Modulation of NLRP3 Inflammasome through Formyl Peptide Receptor 1 (Fpr-1) Pathway as a New Therapeutic Target in Bronchiolitis Obliterans Syndrome

 ,
, 
 ,
,  ,
,  ,
,  ,
,  , and
, and 
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
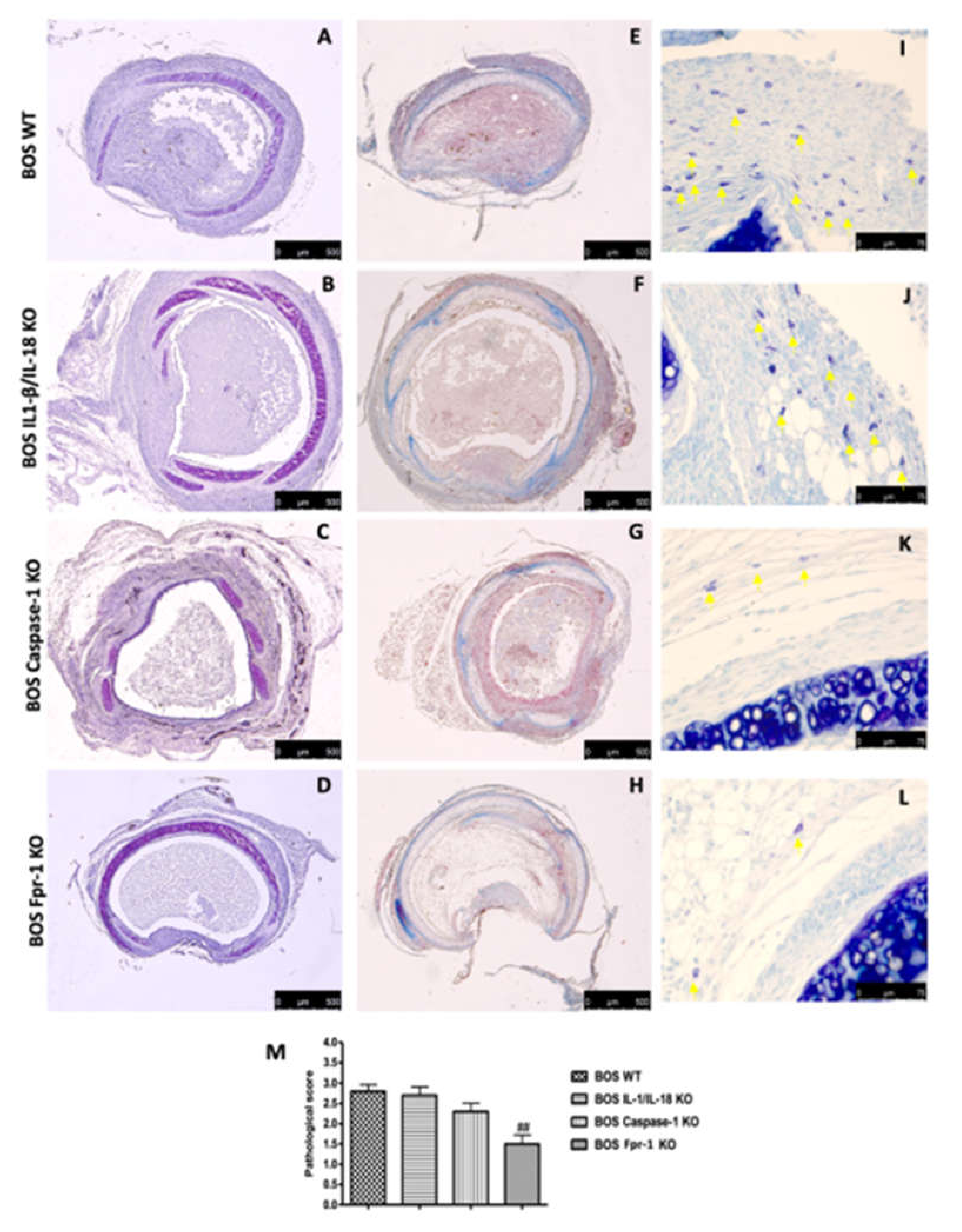
2.1. Histopatology Evaluation and Mast Cell Density in IL-1β/IL-18 KO, Casp-1 KO, and Fpr-1 KO
