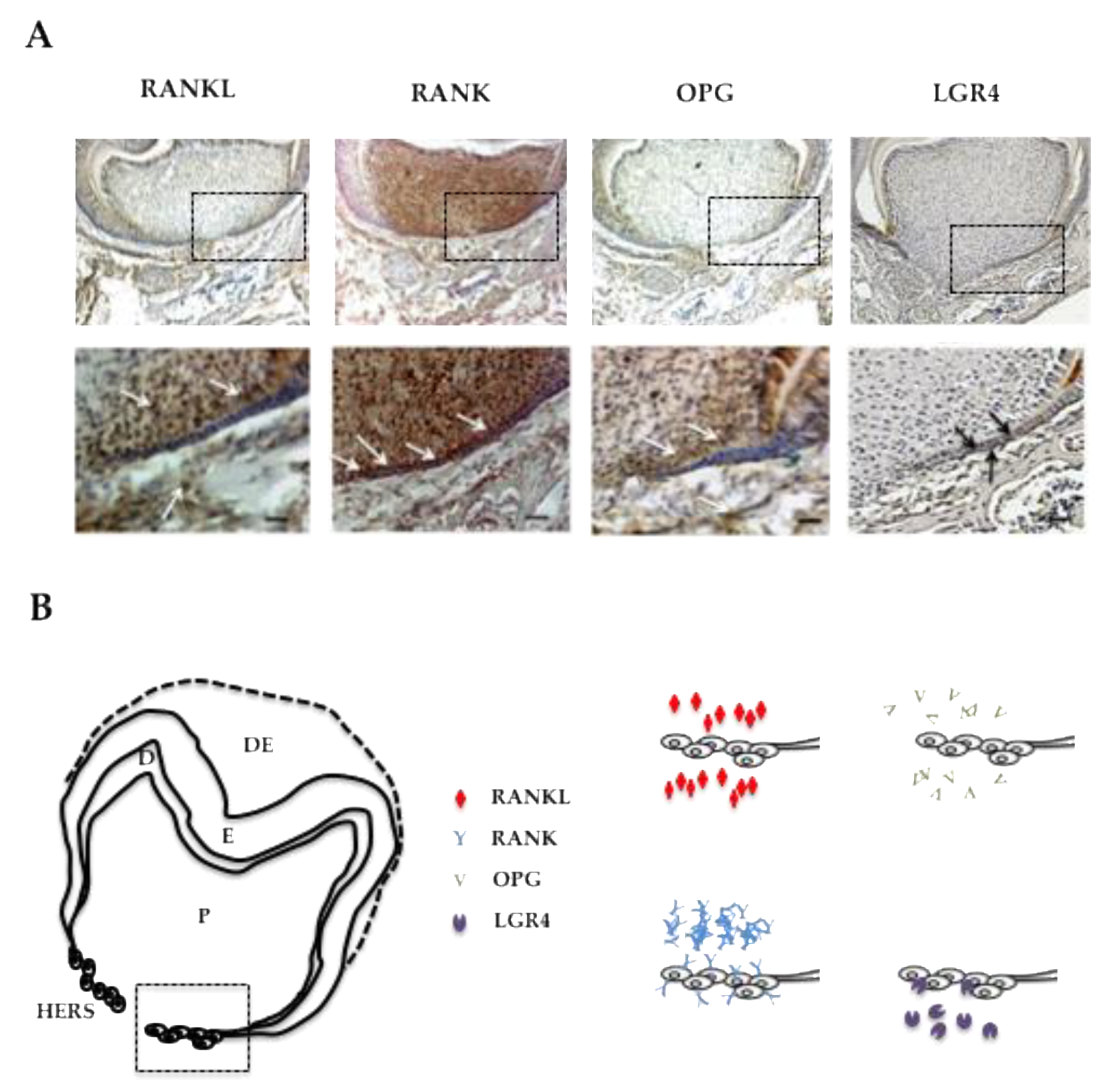
2.3. RANKL Expression Patterns and the Three Receptors, Namely RANK, OPG and LGR4 in the Mandible First Molar Mesial Root of 5-Day-Old Mice Supported the Implication of RANKL in the Proliferation of HERS Cells
2.4. Alterations to HERS Cell Proliferation Associated with the Different Rankl Genotypes
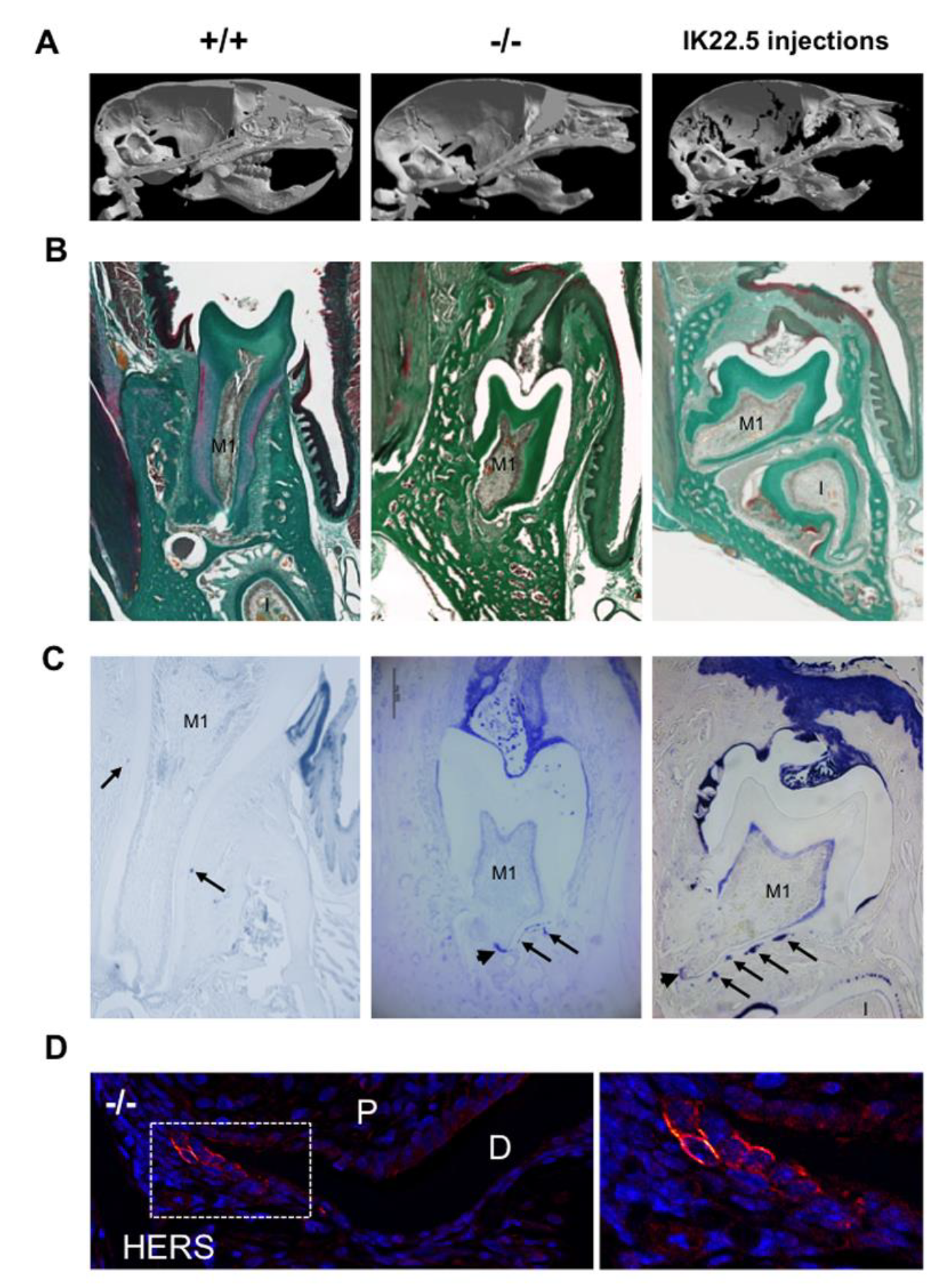
2.5. Comparison of the Mandible first Molar Phenotypes between 35-Day-Old Wild Type, Rankl-/- and RANKL Transitory Defective Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Micro-CT Analyses
4.3. Histology
4.4. Immunohistochemistry
4.5. Immunofluorescence
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HERS | Hertwig’s epithelial root sheath |
| LGR4 | Leucine rich repeat containing G protein-coupled receptor 4 |
| OPG | Osteoprotegerin |
| PCNA | Proliferating cell nuclear antigen |
| P21 | P21Waf-1/Cip-1/Sdi-1 |
| RANK | Receptor activator of NF-κB |
| RANKL | Receptor activator of NF-κB ligand |
References
- Alfaqeeh, S.A.; Gaete, M.; Tucker, A.S. Interactions of the tooth and bone during development. J. Dent. Res. 2013, 92, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Fleischmannova, J.; Matalova, E.; Sharpe, P.T.; Misek, I.; Radlanski, R.J. Formation of the tooth-bone interface. J. Dent. Res. 2010, 89, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Gama, A.; Navet, B.; Vargas, J.W.; Castaneda, B.; Lézot, F. Bone resorption: An actor of dental and periodontal development? Front. Physiol. 2015, 6, 319. [Google Scholar] [CrossRef] [PubMed]
- Lungová, V.; Radlanski, R.J.; Tucker, A.S.; Renz, H.; Míšek, I.; Matalová, E. Tooth-bone morphogenesis during postnatal stages of mouse first molar development. J. Anat. 2011, 218, 699–716. [Google Scholar] [CrossRef] [PubMed]
- Aïoub, M.; Lézot, F.; Molla, M.; Castaneda, B.; Robert, B.; Goubin, G.; Néfussi, J.R.; Berdal, A. Msx2 -/- transgenic mice develop compound amelogenesis imperfecta, dentinogenesis imperfecta and periodental osteopetrosis. Bone 2007, 41, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Berdal, A.; Castaneda, B.; Aïoub, M.; Néfussi, J.R.; Mueller, C.; Descroix, V.; Lézot, F. Osteoclasts in the dental microenvironment: A delicate balance controls dental histogenesis. Cells Tissues Organs (Print) 2011, 194, 238–243. [Google Scholar] [CrossRef]
