Effects of Osiris9a on Silk Properties in Bombyx mori Determined by Transgenic Overexpression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
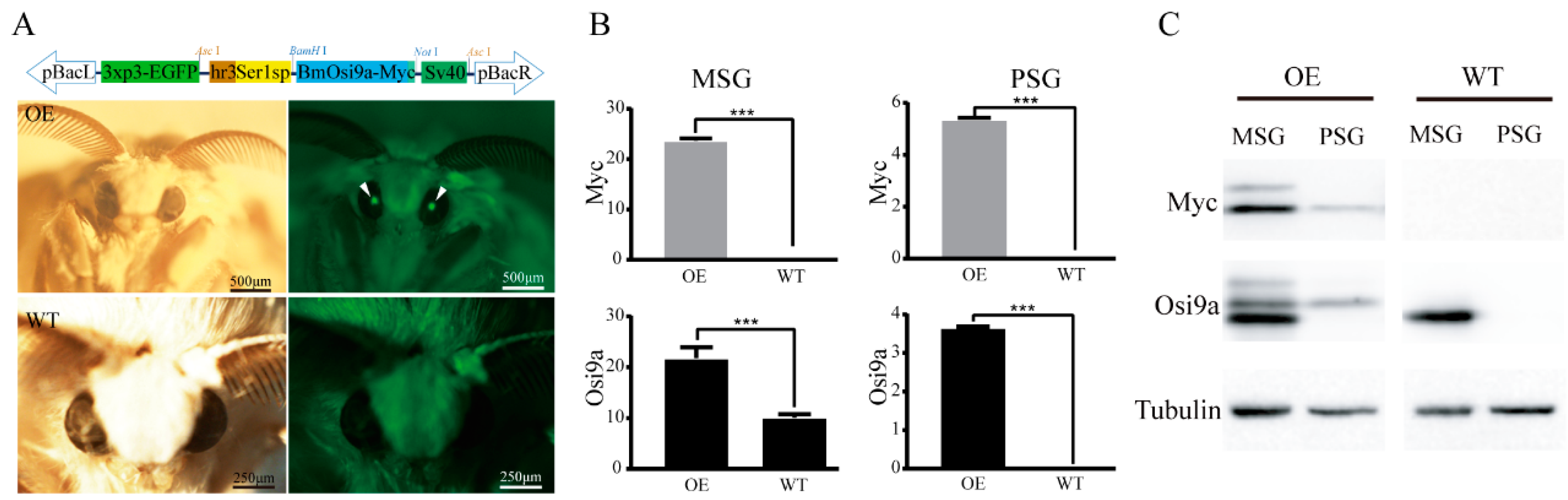
2.1. Transgenic Overexpression of BmOsi9a in the Silk Gland
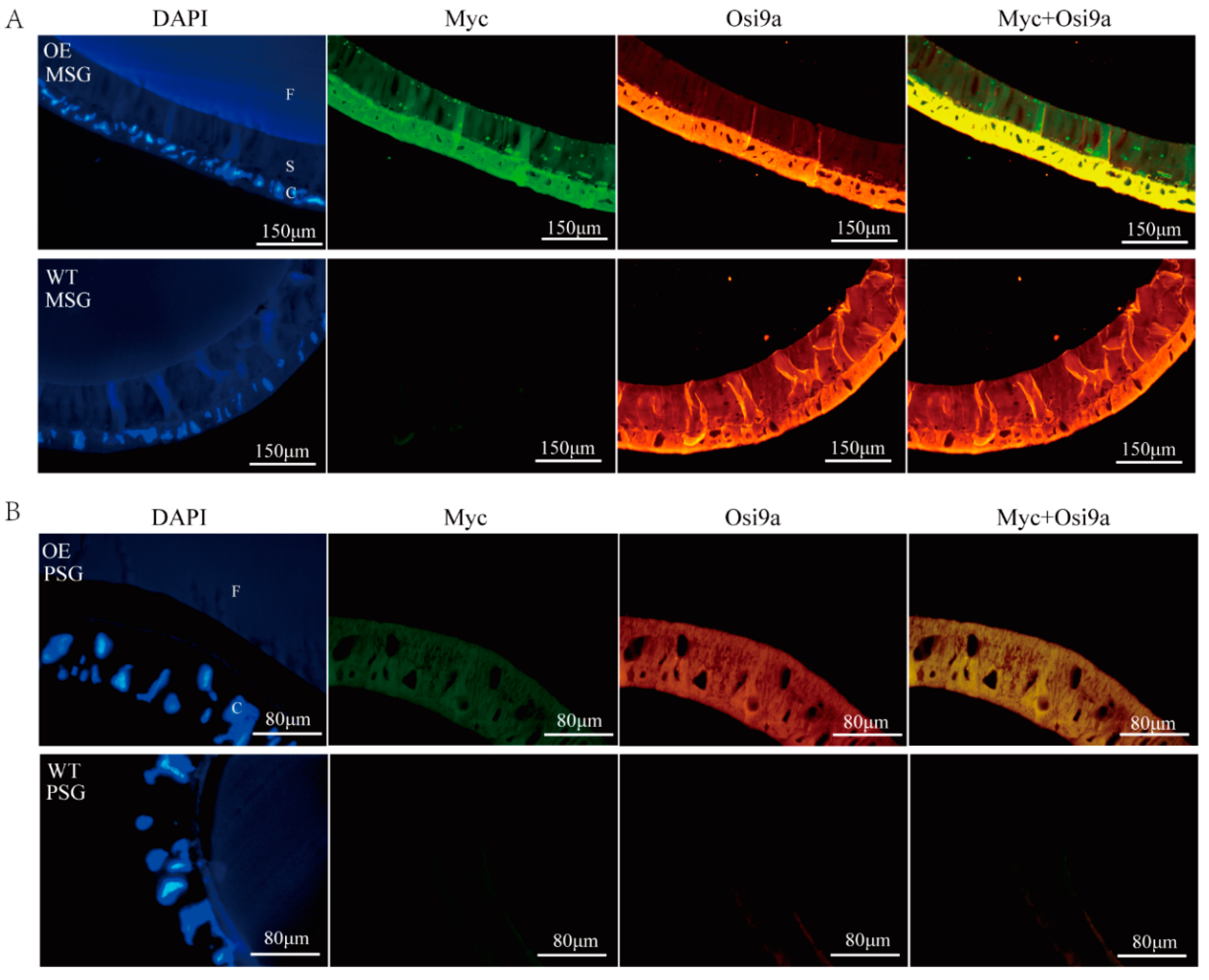
2.2. Location of BmOsi9a in the Silk Gland
2.3. Interaction between BmOsi9a and Ser1
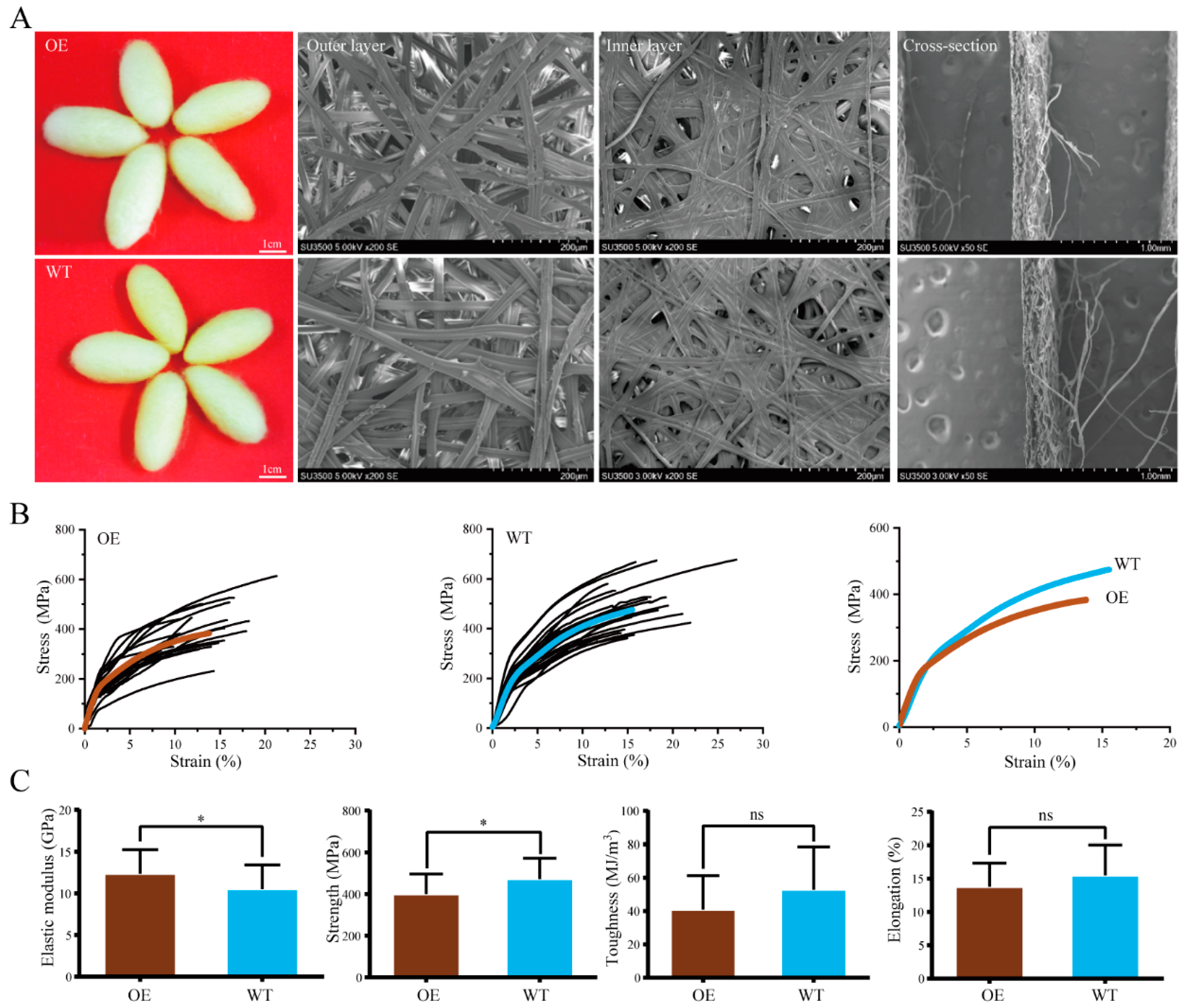
2.4. Mechanical Properties of Silk
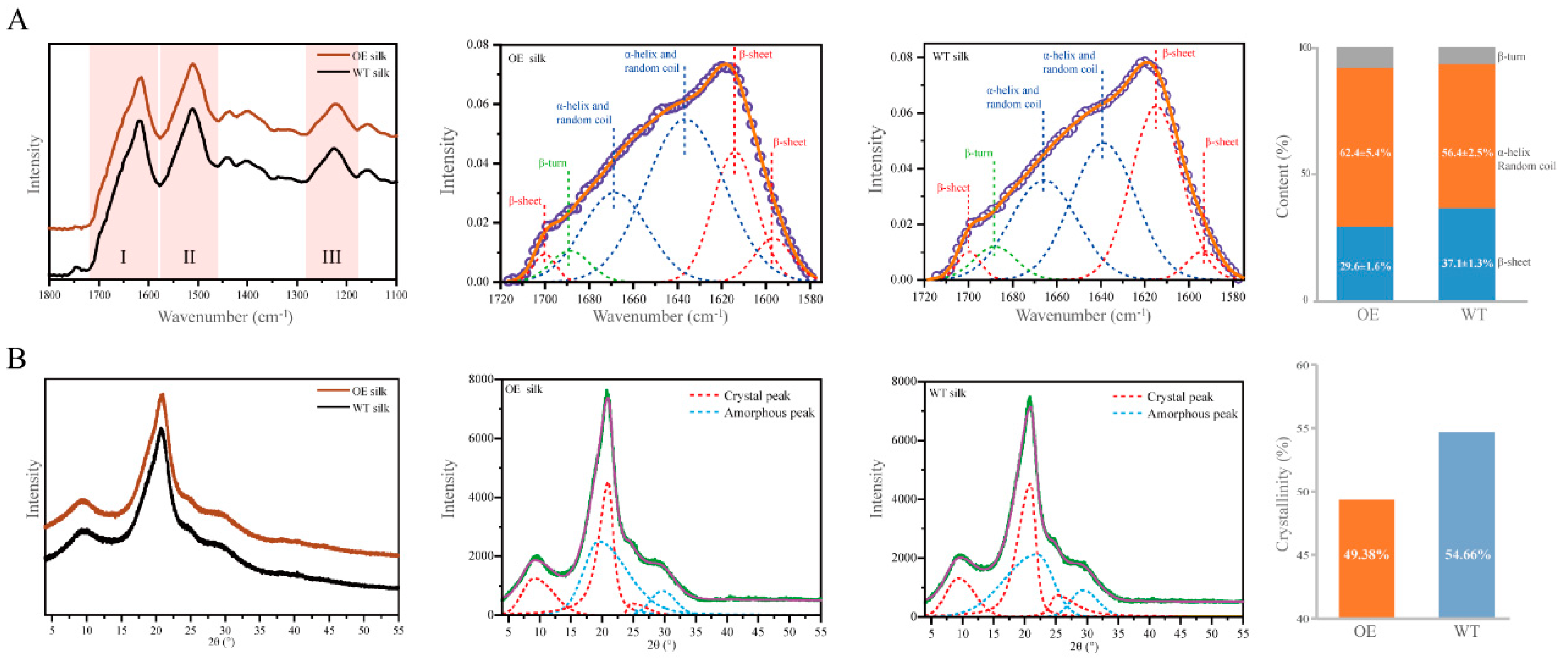
2.5. Secondary Structural and Crystal Morphological Characteristics of Silk Fibers
3. Discussion
4. Materials and Methods
4.1. Insects
4.2. Vector Construction
4.3. Germline Transformation of the Silkworm
4.4. Quantitative Real-Time PCR
4.5. Western Blotting
4.6. Immunohistochemistry and Co-Immunoprecipitation (Co-IP)
4.7. Mechanical Testing of Silk Fiber
4.8. ATR-FTIR and XRD Analyses
4.9. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| MSG | middle silk gland |
| PSG | posterior silk gland |
| Osi | Osiris |
| Ser | sericin |
| qRT-PCR | quantitative real-time PCR |
| Co-IP | co-Immunoprecipitation |
| ATR-FTIR | attenuated total reflectance Fourier-transform infrared spectroscopy |
| XRD | X-ray diffraction |
| OE | overexpression |
| WT | wild-type |
References
- Dorer, D.R.; Rudnick, J.A.; Moriyama, E.N.; Christensen, A.C. A family of genes clustered at the Triplo-lethal locus of Drosophila melanogaster has an unusual evolutionary history and significant synteny with Anopheles gambiae. Genetics 2003, 165, 613–621. [Google Scholar]
- Shah, N.; Dorer, D.R.; Moriyama, E.N.; Christensen, A.C. Evolution of a large, conserved, and syntenic gene family in insects. G3 2012, 2, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Dorer, D.R.; Ezekiel, D.H.; Christensen, A.C. The Triplo-lethal locus of Drosophila: Reexamination of mutants and discovery of a second-site suppressor. Genetics 1995, 141, 1037–1042. [Google Scholar] [PubMed]