2.2. Effects of the Absence of IL-1β/IL-18, Casp-1, and Fpr-1 on the NLRP3 Inflammasome Pathway
2.3. Effects of the Absence of IL-1β/IL-18, Casp-1, and Fpr-1 on Nitrotyrosine Formation and PARP Activation
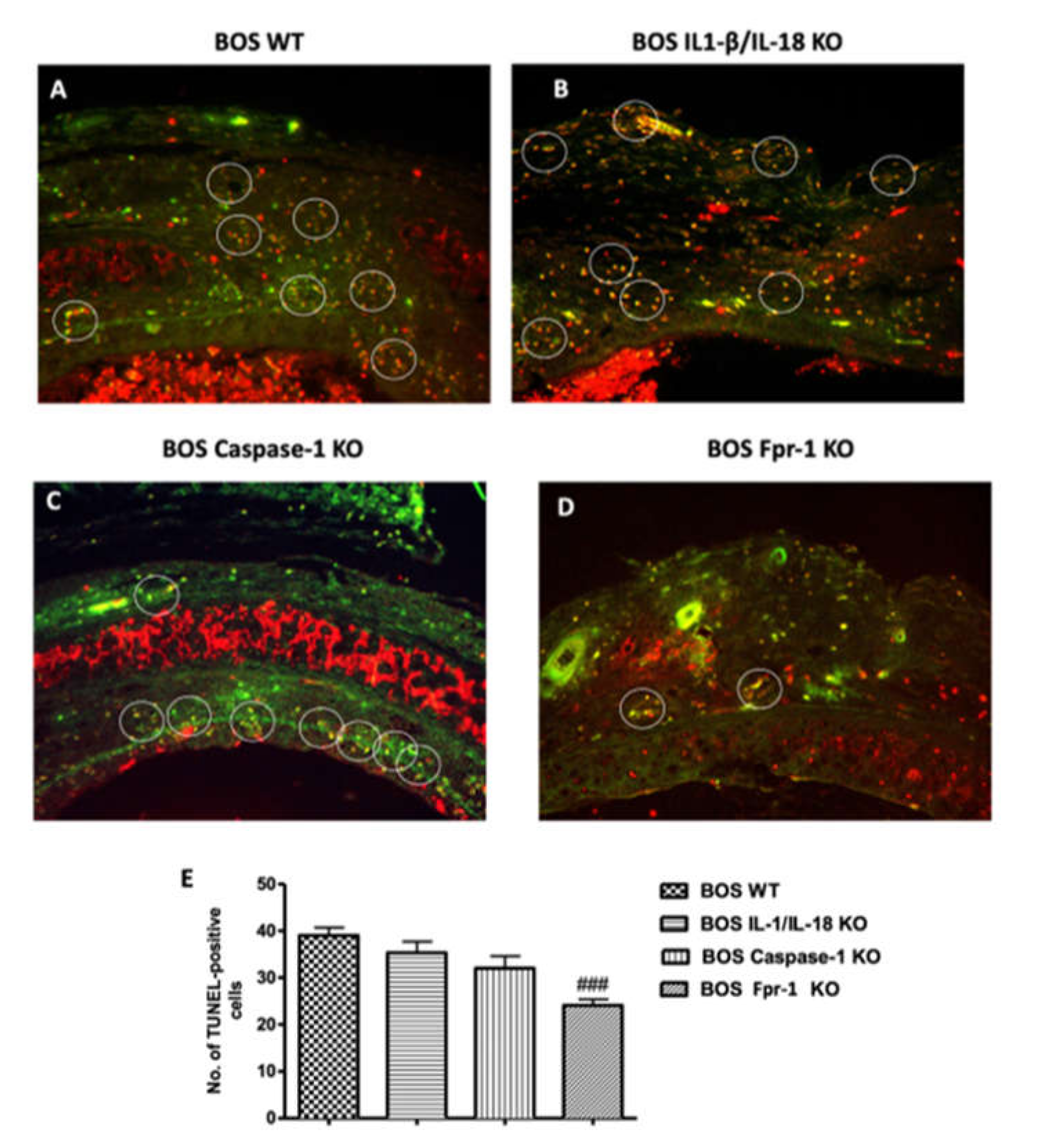
2.4. Effects of the Absence of IL-1β/IL-18, Casp-1, and Fpr-1 on Apoptosis
