Indole-3-Carbinol Derivative DIM Mitigates Carbon Tetrachloride-Induced Acute Liver Injury in Mice by Inhibiting Inflammatory Response, Apoptosis and Regulating Oxidative Stress



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
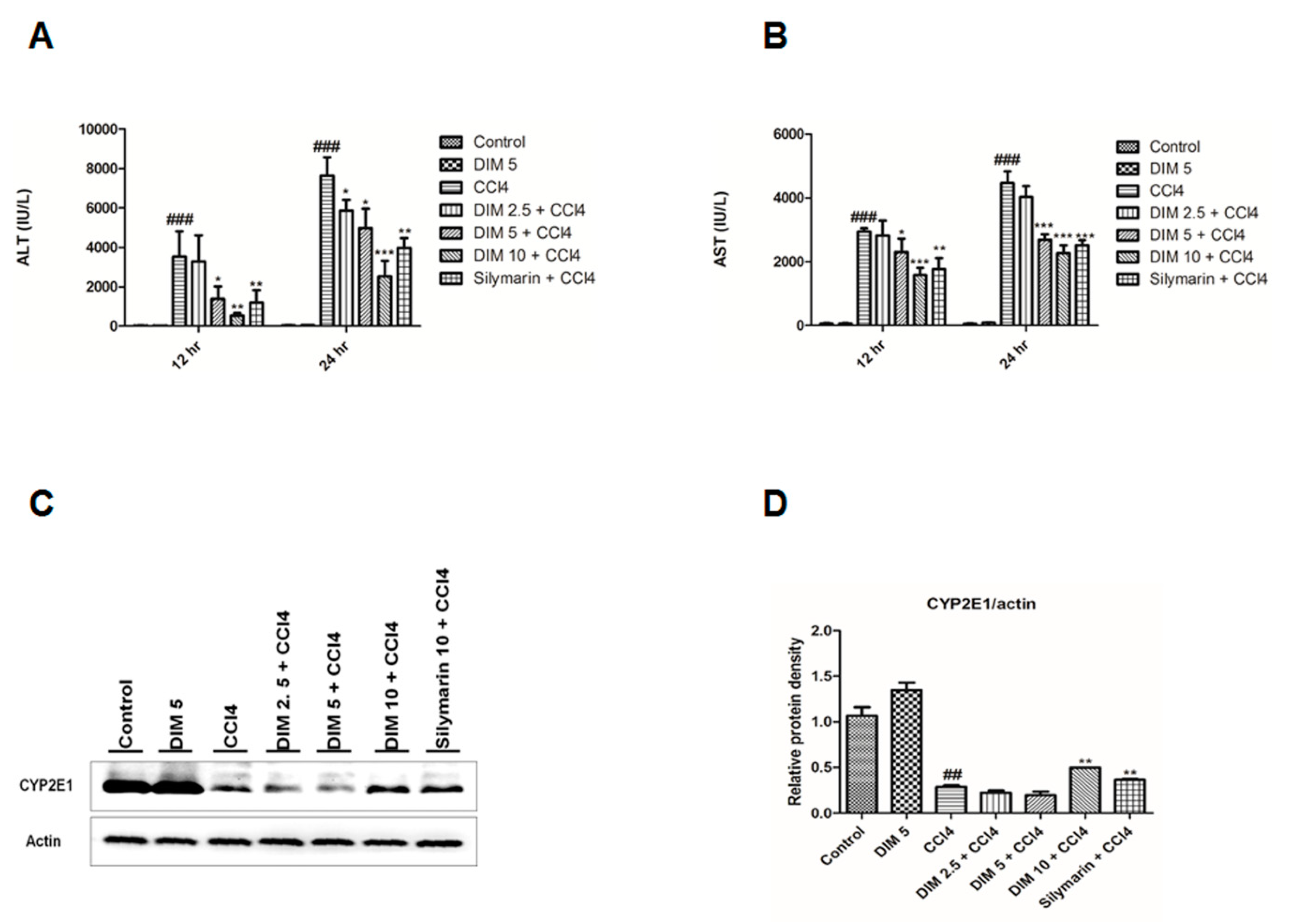
2.1. DIM Inhibits CCl4 Induced Liver Injury
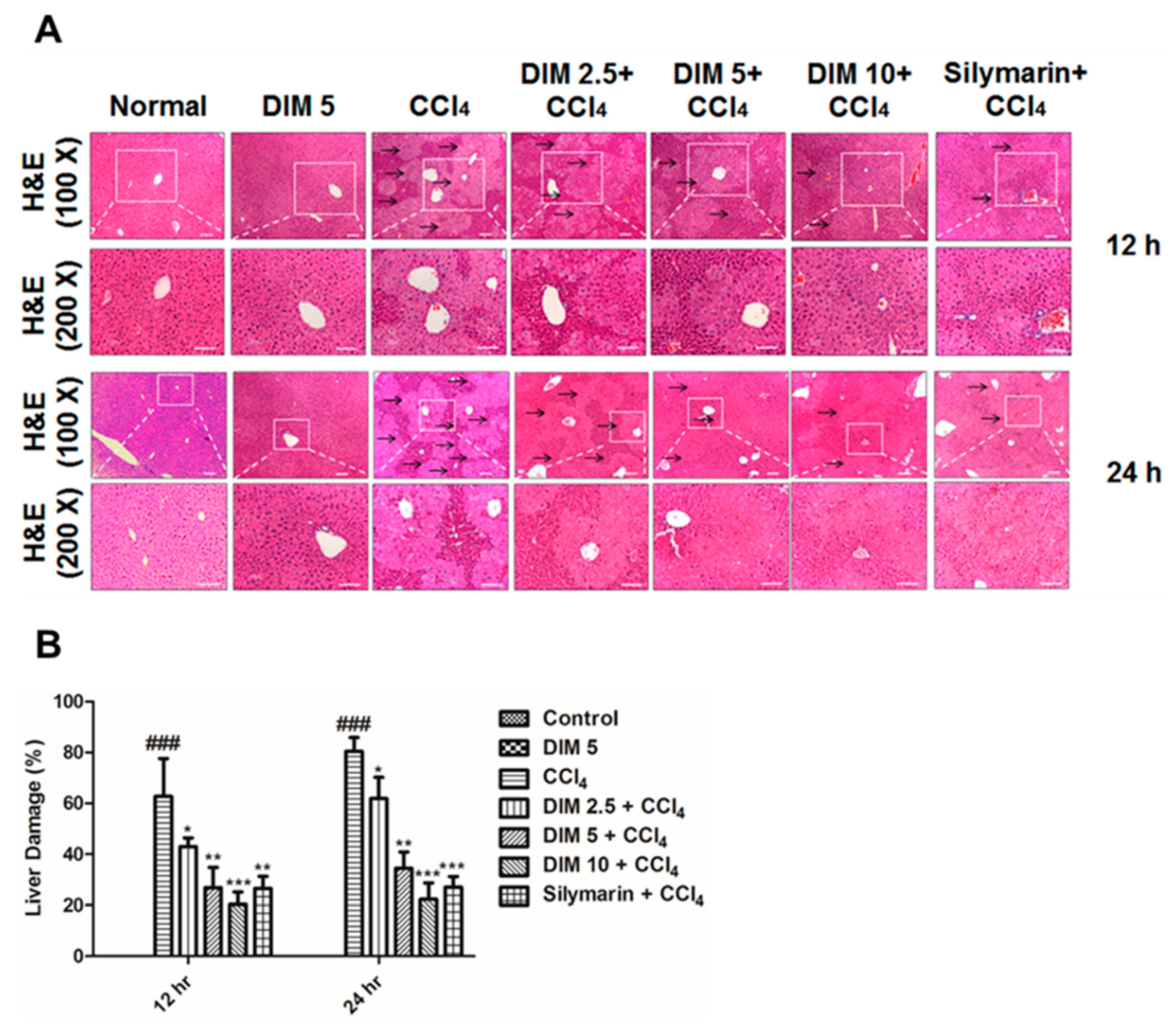
2.2. DIM Mitigates CCl4-Induced Hepatic Histopathological Damage
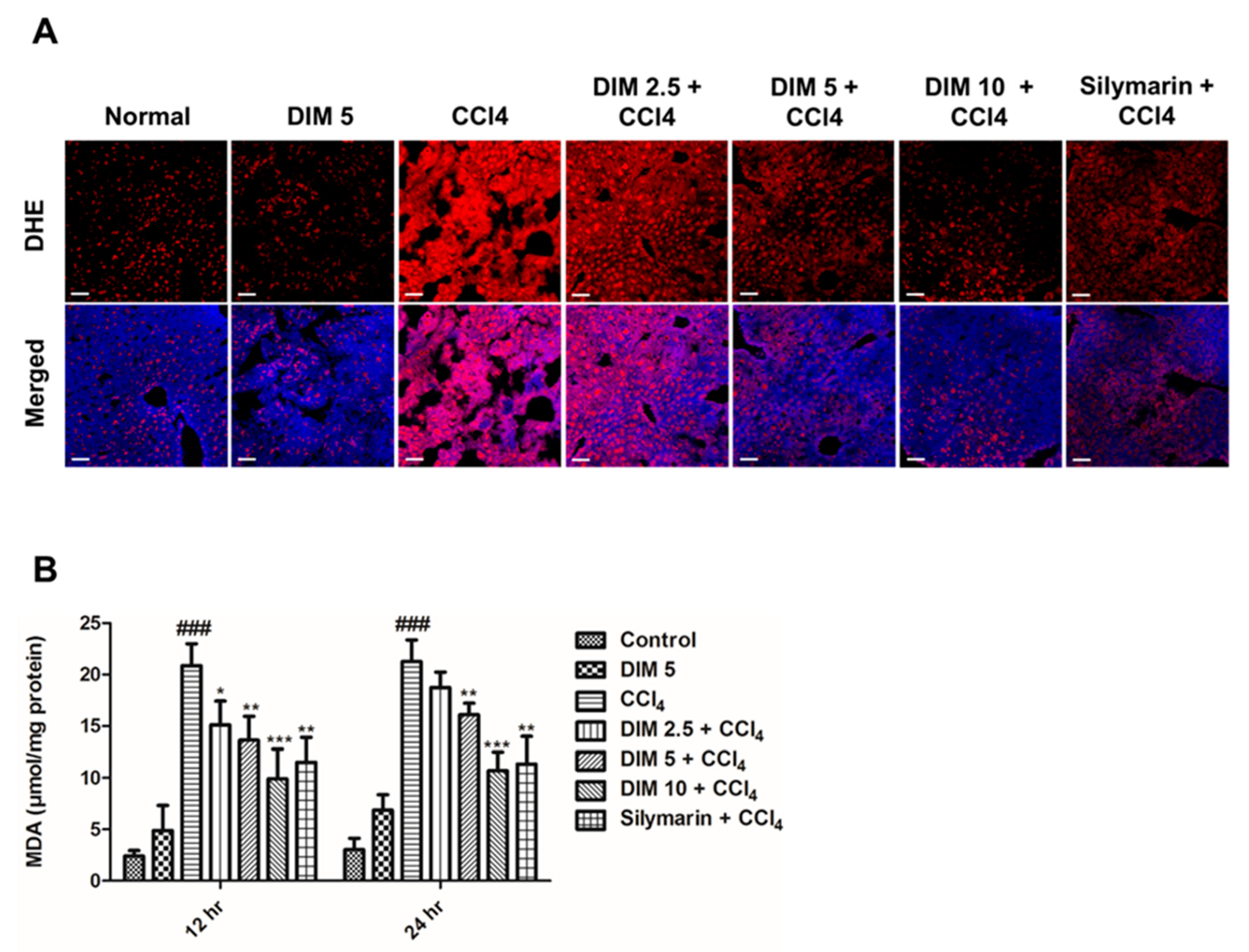
2.3. DIM Pretreatment Inhibits CCl4-Induced Oxidative Stress and ROS Production in Response to CCl4 Administration
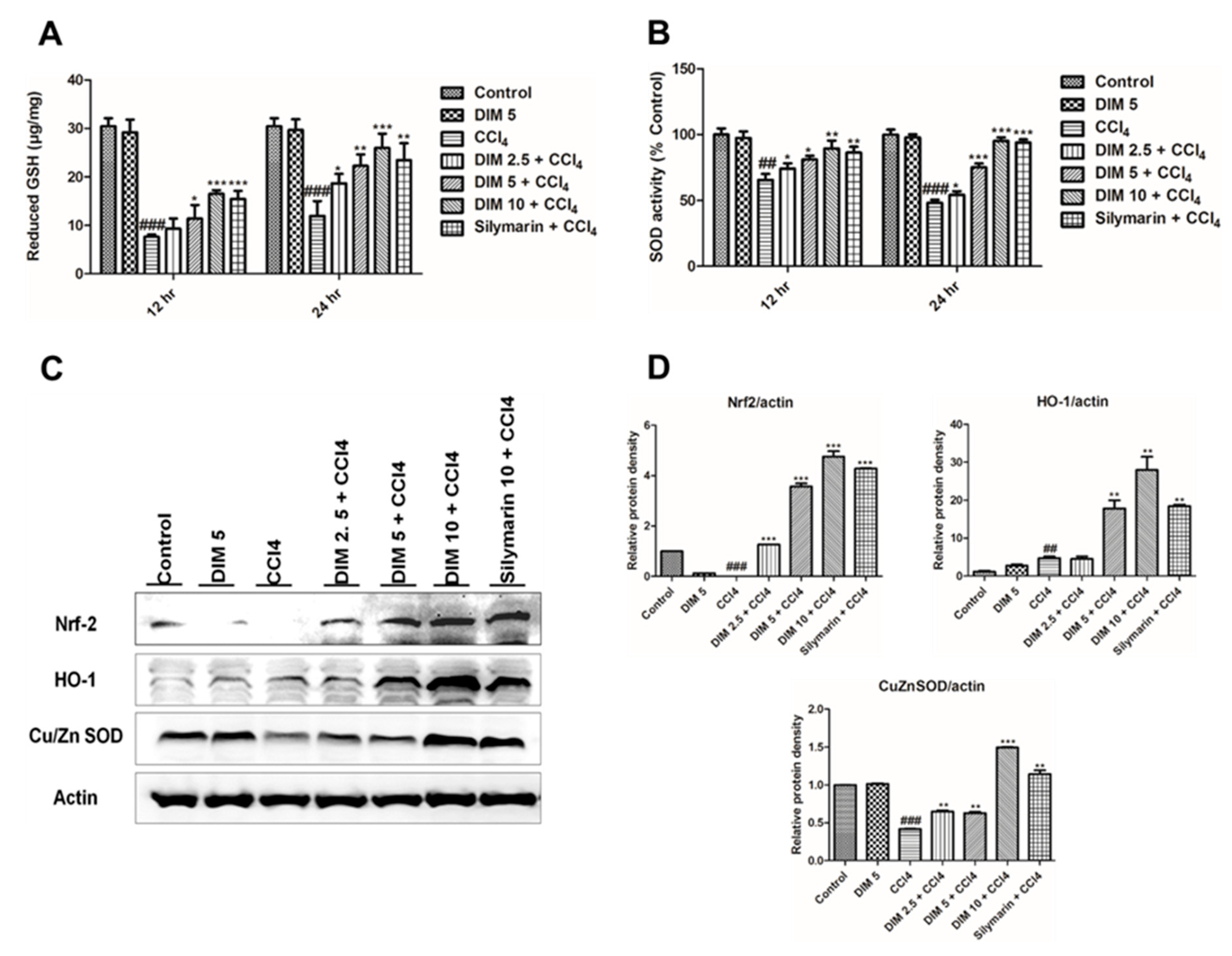
2.4. DIM Pre-Treatment Modulates Antioxidant Activity by Regulating the Nrf2/HO-1 Signaling Pathway and Inhibits Oxidative Stress in Response to CCl4 Administration
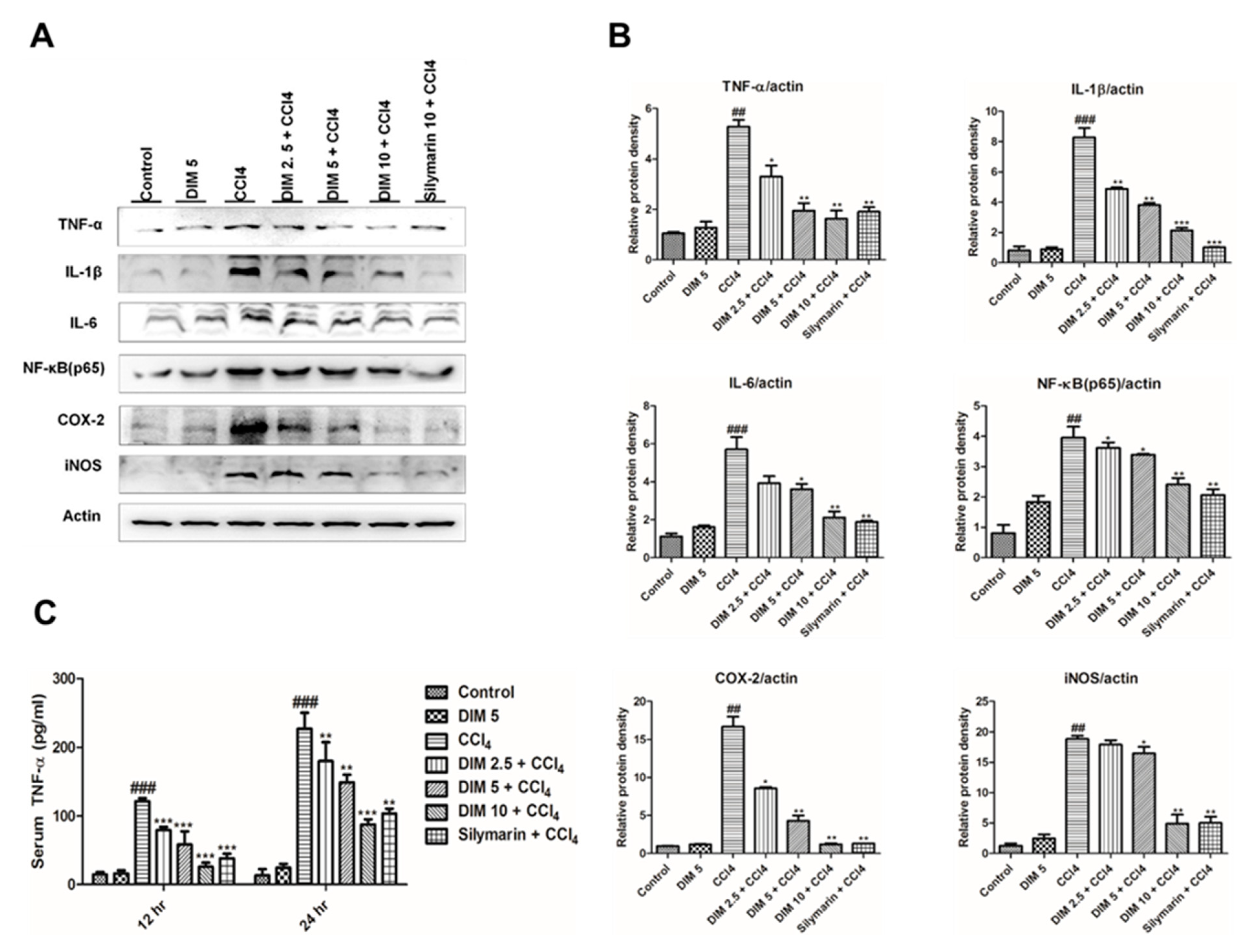
2.5. DIM Pre-Treatment Inhibits CCl4-Induced Inflammatory Mediators and Cytokines
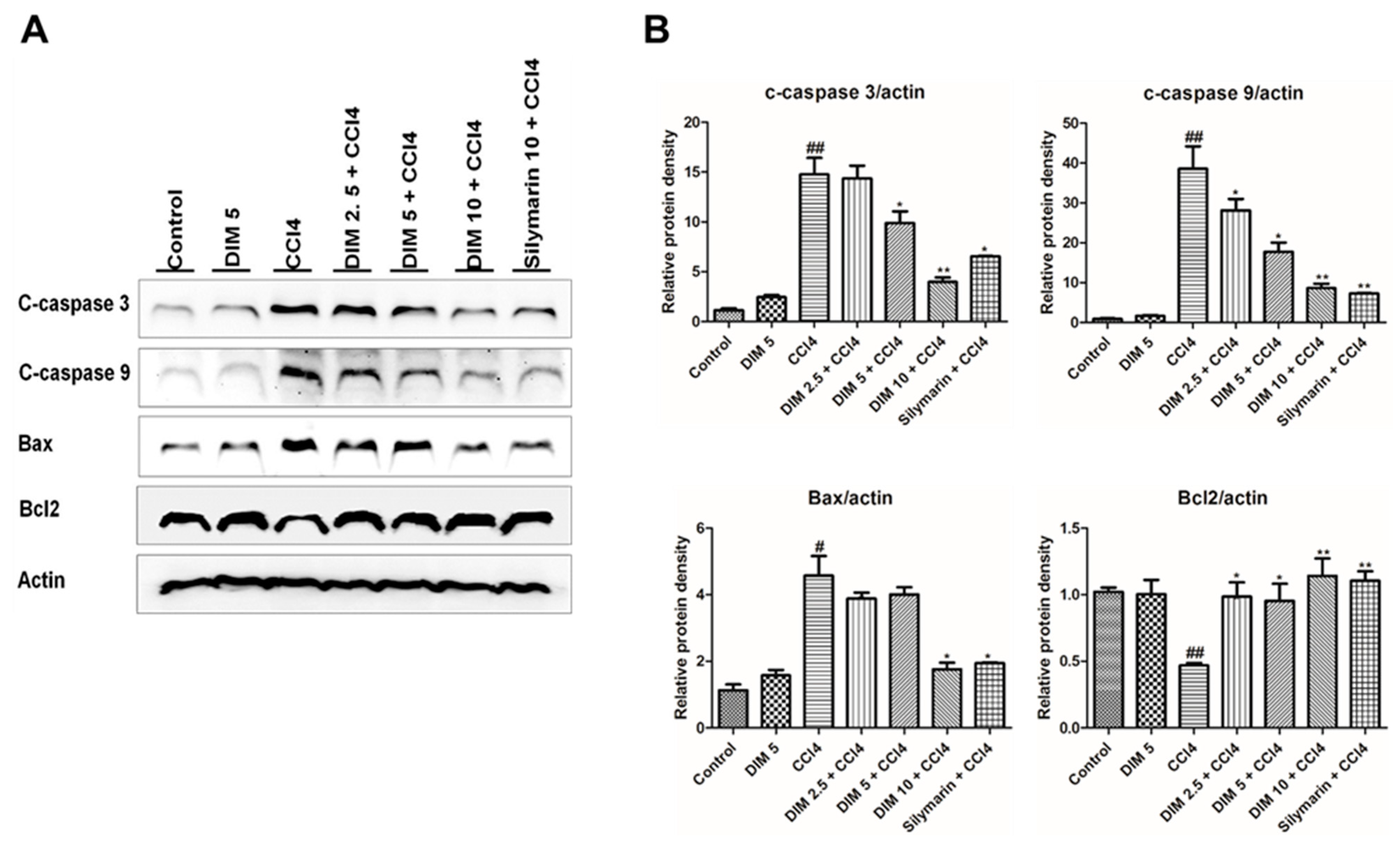
2.6. DIM Pre-Treatments Attenuates CCl4-Induced Hepatocyte Apoptosis in Mice
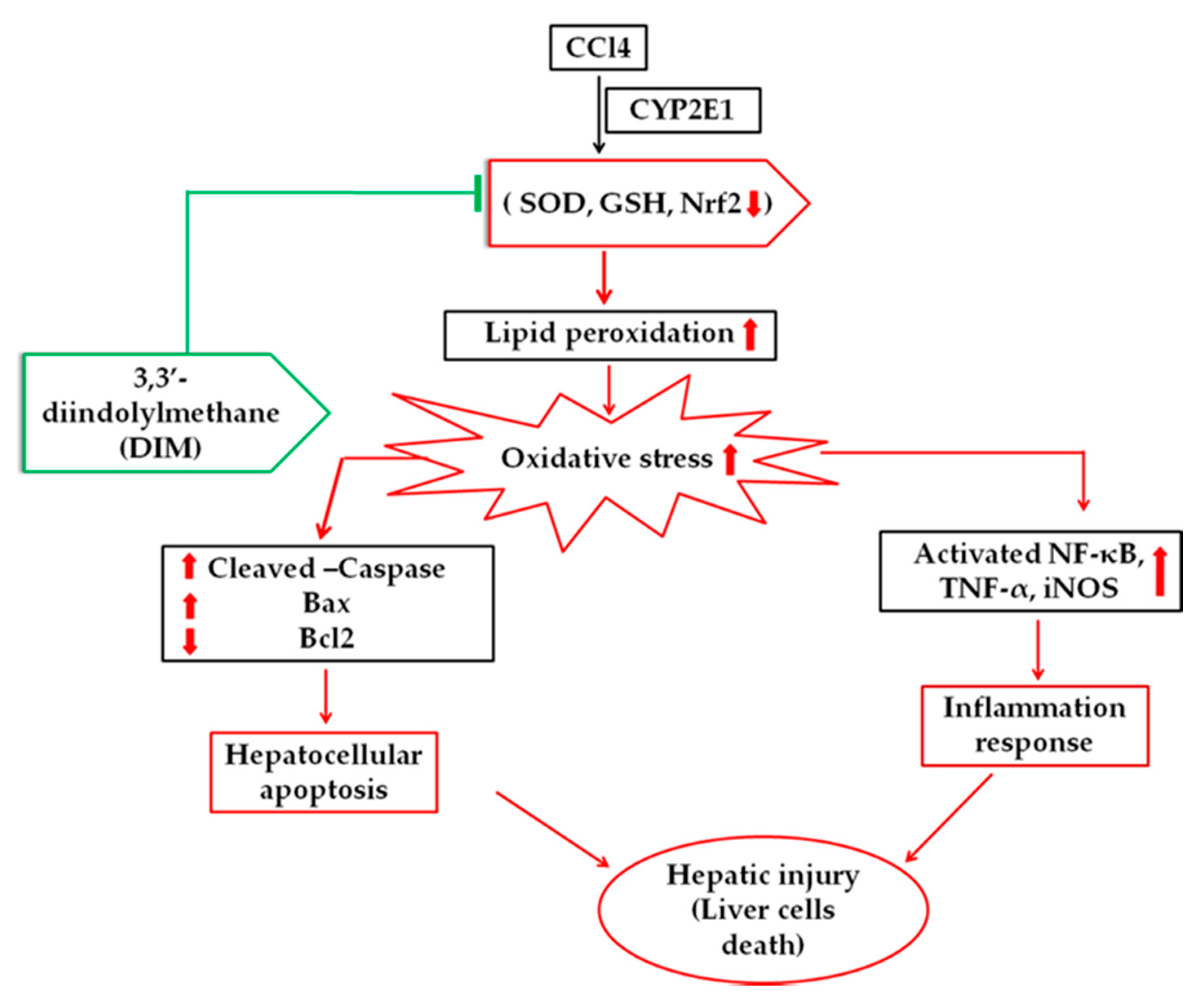
3. Discussion
4. Materials and Methods
4.1. Chemical and Reagents
4.2. Experimental Animals
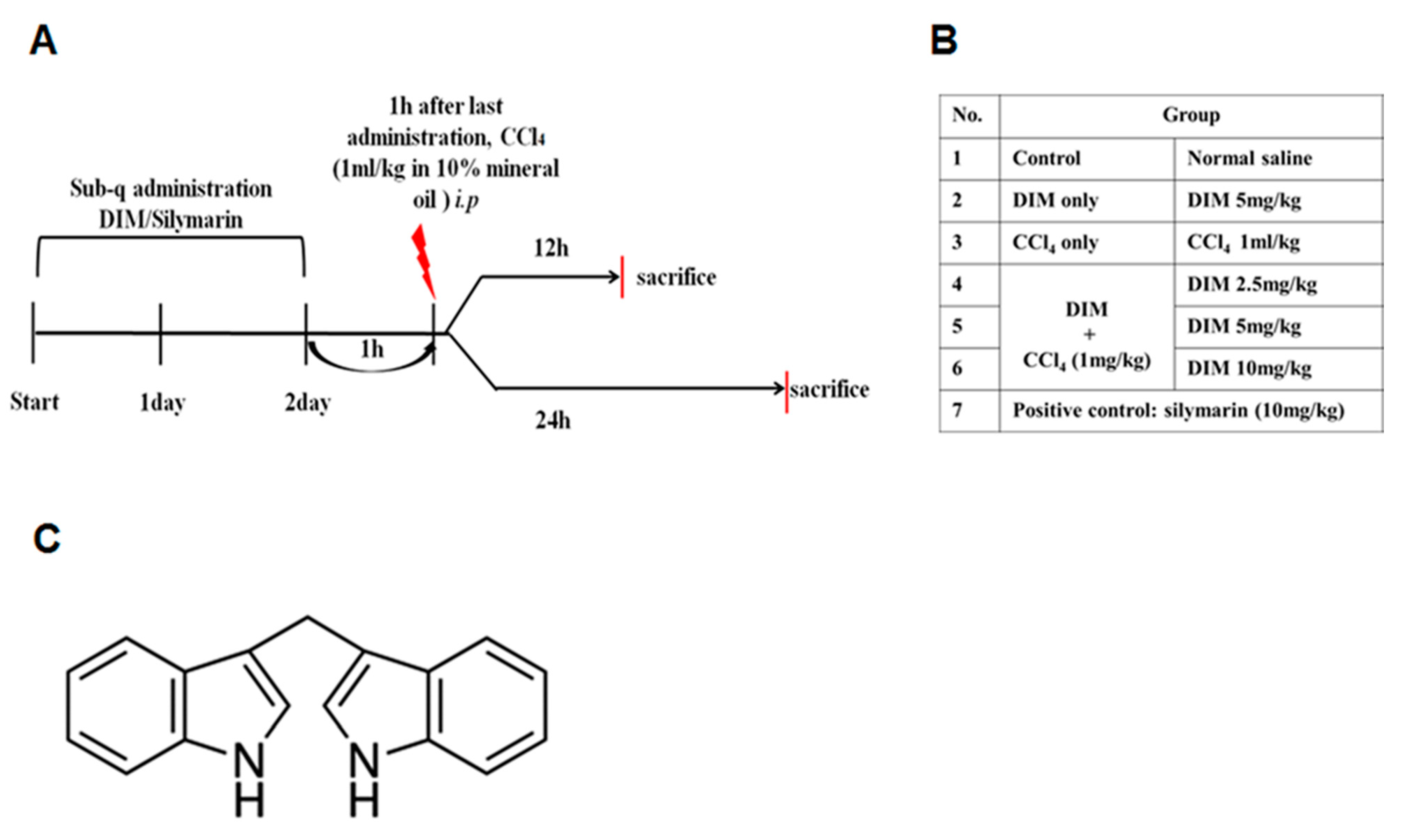
4.3. Animal Model and Treatment with DIM
4.4. Determination of Enzymatic Assay
4.5. Measurements of Reduced Glutathione (GSH) and Superoxide Dismutase (SOD) Levels in Liver Tissue
4.6. Measurements of Malondialdehyde (MDA)
4.7. Measurement of Serum TNF-α
4.8. Liver Histopathology Examination
4.9. Microscopic Detection of Reactive Oxygen Species
4.10. Western Blot Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DIM | 3,3′-Diindolylmethane |
| CCl4 | carbon tetrachloride |
| ROS | reactive oxygen species |
| TNF-α | tumor necrosis factor-α |
| CYP2E1 | cytochrome P450 2E1 |
| Nrf2 | nuclear factor erythroid 2-related factor 2 |
| NF-κB | nuclear factor kappa B |
| HO-1 | heme oxygenase |
| AST | aspartate aminotransferase |
| ALT | alanine aminotransferase |
| GSH | Glutathione |
| MDA | Malondialdehyde |
| DHE | Dihydroethidium |
| SOD | Superoxide dismutase |
| Cu/ZnSOD | Copper, zinc superoxide dismutase |
| COX-2 | Cyclooxygenase-2 |
| iNOS | Inducible nitric oxide synthase |
| IL-6 | Interleukin-6 |
| IL-1β | Interleukin-1β |
| Bcl2 | B-cell lymphoma 2 |
| Bax | Bcl2 associated X protein |
References
- Taub, R. Liver regeneration: From myth to mechanism. Nat. Rev. Mol. Cell Biol. 2004, 5, 836. [Google Scholar] [CrossRef]
- Risal, P.; Hwang, P.H.; Yun, B.S.; Yi, H.K.; Cho, B.H.; Jang, K.Y.; Jeong, Y.J. Hispidin analogue davallialactone attenuates carbon tetrachloride-induced hepatotoxicity in mice. J. Nat. Prod. 2012, 75, 1683–1689. [Google Scholar] [CrossRef] [PubMed]