- Castaneda, B.; Simon, Y.; Ferbus, D.; Robert, B.; Chesneau, J.; Mueller, C.; Berdal, A.; Lézot, F. Role of RANKL (TNFSF11)-dependent osteopetrosis in the dental phenotype of Msx2 null mutant mice. PLoS ONE 2013, 8, e80054. [Google Scholar] [CrossRef]
- Castaneda, B.; Simon, Y.; Jacques, J.; Hess, E.; Choi, Y.-W.; Blin-Wakkach, C.; Mueller, C.; Berdal, A.; Lézot, F. Bone resorption control of tooth eruption and root morphogenesis: Involvement of the receptor activator of NF-κB (RANK). J. Cell. Physiol. 2011, 226, 74–85. [Google Scholar] [CrossRef]
- Vargas-Franco, J.W.; Castaneda, B.; Gama, A.; Mueller, C.G.; Heymann, D.; Rédini, F.; Lézot, F. Genetically-achieved disturbances to the expression levels of TNFSF11 receptors modulate the effects of zoledronic acid on growing mouse skeletons. Biochem. Pharmacol. 2019, 168, 133–148. [Google Scholar] [CrossRef]
- Ohazama, A.; Courtney, J.-M.; Sharpe, P.T. Opg, Rank, and Rankl in tooth development: Co-ordination of odontogenesis and osteogenesis. J. Dent. Res. 2004, 83, 241–244. [Google Scholar] [CrossRef]
- Ozaki, Y.; Koide, M.; Furuya, Y.; Ninomiya, T.; Yasuda, H.; Nakamura, M.; Kobayashi, Y.; Takahashi, N.; Yoshinari, N.; Udagawa, N. Treatment of OPG-deficient mice with WP9QY, a RANKL-binding peptide, recovers alveolar bone loss by suppressing osteoclastogenesis and enhancing osteoblastogenesis. PLoS ONE 2017, 12, e0184904. [Google Scholar] [CrossRef]
- Pacios, S.; Xiao, W.; Mattos, M.; Lim, J.; Tarapore, R.S.; Alsadun, S.; Yu, B.; Wang, C.-Y.; Graves, D.T. Osteoblast Lineage Cells Play an Essential Role in Periodontal Bone Loss Through Activation of Nuclear Factor-Kappa B. Sci. Rep. 2015, 5, 16694. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Z.-F.; Ye, W.; Wang, J.; Li, C.-H.; Liu, J.-H.; Liang, Q.-C.; Li, S.; Xu, K.; Liao, E.-Y. OPG knockout mouse teeth display reduced alveolar bone mass and hypermineralization in enamel and dentin. Arch. Oral Biol. 2010, 55, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Suda, N.; Ohyama, K. Osteoclastogenesis during mouse tooth germ development is mediated by receptor activator of NFKappa-B ligand (RANKL). J. Bone Miner. Metab. 2004, 22, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Navet, B.; Vargas-Franco, J.W.; Gama, A.; Amiaud, J.; Choi, Y.; Yagita, H.; Mueller, C.G.; Rédini, F.; Heymann, D.; Castaneda, B.; et al. Maternal RANKL Reduces the Osteopetrotic Phenotype of Null Mutant Mouse Pups. J. Clin. Med. 2018, 7, 426. [Google Scholar] [CrossRef] [PubMed]