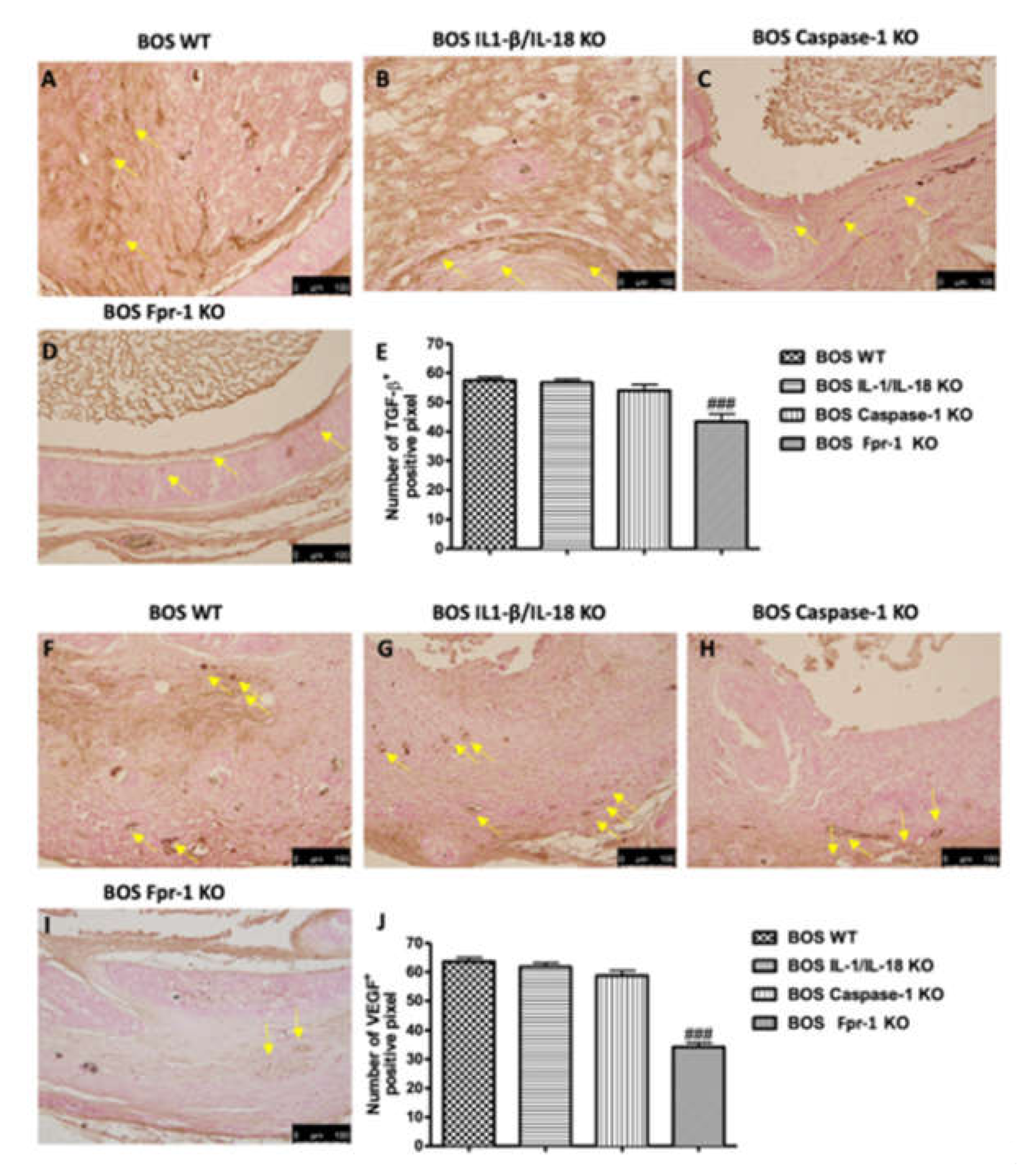
2.5. Effects of the Absence of IL-1β/IL-18, Casp-1, and Fpr-1 on Grow Factors Expression
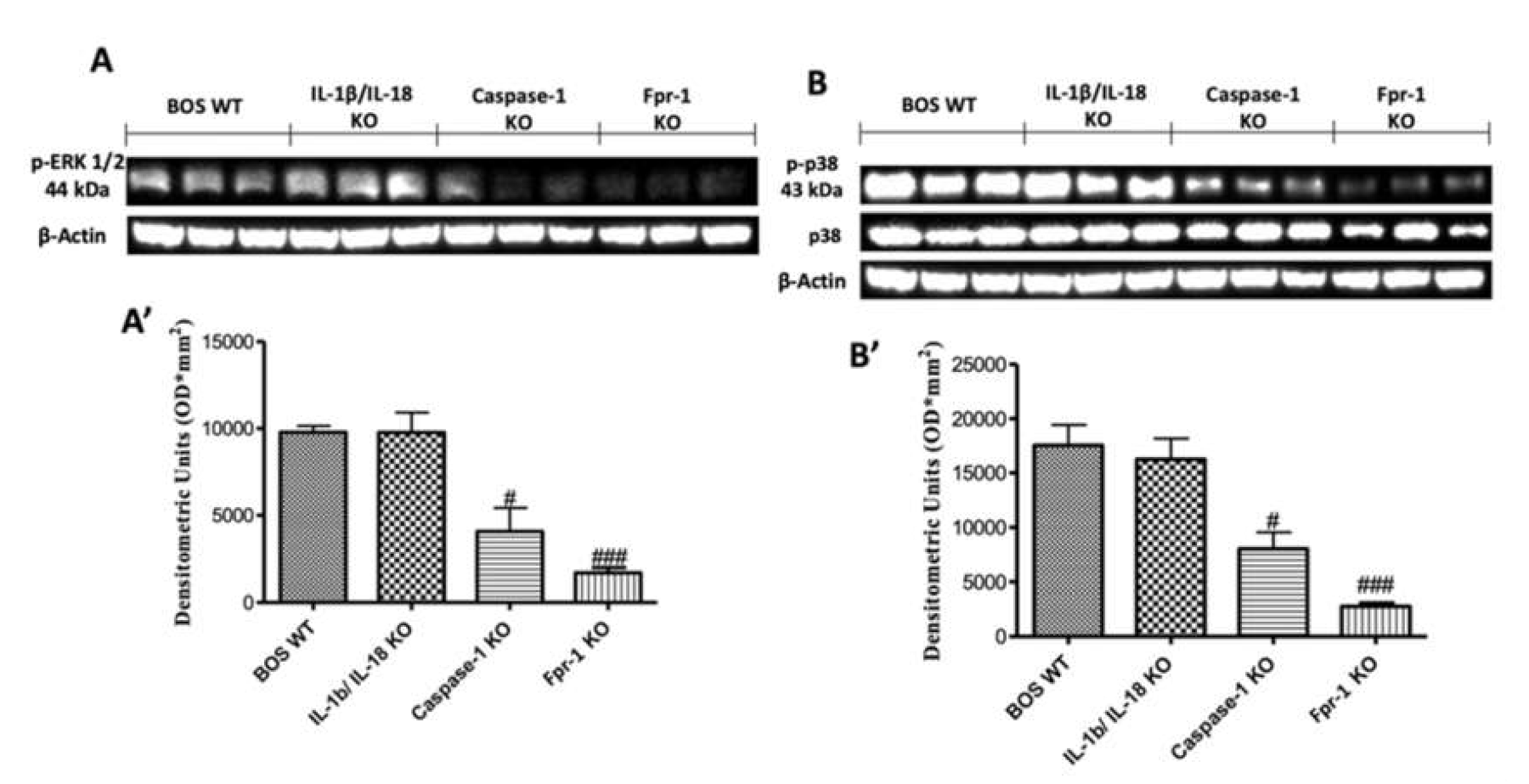
2.6. Effects of the Absence of IL-1β/IL-18, Casp-1, and Fpr-1 on the Mitogen-Activated Protein Kinase (MAPK) Pathway
3. Discussion
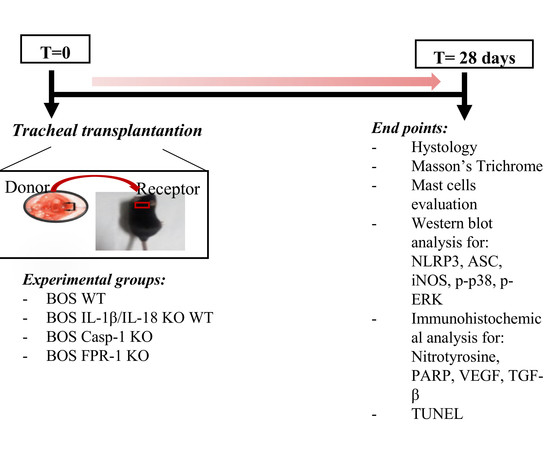
4. Materials and Methods
4.1. Animals
4.2. Animal Model of Tracheal Transplantation
4.3. Experimental Groups
- The BOS WT group: mice were subjected to tracheal transplantation as described above.
- The BOS IL-1β/IL-18 KO group: mice were subjected to tracheal transplantation as described above, as well as the WT group.