- Bernal, W.; Auzinger, G.; Dhawan, A.; Wendon, J. Acute liver failure. Lancet 2010, 376, 190–201. [Google Scholar] [CrossRef]
- Zhan, Y.; Wang, Z.; Yang, P.; Wang, T.; Xia, L.; Zhou, M.; Wang, Y.; Wang, S.; Hua, Z.; Zhang, J. Adenosine 5′-monophosphate ameliorates D-galactosamine/lipopolysaccharide-induced liver injury through an adenosine receptor-independent mechanism in mice. Cell Death Dis. 2014, 5, e985. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Zhang, L.; Zhang, X.; Shi, H.; Wen, T.; Bai, L.; Zheng, S.; Chen, Y.; Chen, D.; Li, L.; et al. Inhibition of glycogen synthase kinase 3beta promotes autophagy to protect mice from acute liver failure mediated by peroxisome proliferator-activated receptor alpha. Cell Death Dis. 2016, 7, e2151. [Google Scholar] [CrossRef]
- Busbee, P.B.; Nagarkatti, M.; Nagarkatti, P.S. Natural indoles, indole-3-carbinol (I3C) and 3,3′-diindolylmethane (DIM), attenuate staphylococcal enterotoxin B-mediated liver injury by downregulating miR-31 expression and promoting caspase-2-mediated apoptosis. PLoS ONE 2015, 10, e0118506. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.-Y.; Wang, N.; Zhang, Z.-J.; Lao, L.; Wong, C.-W.; Feng, Y. The role of oxidative stress and antioxidants in liver diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef]
- Xu, D.; Xu, M.; Jeong, S.; Qian, Y.; Wu, H.; Xia, Q.; Kong, X. The Role of Nrf2 in Liver Disease: Novel Molecular Mechanisms and Therapeutic Approaches. Front. Pharmacol. 2019, 9, 1428. [Google Scholar] [CrossRef]
- Medina, J.; Moreno-Otero, R. Pathophysiological basis for antioxidant therapy in chronic liver disease. Drugs 2005, 65, 2445–2461. [Google Scholar] [CrossRef]
- Dey, A.; Lakshmanan, J. The role of antioxidants and other agents in alleviating hyperglycemia mediated oxidative stress and injury in liver. Food Funct. 2013, 4, 1148–1184. [Google Scholar] [CrossRef]
- Mallikarjuna, K.; Shanmugam, K.R.; Nishanth, K.; Wu, M.C.; Hou, C.W.; Kuo, C.H.; Reddy, K.S. Alcohol-induced deterioration in primary antioxidant and glutathione family enzymes reversed by exercise training in the liver of old rats. Alcohol 2010, 44, 523–529. [Google Scholar] [CrossRef]
- Alam, J.; Stewart, D.; Touchard, C.; Boinapally, S.; Choi, A.M.; Cook, J.L. Nrf2, a Cap’n’Collar transcription factor, regulates induction of the heme oxygenase-1 gene. J. Biol. Chem. 1999, 274, 26071–26078. [Google Scholar] [CrossRef]