- Lézot, F.; Chesneau, J.; Navet, B.; Gobin, B.; Amiaud, J.; Choi, Y.; Yagita, H.; Castaneda, B.; Berdal, A.; Mueller, C.G.; et al. Skeletal consequences of RANKL-blocking antibody (IK22-5) injections during growth: Mouse strain disparities and synergic effect with zoledronic acid. Bone 2015, 73, 51–59. [Google Scholar] [CrossRef]
- Dougall, W.C.; Glaccum, M.; Charrier, K.; Rohrbach, K.; Brasel, K.; De Smedt, T.; Daro, E.; Smith, J.; Tometsko, M.E.; Maliszewski, C.R.; et al. RANK is essential for osteoclast and lymph node development. Genes Dev. 1999, 13, 2412–2424. [Google Scholar] [CrossRef]
- Li, J.; Sarosi, I.; Yan, X.Q.; Morony, S.; Capparelli, C.; Tan, H.L.; McCabe, S.; Elliott, R.; Scully, S.; Van, G.; et al. RANK is the intrinsic hematopoietic cell surface receptor that controls osteoclastogenesis and regulation of bone mass and calcium metabolism. Proc. Natl. Acad. Sci. USA 2000, 97, 1566–1571. [Google Scholar] [CrossRef]
- Luo, J.; Yang, Z.; Ma, Y.; Yue, Z.; Lin, H.; Qu, G.; Huang, J.; Dai, W.; Li, C.; Zheng, C.; et al. LGR4 is a receptor for RANKL and negatively regulates osteoclast differentiation and bone resorption. Nat. Med. 2016, 22, 539–546. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, W.; Zhou, X.; Li, D.; Weng, J.; Yi, Z.; Cho, S.G.; Li, C.; Yi, T.; Wu, X.; et al. Regulation of bone formation and remodeling by G-protein-coupled receptor 48. Development 2009, 136, 2747–2756. [Google Scholar] [CrossRef]
- Sun, P.; Jia, K.; Zheng, C.; Zhu, X.; Li, J.; He, L.; Siwko, S.; Xue, F.; Liu, M.; Luo, J. Loss of Lgr4 inhibits differentiation, migration and apoptosis, and promotes proliferation in bone mesenchymal stem cells. J. Cell. Physiol. 2019, 234, 10855–10867. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Zheng, X.-F.; Yang, Y.-H.; Li, B.; Wang, Y.-R.; Jiang, S.-D.; Jiang, L.-S. LGR4 acts as a key receptor for R-spondin 2 to promote osteogenesis through Wnt signaling pathway. Cell. Signal. 2016, 28, 989–1000. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Odgren, P.R.; Kim, D.K.; Marks, S.C.; Choi, Y. Diverse roles of the tumor necrosis factor family member TRANCE in skeletal physiology revealed by TRANCE deficiency and partial rescue by a lymphocyte-expressed TRANCE transgene. Proc. Natl. Acad. Sci. USA 2000, 97, 10905–10910. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, M.; Porntaveetus, T.; Kawasaki, K.; Oommen, S.; Otsuka-Tanaka, Y.; Hishinuma, M.; Nomoto, T.; Maeda, T.; Takubo, K.; Suda, T.; et al. R-spondins/Lgrs expression in tooth development. Dev. Dyn. 2014, 243, 844–851. [Google Scholar] [CrossRef]
- Yamakami, Y.; Kohashi, K.; Oyama, K.; Mohri, Y.; Hidema, S.; Nishimori, K. LGR4 is required for sequential molar development. Biochem. Biophys. Rep. 2016, 8, 174–183. [Google Scholar] [CrossRef][Green Version]
- Zhou, M.; Guo, S.; Yuan, L.; Zhang, Y.; Zhang, M.; Chen, H.; Lu, M.; Yang, J.; Ma, J. Blockade of LGR4 inhibits proliferation and odonto/osteogenic differentiation of stem cells from apical papillae. J. Mol. Histol. 2017, 48, 389–401. [Google Scholar] [CrossRef]
- Fernandez-Valdivia, R.; Lydon, J.P. From the ranks of mammary progesterone mediators, RANKL takes the spotlight. Mol. Cell. Endocrinol. 2012, 357, 91–100. [Google Scholar] [CrossRef]
- Sobacchi, C.; Menale, C.; Villa, A. The RANKL-RANK Axis: A Bone to Thymus Round Trip. Front. Immunol. 2019, 10, 629. [Google Scholar] [CrossRef]
- Duheron, V.; Hess, E.; Duval, M.; Decossas, M.; Castaneda, B.; Klöpper, J.E.; Amoasii, L.; Barbaroux, J.-B.; Williams, I.R.; Yagita, H.; et al. Receptor activator of NF-kappaB (RANK) stimulates the proliferation of epithelial cells of the epidermo-pilosebaceous unit. Proc. Natl. Acad. Sci. USA 2011, 108, 5342–5347. [Google Scholar] [CrossRef]
- Bargman, R.; Posham, R.; Boskey, A.; Carter, E.; DiCarlo, E.; Verdelis, K.; Raggio, C.; Pleshko, N. High- and low-dose OPG-Fc cause osteopetrosis-like changes in infant mice. Pediatr. Res. 2012, 72, 495–501. [Google Scholar] [CrossRef][Green Version]
- Huang, H.; Wang, J.; Zhang, Y.; Zhu, G.; Li, Y.-P.; Ping, J.; Chen, W. Bone resorption deficiency affects tooth root development in RANKL mutant mice due to attenuated IGF-1 signaling in radicular odontoblasts. Bone 2018, 114, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.Y.; Yoshida, H.; Sarosi, I.; Tan, H.L.; Timms, E.; Capparelli, C.; Morony, S.; Oliveira-dos-Santos, A.J.; Van, G.; Itie, A.; et