- The BOS Casp-1 KO group: mice were subjected to tracheal transplantation as described above, as well as the WT group.
- The BOS Fpr-1 KO group: mice were subjected to tracheal transplantation as described above, as well as the WT group.
4.4. Histopatology and Mast Cell Evaluation
4.5. Western Blot Analysis of Cytosolic and Nuclear Extracts from Tracheal Tissue
4.6. Immunohistochemical Localization of Nitrotyrosine, Poly (ADP-Ribose) Polymerase (PARP), Vascular Endothelial Growth Factor (VEGF), and Transforming Growth Factor-Beta (TGF-β)
4.7. Terminal Deoxynucleotidyl Nick-End Labeling (TUNEL) Assay
4.8. Materials
4.9. Statistical Evaluation
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ASC | apoptosis-associated speck-like protein containing a CARD complex |
| BOS | bronchiolitis Obliterans Syndrome |
| DAG | diacylglycerol |
| Casp-1 | Caspase-1 |
| FPR1 | n-formylpeptide receptor 1 |
| GDP | guanosine diphosphate |
| GPCR | G protein-coupled receptor |
| GTP | guanosine triphosphate |
| IL-1β | interleukin 1β |
| IL-18 | interleukin 18 |
| iNOS | inducible nitric oxide synthase |
| IP3 | 1,4,5-trisphosphate |
| MPO | myeloperoxidase |
| NF-κB | nuclear factor kappa B |
| NLRP3 | NLR Family Pyrin Domain Containing 3 |
| PBS | Phosphate buffered saline |
| PIP2 | 5-bisphosphate |
| PKC | protein kinase C |
| PLCβ | phospholipase C β |
| PMN | polymorphonuclear leukocyte |
| PMSF | phenyl-methyl sulfonyl fluoride |
| ROS | reactive oxygen species |
| SDS | sodium dodecyl sulphate |
References
- Trulock, E.P.; Christie, J.D.; Edwards, L.B.; Boucek, M.M.; Aurora, P.; Taylor, D.O.; Dobbels, F.; Rahmel, A.O.; Keck, B.M.; Hertz, M.I. Registry of the international society for heart and lung transplantation: Twenty-fourth official adult lung and heart-lung transplantation report-2007. J. Heart Lung Transpl. 2007, 26, 782–795. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.; Allen, R. Bronchiolitis obliterans: An update. Curr. Opin. Pulm. Med. 2004, 10, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Yousem, S.A. Lymphocytic bronchitis/bronchiolitis in lung allograft recipients. Am. J. Surg. Pathol. 1993, 17, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Shigenobu, T.; Ohtsuka, T.; Shimoda, M. The prevention of tracheal graft occlusion using pioglitazone: A mouse tracheal transplant model study. Transpl. Immunol. 2019, 53, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Akindipe, O.; Fernandez-Bussy, S.; Jantz, M.; Lu, L.; Deem, A.; Swafford, W.; Staples, E.; Baz, M. Obliterative bronchiolitis in lung allografts removed at retransplant for intractable airway problems. Respirology 2009, 14, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Estenne, M.; Hertz, M.I. Bronchiolitis obliterans after human lung transplantation. Am. J. Respir. Crit. Care Med. 2002, 166, 440–444. [Google Scholar] [CrossRef]
- Morrell, M.R.; Despotis, G.J.; Lublin, D.M.; Patterson, G.A.; Trulock, E.P.; Hachem, R.R. The efficacy of photopheresis for bronchiolitis obliterans syndrome after lung transplantation. J. Heart Lung Transpl. 2010, 29, 424–431. [Google Scholar] [CrossRef]
- Fisher, A.J.; Rutherford, R.M.; Bozzino, J.; Parry, G.; Dark, J.H.; Corris, P.A. The safety and efficacy of total lymphoid irradiation in progressive bronchiolitis obliterans syndrome after lung transplantation. Am. J. Transpl. 2005, 5, 537–543. [Google Scholar] [CrossRef]
- Fusco, R.; Gugliandolo, E.; Biundo, F.; Campolo, M.; Di Paola, R.; Cuzzocrea, S. Inhibition of inflammasome activation improves lung acute injury induced by carrageenan in a mouse model of pleurisy. FASEB J. 2017, 31, 3497–3511. [Google Scholar] [CrossRef]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef]
