Genome-Wide Identification and Characterization of Actin-Depolymerizing Factor (ADF) Family Genes and Expression Analysis of Responses to Various Stresses in Zea Mays L.

 ,
,
Abstract
1. Introduction
2. Results
2.1. Identification of ADF Genes in Maize
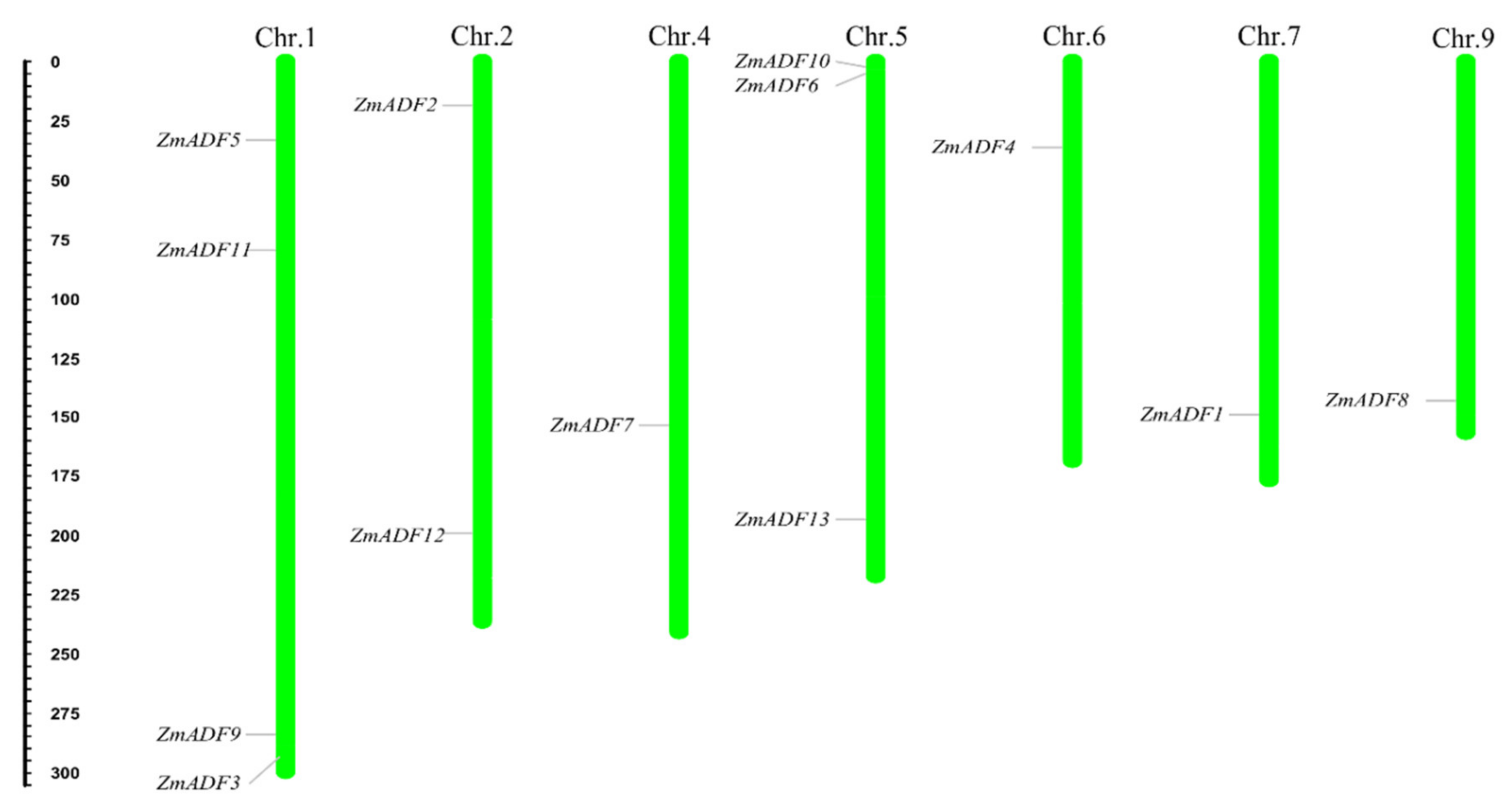
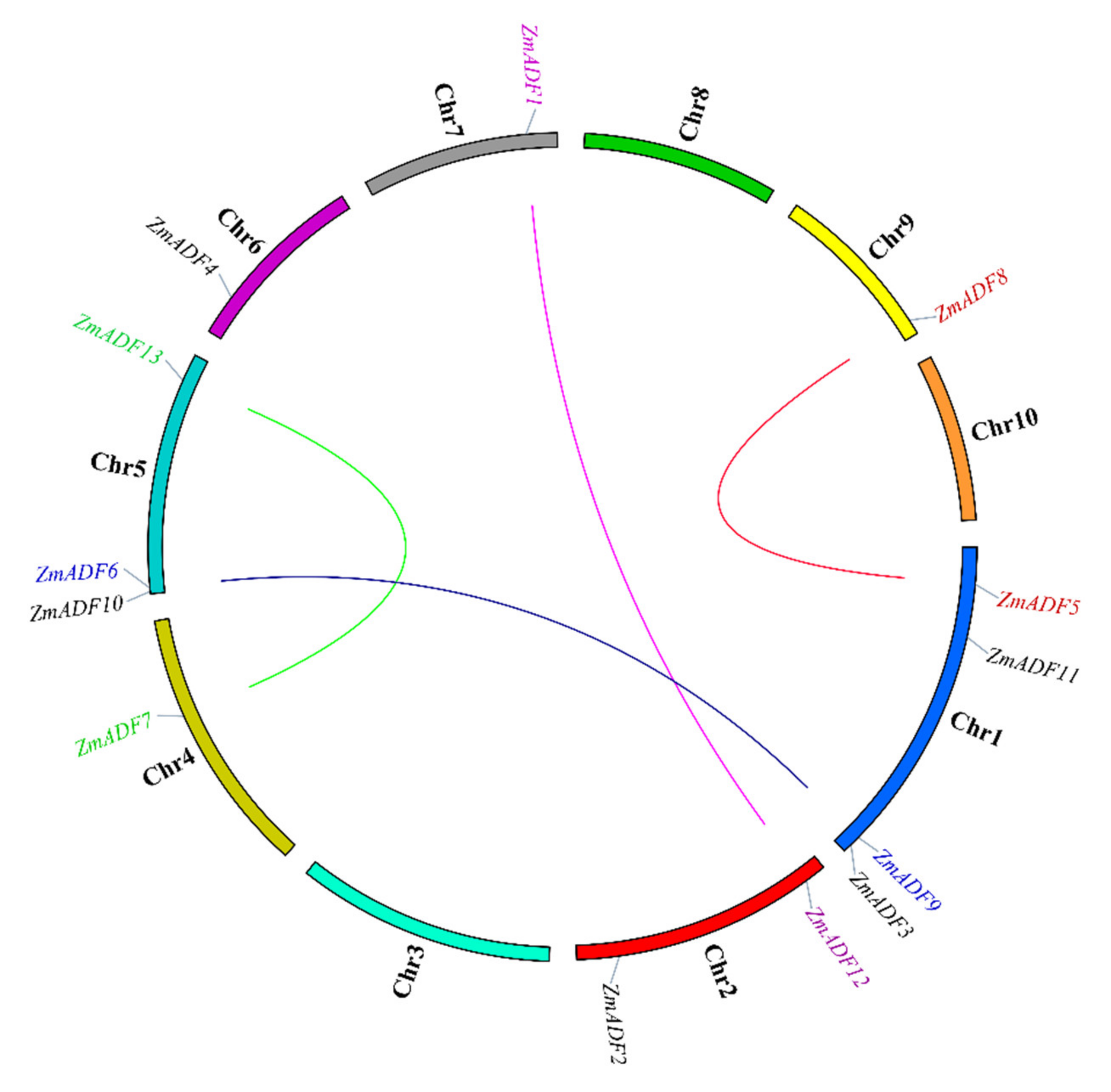
2.2. Chromosome Location and Gene Duplication of ADF Genes in Maize
2.3. Gene Structure of Identified ADF Genes in Maize