al. OPGL is a key regulator of osteoclastogenesis, lymphocyte development and lymph-node organogenesis. Nature 1999, 397, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Odgren, P.R.; Kim, N.; MacKay, C.A.; Mason-Savas, A.; Choi, Y.; Marks, S.C. The role of RANKL (TRANCE/TNFSF11), a tumor necrosis factor family member, in skeletal development: Effects of gene knockout and transgenic rescue. Connect. Tissue Res. 2003, 44 (Suppl. 1), 264–271. [Google Scholar] [CrossRef] [PubMed]
- Navet, B.; Ando, K.; Vargas-Franco, J.W.; Brion, R.; Amiaud, J.; Mori, K.; Yagita, H.; Mueller, C.G.; Verrecchia, F.; Dumars, C.; et al. The Intrinsic and Extrinsic Implications of RANKL/RANK Signaling in Osteosarcoma: From Tumor Initiation to Lung Metastases. Cancers 2018, 10, 398. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rankl+/+ | Rankl+/- | Rankl-/- | RankTg | ||
|---|---|---|---|---|---|
| LINGUAL | 3 days | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 5.8 ± 0.8 |
| 5 days | 11.4 ± 1.1 | 13.2 ± 0.8 | 5.4 ± 1.1 | 17.2 ± 0.8 | |
| 7 days | 17.2 ± 1.5 | 15.0 ± 1.6 | 7.2 ± 0.8 | 13.4 ± 1.1 | |
| BUCCAL | 3 days | 10.0 ± 2.0 | 13.8 ± 1.9 | 0.0 ± 0.0 | 13.6 ± 1.5 |
| 5 days | 14.8 ± 1.3 | 27.4 ± 0.9 | 6.8 ± 1.3 | 34.8 ± 1.1 | |
| 7 days | 22.8 ± 1.9 | 21.0 ± 1.0 | 4.4 ± 1.1 | 12.4 ± 1.8 | |
| PCNA Labeling Intensity | |||||
|---|---|---|---|---|---|
| - | + | ++ | +++ | ||
| Rankl+/+ | 3 days | 1 | 4 | ||
| 4 days | 1 | 3 | 1 | ||
| 5 days | 2 | 2 | 1 | ||
| 6 days | 1 | 2 | 2 | ||
| 7 days | 3 | 2 | |||
| Rankl+/- | 3 days | 1 | 2 | 2 | |
| 4 days | 2 | 1 | 2 | ||
| 5 days | 1 | 3 | 1 | ||
| 6 days | 2 | 2 | 1 | ||
| 7 days | 3 | 2 | |||
| Rankl-/- | 3 days | 3 | 2 | ||
| 4 days | 4 | 1 | |||
| 5 days | 5 | ||||
| 6 days | 5 | ||||
| 7 days | 5 | ||||
| P21 Labeling Intensity | |||||
|---|---|---|---|---|---|
| - | + | ++ | +++ | ||
| Rankl+/+ | 3 days | 1 | 4 | ||
| 4 days | 1 | 4 | |||
| 5 days | 1 | 4 | |||
| 6 days | 1 | 4 | |||
| 7 days | 1 | 3 | 1 | ||
| Rankl+/- | 3 days | 5 | |||
| 4 days | 1 | 4 | |||
| 5 days | 1 | 4 | |||
| 6 days | 1 | 4 | |||
| 7 days | 2 | 3 | |||
| Rankl-/- | 3 days | 5 | |||
| 4 days | 5 | ||||
| 5 days | 5 | ||||
| 6 days | 5 | ||||
| 7 days | 5 | ||||
| Primary Antibodies | |||
|---|---|---|---|
| Anti-mouse RANK | Goat polyclonal IgG | R&D AF692 | 1/20 |