- Sun, Q.; Scott, M.J. Caspase-1 as a multifunctional inflammatory mediator: Noncytokine maturation roles. J. Leukoc. Biol. 2016, 100, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Najafov, A.; Py, B.F. Roles of caspases in necrotic cell death. Cell 2016, 167, 1693–1704. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Siracusa, R.; Cordaro, M.; Peritore, A.F.; Gugliandolo, E.; Mancuso, G.; Midiri, A.; Di Paola, R.; Cuzzocrea, S. Therapeutic potential of dinitrobenzene sulfonic acid (DNBS)-induced colitis in mice by targeting IL-1beta and IL-18. Biochem. Pharmacol. 2018, 155, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Gehrmann, J.; Matsumoto, Y.; Kreutzberg, G.W. Microglia: Intrinsic immuneffector cell of the brain. Brain Res. Brain Res. Rev. 1995, 20, 269–287. [Google Scholar] [CrossRef]
- Yusen, R.D.; Edwards, L.B.; Kucheryavaya, A.Y.; Benden, C.; Dipchand, A.I.; Dobbels, F.; Goldfarb, S.B.; Levvey, B.J.; Lund, L.H.; Meiser, B.; et al. The registry of the international society for heart and lung transplantation: Thirty-first adult lung and heart-lung transplant report—2014; focus theme: Retransplantation. J. Heart Lung Transpl. 2014, 33, 1009–1024. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, K.; Wang, C.; Gong, W.; Yoshimura, T.; Liu, M.; Wang, J.M. Cell surface receptor FPR2 promotes antitumor host defense by limiting M2 polarization of macrophages. Cancer Res. 2013, 73, 550–560. [Google Scholar] [CrossRef]
- Thabut, G.; Mal, H. Outcomes after lung transplantation. J. Thorac. Dis. 2017, 9, 2684–2691. [Google Scholar] [CrossRef]
- Khush, K.K.; Cherikh, W.S.; Chambers, D.C.; Goldfarb, S.; Hayes, D., Jr.; Kucheryavaya, A.Y.; Levvey, B.J.; Meiser, B.; Rossano, J.W.; Stehlik, J.; et al. The international thoracic organ transplant registry of the international society for heart and lung transplantation: Thirty-fifth adult heart transplantation report-2018; focus theme: Multiorgan transplantation. J. Heart Lung Transpl. 2018, 37, 1155–1168. [Google Scholar] [CrossRef]
- Belperio, J.A.; Keane, M.P.; Burdick, M.D.; Gomperts, B.; Xue, Y.Y.; Hong, K.; Mestas, J.; Ardehali, A.; Mehrad, B.; Saggar, R.; et al. Role of CXCR2/CXCR2 ligands in vascular remodeling during bronchiolitis obliterans syndrome. J. Clin. Investig. 2005, 115, 1150–1162. [Google Scholar] [CrossRef]
- McDyer, J.F. Human and murine obliterative bronchiolitis in transplant. Proc. Am. Thorac. Soc. 2007, 4, 37–43. [Google Scholar] [CrossRef]
- Boehler, A.; Kesten, S.; Weder, W.; Speich, R. Bronchiolitis obliterans after lung transplantation: A review. Chest 1998, 114, 1411–1426. [Google Scholar] [CrossRef] [PubMed]
- De Nardo, D.; De Nardo, C.M.; Latz, E. New insights into mechanisms controlling the NLRP3 inflammasome and its role in lung disease. Am. J. Pathol. 2014, 184, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Franchi, L.; Munoz-Planillo, R.; Nunez, G. Sensing and reacting to microbes through the inflammasomes. Nat. Immunol. 2012, 13, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Orihara, Y.; Ikematsu, K.; Tsuda, R.; Nakasono, I. Induction of nitric oxide synthase by traumatic brain injury. Forensic Sci. Int. 2001, 123, 142–149. [Google Scholar] [CrossRef]
- Arcasoy, S.M.; Kotloff, R.M. Lung transplantation. N. Engl. J. Med. 1999, 340, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Dixit, V.M. Mechanisms and functions of inflammasomes. Cell 2014, 157, 1013–1022. [Google Scholar] [CrossRef]
- Man, S.M.; Kanneganti, T.D. Regulation of inflammasome activation. Immunol. Rev. 2015, 265, 6–21. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Nunez, G. Mechanism and regulation of NLRP3 inflammasome activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef]
- Kigerl, K.A.; de Rivero Vaccari, J.P.; Dietrich, W.D.; Popovich, P.G.; Keane, R.W. Pattern recognition receptors and central nervous system repair. Exp. Neurol. 2014, 258, 5–16. [Google Scholar] [CrossRef]