- Andrade Lopez, J.M.; Lanno, S.M.; Auerbach, J.M.; Moskowitz, E.C.; Sligar, L.A.; Wittkopp, P.J.; Coolon, J.D. Genetic basis of octanoic acid resistance in Drosophila sechellia: Functional analysis of a fine-mapped region. Mol. Ecol. 2017, 26, 1148–1160. [Google Scholar] [CrossRef]
- Lanno, S.M.; Gregory, S.M.; Shimshak, S.J.; Alverson, M.K.; Chiu, K.; Feil, A.L.; Findley, M.G.; Forman, T.E.; Gordon, J.T.; Ho, J.; et al. Transcriptomic Analysis of Octanoic Acid Response in Drosophila sechellia Using RNA-Sequencing. G3 2017, 7, 3867–3873. [Google Scholar] [CrossRef] [PubMed]
- Lanno, S.M.; Shimshak, S.J.; Peyser, R.D.; Linde, S.C.; Coolon, J.D. Investigating the role of Osiris genes in Drosophila sechellia larval resistance to a host plant toxin. Ecol. Evol. 2019, 9, 1922–1933. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Song, M.; Hong, S. Negative regulation of the novel norpA(P24) suppressor, diehard4, in the endo-lysosomal trafficking underlies photoreceptor cell degeneration. PLoS Genet. 2013, 9, e1003559. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Scholl, A.; Yang, Y.; McBride, P.; Irwin, K.; Jiang, L. Tracheal expression of Osiris gene family in Drosophila. Gene Expr. Patterns GEP 2018, 28, 87–94. [Google Scholar] [CrossRef]
- Ando, T.; Sekine, S.; Inagaki, S.; Misaki, K.; Badel, L.; Moriya, H.; Sami, M.M.; Itakura, Y.; Chihara, T.; Kazama, H.; et al. Nanopore Formation in the Cuticle of an Insect Olfactory Sensillum. Curr. Biol. 2019, 29, 1512–1520. [Google Scholar] [CrossRef]
- Cornman, R.S.; Lopez, D.; Evans, J.D. Transcriptional response of honey bee larvae infected with the bacterial pathogen Paenibacillus larvae. PLoS ONE 2013, 8, e65424. [Google Scholar] [CrossRef]