- Gupte, A.A.; Lyon, C.J.; Hsueh, W.A. Nuclear factor (erythroid-derived 2)-like-2 factor (Nrf2), a key regulator of the antioxidant response to protect against atherosclerosis and nonalcoholic steatohepatitis. Curr. Diabetes Rep. 2013, 13, 362–371. [Google Scholar] [CrossRef]
- Vomund, S.; Schäfer, A.; Parnham, M.J.; Brüne, B.; von Knethen, A. Nrf2, the master regulator of anti-oxidative responses. Int. J. Mol. Sci. 2017, 18, 2772. [Google Scholar] [CrossRef]
- Chowdhry, S.; Nazmy, M.H.; Meakin, P.J.; Dinkova-Kostova, A.T.; Walsh, S.V.; Tsujita, T.; Dillon, J.F.; Ashford, M.L.J.; Hayes, J.D. Loss of Nrf2 markedly exacerbates nonalcoholic steatohepatitis. Free Radic. Biol. Med. 2010, 48, 357–371. [Google Scholar] [CrossRef]
- Ramadori, P.; Drescher, H.; Erschfeld, S.; Fragoulis, A.; Kensler, T.W.; Wruck, C.J.; Cubero, F.J.; Trautwein, C.; Streetz, K.L.; Kroy, D.C. Genetic Nrf2 overactivation inhibits the deleterious effects induced by hepatocyte-specific c-met deletion during the progression of NASH. Oxid. Med. Cell. Longev. 2017, 2017, 3420286. [Google Scholar] [CrossRef]
- Lee, C.H.; Park, S.W.; Kim, Y.S.; Kang, S.S.; Kim, J.A.; Lee, S.H.; Lee, S.M. Protective mechanism of glycyrrhizin on acute liver injury induced by carbon tetrachloride in mice. Biol. Pharm. Bull. 2007, 30, 1898–1904. [Google Scholar] [CrossRef]
- Fujii, T.; Fuchs, B.C.; Yamada, S.; Lauwers, G.Y.; Kulu, Y.; Goodwin, J.M.; Lanuti, M.; Tanabe, K.K. Mouse model of carbon tetrachloride induced liver fibrosis: Histopathological changes and expression of CD133 and epidermal growth factor. BMC Gastroenterol. 2010, 10, 79. [Google Scholar] [CrossRef]
- Edwards, M.J.; Keller, B.J.; Kauffman, F.C.; Thurman, R.G. The Involvement of Kupffer Cells in Carbon Tetrachloride Toxicity. Toxicol. Appl. Pharmacol. 1993, 119, 275–279. [Google Scholar] [CrossRef]
- Decker, K. Biologically active products of stimulated liver macrophages (Kupffer cells). Eur. J. Biochem. 1990, 192, 245–261. [Google Scholar] [CrossRef]
- Risal, P.; Park, B.H.; Cho, B.H.; Kim, J.C.; Jeong, Y.J. Overexpression of peptidyl-prolyl isomerase Pin1 attenuates hepatocytes apoptosis and secondary necrosis following carbon tetrachloride-induced acute liver injury in mice. Pathol. Int. 2012, 62, 8–15. [Google Scholar] [CrossRef]
- Lee, H.-Y.; Kim, S.-W.; Lee, G.-H.; Choi, M.-K.; Jung, H.-W.; Kim, Y.-J.; Kwon, H.-J.; Chae, H.-J. Turmeric extract and its active compound, curcumin, protect against chronic CCl4-induced liver damage by enhancing antioxidation. BMC Complement. Altern. Med. 2016, 16, 316. [Google Scholar] [CrossRef]
