Abstract
Glioblastoma (GBM) is a particularly devastating tumor with a median survival of about 16 months. Recent research has revealed novel insights into the outstanding heterogeneity of this type of brain cancer. However, all GBM subtypes share the hallmark feature of aggressive invasion into the surrounding tissue. Invasive glioblastoma cells escape surgery and focal therapies and thus represent a major obstacle for curative therapy. This review aims to provide a comprehensive understanding of glioma invasion mechanisms with respect to tumor-cell-intrinsic properties as well as cues provided by the microenvironment. We discuss genetic programs that may influence the dissemination and plasticity of GBM cells as well as their different invasion patterns. We also review how tumor cells shape their microenvironment and how, vice versa, components of the extracellular matrix and factors from non-neoplastic cells influence tumor cell motility. We further discuss different research platforms for modeling invasion. Finally, we highlight the importance of accounting for the complex interplay between tumor cell invasion and treatment resistance in glioblastoma when considering new therapeutic approaches.
1. Introduction
Glioblastoma (GBM) is the most prevalent and malignant primary brain tumor of adults, with a mean incidence of 7 cases per 100,000 per year and a mean overall survival of about 16 months [1]. GBM is a biologically heterogenous tumor that exhibits all of the classic hallmarks of cancer [2], with significant differences between patients. Radio-chemotherapy, supplemented with tumor treating fields (TTFields), is the standard of care [3,4] leading to a mean overall survival of 20.9 month [4]. Personalized therapies that target specific pathogenic events or molecular targets have not been discovered, though subtypes of GBM being prognostically relevant have been defined by genetic [5] and epigenetic [6] methods.
One of the clinical hallmarks of GBM is extensive infiltration of the tumor surrounding parenchyma [7,8]. Gliomas almost never metastasize out of the brain. Invasion happens along pre-existing structures such as blood vessels, white matter tracts and the subarachnoid space [9] and is likely coordinated by specialized cells that may lead collective invasion [10,11]. GBM cells can cross tissue barriers by remodeling their own cytoskeleton and the extracellular matrix (ECM) [9] and invade as individual cells [7] or collectively [12,13].
Reinvasion of distant GBM cells to the primary tumor location and invasion to distant areas may lead to colonization of these sites, thus triggering local and distant bulky relapse. Recurring tumors share many early tumor mutations with their progeny [14], indicating that phylogenetically primary tumor progenitor clones reside in niches from which they escape their dormant state to repopulate locally [15].
Several patterns of invasion have been described [16,17] which depend on (often conserved) genetic programs [18] and interactions with the microenvironment [10]. Invasion can be simulated with GBM brain tumor initiating cells (BTIC) [19,20] as well as differentiated cells [21,22].
Though the mechanisms leading to GBM invasion are increasingly understood in vitro and in animal models, results from the laboratory have not been translated into clinically relevant therapeutic approaches so far [23]. Within this review, we will provide an overview of the current knowledge on GBM cell invasion and provide guidance for future research in this field (Figure 1).
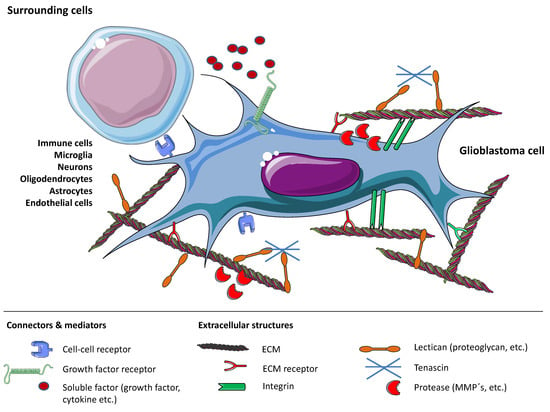
Figure 1.
Overview of mechanisms that enable glioblastoma (GBM) cell invasion by shaping the tumor microenvironment. Processes involve the glioblastoma cell and extracellular matrix (ECM) themselves, as well as connecting molecules and proteases, soluble and bound factors and surrounding cells. Figure drawn with smart.servier.com.
2. Conserved Genetic Programs to Coordinate Invasion in Glioblastoma
GBM BTIC and differentiated cells may exploit conserved genetic programs that are fundamental during neuro- and gliogenesis to migrate through the brain parenchyma [24]. These programs may determine the morphology of invading GBM cells and thereby their ability to move through tight spaces and to react to signals from other prevalent cells and extracellular fluids that are found within the microenvironment.
Migration of both neural progenitor and glioma cells is guided by specific structures, such as white matter tracts [25] and blood vessels [26,27]. Both types of cells typically exhibit a uni- or bipolar morphology that resembles the growth cones of outgrowing axons [28]. Several genes common in normal as well as in GBM progenitor cells have been described that shape cells to make them fit for invasion. One example is myosin II that plays a role in soma translocation of neural progenitor cells during brain development [29]. In concert with dynein, myosin II remodels the microtubule network in a fashion that locates centromeres and the nucleus in direction of the leading process [29]. Similarly, myosin II is also found in the leading, microtubule-rich protrusions of glioma cells, which facilitate invasion and cell-to-cell communication. [30]. Neurodevelopmental molecules necessary for neurite outgrowth and axon guidance are frequently repurposed by glioma cells. Chemo-attractants as well as -repellents show similar effects on normal as well as tumor cells. Pleiotrophin (PTN), for example, is a chemoattractant which normally promotes neurite outgrowth and neuroblast migration. It is produced by neural precursor as well as glioma cells; in the latter, PTN expression levels are inversely correlated with patient overall survival [31]. Genetic knockdown of PTN, on the other hand, strongly reduces glioma invasion [32].
3. Heterogeneity and Plasticity Within Glioblastoma
Clinical evidence suggests that some GBMs are more disseminated than others [33]. GBMs show remarkable interpatient and intratumoral heterogeneities on the genomic [34] and gene expression levels [35,36,37], leading to distinct phenotypes that may likely explain distinct behaviors. For example, mesenchymal-subtype GBM show an increased potential to invade in comparison to its neural, proneural and classical counterparts [5].
This general heterogeneity is mirrored by BTIC that possess stem cell characteristics [38,39] and are considered the most relevant cell population to drive GBM pathogenesis [40]. We and others have shown that BTIC are characterized by self-renewal, clonogenicity and differentiation potential into several lineages [41], and are able to induce tumors in animal models [42,43]. The phenotype of BTIC is primarily coined by their epigenetic state, and further modified by a number of transcription factors, chromatin regulators and associated cellular networks [44].
A number of transcription factors were identified as key regulators of transdifferentiation from an epithelial-like to a mesenchymal state (epithelial–mesenchymal transition, EMT) in a variety of different tumor types, including glioma. [36,45,46]. During EMT, tumor cells upregulate mesenchymal markers [47], lose their epithelial shape, acquire a spindle-like phenotype and thus acquire the ability to invade [48]. At suitable niches, EMT-transformed cells may convert back to an epithelial type, enabling the formation of a new tumor bulk [7,48].
Even though, in glioma, transition to a mesenchymal state emerges rather from a proneural than from an epithelial phenotype, similar factors contribute to this plasticity. Among the most prominent TFs involved in glioma EMT are ZEB1 [49,50], TWIST1 [51], SNAI1/2 [52,53], TAZ/WWTR1 [54] and the so-called “master regulators“ C/EBPβ and STAT3. The latter were shown to induce a mesenchymal gene expression signature and reprogram neural progenitors to a mesenchymal phenotype [36]. Recently, S100A4, a calcium binding protein, has been proposed as an upstream regulator of C/EBPβ and SNAI2 and as a critical mediator of mesenchymal transition in GBMs [50]. In addition, NFκB activation by TNFα was shown to induce mesenchymal differentiation of proneural BTICs via STAT3 and C/EBPβ [55].
Recent publications clearly indicate that glioma subtypes are subject to high plasticity [55,56]. Upon recurrence, a transition to the mesenchymal subtype is often observed and associated with worse overall survival [56,57]. This shift can be rapidly induced by therapeutic drugs or irradiation [58,59,60], which implies that subtype-tailored treatments might lose their activity.
To add even more complexity to the matter, it is not clear for many of the involved processes whether they are predominantly regulated at the genomic, transcriptional, proteomic or metabolomic level. Recently, single-cell profiling has become available for DNA [61], RNA [62], the epigenome [63], proteins [64] and even multiple levels in parallel [65], raising hope that the complexity of processes involved in GBM heterogeneity and plasticity will be soon understood in even more detail.
4. Bulk Vs. Single Cell Analysis
A full understanding of the highly complex GBM ecosystem that drives infiltration may be masked by bulk analysis of the whole tumor. High resolution-analyses at the single cell level may better depict relevant molecular events [66]. On the other side, such information has to be reintegrated to understand the systems-related coordination of tumor invasion.
Reflecting the method of analysis used, it is not entirely clear at this time if single leading cells or the tumor bulk coordinate invasion; several distinct patterns of invasion have been described in GBM [16,17] which relate to genetic programs [18] and factors from the microenvironment [10]. Single cell invasion—that can be amoeboid or mesenchymal [67], in the latter case with a spindle-like cell shape—is a common type of invasion in GBM and depends on a vivid turnover of cell–matrix interactions [68]. In contrast, collective invasion, the predominant pattern in solid tumors, also occurs in GBM, and separates further into multicellular streaming and collective invasion [17]. Guiding structures and chemoattractants [69] as well as a high contractility of cells [70] and close adherence within the tumor bulk may favor collective invasion.
A large number of publications have addressed either single-cell or bulk high-throughput analysis (Table 1). Based on these papers, single-cell methods may be well suited to describe -omics signatures that relate to certain patterns of invasion, whereas bulk analyses are more suited to detect specific driver genes [66,71].

Table 1.
Publications with a focus on glioma cell invasion using bulk and single cell analysis. If not otherwise stated, human tumor material was used. Tumors were harvested from the tumor core, the tumor periphery, or without a specific allocation. Single cells or cells from the tumor bulk were harvested and used for further analysis. RNASeq, RNA-Sequencing; N.d., not described.
5. Mechanisms of Invasion that Relate to GBM cells
GBM cells migrate in a highly coordinated manner [102]. Tumor cells are primarily adherent to adjacent cells, like neurons, astrocytes and endothelial cells, and to the ECM. Adherence is modulated by a variety of different proteins, for example, integrins or cadherins [103]. Integrins are sensitive sensors to the microenvironment and allow tumor cells to modify their attachment behavior [104]. They recruit adaptor molecules and proteins that relay signals into cells, e.g., phosphorylation and dephosphorylation events via the focal adhesion kinase (FAK) [105]. The ECM is shaped by proteolytic factors derived from GBM cells, such as matrix metalloproteinases [106,107], ADAMS [108] and cathepsins [109]. Matrix metalloproteinases (e.g., MMP-2 and MMP-9), enable GBM cells to detach from the microenvironment and to alleviate tension within the ECM [9]. While migrating, tumor cells adapt the composition of their cell membranes [110] and contract, with myosin II being of particular importance if migration happens through tight brain parenchymal spaces [111,112].