- Suetsugu, Y.; Futahashi, R.; Kanamori, H.; Kadono-Okuda, K.; Sasanuma, S.; Narukawa, J.; Ajimura, M.; Jouraku, A.; Namiki, N.; Shimomura, M.; et al. Large scale full-length cDNA sequencing reveals a unique genomic landscape in a lepidopteran model insect, Bombyx mori. G3 2013, 3, 1481–1492. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Deng, H.M.; Zheng, S.C.; Huang, L.H.; Feng, Q.L.; Liu, L. Transcriptomic analysis of developmental features of Bombyx mori wing disc during metamorphosis. BMC Genom. 2014, 15, 820. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.R.; Morandin, C.; Noureddine, M.; Pant, S. Conserved roles of Osiris genes in insect development, polymorphism and protection. J. Evol. Biol. 2018, 31, 516–529. [Google Scholar] [CrossRef] [PubMed]
- Royer, C.; Briolay, J.; Garel, A.; Brouilly, P.; Sasanuma, S.; Sasanuma, M.; Shimomura, M.; Keime, C.; Gandrillon, O.; Huang, Y.; et al. Novel genes differentially expressed between posterior and median silk gland identified by SAGE-aided transcriptome analysis. Insect. Biochem. Mol. Biol. 2011, 41, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hu, W.; Cheng, T.; Peng, Z.; Mita, K.; Xia, Q. Osiris9a is a major component of silk fiber in lepidopteran insects. Insect. Biochem. Mol. Biol. 2017, 89, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Li, S.; Feng, Q. Advances in silkworm studies accelerated by the genome sequencing of Bombyx mori. Annu. Rev. Entomol. 2014, 59, 513–536. [Google Scholar] [CrossRef]
- Peng, Z.; Yang, X.; Liu, C.; Dong, Z.; Wang, F.; Wang, X.; Hu, W.; Zhang, X.; Zhao, P.; Xia, Q. Structural and Mechanical Properties of Silk from Different Instars of Bombyx mori. Biomacromolecules 2019, 20, 1203–1216. [Google Scholar] [CrossRef]