- McGuire, K.P.; Ngoubilly, N.; Neavyn, M.; Lanza-Jacoby, S. 3,3′-diindolylmethane and paclitaxel act synergistically to promote apoptosis in HER2/Neu human breast cancer cells. J. Surg. Res. 2006, 132, 208–213. [Google Scholar] [CrossRef]
- Luo, Q.; Yang, A.; Cao, Q.; Guan, H. 3,3′-Diindolylmethane protects cardiomyocytes from LPS-induced inflammatory response and apoptosis. BMC Pharmacol. Toxicol. 2018, 19, 71. [Google Scholar] [CrossRef]
- Tomar, S.; Nagarkatti, M.; Nagarkatti, P.S. 3,3′-Diindolylmethane attenuates LPS-mediated acute liver failure by regulating miRNAs to target IRAK4 and suppress Toll-like receptor signalling. Br. J. Pharmacol. 2015, 172, 2133–2147. [Google Scholar] [CrossRef]
- Xia, Z.-E.; Xi, J.-L.; Shi, L. 3,3′-Diindolylmethane ameliorates renal fibrosis through the inhibition of renal fibroblast activation in vivo and in vitro. Renal Fail. 2018, 40, 447–454. [Google Scholar] [CrossRef]
- Kaplowitz, N. Mechanisms of liver cell injury. J. Hepatol. 2000, 32, 39–47. [Google Scholar] [CrossRef]
- Reyes-Gordillo, K.; Segovia, J.; Shibayama, M.; Vergara, P.; Moreno, M.G.; Muriel, P. Curcumin protects against acute liver damage in the rat by inhibiting NF-kappaB, proinflammatory cytokines production and oxidative stress. Biochim. Biophys. Acta 2007, 1770, 989–996. [Google Scholar] [CrossRef]
- Cengiz, N.; Kavak, S.; Güzel, A.; Ozbek, H.; Bektaş, H.; Him, A.; Erdoğan, E.; Balahoroğlu, R. Investigation of the hepatoprotective effects of Sesame (Sesamum indicum L.) in carbon tetrachloride-induced liver toxicity. J. Membr. Biol. 2013, 246, 1–6. [Google Scholar] [CrossRef]
- Ashok, B.T.; Chen, Y.G.; Liu, X.; Garikapaty, V.P.; Seplowitz, R.; Tschorn, J.; Roy, K.; Mittelman, A.; Tiwari, R.K. Multiple molecular targets of indole-3-carbinol, a chemopreventive anti-estrogen in breast cancer. Eur. J. Cancer Prev. Off. J. Eur. Cancer Prev. Organ. ECP 2002, 11 (Suppl. 2), S86–S93. [Google Scholar]
- Tahir, M.; Sultana, S. Chrysin modulates ethanol metabolism in wistar rats: A promising role against organ toxicities. Alcohol Alcoholism 2011, 46, 383–392. [Google Scholar] [CrossRef]
- Slater, T.F.; Cheeseman, K.H.; Ingold, K.U. Carbon tetrachloride toxicity as a model for studying free-radical mediated liver injury. Philos.Trans. R. Soc. Lond. Ser. B Biol. Sci. 1985, 311, 633–645. [Google Scholar] [CrossRef]
- Wunjuntuk, K.; Kettawan, A.; Charoenkiatkul, S.; Rungruang, T. Parboiled germinated brown rice protects against CCl4-Induced oxidative stress and liver injury in rats. J. Med. Food 2016, 19, 15–23. [Google Scholar] [CrossRef]