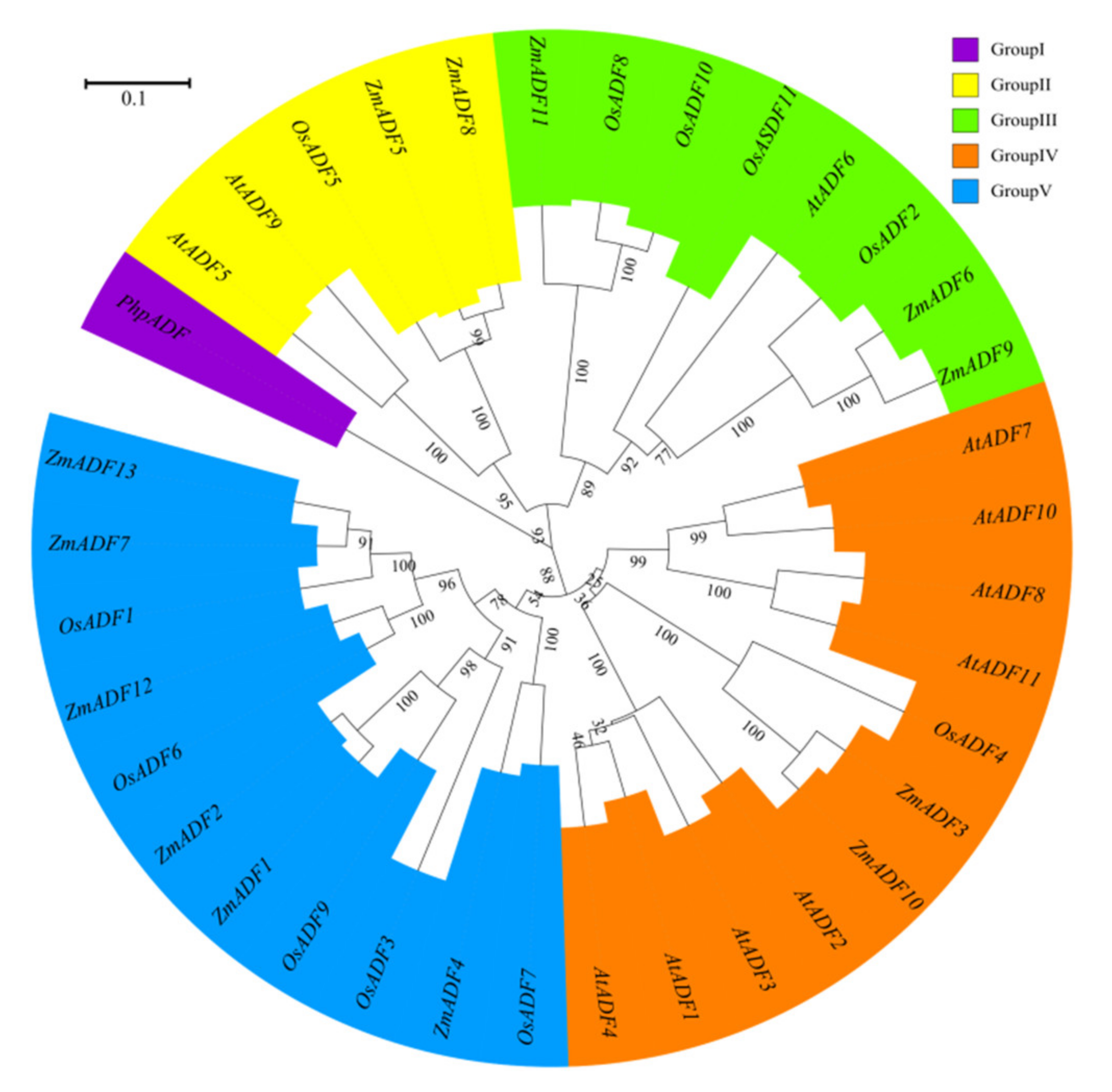
2.4. Phylogenetic Classification of ADF Genes in Monocots, Dicots, and Mosses
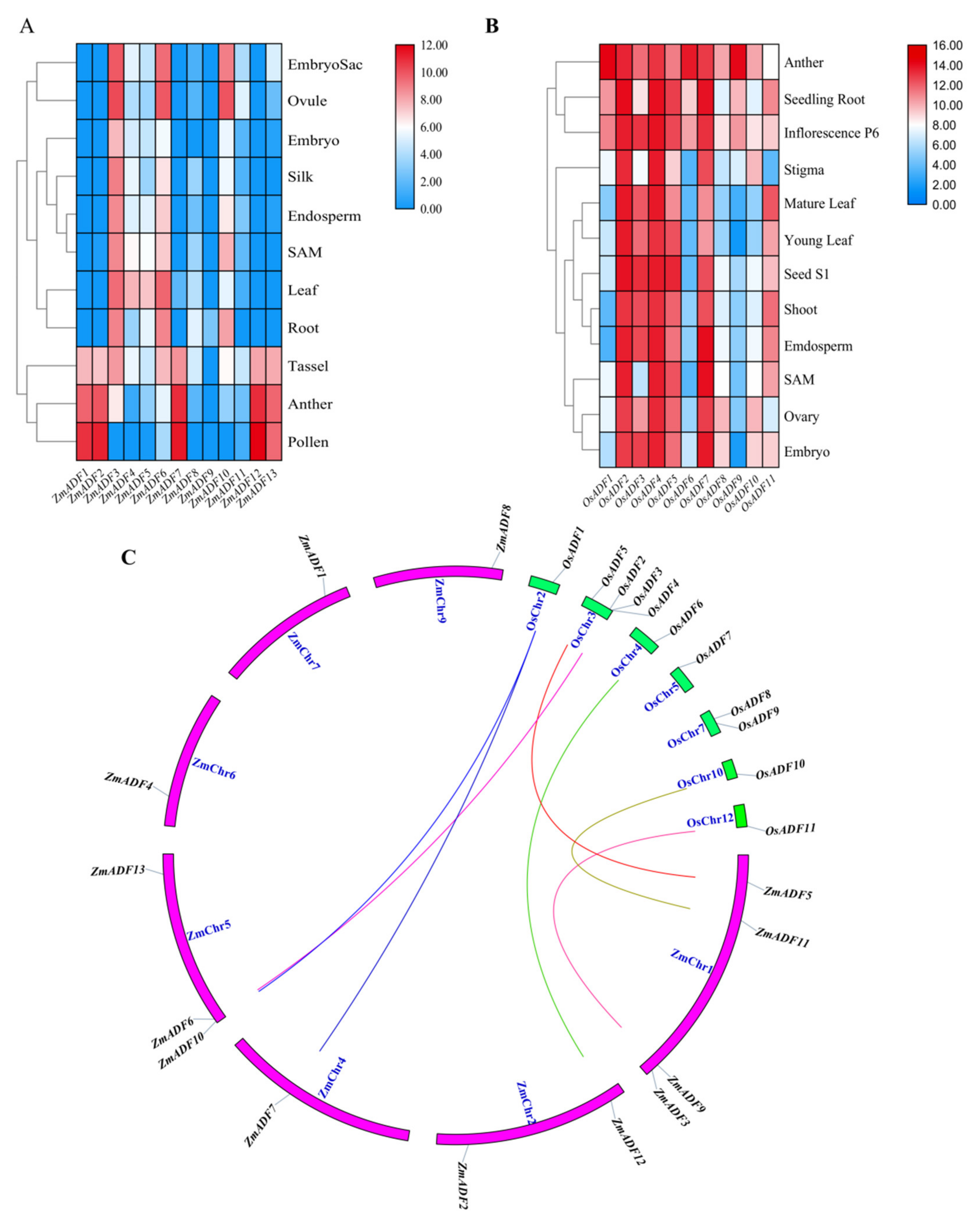
2.5. Expression Profiles of ZmADFs in Different Tissues and Orthologous Genes in Oryza sativa
2.6. Expression Patterns of ZmADFs under Different Abiotic Stresses
3. Discussion
3.1. Classification and Phylogenetic Analysis of the ADF Proteins among Monocots, Dicots and Mosses Indicate their Structural and Functional Similarities