- Weigt, S.S.; Palchevskiy, V.; Belperio, J.A. Inflammasomes and IL-1 biology in the pathogenesis of allograft dysfunction. J. Clin. Investig. 2017, 127, 2022–2029. [Google Scholar] [CrossRef]
- Mattson, M.P.; Meffert, M.K. Roles for NF-kappaB in nerve cell survival, plasticity, and disease. Cell Death Differ. 2006, 13, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Bisignano, C.; Genovese, T.; Mazzon, E.; Wickham, M.S.; Paterniti, I.; Cuzzocrea, S. Natural almond skin reduced oxidative stress and inflammation in an experimental model of inflammatory bowel disease. Int. Immunopharmacol. 2011, 11, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C. Role of poly(ADP-ribose)synthetase in inflammation. Eur. J. Pharmacol. 1998, 350, 1–19. [Google Scholar] [CrossRef]
- Tando, Y.; Ota, C.; Yamada, M.; Kamata, S.; Yamaya, M.; Kano, K.; Okudaira, S.; Aoki, J.; Kubo, H. The role of lysophosphatidic acid on airway epithelial cell denudation in a murine heterotopic tracheal transplant model. Transpl. Direct 2015, 1, e35. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, Y.; Wakeham, A.; Hao, Z.; Toba, H.; Bai, X.; Keshavjee, S.; Mak, T.W.; Liu, M. XB130 deficiency affects tracheal epithelial differentiation during airway repair. PLoS ONE 2014, 9, e108952. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; D’Amico, R.; Cordaro, M.; Gugliandolo, E.; Siracusa, R.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. Absence of formyl peptide receptor 1 causes endometriotic lesion regression in a mouse model of surgically-induced endometriosis. Oncotarget 2018, 9, 31355–31366. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, B.; Harry, L.E.; Paleolog, E.M. Modulating angiogenesis: More vs. less. JAMA 2004, 292, 972–977. [Google Scholar] [CrossRef]
- Koch, A.E. Angiogenesis as a target in rheumatoid arthritis. Ann. Rheum. Dis. 2003, 62 (Suppl. 2), ii60–ii67. [Google Scholar] [CrossRef]
- Ramirez, A.M.; Takagawa, S.; Sekosan, M.; Jaffe, H.A.; Varga, J.; Roman, J. Smad3 deficiency ameliorates experimental obliterative bronchiolitis in a heterotopic tracheal transplantation model. Am. J. Pathol. 2004, 165, 1223–1232. [Google Scholar] [CrossRef]
- Fine, A.; Goldstein, R.H. The effect of transforming growth factor-beta on cell proliferation and collagen formation by lung fibroblasts. J. Biol. Chem. 1987, 262, 3897–3902. [Google Scholar]
- Sporn, M.B.; Roberts, A.B.; Wakefield, L.M.; de Crombrugghe, B. Some recent advances in the chemistry and biology of transforming growth factor-beta. J. Cell Biol. 1987, 105, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Mora, A.L.; LaVoy, J.; McKean, M.; Stecenko, A.; Brigham, K.L.; Parker, R.; Rojas, M. Prevention of NF-kappaB activation in vivo by a cell-permeable NF-kappaB inhibitor peptide. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 289, L536–L544. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nagaraj, N.S.; Datta, P.K. Targeting the transforming growth factor-beta signaling pathway in human cancer. Expert Opin. Investig. Drugs 2010, 19, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Rice, A.B.; Ingram, J.L.; Bonner, J.C. p38 mitogen-activated protein kinase regulates growth factor-induced mitogenesis of rat pulmonary myofibroblasts. Am. J. Respir. Cell Mol. Biol. 2002, 27, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Hocevar, B.A.; Brown, T.L.; Howe, P.H. TGF-beta induces fibronectin synthesis through a c-Jun N-terminal kinase-dependent, Smad4-independent pathway. EMBO J. 1999, 18, 1345–1356. [Google Scholar] [CrossRef]
- Kelly, K.E.; Hertz, M.I.; Mueller, D.L. T-cell and major histocompatibility complex requirements for obliterative airway disease in heterotopically transplanted murine tracheas. Transplantation 1998, 66, 764–771. [Google Scholar] [CrossRef]