- Al-Rasheed, N.; Faddah, L.; Al-Rasheed, N.; Bassiouni, Y.A.; Hasan, I.H.; Mahmoud, A.M.; Mohamad, R.A.; Yacoub, H.I. Protective effects of silymarin, alone or in combination with chlorogenic acid and/or melatonin, against carbon tetrachloride-induced hepatotoxicity. Pharmacogn. Mag. 2016, 12, S337–S345. [Google Scholar] [CrossRef][Green Version]
- Dai, Y.; Cederbaum, A.I. Inactivation and degradation of human cytochrome P4502E1 by CCl4 in a transfected HepG2 cell line. J. Pharmacol. Exp. Ther. 1995, 275, 1614–1622. [Google Scholar]
- Ai, G.; Liu, Q.; Hua, W.; Huang, Z.; Wang, D. Hepatoprotective evaluation of the total flavonoids extracted from flowers of Abelmoschus manihot (L.) Medic: In vitro and in vivo studies. J. Ethnopharmacol. 2013, 146, 794–802. [Google Scholar] [CrossRef]
- Zhang, W.; Dong, Z.; Chang, X.; Zhang, C.; Rong, G.; Gao, X.; Zeng, Z.; Wang, C.; Chen, Y.; Rong, Y.; et al. Protective effect of the total flavonoids from Apocynum venetum L. on carbon tetrachloride-induced hepatotoxicity in vitro and in vivo. J. Physiol. Biochem. 2018, 74, 301–312. [Google Scholar] [CrossRef]
- Plaa, G.L. Chlorinated Methanes and Liver Injury: Highlights of the Past 50 Years. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 43–65. [Google Scholar] [CrossRef]
- Ismail, A.F.M.; Salem, A.A.M.; Eassawy, M.M.T. Hepatoprotective effect of grape seed oil against carbon tetrachloride induced oxidative stress in liver of γ-irradiated rat. J. Photochem. Photobiol. B Biol. 2016, 160, 1–10. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Nordberg, J.; Arnér, E.S.J. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system1 1This review is based on the licentiate thesis “Thioredoxin reductase—Interactions with the redox active compounds 1-chloro-2,4-dinitrobenzene and lipoic acid” by Jonas Nordberg, 2001, Karolinska Institute, Stockholm, ISBN 91–631–1064–4. Free Radic. Biol. Med. 2001, 31, 1287–1312. [Google Scholar] [CrossRef]
- Nielsen, F.; Mikkelsen, B.B.; Nielsen, J.B.; Andersen, H.R.; Grandjean, P. Plasma malondialdehyde as biomarker for oxidative stress: Reference interval and effects of life-style factors. Clin. Chem. 1997, 43, 1209–1214. [Google Scholar] [CrossRef]
- Levings, D.C.; Wang, X.; Kohlhase, D.; Bell, D.A.; Slattery, M. A distinct class of antioxidant response elements is consistently activated in tumors with NRF2 mutations. Redox Biol. 2018, 19, 235–249. [Google Scholar] [CrossRef]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2:INrf2 (Keap1) signaling in oxidative stress. Free Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef]