3.2. To Protect Or Promote the Reproductive Development, which is more Important?
4. Materials and Methods
4.1. Identification and Analysis of ADF Family Genes
4.2. Phylogenetic Analysis of Maize ADF Genes
4.3. Analysis of Chromosome Location and gene Duplication
4.4. Gene Structure Analysis
4.5. Expression Pattern Analysis in Different Tissues of ZmADF Genes and Orthologous Genes in Oryza sativa
4.6. Plant Materials and qPCR Expression Analysis under Diverse Abiotic Stresses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADF | Actin-depolymerizing factor |
| SAM | Shoot apical meristem |
| GPA | Green peach aphid |
| ABP | Actin-binding protein |
| ORF | Open reading frame |
| CDS | Coding sequences |
| ROS | Reactive oxygen species |
| GRAVY | Grand average of hydropathy |
| PI | Iso-electric point |
| Mw | Molecular weight |
| HMM | Hidden Markov Model |
| PMCs | Pollen mother cells; 2,4-D: 2,4-dichlorophenoxyacetic acid |
| ABA | Abscisic acid |
| GA3 | Gibberellic acid |
| IAA | Indole-3-acetic acid |
| JA | Jasmonic acid |
| HR | Hypersensitive response |
| qRT-PCR | Quantitative real-time PCR |
References
- Huang, X.; Xu, Y.; Guo, G.; Lin, Q.; Ye, Z.; Yuan, L.; Sun, Z.; Ni, W. Molecular characterization of an actin depolymerizing factor from Cryptocaryon Irrit. Parasitology 2013, 140, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Makioka, A.; Kumagai, M.; Hiranuka, K.; Kobayashi, S.; Takeuchi, T. Entamoeba invadens: Identification of ADF/cofilin and their expression analysis in relation to encystation and excystation. Exp. Parasitol. 2011, 127, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, B.W.; Bamburg, J.R. ADF/cofilin: A functional node in cell biology. Trends Cell Biol. 2010, 20, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Nishida, E.; Maekawa, S.; Sakai, H. Cofilin, a protein in porcine brain that binds to actin filaments and inhibits their interactions with myosin and tropomyosin. Biochemistry 1984, 23, 5307–5313. [Google Scholar] [CrossRef] [PubMed]
- Inada, N. Plant actin depolymerizing factor: Actin microfilament disassembly and more. J. Plant Res. 2017, 130, 227–238. [Google Scholar] [CrossRef]
- Staiger, C.J.; Gibbon, B.C.; Kovar, D.R.; Zonia, L.E. Profilin and actin-depolymerizing factor: Modulators of actin organization in plants. Trends Plant Sci. 1997, 2, 275–281. [Google Scholar] [CrossRef]
- Feng, Y.; Liu, Q.; Xue, Q. Comparative study of rice and Arabidopsis actin-depolymerizing factors gene families. J. Plant Physiol. 2006, 163, 69–79. [Google Scholar] [CrossRef]
- Ruzicka, D.R.; Kandasamy, M.K.; McKinney, E.C.; Burgos-Rivera, B.; Meagher, R.B. The ancient subclasses of Arabidopsis Actin Depolymerizing Factor genes exhibit novel and differential expression. Plant J. Cell Mol. Biol. 2007, 52, 460–472. [Google Scholar] [CrossRef]
- Nan, Q.; Qian, D.; Niu, Y.; He, Y.; Tong, S.; Niu, Z.; Ma, J.; Yang, Y.; An, L.; Wan, D.; et al. Plant Actin-Depolymerizing Factors Possess Opposing Biochemical Properties Arising from Key Amino Acid Changes throughout Evolution. Plant Cell 2017, 29, 395–408. [Google Scholar] [CrossRef]
- Roy-Zokan, E.M.; Dyer, K.A.; Meagher, R.B. Phylogenetic Patterns of Codon Evolution in the ACTIN-DEPOLYMERIZING FACTOR/COFILIN (ADF/CFL) Gene Family. PLoS ONE 2015, 10, e0145917. [Google Scholar] [CrossRef]