- Xu, J.; Torres, E.; Mora, A.L.; Shim, H.; Ramirez, A.; Neujahr, D.; Brigham, K.L.; Rojas, M. Attenuation of obliterative bronchiolitis by a CXCR4 antagonist in the murine heterotopic tracheal transplant model. J. Heart Lung Transpl. 2008, 27, 1302–1310. [Google Scholar] [CrossRef]
- Hertz, M.I.; Jessurun, J.; King, M.B.; Savik, S.K.; Murray, J.J. Reproduction of the obliterative bronchiolitis lesion after heterotopic transplantation of mouse airways. Am. J. Pathol. 1993, 142, 1945–1951. [Google Scholar]
- Murakawa, T.; Kerklo, M.M.; Zamora, M.R.; Wei, Y.; Gill, R.G.; Henson, P.M.; Grover, F.L.; Nicolls, M.R. Simultaneous LFA-1 and CD40 ligand antagonism prevents airway remodeling in orthotopic airway transplantation: Implications for the role of respiratory epithelium as a modulator of fibrosis. J. Immunol. 2005, 174, 3869–3879. [Google Scholar] [CrossRef]
- Petrosino, S.; Campolo, M.; Impellizzeri, D.; Paterniti, I.; Allara, M.; Gugliandolo, E.; D’Amico, R.; Siracusa, R.; Cordaro, M.; Esposito, E.; et al. 2-pentadecyl-2-oxazoline, the oxazoline of pea, modulates carrageenan-induced acute inflammation. Front. Pharmacol. 2017, 8, 308. [Google Scholar] [CrossRef]
- D’Amico, R.; Fusco, R.; Gugliandolo, E.; Cordaro, M.; Siracusa, R.; Impellizzeri, D.; Peritore, A.F.; Crupi, R.; Cuzzocrea, S.; Di Paola, R. Effects of a new compound containing Palmitoylethanolamide and Baicalein in myocardial ischaemia/reperfusion injury in vivo. Phytomedicine 2019, 54, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Sawant, S.; Gokulan, R.; Dongre, H.; Vaidya, M.; Chaukar, D.; Prabhash, K.; Ingle, A.; Joshi, S.; Dange, P.; Joshi, S.; et al. Prognostic role of Oct4, CD44 and c-Myc in radio-chemo-resistant oral cancer patients and their tumourigenic potential in immunodeficient mice. Clin. Oral Investig. 2016, 20, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Varghese, F.; Bukhari, A.B.; Malhotra, R.; De, A. IHC Profiler: An open source plugin for the quantitative evaluation and automated scoring of immunohistochemistry images of human tissue samples. PLoS ONE 2014, 9, e96801. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Amico, R.; Fusco, R.; Cordaro, M.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Scuto, M.; Cuzzocrea, S.; Di Paola, R.; et al. Modulation of NLRP3 Inflammasome through Formyl Peptide Receptor 1 (Fpr-1) Pathway as a New Therapeutic Target in Bronchiolitis Obliterans Syndrome. Int. J. Mol. Sci. 2020, 21, 2144. https://doi.org/10.3390/ijms21062144
D’Amico R, Fusco R, Cordaro M, Siracusa R, Peritore AF, Gugliandolo E, Crupi R, Scuto M, Cuzzocrea S, Di Paola R, et al. Modulation of NLRP3 Inflammasome through Formyl Peptide Receptor 1 (Fpr-1) Pathway as a New Therapeutic Target in Bronchiolitis Obliterans Syndrome. International Journal of Molecular Sciences. 2020; 21(6):2144. https://doi.org/10.3390/ijms21062144
Chicago/Turabian StyleD’Amico, Ramona, Roberta Fusco, Marika Cordaro, Rosalba Siracusa, Alessio Filippo Peritore, Enrico Gugliandolo, Rosalia Crupi, Maria Scuto, Salvatore Cuzzocrea, Rosanna Di Paola, and et al. 2020. "Modulation of NLRP3 Inflammasome through Formyl Peptide Receptor 1 (Fpr-1) Pathway as a New Therapeutic Target in Bronchiolitis Obliterans Syndrome" International Journal of Molecular Sciences 21, no. 6: 2144. https://doi.org/10.3390/ijms21062144
APA StyleD’Amico, R., Fusco, R., Cordaro, M., Siracusa, R., Peritore, A. F., Gugliandolo, E., Crupi, R., Scuto, M., Cuzzocrea, S., Di Paola, R., & Impellizzeri, D. (2020). Modulation of NLRP3 Inflammasome through Formyl Peptide Receptor 1 (Fpr-1) Pathway as a New Therapeutic Target in Bronchiolitis Obliterans Syndrome. International Journal of Molecular Sciences, 21(6), 2144. https://doi.org/10.3390/ijms21062144