- Ma, Q.; He, X. Molecular basis of electrophilic and oxidative defense: Promises and perils of Nrf2. Pharmacol. Rev. 2012, 64, 1055–1081. [Google Scholar] [CrossRef]
- Abdel Moneim, A.E. Indigofera oblongifolia prevents lead acetate-induced hepatotoxicity, oxidative stress, fibrosis and apoptosis in rats. PLoS ONE 2016, 11, e0158965. [Google Scholar] [CrossRef]
- Cai, Z.; Lou, Q.; Wang, F.; Li, E.; Sun, J.; Fang, H.; Xi, J.; Ju, L. N-acetylcysteine protects against liver injure induced by carbon tetrachloride via activation of the Nrf2/HO-1 pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 8655–8662. [Google Scholar]
- Ji, S.; Li, Z.; Song, W.; Wang, Y.; Liang, W.; Li, K.; Tang, S.; Wang, Q.; Qiao, X.; Zhou, D.; et al. Bioactive constituents of glycyrrhiza uralensis (Licorice): Discovery of the effective components of a traditional herbal medicine. J. Nat. Prod. 2016, 79, 281–292. [Google Scholar] [CrossRef]
- Chen, S.; Zou, L.; Li, L.; Wu, T. The protective effect of glycyrrhetinic acid on carbon tetrachloride-induced chronic liver fibrosis in mice via upregulation of Nrf2. PLoS ONE 2013, 8, e53662. [Google Scholar] [CrossRef]
- Cao, Y.-W.; Jiang, Y.; Zhang, D.-Y.; Wang, M.; Chen, W.-S.; Su, H.; Wang, Y.-T.; Wan, J.-B. Protective effects of penthorum chinense pursh against chronic ethanol-induced liver injury in mice. J. Ethnopharmacol. 2015, 161, 92–98. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, X.-J.; Feng, R.; Jiang, Y.; Zhang, D.-Y.; He, C.; Li, P.; Wan, J.-B. Hepatoprotective properties of Penthorum chinense Pursh against carbon tetrachloride-induced acute liver injury in mice. Chin. Med. 2017, 12, 32. [Google Scholar] [CrossRef]
- Sass, G.; Heinlein, S.; Agli, A.; Bang, R.; Schümann, J.; Tiegs, G. Cytokine expression in three mouse models of experimental hepatitis. Cytokine 2002, 19, 115–120. [Google Scholar] [CrossRef]
- Ning, C.; Gao, X.; Wang, C.; Huo, X.; Liu, Z.; Sun, H.; Yang, X.; Sun, P.; Ma, X.; Meng, Q.; et al. Hepatoprotective effect of ginsenoside Rg1 from Panax ginseng on carbon tetrachloride-induced acute liver injury by activating Nrf2 signaling pathway in mice. Environ. Toxicol. 2018, 33, 1050–1060. [Google Scholar] [CrossRef]
- Germoush, M.O.; Othman, S.I.; Al-Qaraawi, M.A.; Al-Harbi, H.M.; Hussein, O.E.; Al-Basher, G.; Alotaibi, M.F.; Elgebaly, H.A.; Sandhu, M.A.; Allam, A.A.; et al. Umbelliferone prevents oxidative stress, inflammation and hematological alterations, and modulates glutamate-nitric oxide-cGMP signaling in hyperammonemic rats. Biomed. Pharmacother. 2018, 102, 392–402. [Google Scholar] [CrossRef]