- Dong, Z.; Zhao, P.; Wang, C.; Zhang, Y.; Chen, J.; Wang, X.; Lin, Y.; Xia, Q. Comparative proteomics reveal diverse functions and dynamic changes of Bombyx mori silk proteins spun from different development stages. J. Proteome Res. 2013, 12, 5213–5222. [Google Scholar] [CrossRef]
- Xu, H.; Yuan, L.; Wang, F.; Wang, Y.; Wang, R.; Song, C.; Xia, Q.; Zhao, P. Overexpression of recombinant infectious bursal disease virus (IBDV) capsid protein VP2 in the middle silk gland of transgenic silkworm. Transgenic Res. 2014, 23, 809–816. [Google Scholar] [CrossRef]
- Wang, F.; Xu, H.; Yuan, L.; Ma, S.; Wang, Y.; Duan, X.; Duan, J.; Xiang, Z.; Xia, Q. An optimized sericin-1 expression system for mass-producing recombinant proteins in the middle silk glands of transgenic silkworms. Transgenic Res. 2013, 22, 925–938. [Google Scholar] [CrossRef]
- Peng, Z.; Liu, C.; Zhang, L.; Li, W.; Hu, W.; Ma, S.; Xia, Q. A Simple Method for the Cross-Section Area Determination of Single Profiled Fibers and Its Application. Microsc. Microanal. 2018, 24, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Ling, S.; Qin, Z.; Li, C.; Huang, W.; Kaplan, D.L.; Buehler, M.J. Polymorphic regenerated silk fibers assembled through bioinspired spinning. Nat. Commun. 2017, 8, 1387. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Y.; Liu, Q.; Chen, Q.; Xia, Q.; Zhao, P. In vivo effects of metal ions on conformation and mechanical performance of silkworm silks. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Bowen, C.H.; Dai, B.; Sargent, C.J.; Bai, W.; Ladiwala, P.; Feng, H.; Huang, W.; Kaplan, D.L.; Galazka, J.M.; Zhang, F. Recombinant Spidroins Fully Replicate Primary Mechanical Properties of Natural Spider Silk. Biomacromolecules 2018, 19, 3853–3860. [Google Scholar] [CrossRef]