- Khatun, K.; Robin, A.H.; Park, J.I.; Kim, C.K.; Lim, K.B.; Kim, M.B.; Lee, D.J.; Nou, I.S.; Chung, M.Y. Genome-Wide Identification, Characterization and Expression Profiling of ADF Family Genes in Solanum lycopersicum L. Genes 2016, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.H.; Kost, B.; Xia, G.; Chua, N.H. Molecular identification and characterization of the Arabidopsis AtADF1, AtADF5 and AtADF6 genes. Plant Mol. Biol. 2001, 45, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Lopez, I.; Anthony, R.G.; Maciver, S.K.; Jiang, C.J.; Khan, S.; Weeds, A.G.; Hussey, P.J. Pollen specific expression of maize genes encoding actin depolymerizing factor-like proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 7415–7420. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Rivera, B.; Ruzicka, D.R.; Deal, R.B.; McKinney, E.C.; King-Reid, L.; Meagher, R.B. ACTIN DEPOLYMERIZING FACTOR9 controls development and gene expression in Arabidopsis. Plant Mol. Biol. 2008, 68, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Allwood, E.G.; Anthony, R.G.; Smertenko, A.P.; Reichelt, S.; Drobak, B.K.; Doonan, J.H.; Weeds, A.G.; Hussey, P.J. Regulation of the Pollen-Specific Actin-Depolymerizing Factor LlADF1. Plant Cell 2002, 14, 2915–2927. [Google Scholar] [CrossRef]
- Bou Daher, F.; van Oostende, C.; Geitmann, A. Spatial and temporal expression of actin depolymerizing factors ADF7 and ADF10 during male gametophyte development in Arabidopsis thaliana. Plant Cell Physiol. 2011, 52, 1177–1192. [Google Scholar] [CrossRef]
- Chi, J.; Wang, X.; Zhou, H.; Zhang, G.; Sun, Y.; Li, Z.; Ma, Z. Molecular cloning and characterization of the actin-depolymerizing factor gene in Gossypium barbadense. Genes Genet. Syst. 2008, 83, 383–391. [Google Scholar] [CrossRef]
- Wang, H.Y.; Wang, J.; Gao, P.; Jiao, G.L.; Zhao, P.M.; Li, Y.; Wang, G.L.; Xia, G.X. Down-regulation of GhADF1 gene expression affects cotton fibre properties. Plant Biotechnol. J. 2009, 7, 13–23. [Google Scholar] [CrossRef]
- Clement, M.; Ketelaar, T.; Rodiuc, N.; Banora, M.Y.; Smertenko, A.; Engler, G.; Abad, P.; Hussey, P.J.; de Almeida Engler, J. Actin-depolymerizing factor2-mediated actin dynamics are essential for root-knot nematode infection of Arabidopsis. Plant Cell 2009, 21, 2963–2979. [Google Scholar] [CrossRef]
- Tian, M.; Chaudhry, F.; Ruzicka, D.R.; Meagher, R.B.; Staiger, C.J.; Day, B. Arabidopsis actin-depolymerizing factor AtADF4 mediates defense signal transduction triggered by the Pseudomonas syringae effector AvrPphB. Plant Physiol. 2009, 150, 815–824. [Google Scholar] [CrossRef]
- Fu, Y.; Duan, X.; Tang, C.; Li, X.; Voegele, R.T.; Wang, X.; Wei, G.; Kang, Z. TaADF7, an actin-depolymerizing factor; contributes to wheat resistance against Puccinia striiformis f. sp. tritici. Plant J. 2014, 78, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Deng, L.; Chang, D.; Chen, S.; Wang, X.; Kang, Z. TaADF3, an Actin-Depolymerizing Factor, Negatively Modulates Wheat Resistance Against Puccinia striiformis. Front. Plant Sci. 2015, 6, 1214. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Hua, Y.; Wang, J.; Huo, Y.; Shimono, M.; Day, B.; Ma, Q. TaADF4, an actin-depolymerizing factor from wheat, is required for resistance to the stripe rust pathogen Puccinia striiformis f. sp. tritici. Plant J. 2017, 89, 1210–1224. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.C.; Huang, W.L.; Hong, C.Y.; Lur, H.S.; Chang, M.C. Comprehensive analysis of differentially expressed rice actin depolymerizing factor gene family and heterologous overexpression of OsADF3 confers Arabidopsis Thaliana drought tolerance. Rice (N Y) 2012, 5, 33. [Google Scholar] [CrossRef]