GBM cells also attract cells such as microglia, astrocytes and endothelial cells that secrete proteases to enhance migration [113]. By moving along blood vessels, GBM tumor cells degrade astrocyte connections to the basement membrane and thereby change the homeostasis in proximity of the blood vessels [114]. The focus of attention has shifted increasingly to communication between GBM cells and other cells in their microenvironment after so-called tumor microtubes have been detected recently that allow precise communication in between tumor cells and their neighbors [30,115].
Invasive programs are coordinated by overexpression of transcription factors like STAT3 and C/EBPβ, which have been shown to be synergistic initiators and master regulators of mesenchymal transformation in BTICs [36]. NF-κB may enhance and c-Myc may decrease the invasive activity in the migratory cell population [94]. Accordingly, promoter analysis of genes upregulated in invasion-leading cells revealed binding sites for NF-κB and C/EBPβ [116]. Additional programs that may influence invasion have also been described [45,46].
6. Mechanisms of Invasion that Relate to the GBM Microenvironment
GBM invasion results from a continuous bidirectional interaction between tumor cells and their microenvironment [117]. The microenvironment consists of several ECM components, fluids, soluble factors such as chemokines and cytokines, and cells such as neurons, astrocytes, oligodendrocytes, endothelial cells and immune cells [118].
6.1. Extracellular Matrix
ECM molecules provide structural support and act as a guiding scaffold or barrier [21,119]. The brain parenchyma, in contrast to other ECMs, lacks rigid components such as collagens, fibrinogen and laminin, and instead consists of proteoglycans [120], hyaluronic acid [121] and tenascin-C [120].
We and others have shown that the ECM itself, as well as cell–cell and cell–ECM connecting molecules and proteases, have an impact on glioma cell migration [122,123,124,125]. Soluble factors such as growth factors [126] also guide invasion and contribute to the fate of BTICs and differentiated tumor cells [127]. Several chemoattractants such as bradykinin [128] and vascular endothelial growth factor (VEGF) [129] play relevant roles in this context.
6.2. Adapted Niches within the Brain Parenchyma
The composition of the ECM generates specific stem cell niches and facilitates migration and invasion in distinct ways [130,131].
The most prominent example of a niche within the diseased brain is the hypoxic niche. A hypoxic environment stimulates the expression of the hypoxia-inducible factors (HIFs) HIF1α and HIF2α that favor an invasive phenotype [132]. An acidic environment also stimulates HIF function [133] and activates MMPs [134], therefore inducing similar mechanisms.
Another example of a brain-intrinsic niche that may enhance invasion is the angiogenic niche. Endothelial cells interact closely with BTICs and secrete factors that maintain these cells in a stem-cell-like state [127] and enable invasion [135].
6.3. Non-neoplastic Cells
Amongst the nonimmunogenic cell types that are prevalent in the brain, endothelial cells are important inducers of invasion, especially in the angiogenic niche. The activation of angiopoietin 1 and its crosstalk with Tie2 increases invasion [136].
The most important immunogenic cell type able to promote invasion in GBM are microglia. GBM progenitor cells closely interact with microglia [137] and thereby enable invasion, e.g., by induction of MMPs [138], induction of growth factors and release of ECM components [139]. Several proinvasive signaling pathways such as Pyk2 [140], osteopontin-cd44 [141], EGFR [142] and TGF-β [143] are also induced by the interaction of GBM with microglia.
7. In Vitro Models to Investigate Invasion in Glioblastoma
In vivo animal models allow the study of GMB invasion within the microenvironment of a living host. However, they are often complex, expensive, time consuming, impaired by a low reproducibility and associated with difficulty in dissecting the different components involved in the process.
Therefore, a number of in vitro models have been developed in recent years [144]. These models are constructed to mirror features of the human disease such as widespread dissemination of tumor cells and should cover sufficient timeframes to allow population of distant sites and repopulation of the primary tumor area [25]. In addition, they are autologous in the best case and feature not only BTICs and differentiated tumor cells, but also their microenvironment.
7.1. Two-Dimensional Models
Two-dimensional models are based on monolayers of GBM tumor cells on surfaces that are coated by, for example, ECM molecules. These models are simple to use but have important limitations. Cells in monolayers do not invade (but migrate) and are separated from their natural microenvironment, therefore behaving differently [145]. Transwell assays allow a higher level of complexity but often produce a wide range of results with high standard deviations and have to be interpreted with caution [146]. In summary, no convincing two-dimensional model has been developed for live-imaging of invading human BTICs so far [147].
7.2. Three-Dimensional Models
Three-dimensional models can be used to investigate intraparenchymal [114] and perivascular invasion [148]. Typically, collagen-based 3D assays are used, however these often lack essential brain ECM elements such as laminin, hyaluronic acid, proteoglycans and fibrous glycoproteins [149,150].
Tumor spheroids on the other hand are generated as organotypic multicellular spheroids that are obtained by culturing resected fragments of tumors [151]. They retain part of the original morphology [152] and reflect intratumoral heterogeneity [153]. Tumor organoids are another recently developed model to culture tumor cells within matrigel for up to several months. They also consist of heterogenous areas and mimic tumor development and gradients [154,155].
As a three-dimensional ex vivo model, organotypic slice cultures (OBSC) have been developed to allow long-term imaging of cancer cell migration and invasion [156,157,158,159] and to describe specific pathogenic events [160]. In addition, the OBSC-model allows the monitoring of tumor cell invasion in real time. OBSCs fill the gap between 2D cell cultures and in vivo studies and reduce the number of animals needed and the burden on them [159]. The model also allows the discrimination of events into distinct molecular and cellular levels [161] and to evaluate therapeutic interventions in situ. Functional cDNA libraries can be obtained from as few as a single cell extracted from OBSCs [11,162]. Immunohistochemical (IHC) stainings of OBSCs after as long as 21 days in situ allow to monitor relevant environmental factors [163].
The combination of an orthotopic GBM model and time-lapse in vivo microscopy of human brain tumor initiating cells through a cranial window allows in-depth analysis of tumor cell infiltration to acquire a better insight of GBM tumor cells invasion dynamics within their environment [20]. Mice can be imaged repeatedly [164], and images can be processed in a way that allows the description of features such as movement direction, persistence of direction, speed and velocity [20]. This model, specifically if allograft tumor cells are used, is close to nature, but is technically complex and puts a high burden on animals.
8. Emerging Novel Technologies and Approaches for Studying Invasive GBM
Models of tumor progression offer the possibility to predict the behavior of tumor invasiveness and, consequently, to define more accurate prognostic estimates and therapeutic strategies. In the last 20 years, several in silico models have been developed to elucidate the main mechanisms that induce glioma proliferation and invasion [165]. Although useful to investigate specific aspects of glioma invasion, many of these models have been derived from in vitro experiments, therefore limiting their use in clinical settings.
The advent of high-throughput data generation, largely coincident with the development of next-generation sequencing, has boosted the knowledge of many cancers—including gliomas—allowing computational modeling of large datasets to assess and predict tumor dynamics. For example, the Cancer Genome Atlas (TGCA) and Repository for Molecular BRAin Neoplasia DaTa (REMBRANDT) are two large-scale efforts containing genomics, transcriptomics, DNA methylomics and proteomics data as well as clinical data [166,167]. Such high dimensional data have been used successfully to cluster gliomas in different groups and to identify known molecular characteristics commonly used in clinical evaluation [168].
Recently, the Ivy Glioblastoma Atlas Project (Ivy-GAP) has generated a transcriptional atlas for known anatomical features of GBM [169]. Starting from the Ivy-GAP spatial information and data derived from genome and transcriptome sequencing, it has been possible to generate computational models that are able to explain the dynamics of GBM [170]. It has thereby been shown that GBM follows a divergent evolution with substantial genomic differences between initial versus recurrent tumors, with the latter presenting more comprehensive and extensive alterations in, e.g., RTK-MAPK, RTK-PI3K, INK4-Rb and ARF-p53 pathways [14]. Similar, evolutionary gene expression patterns can lead to a more invasive phenotype, e.g., by activation of TGFβ by overexpression of LTBP4 [56]. These patterns can by even more dissociated by dissecting infiltrative GBM cells at a single cell level, rendering possible the identification of genes involved in the invasion of the interstitial matrix [87].
To understand infiltrating single cells on a deeper level, novel targeted approaches for single cell sequencing have been developed. For example, topographic single cell sequencing (TSCS) is a method that combines laser microdissection and cell catapulting [171] with single cell DNA sequencing, while preserving spatial information. The application of TSCS in mammary ductal carcinoma in situ (DCIS) has highlighted that initially rare invading clones shift the mutation frequencies of specific genes to better escape the basement membrane and establish an invasive tumor mass [172]. TSCS, if applied to GBM, could therefore be useful in the near future to more precisely study the gene signature of single infiltrating cells and to identify mechanisms of GBM invasion and resistance, as well as to pave the way for new drugs capable of targeting these cells. Information derived from these data could also suggest combinatorial therapeutic approaches by targeting multiple cell subtypes according to their molecular characteristics.
Although DNA-seq and RNA-seq are useful to unravel cancer heterogeneity, clonal evolution and mechanisms of invasiveness in gliomas, they do not provide insight into cell-to-cell communications and interactions with the tumor microenvironment. Recently, spatial transcriptomics has therefore also interrogated cell-to-cell interactions in their natural context, revealing an unexplored landscape of heterogeneity in different types of cancers [173,174,175]. For example, spatial transcriptomics signatures in breast cancer have supported clinical diagnostics, offering the possibility to distinguish DCIS and invasive ductal carcinoma (IDC) [173], or to identify specific areas where certain transcripts (e.g., TMSB10) can predict an invasive phenotype [176]. An application of these techniques to GBM might therefore be of great relevance to explore tissue areas that surround the tumor core. Moreover, a comparison of spatial transcriptomics data over time in individual patients could give hints at which stage of the disease the invasive phenotype occurred, if known models of tumor evolution, e.g., branched or linear, are applied.
Spatial transcriptomics may also unravel expression patterns in the inflammatory environment of cancers. Recently, a dissection of prostate cancer tissue has shown that reactive stroma and inflammation are present at early stages and promote tumor growth by activation of oxidative stress and integrin-linked kinase (ILK) signaling [174]. Similarly, gene expression gradients have been observed in malignant melanoma, where it has been shown that transcriptional programs in the cancer core overlap with surrounding lymphoid tissues [175]. Lastly, the application of spatial transcriptomics to synovial biopsies from patients with rheumatoid arthritis and spondyloarthritis has allowed to decipher inflammatory signatures and to predict immune cell patterns [177]. In GBM, heterogeneous populations of tumor-associated macrophages (TAM), regulatory T-cells and dendritic cells with immunosuppressive capacity have been recently described [178]. TAM thereby support tumor cell migration by producing a wide array of cytokines and growth factors in response to signals derived from glioma cells [179]. Nevertheless, it is still not clear which exact molecular mechanisms are at the basis of this interaction and if microglia and TAM acquire new phenotypes to support the invasion of glioma cells. The mentioned interplays could be better characterized if the cells would be analyzed in their spatial context.