- Lang, G.; Neugirg, B.R.; Kluge, D.; Fery, A.; Scheibel, T. Mechanical Testing of Engineered Spider Silk Filaments Provides Insights into Molecular Features on a Mesoscale. ACS Appl. Mater Interfaces 2017, 9, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Ashton, N.N.; Stewart, R.J. Aquatic caddisworm silk is solidified by environmental metal ions during the natural fiber-spinning process. FASEB J. 2019, 33, 572–583. [Google Scholar] [CrossRef]
- Cebe, P.; Partlow, B.P.; Kaplan, D.L.; Wurm, A.; Zhuravlev, E.; Schick, C. Silk I and Silk II studied by fast scanning calorimetry. Acta Biomater 2017, 55, 323–332. [Google Scholar] [CrossRef]
- McGill, M.; Holland, G.P.; Kaplan, D.L. Experimental Methods for Characterizing the Secondary Structure and Thermal Properties of Silk Proteins. Macromol. Rapid Commun. 2019, 40, e1800390. [Google Scholar] [CrossRef]
- Dong, A.; Huang, P.; Caughey, W.S. Protein secondary structures in water from second-derivative amide I infrared spectra. Biochemistry 1990, 29, 3303–3308. [Google Scholar] [CrossRef]
- Tomita, M.; Hino, R.; Ogawa, S.; Iizuka, M.; Adachi, T.; Shimizu, K.; Sotoshiro, H.; Yoshizato, K. A germline transgenic silkworm that secretes recombinant proteins in the sericin layer of cocoon. Transgenic Res. 2007, 16, 449–465. [Google Scholar] [CrossRef]
- Ogawa, S.; Tomita, M.; Shimizu, K.; Yoshizato, K. Generation of a transgenic silkworm that secretes recombinant proteins in the sericin layer of cocoon: Production of recombinant human serum albumin. J. Biotechnol. 2007, 128, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, R.; Luo, Q.; Qu, D.; Chen, T.; Yao, O.; Xu, H. Expression and characterization of recombinant human VEGF165 in the middle silk gland of transgenic silkworms. Transgenic Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Tanaka, K.; Arisaka, F.; Kimura, S.; Ohtomo, K.; Mizuno, S. Silk fibroin of Bombyx mori is secreted, assembling a high molecular mass elementary unit consisting of H-chain, L-chain, and P25, with a 6:6:1 molar ratio. J. Biol. Chem. 2000, 275, 40517–40528. [Google Scholar] [CrossRef] [PubMed]