- Utaipan, T.; Suksamrarn, A.; Kaemchantuek, P.; Chokchaisiri, R.; Stremmel, W.; Chamulitrat, W.; Chunglok, W. Diterpenoid trigonoreidon B isolated from Trigonostemon reidioides alleviates inflammation in models of LPS-stimulated murine macrophages and inflammatory liver injury in mice. Biomed. Pharmacother. 2018, 101, 961–971. [Google Scholar] [CrossRef]
- Kiso, K.; Ueno, S.; Fukuda, M.; Ichi, I.; Kobayashi, K.; Sakai, T.; Fukui, K.; Kojo, S. The Role of Kupffer Cells in Carbon Tetrachloride Intoxication in Mice. Biol. Pharm. Bull. 2012, 35, 980–983. [Google Scholar] [CrossRef]
- Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Nitric Oxide Production and Signaling in Inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 471–479. [Google Scholar] [CrossRef]
- Shi, J.; Aisaki, K.; Ikawa, Y.; Wake, K. Evidence of hepatocyte apoptosis in rat liver after the administration of carbon tetrachloride. Am. J. Pathol. 1998, 153, 515–525. [Google Scholar] [CrossRef]
- Aram, G.; Potter, J.J.; Liu, X.; Wang, L.; Torbenson, M.S.; Mezey, E. Deficiency of nicotinamide adenine dinucleotide phosphate, reduced form oxidase enhances hepatocellular injury but attenuates fibrosis after chronic carbon tetrachloride administration. Hepatology 2009, 49, 911–919. [Google Scholar] [CrossRef]
- Guo, X.-L.; Liang, B.; Wang, X.-W.; Fan, F.-G.; Jin, J.; Lan, R.; Yang, J.-H.; Wang, X.-C.; Jin, L.; Cao, Q. Glycyrrhizic acid attenuates CCl₄-induced hepatocyte apoptosis in rats via a p53-mediated pathway. World J. Gastroenterol. 2013, 19, 3781–3791. [Google Scholar] [CrossRef]
- Czabotar, P.E.; Lessene, G.; Strasser, A.; Adams, J.M. Control of apoptosis by the BCL-2 protein family: Implications for physiology and therapy. Nat. Rev. Mol. Cell Biol. 2014, 15, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Shirali, S.; Aghaei, M.; Shabani, M.; Fathi, M.; Sohrabi, M.; Moeinifard, M. Adenosine induces cell cycle arrest and apoptosis via cyclinD1/Cdk4 and Bcl-2/Bax pathways in human ovarian cancer cell line OVCAR-3. Tumor Biol. 2013, 34, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Lin, Y.; Li, H.; Liu, J.; Sheng, X.; Zhang, W. 2,5-Hexanedione induces human ovarian granulosa cell apoptosis through BCL-2, BAX, and CASPASE-3 signaling pathways. Arch. Toxicol. 2012, 86, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Jung, W.-K.; Jeong, M.H.; Yoon, T.R.; Kim, H.K. Sanguinarine Induces Apoptosis of HT-29 Human Colon Cancer Cells via the Regulation of Bax/Bcl-2 Ratio and Caspase-9-Dependent Pathway. Int. J. Toxicol. 2012, 31, 70–77. [Google Scholar] [CrossRef]