A bioinformatic integration of the mentioned scRNA-Seq and spatial transcriptomics data has been recently developed and applied to infer the tissue architecture in nonglioma tumors, for example in pancreatic ductal adenocarcinoma [180]. Such integrated approaches have also allowed to reveal the cellular and spatial organization of bone marrow niches at an unprecedented resolution [181]. By integrating cell-type markers derived from scRNA-seq data, it is possible to determine relative cell-type proportions in spatial transcriptomics data and, possibly, to identify rare tumor cell types and patient-specific pathogenic factors. It is noteworthy that with improved resolution at single cell levels such approaches could also be applied in GBM to identify single-cell relationships in the tumor microenvironment to pave the way for more individualized therapy decisions.
In the clinical setting, high-quality MRI data are nowadays transformed into radiomics, a method of use for building predictive models for diagnosis, prognosis and therapeutic response [182,183]. Radiogenomics—an advanced field of radiomics—aims to associate and overlay clinical and molecular features with imaging data in an attempt to describe intratumor heterogeneity [184,185]. As an example, overlapping of miRNA data derived from the TCGA with a MRI technique called fluid-attenuated inversion recovery (MRI-FLAIR) has shown concordance between peritumoral FLAIR intensity and downregulation of miRNA regulating genes involved in cellular migration and invasion, such as POSTN and CXCL12 [186]. In addition, in patients with deep white matter track involvement and ependymal invasion, it has been observed that MYC overexpression and inhibition of NF-KB inhibitor-alpha (NFKBIA) support an invasive imaging signature [187]. Similarly, the correlation of imaging from nonenhancing parenchyma with the spatially matched genetic status has shown an association with copy number aberration (CNA) changes in driver genes of GBM, such as PDGFRA, PTEN, TP53, RB1 and EGFR [188]. Further advances in radiogenomics could be of great help in the near future, especially in the follow-up phase of GBM when repeated biopsies or extensive genomic and transcriptomic analysis are not possible. In addition, 3D models that simulate response to radiotherapy have also been developed and validated in small number of patients, based on modern imaging techniques [189].
In summary, these multidimensional approaches may enable much better models and concepts for the clinical behavior of invasive GBM, allow to further fine-tune the landscape of an individual’s GBM and support the development of more precisely targeted and therefore personalized therapies.
9. Overcoming Treatment Resistance by Targeting Invasion in GBM
Molecular mechanisms of invasion and drug resistance overlap to a high extent and are influenced by cell-intrinsic and microenvironmental factors. An invasive tumor phenotype may therefore also increase resistance to antitumor systemic therapy and vice versa. Glioma cells stimulated to migrate showed downregulation of proliferation- and apoptosis-associated genes while genes involved in cell motility became overexpressed [190]. Similar results were shown by Molina et al., who found elevated MAPK signaling in the tumor core, corresponding to a proliferative state. Conversely, invading glioma cells downregulated MAPK and upregulated AKT, thereby activating prosurvival signaling cascades [191].
In general, members of pivotal signaling pathways such as p53, the Ras/MAPK, PI3K and mTOR pathways, growth factors, chemokines and integrins are hubs of mechanisms that jointly support tumor cell survival and resistance as well as migration and invasion. One prominent example is the tumor-suppressive transcription factor (TF) p53, which is altered in cancer, including glioma, via loss- or gain-of-function mutations. In both cases, apoptosis and cell cycle arrest are impaired via, e.g., downregulation of proapoptotic molecules like Bax and lack of p21 activation, respectively [192,193]. Concomitantly, epithelial-to-mesenchymal transition (EMT) is induced by upregulation of TFs Twist and Slug [194]. Another important example of such a pathway is the binding of integrins to ECM components that leads to downstream activation of GSK3, PI3K/AKT, STAT3 and FAK, resulting in enhanced survival [195,196] as well as chemo- and radio-resistance [196,197,198]. Cytoskeletal rearrangements are also induced via FAK, leading to altered ECM adhesion and motility [199]. There are many additional pathways and factors associated with therapy resistance and tumor cell motility, all of which highly relevant for both the induction of resistance and invasion. Most of them are summarized in recent excellent reviews [119,200].
In the future, it will be highly important to precisely define focal points that influence both invasion and resistance to therapy to select high priority candidates for a targeted drug intervention in patients with GBM.
10. Conclusions
Malignant gliomas are, amongst other pathogenetic features, characterized by a unique ability to invade. From a clinical point of view, invasion renders gliomas microscopically nonresectable and presents a major obstacle for curative treatment. The immense interpatient and intratumoral heterogeneity of gliomas, as well as the pivotal role of the tumor microenvironment that makes a profound understanding of the involved mechanisms difficult, are increasingly appreciated in this context. Even so, research on the complex interplay between tumor cells of different subtypes and the surrounding brain parenchyma is still at its infancy. Current standard therapies are unable to target infiltrative tumor cells, and effective anti-invasive treatment strategies are not available so far. Therefore, there is urgent clinical need to investigate the mechanisms leading to invasion of glioma in more detail. Sophisticated sequencing approaches and advancing new in vitro and in vivo models will hopefully provide a better understanding of these interactions and soon present new targets for anti-invasive therapeutic strategies.
Funding
This research received no external funding.
Acknowledgments
We thank the Wilhelm Sander-Stiftung, Munich, Germany (grants 2009.800.1 and .2) for financial support to P.H.
Conflicts of Interest
V.L. is by now a paid employee of Novocure. All other authors declare no conflict of interest. The founding sponsors had no role in the design of the manuscript, interpretation of data, writing of the manuscript, and in the decision to publish the results.
Abbreviations
| BTIC | Brain tumor initiating cells |
| GBM | Glioblastoma |
| ECM | Extracellular matrix |
| EMT | Epithelial-mesenchymal transition |
| OBSC | Organotypic brain slice culture |
References
- Davis, F.G.; Freels, S.; Grutsch, J.; Barlas, S.; Brem, S. Survival rates in patients with primary malignant brain tumors stratified by patient age and tumor histological type: An analysis based on Surveillance, Epidemiology, and End Results (SEER) data, 1973-1991. J. Neurosurg. 1998, 88, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Hegi, M.E.; Mason, W.P.; Van den Bent, M.J.; Taphoorn, M.J.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Fisher, B.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef]
- Stupp, R.; Taillibert, S.; Kanner, A.; Read, W.; Steinberg, D.; Lhermitte, B.; Toms, S.; Idbaih, A.; Ahluwalia, M.S.; Fink, K.; et al. Effect of Tumor-Treating Fields Plus Maintenance Temozolomide vs Maintenance Temozolomide Alone on Survival in Patients With Glioblastoma: A Randomized Clinical Trial. JAMA 2017, 318, 2306–2316. [Google Scholar] [CrossRef]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef]
- Noushmehr, H.; Weisenberger, D.J.; Diefes, K.; Phillips, H.S.; Pujara, K.; Berman, B.P.; Pan, F.; Pelloski, C.E.; Sulman, E.P.; Bhat, K.P.; et al. Identification of a CpG island methylator phenotype that defines a distinct subgroup of glioma. Cancer Cell 2010, 17, 510–522. [Google Scholar] [CrossRef]
- Claes, A.; Idema, A.J.; Wesseling, P. Diffuse glioma growth: A guerilla war. Acta Neuropathol. 2007, 114, 443–458. [Google Scholar] [CrossRef]
- Li, C.; Wang, S.; Yan, J.L.; Torheim, T.; Boonzaier, N.R.; Sinha, R.; Matys, T.; Markowetz, F.; Price, S.J. Characterizing tumor invasiveness of glioblastoma using multiparametric magnetic resonance imaging. J. Neurosurg. 2019, 1–8. [Google Scholar] [CrossRef]
- Cuddapah, V.A.; Robel, S.; Watkins, S.; Sontheimer, H. A neurocentric perspective on glioma invasion. Nat. Rev. Neurosci. 2014, 15, 455–465. [Google Scholar] [CrossRef]
- Vitorino, P.; Meyer, T. Modular control of endothelial sheet migration. Genes Dev. 2008, 22, 3268–3281. [Google Scholar] [CrossRef]
- Hartmann, C.H.; Klein, C.A. Gene expression profiling of single cells on large-scale oligonucleotide arrays. Nucleic Acids Res. 2006, 34, e143. [Google Scholar] [CrossRef] [PubMed]
- Friedl, P.; Wolf, K. Tumour-cell invasion and migration: Diversity and escape mechanisms. Nat. Rev. Cancer 2003, 3, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K.; Friedl, P. Molecular mechanisms of cancer cell invasion and plasticity. Br. J. Dermatol. 2006, 154 Suppl 1, 11–15. [Google Scholar] [CrossRef]