- Horn, C.; Wimmer, E.A. A versatile vector set for animal transgenesis. Dev. Genes Evol. 2000, 210, 630–637. [Google Scholar] [CrossRef]
- Tamura, T.; Thibert, C.; Royer, C.; Kanda, T.; Abraham, E.; Kamba, M.; Komoto, N.; Thomas, J.L.; Mauchamp, B.; Chavancy, G.; et al. Germline transformation of the silkworm Bombyx mori L. using a piggyBac transposon-derived vector. Nat. Biotechnol. 2000, 18, 81–84. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Xie, K.; Yi, Q.; Chen, Q.; Wang, X.; Shen, H.; Xia, Q.; Zhao, P. Ca2+ and endoplasmic reticulum Ca2+-ATPase regulate the formation of silk fibers with favorable mechanical properties. J. Insect. Physiol. 2015, 73, 53–59. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, T.; Zhang, X.; Peng, Z.; Fan, Y.; Zhang, L.; Liu, C. Effects of Osiris9a on Silk Properties in Bombyx mori Determined by Transgenic Overexpression. Int. J. Mol. Sci. 2020, 21, 1888. https://doi.org/10.3390/ijms21051888
Cheng T, Zhang X, Peng Z, Fan Y, Zhang L, Liu C. Effects of Osiris9a on Silk Properties in Bombyx mori Determined by Transgenic Overexpression. International Journal of Molecular Sciences. 2020; 21(5):1888. https://doi.org/10.3390/ijms21051888
Chicago/Turabian StyleCheng, Tingcai, Xia Zhang, Zhangchuan Peng, Yinfeng Fan, Lin Zhang, and Chun Liu. 2020. "Effects of Osiris9a on Silk Properties in Bombyx mori Determined by Transgenic Overexpression" International Journal of Molecular Sciences 21, no. 5: 1888. https://doi.org/10.3390/ijms21051888
APA StyleCheng, T., Zhang, X., Peng, Z., Fan, Y., Zhang, L., & Liu, C. (2020). Effects of Osiris9a on Silk Properties in Bombyx mori Determined by Transgenic Overexpression. International Journal of Molecular Sciences, 21(5), 1888. https://doi.org/10.3390/ijms21051888