- Spampanato, C.; De Maria, S.; Sarnataro, M.; Giordano, E.; Zanfardino, M.; Baiano, S.; Cartenì, M.; Morelli, F. Simvastatin inhibits cancer cell growth by inducing apoptosis correlated to activation of Bax and down-regulation of BCL-2 gene expression. Int. J. Oncol. 2012, 40, 935–941. [Google Scholar] [CrossRef]
- Mahmoodzadeh, Y.; Mazani, M.; Rezagholizadeh, L. Hepatoprotective effect of methanolic Tanacetum parthenium extract on CCl4-induced liver damage in rats. Toxicol. Rep. 2017, 4, 455–462. [Google Scholar] [CrossRef]
- Niu, X.; Liu, F.; Li, W.; Zhi, W.; Yao, Q.; Zhao, J.; Yang, G.; Wang, X.; Qin, L.; He, Z. Hepatoprotective effect of fraxin against carbon tetrachloride-induced hepatotoxicity in vitro and in vivo through regulating hepatic antioxidant, inflammation response and the MAPK-NF-κB signaling pathway. Biomed. Pharmacother. 2017, 95, 1091–1102. [Google Scholar] [CrossRef]
- Lehwald, N.; Tao, G.-Z.; Jang, K.Y.; Sorkin, M.; Knoefel, W.T.; Sylvester, K.G. Wnt-β-catenin signaling protects against hepatic ischemia and reperfusion injury in mice. Gastroenterology 2011, 141, 707–718.e5. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munakarmi, S.; Chand, L.; Shin, H.B.; Jang, K.Y.; Jeong, Y.J. Indole-3-Carbinol Derivative DIM Mitigates Carbon Tetrachloride-Induced Acute Liver Injury in Mice by Inhibiting Inflammatory Response, Apoptosis and Regulating Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 2048. https://doi.org/10.3390/ijms21062048
Munakarmi S, Chand L, Shin HB, Jang KY, Jeong YJ. Indole-3-Carbinol Derivative DIM Mitigates Carbon Tetrachloride-Induced Acute Liver Injury in Mice by Inhibiting Inflammatory Response, Apoptosis and Regulating Oxidative Stress. International Journal of Molecular Sciences. 2020; 21(6):2048. https://doi.org/10.3390/ijms21062048
Chicago/Turabian StyleMunakarmi, Suvesh, Lokendra Chand, Hyun Beak Shin, Kyu Yun Jang, and Yeon Jun Jeong. 2020. "Indole-3-Carbinol Derivative DIM Mitigates Carbon Tetrachloride-Induced Acute Liver Injury in Mice by Inhibiting Inflammatory Response, Apoptosis and Regulating Oxidative Stress" International Journal of Molecular Sciences 21, no. 6: 2048. https://doi.org/10.3390/ijms21062048
APA StyleMunakarmi, S., Chand, L., Shin, H. B., Jang, K. Y., & Jeong, Y. J. (2020). Indole-3-Carbinol Derivative DIM Mitigates Carbon Tetrachloride-Induced Acute Liver Injury in Mice by Inhibiting Inflammatory Response, Apoptosis and Regulating Oxidative Stress. International Journal of Molecular Sciences, 21(6), 2048. https://doi.org/10.3390/ijms21062048