- Kim, J.; Lee, I.H.; Cho, H.J.; Park, C.K.; Jung, Y.S.; Kim, Y.; Nam, S.H.; Kim, B.S.; Johnson, M.D.; Kong, D.S.; et al. Spatiotemporal Evolution of the Primary Glioblastoma Genome. Cancer Cell 2015, 28, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.C.; Veeravagu, A.; Hsu, A.R.; Tse, V.C. Recurrent glioblastoma multiforme: A review of natural history and management options. Neurosurg Focus 2006, 20, E5. [Google Scholar] [CrossRef] [PubMed]
- Friedl, P.; Gilmour, D. Collective cell migration in morphogenesis, regeneration and cancer. Nat. Rev. Mol. Cell Biol. 2009, 10, 445–457. [Google Scholar] [CrossRef]
- Friedl, P.; Locker, J.; Sahai, E.; Segall, J.E. Classifying collective cancer cell invasion. Nat. Cell Biol. 2012, 14, 777–783. [Google Scholar] [CrossRef]
- Cheung, K.J.; Gabrielson, E.; Werb, Z.; Ewald, A.J. Collective invasion in breast cancer requires a conserved basal epithelial program. Cell 2013, 155, 1639–1651. [Google Scholar] [CrossRef]
- Sanai, N.; Alvarez-Buylla, A.; Berger, M.S. Neural stem cells and the origin of gliomas. N. Engl. J. Med. 2005, 353, 811–822. [Google Scholar] [CrossRef]
- Alieva, M.; Leidgens, V.; Riemenschneider, M.J.; Klein, C.A.; Hau, P.; Van Rheenen, J. Intravital imaging of glioma border morphology reveals distinctive cellular dynamics and contribution to tumor cell invasion. Sci. Rep. 2019, 9, 2054. [Google Scholar] [CrossRef]
- Friedl, P.; Alexander, S. Cancer invasion and the microenvironment: Plasticity and reciprocity. Cell 2011, 147, 992–1009. [Google Scholar] [CrossRef] [PubMed]
- Nevo, I.; Woolard, K.; Cam, M.; Li, A.; Webster, J.D.; Kotliarov, Y.; Kim, H.S.; Ahn, S.; Walling, J.; Kotliarova, S.; et al. Identification of molecular pathways facilitating glioma cell invasion in situ. PLoS ONE 2014, 9, e111783. [Google Scholar] [CrossRef]
- Perrin, S.L.; Samuel, M.S.; Koszyca, B.; Brown, M.P.; Ebert, L.M.; Oksdath, M.; Gomez, G.A. Glioblastoma heterogeneity and the tumour microenvironment: Implications for preclinical research and development of new treatments. Biochem. Soc. Trans. 2019, 47, 625–638. [Google Scholar] [CrossRef] [PubMed]
- Ming, G.L.; Song, H. Adult neurogenesis in the mammalian brain: Significant answers and significant questions. Neuron 2011, 70, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Sahm, F.; Capper, D.; Jeibmann, A.; Habel, A.; Paulus, W.; Troost, D.; Von Deimling, A. Addressing diffuse glioma as a systemic brain disease with single-cell analysis. Arch. Neurol. 2012, 69, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Le Magueresse, C.; Alfonso, J.; Bark, C.; Eliava, M.; Khrulev, S.; Monyer, H. Subventricular zone-derived neuroblasts use vasculature as a scaffold to migrate radially to the cortex in neonatal mice. Cereb. Cortex 2012, 22, 2285–2296. [Google Scholar] [CrossRef] [PubMed]
- Winkler, F.; Kienast, Y.; Fuhrmann, M.; Von Baumgarten, L.; Burgold, S.; Mitteregger, G.; Kretzschmar, H.; Herms, J. Imaging glioma cell invasion in vivo reveals mechanisms of dissemination and peritumoral angiogenesis. Glia 2009, 57, 1306–1315. [Google Scholar] [CrossRef]
- Lowery, L.A.; Van Vactor, D. The trip of the tip: Understanding the growth cone machinery. Nat. Rev. Mol. Cell Biol. 2009, 10, 332–343. [Google Scholar] [CrossRef]
- Tsai, J.W.; Bremner, K.H.; Vallee, R.B. Dual subcellular roles for LIS1 and dynein in radial neuronal migration in live brain tissue. Nat. Neurosci. 2007, 10, 970–979. [Google Scholar] [CrossRef]
- Osswald, M.; Jung, E.; Sahm, F.; Solecki, G.; Venkataramani, V.; Blaes, J.; Weil, S.; Horstmann, H.; Wiestler, B.; Syed, M.; et al. Brain tumour cells interconnect to a functional and resistant network. Nature 2015, 528, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Kundu, S.; Feenstra, T.; Li, X.; Jin, C.; Laaniste, L.; El Hassan, T.E.; Ohlin, K.E.; Yu, D.; Olofsson, T.; et al. Pleiotrophin promotes vascular abnormalization in gliomas and correlates with poor survival in patients with astrocytomas. Sci. Signal. 2015, 8, ra125. [Google Scholar] [CrossRef] [PubMed]
- Qin, E.Y.; Cooper, D.D.; Abbott, K.L.; Lennon, J.; Nagaraja, S.; Mackay, A.; Jones, C.; Vogel, H.; Jackson, P.K.; Monje, M. Neural Precursor-Derived Pleiotrophin Mediates Subventricular Zone Invasion by Glioma. Cell 2017, 170, 845–859. [Google Scholar] [CrossRef]
- Chamberlain, M.C. Radiographic patterns of relapse in glioblastoma. J. Neurooncol. 2011, 101, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Sottoriva, A.; Spiteri, I.; Piccirillo, S.G.; Touloumis, A.; Collins, V.P.; Marioni, J.C.; Curtis, C.; Watts, C.; Tavare, S. Intratumor heterogeneity in human glioblastoma reflects cancer evolutionary dynamics. Proc. Natl. Acad. Sci. USA 2013, 110, 4009–4014. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.P.; Tirosh, I.; Trombetta, J.J.; Shalek, A.K.; Gillespie, S.M.; Wakimoto, H.; Cahill, D.P.; Nahed, B.V.; Curry, W.T.; Martuza, R.L.; et al. Single-cell RNA-seq highlights intratumoral heterogeneity in primary glioblastoma. Science 2014, 344, 1396–1401. [Google Scholar] [CrossRef] [PubMed]
- Carro, M.S.; Lim, W.K.; Alvarez, M.J.; Bollo, R.J.; Zhao, X.; Snyder, E.Y.; Sulman, E.P.; Anne, S.L.; Doetsch, F.; Colman, H.; et al. The transcriptional network for mesenchymal transformation of brain tumours. Nature 2010, 463, 318–325. [Google Scholar] [CrossRef]
- Galli, R.; Binda, E.; Orfanelli, U.; Cipelletti, B.; Gritti, A.; De Vitis, S.; Fiocco, R.; Foroni, C.; Dimeco, F.; Vescovi, A. Isolation and characterization of tumorigenic, stem-like neural precursors from human glioblastoma. Cancer Res. 2004, 64, 7011–7021. [Google Scholar] [CrossRef]
- Singh, S.K.; Clarke, I.D.; Terasaki, M.; Bonn, V.E.; Hawkins, C.; Squire, J.; Dirks, P.B. Identification of a cancer stem cell in human brain tumors. Cancer Res. 2003, 63, 5821–5828. [Google Scholar]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.A.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of human brain tumour initiating cells. Nature 2004, 432, 396–401. [Google Scholar] [CrossRef]
- Leidgens, V.; Proske, J.; Rauer, L.; Moeckel, S.; Renner, K.; Bogdahn, U.; Riemenschneider, M.J.; Proescholdt, M.; Vollmann-Zwerenz, A.; Hau, P.; et al. Stattic and metformin inhibit brain tumor initiating cells by reducing STAT3-phosphorylation. Oncotarget 2017, 8, 8250–8263. [Google Scholar] [CrossRef]
- Beier, D.; Hau, P.; Proescholdt, M.; Lohmeier, A.; Wischhusen, J.; Oefner, P.J.; Aigner, L.; Brawanski, A.; Bogdahn, U.; Beier, C.P. CD133(+) and CD133(−) glioblastoma-derived cancer stem cells show differential growth characteristics and molecular profiles. Cancer Res. 2007, 67, 4010–4015. [Google Scholar] [CrossRef] [PubMed]
- Beier, D.; Rohrl, S.; Pillai, D.R.; Schwarz, S.; Kunz-Schughart, L.A.; Leukel, P.; Proescholdt, M.; Brawanski, A.; Bogdahn, U.; Trampe-Kieslich, A.; et al. Temozolomide preferentially depletes cancer stem cells in glioblastoma. Cancer Res. 2008, 68, 5706–5715. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Srikanth, M.; Kessler, J.A. Cancer stem cells and glioma. Nat. Clin. Pract. Neurol. 2008, 4, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Suva, M.L.; Riggi, N.; Bernstein, B.E. Epigenetic reprogramming in cancer. Science 2013, 339, 1567–1570. [Google Scholar] [CrossRef]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef]
- Nieto, M.A.; Huang, R.Y.; Jackson, R.A.; Thiery, J.P. Emt: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef]
- Kong, D.; Li, Y.; Wang, Z.; Sarkar, F.H. Cancer Stem Cells and Epithelial-to-Mesenchymal Transition (EMT)-Phenotypic Cells: Are They Cousins or Twins? Cancers (Basel) 2011, 3, 716–729. [Google Scholar] [CrossRef]
- Singh, A.; Settleman, J. EMT, cancer stem cells and drug resistance: An emerging axis of evil in the war on cancer. Oncogene 2010, 29, 4741–4751. [Google Scholar] [CrossRef]
- Siebzehnrubl, F.A.; Silver, D.J.; Tugertimur, B.; Deleyrolle, L.P.; Siebzehnrubl, D.; Sarkisian, M.R.; Devers, K.G.; Yachnis, A.T.; Kupper, M.D.; Neal, D.; et al. The ZEB1 pathway links glioblastoma initiation, invasion and chemoresistance. EMBO Mol. Med. 2013, 5, 1196–1212. [Google Scholar] [CrossRef]
- Chow, K.H.; Park, H.J.; George, J.; Yamamoto, K.; Gallup, A.D.; Graber, J.H.; Chen, Y.; Jiang, W.; Steindler, D.A.; Neilson, E.G.; et al. S100A4 Is a Biomarker and Regulator of Glioma Stem Cells That Is Critical for Mesenchymal Transition in Glioblastoma. Cancer Res. 2017, 77, 5360–5373. [Google Scholar] [CrossRef]
- Mikheeva, S.A.; Mikheev, A.M.; Petit, A.; Beyer, R.; Oxford, R.G.; Khorasani, L.; Maxwell, J.P.; Glackin, C.A.; Wakimoto, H.; Gonzalez-Herrero, I.; et al. TWIST1 promotes invasion through mesenchymal change in human glioblastoma. Molecular cancer 2010, 9, 194. [Google Scholar] [CrossRef] [PubMed]
- Kuhnol, C.D.; Wurfel, C.; Staege, M.S.; Kramm, C. Snail homolog 1 is involved in epithelial-mesenchymal transition-like processes in human glioblastoma cells. Oncol. Lett. 2017, 13, 3882–3888. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yang, H.W.; Menon, L.G.; Black, P.M.; Carroll, R.S.; Johnson, M.D. SNAI2/Slug promotes growth and invasion in human gliomas. BMC Cancer 2010, 10, 301. [Google Scholar] [CrossRef] [PubMed]