| Anti-human/mouse RANKL | Rabbit polyclonal IgG | Abcam ab62516 | 1/20 |
| Anti-mouse OPG | Goat polyclonal IgG | R&D AF805 | 1/10 |
| Anti-mouse LGR4 | Rabbit polyclonal IgG | ThermoFischer PA5-67868 | 1/100 |
| Anti-mouse PCNA | Rabbit polyclonal IgG | Abcam ab2426 | 1/500 |
| Anti-mouse P21 | Rabbit polyclonal IgG | Santa Cruz Biotechnology SC-397 | 1/50 |
| Anti-mouse KERATIN-14 | Rabbit polyclonal IgG | Covance AF64 | 1/500 - 1/1000 |
| Secondary antibodies | |||
| Bovine anti-goat | Biotin-SP conjugated polyclonal whole IgG | Jakson Immuno Research 805-065-180 | 1/400 |
| Goat anti-rabbit | Peroxidase conjugated polyclonal IgG | Dako P0448 | 1/200 - 1/500 |
| Horse anti-rabbit | Biotin conjugated polyclonal IgG | Vector laboratories BA-1100 | 7/1000 |
| Goat anti-rabbit | Alexa Fluor 594 conjugated polyclonal IgG | Life Technologies A-11072 | 1/500 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gama, A.; Vargas-Franco, J.W.; Sánchez Mesa, D.C.; Restrepo Bedoya, E.; Amiaud, J.; Babajko, S.; Berdal, A.; Acevedo, A.C.; Heymann, D.; Lézot, F.; et al. Origins of Alterations to Rankl Null Mutant Mouse Dental Root Development. Int. J. Mol. Sci. 2020, 21, 2201. https://doi.org/10.3390/ijms21062201
Gama A, Vargas-Franco JW, Sánchez Mesa DC, Restrepo Bedoya E, Amiaud J, Babajko S, Berdal A, Acevedo AC, Heymann D, Lézot F, et al. Origins of Alterations to Rankl Null Mutant Mouse Dental Root Development. International Journal of Molecular Sciences. 2020; 21(6):2201. https://doi.org/10.3390/ijms21062201
Chicago/Turabian StyleGama, Andrea, Jorge William Vargas-Franco, Diana Carolina Sánchez Mesa, Elizabeth Restrepo Bedoya, Jérome Amiaud, Sylvie Babajko, Ariane Berdal, Ana Carolina Acevedo, Dominique Heymann, Frédéric Lézot, and et al. 2020. "Origins of Alterations to Rankl Null Mutant Mouse Dental Root Development" International Journal of Molecular Sciences 21, no. 6: 2201. https://doi.org/10.3390/ijms21062201
APA StyleGama, A., Vargas-Franco, J. W., Sánchez Mesa, D. C., Restrepo Bedoya, E., Amiaud, J., Babajko, S., Berdal, A., Acevedo, A. C., Heymann, D., Lézot, F., & Castaneda, B. (2020). Origins of Alterations to Rankl Null Mutant Mouse Dental Root Development. International Journal of Molecular Sciences, 21(6), 2201. https://doi.org/10.3390/ijms21062201