- Bhat, K.P.; Salazar, K.L.; Balasubramaniyan, V.; Wani, K.; Heathcock, L.; Hollingsworth, F.; James, J.D.; Gumin, J.; Diefes, K.L.; Kim, S.H.; et al. The transcriptional coactivator TAZ regulates mesenchymal differentiation in malignant glioma. Genes Dev. 2011, 25, 2594–2609. [Google Scholar] [CrossRef]
- Bhat, K.P.L.; Balasubramaniyan, V.; Vaillant, B.; Ezhilarasan, R.; Hummelink, K.; Hollingsworth, F.; Wani, K.; Heathcock, L.; James, J.D.; Goodman, L.D.; et al. Mesenchymal differentiation mediated by NF-kappaB promotes radiation resistance in glioblastoma. Cancer Cell 2013, 24, 331–346. [Google Scholar] [CrossRef]
- Wang, J.; Cazzato, E.; Ladewig, E.; Frattini, V.; Rosenbloom, D.I.; Zairis, S.; Abate, F.; Liu, Z.; Elliott, O.; Shin, Y.J.; et al. Clonal evolution of glioblastoma under therapy. Nat. Genet. 2016, 48, 768–776. [Google Scholar] [CrossRef]
- Wang, Q.; Hu, B.; Hu, X.; Kim, H.; Squatrito, M.; Scarpace, L.; DeCarvalho, A.C.; Lyu, S.; Li, P.; Li, Y.; et al. Tumor Evolution of Glioma-Intrinsic Gene Expression Subtypes Associates with Immunological Changes in the Microenvironment. Cancer Cell 2017, 32, 42–56. [Google Scholar] [CrossRef]
- Segerman, A.; Niklasson, M.; Haglund, C.; Bergstrom, T.; Jarvius, M.; Xie, Y.; Westermark, A.; Sonmez, D.; Hermansson, A.; Kastemar, M.; et al. Clonal Variation in Drug and Radiation Response among Glioma-Initiating Cells Is Linked to Proneural-Mesenchymal Transition. Cell Rep. 2016, 17, 2994–3009. [Google Scholar] [CrossRef]
- Halliday, J.; Helmy, K.; Pattwell, S.S.; Pitter, K.L.; LaPlant, Q.; Ozawa, T.; Holland, E.C. In vivo radiation response of proneural glioma characterized by protective p53 transcriptional program and proneural-mesenchymal shift. Proc. Natl. Acad. Sci. USA 2014, 111, 5248–5253. [Google Scholar] [CrossRef]
- Neftel, C.; Laffy, J.; Filbin, M.G.; Hara, T.; Shore, M.E.; Rahme, G.J.; Richman, A.R.; Silverbush, D.; Shaw, M.L.; Hebert, C.M.; et al. An Integrative Model of Cellular States, Plasticity, and Genetics for Glioblastoma. Cell 2019, 178, 835–849. [Google Scholar] [CrossRef]
- Gawad, C.; Koh, W.; Quake, S.R. Single-cell genome sequencing: Current state of the science. Nat. Rev. Genet. 2016, 17, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Ziegenhain, C.; Vieth, B.; Parekh, S.; Reinius, B.; Guillaumet-Adkins, A.; Smets, M.; Leonhardt, H.; Heyn, H.; Hellmann, I.; Enard, W. Comparative Analysis of Single-Cell RNA Sequencing Methods. Mol. Cell 2017, 65, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Schwartzman, O.; Tanay, A. Single-cell epigenomics: Techniques and emerging applications. Nat. Rev. Genet. 2015, 16, 716–726. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Singh, A.K. Microfluidic Flow Cytometry for Single-Cell Protein Analysis. Methods Mol. Biol. 2015, 1346, 69–83. [Google Scholar] [CrossRef]
- Macaulay, I.C.; Ponting, C.P.; Voet, T. Single-Cell Multiomics: Multiple Measurements from Single Cells. Trends Genet. 2017, 33, 155–168. [Google Scholar] [CrossRef]
- Suva, M.L.; Tirosh, I. Single-Cell RNA Sequencing in Cancer: Lessons Learned and Emerging Challenges. Mol. Cell 2019, 75, 7–12. [Google Scholar] [CrossRef]
- Tsuji, T.; Ibaragi, S.; Hu, G.F. Epithelial-mesenchymal transition and cell cooperativity in metastasis. Cancer Res. 2009, 69, 7135–7139. [Google Scholar] [CrossRef]
- Friedl, P.; Wolf, K. Tube travel: The role of proteases in individual and collective cancer cell invasion. Cancer Res. 2008, 68, 7247–7249. [Google Scholar] [CrossRef]
- Roussos, E.T.; Condeelis, J.S.; Patsialou, A. Chemotaxis in cancer. Nat. Rev. Cancer 2011, 11, 573–587. [Google Scholar] [CrossRef]
- Sanz-Moreno, V.; Marshall, C.J. The plasticity of cytoskeletal dynamics underlying neoplastic cell migration. Curr. Opin. Cell Biol. 2010, 22, 690–696. [Google Scholar] [CrossRef]
- Tirosh, I.; Suva, M.L. Dissecting human gliomas by single-cell RNA sequencing. Neuro-oncology 2018, 20, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Daubon, T.; Leon, C.; Clarke, K.; Andrique, L.; Salabert, L.; Darbo, E.; Pineau, R.; Guerit, S.; Maitre, M.; Dedieu, S.; et al. Deciphering the complex role of thrombospondin-1 in glioblastoma development. Nat. Commun. 2019, 10, 1146. [Google Scholar] [CrossRef] [PubMed]
- Fayzullin, A.; Sandberg, C.J.; Spreadbury, M.; Saberniak, B.M.; Grieg, Z.; Skaga, E.; Langmoen, I.A.; Vik-Mo, E.O. Phenotypic and Expressional Heterogeneity in the Invasive Glioma Cells. Transl. Oncol. 2019, 12, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Kruthika, B.S.; Jain, R.; Arivazhagan, A.; Bharath, R.D.; Yasha, T.C.; Kondaiah, P.; Santosh, V. Transcriptome profiling reveals PDZ binding kinase as a novel biomarker in peritumoral brain zone of glioblastoma. J. Neurooncol. 2019, 141, 315–325. [Google Scholar] [CrossRef]
- Minata, M.; Audia, A.; Shi, J.; Lu, S.; Bernstock, J.; Pavlyukov, M.S.; Das, A.; Kim, S.H.; Shin, Y.J.; Lee, Y.; et al. Phenotypic Plasticity of Invasive Edge Glioma Stem-like Cells in Response to Ionizing Radiation. Cell Rep. 2019, 26, 1893–1905. [Google Scholar] [CrossRef]
- Tong, X.; Yang, P.; Wang, K.; Liu, Y.; Liu, X.; Shan, X.; Huang, R.; Zhang, K.; Wang, J. Survivin is a prognostic indicator in glioblastoma and may be a target of microRNA-218. Oncol. Lett. 2019, 18, 359–367. [Google Scholar] [CrossRef]
- Wang, C.C.; Hueng, D.Y.; Huang, A.F.; Chen, W.L.; Huang, S.M.; Yi-Hsin Chan, J. CD164 regulates proliferation, progression, and invasion of human glioblastoma cells. Oncotarget 2019, 10, 2041–2054. [Google Scholar] [CrossRef]
- Wang, X.; Sun, Y.; Xu, T.; Qian, K.; Huang, B.; Zhang, K.; Song, Z.; Qian, T.; Shi, J.; Li, L. HOXB13 promotes proliferation, migration, and invasion of glioblastoma through transcriptional upregulation of lncRNA HOXC-AS3. J. Cell Biochem. 2019, 120, 15527–15537. [Google Scholar] [CrossRef]
- Zeng, C.; Xing, W.; Liu, Y. Identification of UGP2 as a progression marker that promotes cell growth and motility in human glioma. J. Cell Biochem. 2019, 120, 12489–12499. [Google Scholar] [CrossRef]
- Franceschi, S.; Corsinovi, D.; Lessi, F.; Tantillo, E.; Aretini, P.; Menicagli, M.; Scopelliti, C.; Civita, P.; Pasqualetti, F.; Naccarato, A.G.; et al. Mitochondrial enzyme GLUD2 plays a critical role in glioblastoma progression. EBioMedicine 2018, 37, 56–67. [Google Scholar] [CrossRef]
- Griveau, A.; Seano, G.; Shelton, S.J.; Kupp, R.; Jahangiri, A.; Obernier, K.; Krishnan, S.; Lindberg, O.R.; Yuen, T.J.; Tien, A.C.; et al. A Glial Signature and Wnt7 Signaling Regulate Glioma-Vascular Interactions and Tumor Microenvironment. Cancer Cell 2018, 33, 874–889. [Google Scholar] [CrossRef] [PubMed]
- Gulluoglu, S.; Tuysuz, E.C.; Sahin, M.; Kuskucu, A.; Kaan Yaltirik, C.; Ture, U.; Kucukkaraduman, B.; Akbar, M.W.; Gure, A.O.; Bayrak, O.F.; et al. Simultaneous miRNA and mRNA transcriptome profiling of glioblastoma samples reveals a novel set of OncomiR candidates and their target genes. Brain Res. 2018, 1700, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Tsai, J.T.; Chao, T.Y.; Ma, H.I.; Liu, W.H. The STAT3/Slug Axis Enhances Radiation-Induced Tumor Invasion and Cancer Stem-like Properties in Radioresistant Glioblastoma. Cancers (Basel) 2018, 10, 512. [Google Scholar] [CrossRef] [PubMed]
- Moncayo, G.; Grzmil, M.; Smirnova, T.; Zmarz, P.; Huber, R.M.; Hynx, D.; Kohler, H.; Wang, Y.; Hotz, H.R.; Hynes, N.E.; et al. SYK inhibition blocks proliferation and migration of glioma cells and modifies the tumor microenvironment. Neuro-oncology 2018, 20, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Otani, Y.; Ichikawa, T.; Kurozumi, K.; Inoue, S.; Ishida, J.; Oka, T.; Shimizu, T.; Tomita, Y.; Hattori, Y.; Uneda, A.; et al. Fibroblast growth factor 13 regulates glioma cell invasion and is important for bevacizumab-induced glioma invasion. Oncogene 2018, 37, 777–786. [Google Scholar] [CrossRef]
- Yuan, J.; Levitin, H.M.; Frattini, V.; Bush, E.C.; Boyett, D.M.; Samanamud, J.; Ceccarelli, M.; Dovas, A.; Zanazzi, G.; Canoll, P.; et al. Single-cell transcriptome analysis of lineage diversity in high-grade glioma. Genome Med. 2018, 10, 57. [Google Scholar] [CrossRef]
- Darmanis, S.; Sloan, S.A.; Croote, D.; Mignardi, M.; Chernikova, S.; Samghababi, P.; Zhang, Y.; Neff, N.; Kowarsky, M.; Caneda, C.; et al. Single-Cell RNA-Seq Analysis of Infiltrating Neoplastic Cells at the Migrating Front of Human Glioblastoma. Cell Rep. 2017, 21, 1399–1410. [Google Scholar] [CrossRef]
- Che Mat, M.F.; Abdul Murad, N.A.; Ibrahim, K.; Mohd Mokhtar, N.; Wan Ngah, W.Z.; Harun, R.; Jamal, R. Silencing of PROS1 induces apoptosis and inhibits migration and invasion of glioblastoma multiforme cells. Int. J. Oncol. 2016, 49, 2359–2366. [Google Scholar] [CrossRef][Green Version]
- Zhou, P.; Erfani, S.; Liu, Z.; Jia, C.; Chen, Y.; Xu, B.; Deng, X.; Alfaro, J.E.; Chen, L.; Napier, D.; et al. CD151-alpha3beta1 integrin complexes are prognostic markers of glioblastoma and cooperate with EGFR to drive tumor cell motility and invasion. Oncotarget 2015, 6, 29675–29693. [Google Scholar] [CrossRef]
- Murray, D.W.; Didier, S.; Chan, A.; Paulino, V.; Van Aelst, L.; Ruggieri, R.; Tran, N.L.; Byrne, A.T.; Symons, M. Guanine nucleotide exchange factor Dock7 mediates HGF-induced glioblastoma cell invasion via Rac activation. Br. J. Cancer 2014, 110, 1307–1315. [Google Scholar] [CrossRef]
- Park, J.B.; Agnihotri, S.; Golbourn, B.; Bertrand, K.C.; Luck, A.; Sabha, N.; Smith, C.A.; Byron, S.; Zadeh, G.; Croul, S.; et al. Transcriptional profiling of GBM invasion genes identifies effective inhibitors of the LIM kinase-Cofilin pathway. Oncotarget 2014, 5, 9382–9395. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Han, L.; Dong, Y.; Tian, J.; Huang, W.; Liu, Z.; Jia, X.; Jiang, T.; Zhang, J.; Li, X.; et al. JAK2/STAT3 targeted therapy suppresses tumor invasion via disruption of the EGFRvIII/JAK2/STAT3 axis and associated focal adhesion in EGFRvIII-expressing glioblastoma. Neuro-oncology 2014, 16, 1229–1243. [Google Scholar] [CrossRef] [PubMed]
- Tie, X.; Han, S.; Meng, L.; Wang, Y.; Wu, A. NFAT1 is highly expressed in, and regulates the invasion of, glioblastoma multiforme cells. PLoS ONE 2013, 8, e66008. [Google Scholar] [CrossRef] [PubMed]
- Dhruv, H.D.; McDonough Winslow, W.S.; Armstrong, B.; Tuncali, S.; Eschbacher, J.; Kislin, K.; Loftus, J.C.; Tran, N.L.; Berens, M.E. Reciprocal activation of transcription factors underlies the dichotomy between proliferation and invasion of glioma cells. PLoS ONE 2013, 8, e72134. [Google Scholar] [CrossRef]
- De Rosa, A.; Pellegatta, S.; Rossi, M.; Tunici, P.; Magnoni, L.; Speranza, M.C.; Malusa, F.; Miragliotta, V.; Mori, E.; Finocchiaro, G.; et al. A radial glia gene marker, fatty acid binding protein 7 (FABP7), is involved in proliferation and invasion of glioblastoma cells. PLoS ONE 2012, 7, e52113. [Google Scholar] [CrossRef]
- Toussaint, L.G., 3rd; Nilson, A.E.; Goble, J.M.; Ballman, K.V.; James, C.D.; Lefranc, F.; Kiss, R.; Uhm, J.H. Galectin-1, a gene preferentially expressed at the tumor margin, promotes glioblastoma cell invasion. Mol. Cancer 2012, 11, 32. [Google Scholar] [CrossRef]
- Lo, H.W.; Zhu, H.; Cao, X.; Aldrich, A.; Ali-Osman, F. A novel splice variant of GLI1 that promotes glioblastoma cell migration and invasion. Cancer Res. 2009, 69, 6790–6798. [Google Scholar] [CrossRef]
- Baldwin, R.M.; Parolin, D.A.; Lorimer, I.A. Regulation of glioblastoma cell invasion by PKC iota and RhoB. Oncogene 2008, 27, 3587–3595. [Google Scholar] [CrossRef]
- Demuth, T.; Rennert, J.L.; Hoelzinger, D.B.; Reavie, L.B.; Nakada, M.; Beaudry, C.; Nakada, S.; Anderson, E.M.; Henrichs, A.N.; McDonough, W.S.; et al. Glioma cells on the run - the migratory transcriptome of 10 human glioma cell lines. BMC Genomics 2008, 9, 54. [Google Scholar] [CrossRef]
- Asano, K.; Duntsch, C.D.; Zhou, Q.; Weimar, J.D.; Bordelon, D.; Robertson, J.H.; Pourmotabbed, T. Correlation of N-cadherin expression in high grade gliomas with tissue invasion. J. Neurooncol. 2004, 70, 3–15. [Google Scholar] [CrossRef]
- Wang, H.; Wang, H.; Shen, W.; Huang, H.; Hu, L.; Ramdas, L.; Zhou, Y.H.; Liao, W.S.; Fuller, G.N.; Zhang, W. Insulin-like growth factor binding protein 2 enhances glioblastoma invasion by activating invasion-enhancing genes. Cancer Res. 2003, 63, 4315–4321. [Google Scholar] [PubMed]
- Ridley, A.J.; Schwartz, M.A.; Burridge, K.; Firtel, R.A.; Ginsberg, M.H.; Borisy, G.; Parsons, J.T.; Horwitz, A.R. Cell migration: Integrating signals from front to back. Science 2003, 302, 1704–1709. [Google Scholar] [CrossRef] [PubMed]
- Demuth, T.; Berens, M.E. Molecular mechanisms of glioma cell migration and invasion. J. Neurooncol. 2004, 70, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Wolfenson, H.; Lavelin, I.; Geiger, B. Dynamic regulation of the structure and functions of integrin adhesions. Dev. Cell 2013, 24, 447–458. [Google Scholar] [CrossRef]
- Hu, B.; Jarzynka, M.J.; Guo, P.; Imanishi, Y.; Schlaepfer, D.D.; Cheng, S.Y. Angiopoietin 2 induces glioma cell invasion by stimulating matrix metalloprotease 2 expression through the alphavbeta1 integrin and focal adhesion kinase signaling pathway. Cancer Res. 2006, 66, 775–783. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, P.; Yang, L.; Yu, X.; Ye, X.; Yang, J.; Qian, C.; Zhang, X.; Cui, Y.H.; Bian, X.W. Activation of toll-like receptor 2 promotes invasion by upregulating MMPs in glioma stem cells. Am. J. Transl. Res. 2015, 7, 607–615. [Google Scholar]
- Inoue, A.; Takahashi, H.; Harada, H.; Kohno, S.; Ohue, S.; Kobayashi, K.; Yano, H.; Tanaka, J.; Ohnishi, T. Cancer stem-like cells of glioblastoma characteristically express MMP-13 and display highly invasive activity. Int. J. Oncol. 2010, 37, 1121–1131. [Google Scholar] [CrossRef]
- Seals, D.F.; Courtneidge, S.A. The ADAMs family of metalloproteases: Multidomain proteins with multiple functions. Genes Dev. 2003, 17, 7–30. [Google Scholar] [CrossRef]
- Gole, B.; Huszthy, P.C.; Popovic, M.; Jeruc, J.; Ardebili, Y.S.; Bjerkvig, R.; Lah, T.T. The regulation of cysteine cathepsins and cystatins in human gliomas. Int. J. Cancer 2012, 131, 1779–1789. [Google Scholar] [CrossRef]
- Soroceanu, L.; Manning, T.J., Jr.; Sontheimer, H. Modulation of glioma cell migration and invasion using Cl(−) and K(+) ion channel blockers. J. Neurosci. 1999, 19, 5942–5954. [Google Scholar] [CrossRef]
- Beadle, C.; Assanah, M.C.; Monzo, P.; Vallee, R.; Rosenfeld, S.S.; Canoll, P. The role of myosin II in glioma invasion of the brain. Mol. Biol. Cell 2008, 19, 3357–3368. [Google Scholar] [CrossRef] [PubMed]
- Picariello, H.S.; Kenchappa, R.S.; Rai, V.; Crish, J.F.; Dovas, A.; Pogoda, K.; McMahon, M.; Bell, E.S.; Chandrasekharan, U.; Luu, A.; et al. Myosin IIA suppresses glioblastoma development in a mechanically sensitive manner. Proc. Natl. Acad. Sci. USA 2019, 116, 15550–15559. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, D.; Annovazzi, L.; Casalone, C.; Corona, C.; Mellai, M. Glioblastoma: Microenvironment and Niche Concept. Cancers (Basel) 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Watkins, S.; Robel, S.; Kimbrough, I.F.; Robert, S.M.; Ellis-Davies, G.; Sontheimer, H. Disruption of astrocyte-vascular coupling and the blood-brain barrier by invading glioma cells. Nat. Commun. 2014, 5, 4196. [Google Scholar] [CrossRef]
- Venkataramani, V.; Tanev, D.I.; Strahle, C.; Studier-Fischer, A.; Fankhauser, L.; Kessler, T.; Korber, C.; Kardorff, M.; Ratliff, M.; Xie, R.; et al. Glutamatergic synaptic input to glioma cells drives brain tumour progression. Nature 2019, 573, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Messeguer, X.; Escudero, R.; Farre, D.; Nunez, O.; Martinez, J.; Alba, M.M. PROMO: Detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 2002, 18, 333–334. [Google Scholar] [CrossRef]
- Yi, Y.; Hsieh, I.Y.; Huang, X.; Li, J.; Zhao, W. Glioblastoma Stem-Like Cells: Characteristics, Microenvironment, and Therapy. Front. Pharmacol. 2016, 7, 477. [Google Scholar] [CrossRef]
- Li, G.; Qin, Z.; Chen, Z.; Xie, L.; Wang, R.; Zhao, H. Tumor Microenvironment in Treatment of Glioma. Open Med. (Wars) 2017, 12, 247–251. [Google Scholar] [CrossRef]
- McGranahan, N.; Swanton, C. Clonal Heterogeneity and Tumor Evolution: Past, Present, and the Future. Cell 2017, 168, 613–628. [Google Scholar] [CrossRef]
- Zimmermann, D.R.; Dours-Zimmermann, M.T. Extracellular matrix of the central nervous system: From neglect to challenge. Histochem. Cell Biol. 2008, 130, 635–653. [Google Scholar] [CrossRef]
- Park, J.B.; Kwak, H.J.; Lee, S.H. Role of hyaluronan in glioma invasion. Cell Adh. Migr. 2008, 2, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Arslan, F.; Bosserhoff, A.K.; Nickl-Jockschat, T.; Doerfelt, A.; Bogdahn, U.; Hau, P. The role of versican isoforms V0/V1 in glioma migration mediated by transforming growth factor-beta2. Br. J. Cancer 2007, 96, 1560–1568. [Google Scholar] [CrossRef] [PubMed]
- Baumann, F.; Leukel, P.; Doerfelt, A.; Beier, C.P.; Dettmer, K.; Oefner, P.J.; Kastenberger, M.; Kreutz, M.; Nickl-Jockschat, T.; Bogdahn, U.; et al. Lactate promotes glioma migration by TGF-beta2-dependent regulation of matrix metalloproteinase-2. Neuro-Oncology 2009, 11, 368–380. [Google Scholar] [CrossRef] [PubMed]
- Hau, P.; Kunz-Schughart, L.A.; Rummele, P.; Arslan, F.; Dorfelt, A.; Koch, H.; Lohmeier, A.; Hirschmann, B.; Muller, A.; Bogdahn, U.; et al. Tenascin-C protein is induced by transforming growth factor-beta1 but does not correlate with time to tumor progression in high-grade gliomas. J. Neurooncol. 2006, 77, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Onken, J.; Moeckel, S.; Leukel, P.; Leidgens, V.; Baumann, F.; Bogdahn, U.; Vollmann-Zwerenz, A.; Hau, P. Versican isoform V1 regulates proliferation and migration in high-grade gliomas. J. Neurooncol. 2014, 120, 73–83. [Google Scholar] [CrossRef]
- Medema, J.P. Cancer stem cells: The challenges ahead. Nat. Cell Biol. 2013, 15, 338–344. [Google Scholar] [CrossRef]
- Calabrese, C.; Poppleton, H.; Kocak, M.; Hogg, T.L.; Fuller, C.; Hamner, B.; Oh, E.Y.; Gaber, M.W.; Finklestein, D.; Allen, M.; et al. A perivascular niche for brain tumor stem cells. Cancer Cell 2007, 11, 69–82. [Google Scholar] [CrossRef]
- Montana, V.; Sontheimer, H. Bradykinin promotes the chemotactic invasion of primary brain tumors. J. Neurosci. 2011, 31, 4858–4867. [Google Scholar] [CrossRef]
- Ping, Y.F.; Yao, X.H.; Jiang, J.Y.; Zhao, L.T.; Yu, S.C.; Jiang, T.; Lin, M.C.; Chen, J.H.; Wang, B.; Zhang, R.; et al. The chemokine CXCL12 and its receptor CXCR4 promote glioma stem cell-mediated VEGF production and tumour angiogenesis via PI3K/AKT signalling. J. Pathol. 2011, 224, 344–354. [Google Scholar] [CrossRef]
- Hynes, R.O. The extracellular matrix: Not just pretty fibrils. Science 2009, 326, 1216–1219. [Google Scholar] [CrossRef]
- Nguyen-Ngoc, K.V.; Cheung, K.J.; Brenot, A.; Shamir, E.R.; Gray, R.S.; Hines, W.C.; Yaswen, P.; Werb, Z.; Ewald, A.J. ECM microenvironment regulates collective migration and local dissemination in normal and malignant mammary epithelium. Proc. Natl. Acad. Sci. USA 2012, 109, E2595–E2604. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bao, S.; Wu, Q.; Wang, H.; Eyler, C.; Sathornsumetee, S.; Shi, Q.; Cao, Y.; Lathia, J.; McLendon, R.E.; et al. Hypoxia-inducible factors regulate tumorigenic capacity of glioma stem cells. Cancer Cell 2009, 15, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Filatova, A.; Seidel, S.; Bogurcu, N.; Graf, S.; Garvalov, B.K.; Acker, T. Acidosis Acts through HSP90 in a PHD/VHL-Independent Manner to Promote HIF Function and Stem Cell Maintenance in Glioma. Cancer Res. 2016, 76, 5845–5856. [Google Scholar] [CrossRef] [PubMed]
- Cong, D.; Zhu, W.; Shi, Y.; Pointer, K.B.; Clark, P.A.; Shen, H.; Kuo, J.S.; Hu, S.; Sun, D. Upregulation of NHE1 protein expression enables glioblastoma cells to escape TMZ-mediated toxicity via increased H(+) extrusion, cell migration and survival. Carcinogenesis 2014, 35, 2014–2024. [Google Scholar] [CrossRef] [PubMed]
- Roos, A.; Ding, Z.; Loftus, J.C.; Tran, N.L. Molecular and Microenvironmental Determinants of Glioma Stem-Like Cell Survival and Invasion. Front. Oncol. 2017, 7, 120. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Martin, V.; Fueyo, J.; Lee, O.H.; Xu, J.; Cortes-Santiago, N.; Alonso, M.M.; Aldape, K.; Colman, H.; Gomez-Manzano, C. Tie2/TEK modulates the interaction of glioma and brain tumor stem cells with endothelial cells and promotes an invasive phenotype. Oncotarget 2010, 1, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Dzaye, O.; Hu, F.; Derkow, K.; Haage, V.; Euskirchen, P.; Harms, C.; Lehnardt, S.; Synowitz, M.; Wolf, S.A.; Kettenmann, H. Glioma Stem Cells but Not Bulk Glioma Cells Upregulate IL-6 Secretion in Microglia/Brain Macrophages via Toll-like Receptor 4 Signaling. J. Neuropathol. Exp. Neurol. 2016, 75, 429–440. [Google Scholar] [CrossRef]
- Markovic, D.S.; Vinnakota, K.; Chirasani, S.; Synowitz, M.; Raguet, H.; Stock, K.; Sliwa, M.; Lehmann, S.; Kalin, R.; Van Rooijen, N.; et al. Gliomas induce and exploit microglial MT1-MMP expression for tumor expansion. Proc. Natl. Acad. Sci. USA 2009, 106, 12530–12535. [Google Scholar] [CrossRef]
- Placone, A.L.; Quinones-Hinojosa, A.; Searson, P.C. The role of astrocytes in the progression of brain cancer: Complicating the picture of the tumor microenvironment. Tumour Biology 2016, 37, 61–69. [Google Scholar] [CrossRef]
- Rolon-Reyes, K.; Kucheryavykh, Y.V.; Cubano, L.A.; Inyushin, M.; Skatchkov, S.N.; Eaton, M.J.; Harrison, J.K.; Kucheryavykh, L.Y. Microglia Activate Migration of Glioma Cells through a Pyk2 Intracellular Pathway. PLoS ONE 2015, 10, e0131059. [Google Scholar] [CrossRef]
- Pietras, A.; Katz, A.M.; Ekstrom, E.J.; Wee, B.; Halliday, J.J.; Pitter, K.L.; Werbeck, J.L.; Amankulor, N.M.; Huse, J.T.; Holland, E.C. Osteopontin-CD44 signaling in the glioma perivascular niche enhances cancer stem cell phenotypes and promotes aggressive tumor growth. Cell Stem. Cell 2014, 14, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Coniglio, S.J.; Eugenin, E.; Dobrenis, K.; Stanley, E.R.; West, B.L.; Symons, M.H.; Segall, J.E. Microglial stimulation of glioblastoma invasion involves epidermal growth factor receptor (EGFR) and colony stimulating factor 1 receptor (CSF-1R) signaling. Mol. Med. 2012, 18, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.Z.; Xu, S.L.; Xin, Y.H.; Yu, S.C.; Ping, Y.F.; Chen, L.; Xiao, H.L.; Wang, B.; Yi, L.; Wang, Q.L.; et al. Tumor-associated microglia/macrophages enhance the invasion of glioma stem-like cells via TGF-beta1 signaling pathway. J. Immunol. 2012, 189, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Katt, M.E.; Placone, A.L.; Wong, A.D.; Xu, Z.S.; Searson, P.C. In Vitro Tumor Models: Advantages, Disadvantages, Variables, and Selecting the Right Platform. Front. Bioeng. Biotechnol. 2016, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.M.; Cukierman, E. Modeling tissue morphogenesis and cancer in 3D. Cell 2007, 130, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Rape, A.; Ananthanarayanan, B.; Kumar, S. Engineering strategies to mimic the glioblastoma microenvironment. Adv. Drug Deliv. Rev. 2014, 79–80, 172–183. [Google Scholar] [CrossRef]
- Valster, A.; Tran, N.L.; Nakada, M.; Berens, M.E.; Chan, A.Y.; Symons, M. Cell migration and invasion assays. Methods 2005, 37, 208–215. [Google Scholar] [CrossRef]
- Watkins, S.; Sontheimer, H. Hydrodynamic cellular volume changes enable glioma cell invasion. J. Neurosci. 2011, 31, 17250–17259. [Google Scholar] [CrossRef]
- Bonneh-Barkay, D.; Wiley, C.A. Brain extracellular matrix in neurodegeneration. Brain Pathol. 2009, 19, 573–585. [Google Scholar] [CrossRef]
- Lau, L.W.; Cua, R.; Keough, M.B.; Haylock-Jacobs, S.; Yong, V.W. Pathophysiology of the brain extracellular matrix: A new target for remyelination. Nat. Rev. Neurosci. 2013, 14, 722–729. [Google Scholar] [CrossRef]
- Weiswald, L.B.; Bellet, D.; Dangles-Marie, V. Spherical cancer models in tumor biology. Neoplasia 2015, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bjerkvig, R.; Tonnesen, A.; Laerum, O.D.; Backlund, E.O. Multicellular tumor spheroids from human gliomas maintained in organ culture. J. Neurosurg. 1990, 72, 463–475. [Google Scholar] [CrossRef]
- Mahesparan, R.; Tysnes, B.B.; Read, T.A.; Enger, P.O.; Bjerkvig, R.; Lund-Johansen, M. Extracellular matrix-induced cell migration from glioblastoma biopsy specimens in vitro. Acta Neuropathol. 1999, 97, 231–239. [Google Scholar] [CrossRef]
- Hubert, C.G.; Rivera, M.; Spangler, L.C.; Wu, Q.; Mack, S.C.; Prager, B.C.; Couce, M.; McLendon, R.E.; Sloan, A.E.; Rich, J.N. A Three-Dimensional Organoid Culture System Derived from Human Glioblastomas Recapitulates the Hypoxic Gradients and Cancer Stem Cell Heterogeneity of Tumors Found In Vivo. Cancer Res. 2016, 76, 2465–2477. [Google Scholar] [CrossRef] [PubMed]
- Linkous, A.; Balamatsias, D.; Snuderl, M.; Edwards, L.; Miyaguchi, K.; Milner, T.; Reich, B.; Cohen-Gould, L.; Storaska, A.; Nakayama, Y.; et al. Modeling Patient-Derived Glioblastoma with Cerebral Organoids. Cell Rep. 2019, 26, 3203–3211. [Google Scholar] [CrossRef] [PubMed]
- Gogolla, N.; Galimberti, I.; DePaola, V.; Caroni, P. Long-term live imaging of neuronal circuits in organotypic hippocampal slice cultures. Nat. Protoc. 2006, 1, 1223–1226. [Google Scholar] [CrossRef] [PubMed]
- Gogolla, N.; Galimberti, I.; DePaola, V.; Caroni, P. Preparation of organotypic hippocampal slice cultures for long-term live imaging. Nat. Protoc. 2006, 1, 1165–1171. [Google Scholar] [CrossRef] [PubMed]
- Aaberg-Jessen, C.; Norregaard, A.; Christensen, K.; Pedersen, C.B.; Andersen, C.; Kristensen, B.W. Invasion of primary glioma- and cell line-derived spheroids implanted into corticostriatal slice cultures. Int. J. Clin. Exp. Pathol. 2013, 6, 546–560. [Google Scholar] [PubMed]
- Humpel, C. Organotypic brain slice cultures: A review. Neuroscience 2015, 305, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Eyupoglu, I.Y.; Hahnen, E.; Heckel, A.; Siebzehnrubl, F.A.; Buslei, R.; Fahlbusch, R.; Blumcke, I. Malignant glioma-induced neuronal cell death in an organotypic glioma invasion model. J. Neurosurg. 2005, 102, 738–744. [Google Scholar] [CrossRef]
- Haeger, A.; Krause, M.; Wolf, K.; Friedl, P. Cell jamming: Collective invasion of mesenchymal tumor cells imposed by tissue confinement. Bioch. Biophy. Acta 2014, 1840, 2386–2395. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.A.; Seidl, S.; Petat-Dutter, K.; Offner, S.; Geigl, J.B.; Schmidt-Kittler, O.; Wendler, N.; Passlick, B.; Huber, R.M.; Schlimok, G.; et al. Combined transcriptome and genome analysis of single micrometastatic cells. Nat. Biotechnol. 2002, 20, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Gogolla, N.; Galimberti, I.; DePaola, V.; Caroni, P. Staining protocol for organotypic hippocampal slice cultures. Nat. Protoc. 2006, 1, 2452–2456. [Google Scholar] [CrossRef] [PubMed]
- Alieva, M.; Margarido, A.S.; Wieles, T.; Abels, E.R.; Colak, B.; Boquetale, C.; Jan Noordmans, H.; Snijders, T.J.; Broekman, M.L.; Van Rheenen, J. Preventing inflammation inhibits biopsy-mediated changes in tumor cell behavior. Sci. Rep. 2017, 7, 7529. [Google Scholar] [CrossRef]
- Alfonso, J.C.L.; Talkenberger, K.; Seifert, M.; Klink, B.; Hawkins-Daarud, A.; Swanson, K.R.; Hatzikirou, H.; Deutsch, A. The biology and mathematical modelling of glioma invasion: A review. J. R. Soc. Interface 2017, 14. [Google Scholar] [CrossRef]
- Kong, J.; Cooper, L.A.; Wang, F.; Gutman, D.A.; Gao, J.; Chisolm, C.; Sharma, A.; Pan, T.; Van Meir, E.G.; Kurc, T.M.; et al. Integrative, multimodal analysis of glioblastoma using TCGA molecular data, pathology images, and clinical outcomes. IEEE Trans. Biomed. Eng. 2011, 58, 3469–3474. [Google Scholar] [CrossRef]
- Madhavan, S.; Zenklusen, J.C.; Kotliarov, Y.; Sahni, H.; Fine, H.A.; Buetow, K. Rembrandt: Helping personalized medicine become a reality through integrative translational research. Mol. Cancer Res. 2009, 7, 157–167. [Google Scholar] [CrossRef]
- Bolouri, H.; Zhao, L.P.; Holland, E.C. Big data visualization identifies the multidimensional molecular landscape of human gliomas. Proc. Natl. Acad. Sci. USA 2016, 113, 5394–5399. [Google Scholar] [CrossRef]
- Puchalski, R.B.; Shah, N.; Miller, J.; Dalley, R.; Nomura, S.R.; Yoon, J.G.; Smith, K.A.; Lankerovich, M.; Bertagnolli, D.; Bickley, K.; et al. An anatomic transcriptional atlas of human glioblastoma. Science 2018, 360, 660–663. [Google Scholar] [CrossRef]
- Celiku, O.; Gilbert, M.R.; Lavi, O. Computational modeling demonstrates that glioblastoma cells can survive spatial environmental challenges through exploratory adaptation. Nat. Commun. 2019, 10, 5704. [Google Scholar] [CrossRef]
- Datta, S.; Malhotra, L.; Dickerson, R.; Chaffee, S.; Sen, C.K.; Roy, S. Laser capture microdissection: Big data from small samples. Histol. Histopathol. 2015, 30, 1255–1269. [Google Scholar] [CrossRef] [PubMed]
- Casasent, A.K.; Schalck, A.; Gao, R.; Sei, E.; Long, A.; Pangburn, W.; Casasent, T.; Meric-Bernstam, F.; Edgerton, M.E.; Navin, N.E. Multiclonal Invasion in Breast Tumors Identified by Topographic Single Cell Sequencing. Cell 2018, 172, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Yoosuf, N.; Navarro, J.F.; Salmen, F.; Stahl, P.L.; Daub, C.O. Identification and transfer of spatial transcriptomics signatures for cancer diagnosis. Breast Cancer Res. 2020, 22, 6. [Google Scholar] [CrossRef] [PubMed]
- Berglund, E.; Maaskola, J.; Schultz, N.; Friedrich, S.; Marklund, M.; Bergenstrahle, J.; Tarish, F.; Tanoglidi, A.; Vickovic, S.; Larsson, L.; et al. Spatial maps of prostate cancer transcriptomes reveal an unexplored landscape of heterogeneity. Nat. Commun. 2018, 9, 2419. [Google Scholar] [CrossRef] [PubMed]
- Thrane, K.; Eriksson, H.; Maaskola, J.; Hansson, J.; Lundeberg, J. Spatially Resolved Transcriptomics Enables Dissection of Genetic Heterogeneity in Stage III Cutaneous Malignant Melanoma. Cancer Res. 2018, 78, 5970–5979. [Google Scholar] [CrossRef] [PubMed]
- Vickovic, S.; Eraslan, G.; Salmen, F.; Klughammer, J.; Stenbeck, L.; Schapiro, D.; Aijo, T.; Bonneau, R.; Bergenstrahle, L.; Navarro, J.F.; et al. High-definition spatial transcriptomics for in situ tissue profiling. Nat. Methods 2019, 16, 987–990. [Google Scholar] [CrossRef]
- Carlberg, K.; Korotkova, M.; Larsson, L.; Catrina, A.I.; Stahl, P.L.; Malmstrom, V. Exploring inflammatory signatures in arthritic joint biopsies with Spatial Transcriptomics. Sci. Rep. 2019, 9, 18975. [Google Scholar] [CrossRef]
- Pombo Antunes, A.R.; Scheyltjens, I.; Duerinck, J.; Neyns, B.; Movahedi, K.; Van Ginderachter, J.A. Understanding the glioblastoma immune microenvironment as basis for the development of new immunotherapeutic strategies. Elife 2020. [Google Scholar] [CrossRef]
- Hambardzumyan, D.; Gutmann, D.H.; Kettenmann, H. The role of microglia and macrophages in glioma maintenance and progression. Nat. Neurosci. 2016, 19, 20–27. [Google Scholar] [CrossRef]
- Moncada, R.; Barkley, D.; Wagner, F.; Chiodin, M.; Devlin, J.C.; Baron, M.; Hajdu, C.H.; Simeone, D.M.; Yanai, I. Integrating microarray-based spatial transcriptomics and single-cell RNA-seq reveals tissue architecture in pancreatic ductal adenocarcinomas. Nat. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Baccin, C.; Al-Sabah, J.; Velten, L.; Helbling, P.M.; Grunschlager, F.; Hernandez-Malmierca, P.; Nombela-Arrieta, C.; Steinmetz, L.M.; Trumpp, A.; Haas, S. Combined single-cell and spatial transcriptomics reveal the molecular, cellular and spatial bone marrow niche organization. Nat. Cell Biol. 2020, 22, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Shboul, Z.A.; Alam, M.; Vidyaratne, L.; Pei, L.; Elbakary, M.I.; Iftekharuddin, K.M. Feature-Guided Deep Radiomics for Glioblastoma Patient Survival Prediction. Front. Neurosci. 2019, 13, 966. [Google Scholar] [CrossRef] [PubMed]
- Elshafeey, N.; Kotrotsou, A.; Hassan, A.; Elshafei, N.; Hassan, I.; Ahmed, S.; Abrol, S.; Agarwal, A.; El Salek, K.; Bergamaschi, S.; et al. Multicenter study demonstrates radiomic features derived from magnetic resonance perfusion images identify pseudoprogression in glioblastoma. Nat. Commun. 2019, 10, 3170. [Google Scholar] [CrossRef] [PubMed]
- Chaddad, A.; Kucharczyk, M.J.; Daniel, P.; Sabri, S.; Jean-Claude, B.J.; Niazi, T.; Abdulkarim, B. Radiomics in Glioblastoma: Current Status and Challenges Facing Clinical Implementation. Front. Oncol. 2019, 9, 374. [Google Scholar] [CrossRef]
- Zinn, P.O.; Singh, S.K.; Kotrotsou, A.; Hassan, I.; Thomas, G.; Luedi, M.M.; Elakkad, A.; Elshafeey, N.; Idris, T.; Mosley, J.; et al. A Coclinical Radiogenomic Validation Study: Conserved Magnetic Resonance Radiomic Appearance of Periostin-Expressing Glioblastoma in Patients and Xenograft Models. Clin. Cancer Res. 2018, 24, 6288–6299. [Google Scholar] [CrossRef]
- Zinn, P.O.; Mahajan, B.; Sathyan, P.; Singh, S.K.; Majumder, S.; Jolesz, F.A.; Colen, R.R. Radiogenomic mapping of edema/cellular invasion MRI-phenotypes in glioblastoma multiforme. PLoS ONE 2011, 6, e25451. [Google Scholar] [CrossRef]
- Colen, R.R.; Vangel, M.; Wang, J.; Gutman, D.A.; Hwang, S.N.; Wintermark, M.; Jain, R.; Jilwan-Nicolas, M.; Chen, J.Y.; Raghavan, P.; et al. Imaging genomic mapping of an invasive MRI phenotype predicts patient outcome and metabolic dysfunction: A TCGA glioma phenotype research group project. BMC Med. Genomics 2014, 7, 30. [Google Scholar] [CrossRef]
- Hu, L.S.; Ning, S.; Eschbacher, J.M.; Baxter, L.C.; Gaw, N.; Ranjbar, S.; Plasencia, J.; Dueck, A.C.; Peng, S.; Smith, K.A.; et al. Radiogenomics to characterize regional genetic heterogeneity in glioblastoma. Neuro-Oncology 2017, 19, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Protopapa, M.; Zygogianni, A.; Stamatakos, G.S.; Antypas, C.; Armpilia, C.; Uzunoglu, N.K.; Kouloulias, V. Clinical implications of in silico mathematical modeling for glioblastoma: A critical review. J. Neurooncol. 2018, 136, 1–11. [Google Scholar] [CrossRef]
- Mariani, L.; Beaudry, C.; McDonough, W.S.; Hoelzinger, D.B.; Demuth, T.; Ross, K.R.; Berens, T.; Coons, S.W.; Watts, G.; Trent, J.M.; et al. Glioma cell motility is associated with reduced transcription of proapoptotic and proliferation genes: A cDNA microarray analysis. J. Neurooncol. 2001, 53, 161–176. [Google Scholar] [CrossRef]
- Molina, J.R.; Hayashi, Y.; Stephens, C.; Georgescu, M.M. Invasive glioblastoma cells acquire stemness and increased Akt activation. Neoplasia 2010, 12, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, T.; Reed, J.C. Tumor suppressor p53 is a direct transcriptional activator of the human bax gene. Cell 1995, 80, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Waldman, T.; Kinzler, K.W.; Vogelstein, B. p21 is necessary for the p53-mediated G1 arrest in human cancer cells. Cancer Res. 1995, 55, 5187–5190. [Google Scholar] [PubMed]
- Muller, P.A.; Vousden, K.H.; Norman, J.C. p53 and its mutants in tumor cell migration and invasion. J. Cell Biol. 2011, 192, 209–218. [Google Scholar] [CrossRef] [PubMed]
- De Toni-Costes, F.; Despeaux, M.; Bertrand, J.; Bourogaa, E.; Ysebaert, L.; Payrastre, B.; Racaud-Sultan, C. A New alpha5beta1 integrin-dependent survival pathway through GSK3beta activation in leukemic cells. PLoS ONE 2010, 5, e9807. [Google Scholar] [CrossRef] [PubMed]
- Eke, I.; Koch, U.; Hehlgans, S.; Sandfort, V.; Stanchi, F.; Zips, D.; Baumann, M.; Shevchenko, A.; Pilarsky, C.; Haase, M.; et al. PINCH1 regulates Akt1 activation and enhances radioresistance by inhibiting PP1alpha. J. Clin. Investig. 2010, 120, 2516–2527. [Google Scholar] [CrossRef] [PubMed]
- Damiano, J.S.; Cress, A.E.; Hazlehurst, L.A.; Shtil, A.A.; Dalton, W.S. Cell adhesion mediated drug resistance (CAM-DR): Role of integrins and resistance to apoptosis in human myeloma cell lines. Blood 1999, 93, 1658–1667. [Google Scholar] [CrossRef]
- Cordes, N.; Meineke, V. Cell adhesion-mediated radioresistance (CAM-RR). Extracellular matrix-dependent improvement of cell survival in human tumor and normal cells in vitro. Strahlenther. Onkol. 2003, 179, 337–344. [Google Scholar] [CrossRef]
- Mitra, S.K.; Schlaepfer, D.D. Integrin-regulated FAK-Src signaling in normal and cancer cells. Curr. Opin. Cell Biol. 2006, 18, 516–523. [Google Scholar] [CrossRef]
- Klemm, F.; Joyce, J.A. Microenvironmental regulation of therapeutic response in cancer. Trends Cell Biol. 2015, 25, 198–213. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).