- Dong, C.H.; Tang, W.P.; Liu, J.Y. Arabidopsis AtADF1 is functionally affected by mutations on actin binding sites. J. Integr. Plant Biol. 2013, 55, 250–261. [Google Scholar] [CrossRef]
- Dong, C.H.; Xia, G.X.; Hong, Y.; Ramachandran, S.; Kost, B.; Chua, N.H. ADF proteins are involved in the control of flowering and regulate F-actin organization, cell expansion, and organ growth in Arabidopsis. Plant Cell 2001, 13, 1333–1346. [Google Scholar] [CrossRef]
- Henty, J.L.; Bledsoe, S.W.; Khurana, P.; Meagher, R.B.; Day, B.; Blanchoin, L.; Staiger, C.J. Arabidopsis Actin Depolymerizing Factor4 Modulates the Stochastic Dynamic Behavior of Actin Filaments in the Cortical Array of Epidermal Cells. Plant Cell 2011, 23, 3711–3726. [Google Scholar] [CrossRef]
- Inada, N.; Higaki, T.; Hasezawa, S. Nuclear Function of Subclass I Actin-Depolymerizing Factor Contributes to Susceptibility in Arabidopsis to an Adapted Powdery Mildew Fungus. Plant Physiol. 2016, 170, 1420–1434. [Google Scholar] [CrossRef]
- Mondal, H.A.; Louis, J.; Archer, L.; Patel, M.; Nalam, V.J.; Sarowar, S.; Sivapalan, V.; Root, D.D.; Shah, J. Arabidopsis ACTIN-DEPOLYMERIZING FACTOR3 is required for controlling aphid feeding from the phloem. Plant Physiol. 2018, 176, 879–890. [Google Scholar] [CrossRef]
- Zhu, J.; Nan, Q.; Qin, T.; Qian, D.; Mao, T.; Yuan, S.; Wu, X.; Niu, Y.; Bai, Q.; An, L.; et al. Higher-Ordered Actin Structures Remodeled by Arabidopsis ACTIN-DEPOLYMERIZING FACTOR5 Are Important for Pollen Germination and Pollen Tube Growth. Mol. Plant 2017, 10, 1065–1081. [Google Scholar] [CrossRef]
- Zheng, Y.; Xie, Y.; Jiang, Y.; Qu, X.; Huang, S. Arabidopsis actin-depolymerizing factor7 severs actin filaments and regulates actin cable turnover to promote normal pollen tube growth. Plant Cell 2013, 25, 3405–3423. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, F.; Carpentier, E.; Cope, M.J.; Monroy, A.F.; Sarhan, F. Regulation of a wheat actin-depolymerizing factor during cold acclimation. Plant Physiol. 2001, 125, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.J.; Weeds, A.G.; Khan, S.; Hussey, P.J. F-actin and G-actin binding are uncoupled by mutation of conserved tyrosine residues in maize actin depolymerizing factor (ZmADF). Proc. Natl. Acad. Sci. USA 1997, 94, 9973–9978. [Google Scholar] [CrossRef] [PubMed]
- Smertenko, A.P.; Allwood, E.G.; Khan, S.; Jiang, C.J.; Maciver, S.K.; Weeds, A.G.; Hussey, P.J. Interaction of pollen-specific actin-depolymerizing factor with actin. Plant J. 2001, 25, 203–212. [Google Scholar] [CrossRef]
- Soltis, P.S.; Soltis, D.E.; Savolainen, V.; Crane, P.R.; Barraclough, T.G. Rate heterogeneity among lineages of tracheophytes: Integration of molecular and fossil data and evidence for molecular living fossils. Proc. Nat. Acad. Sci. USA 2002, 99, 4430–4435. [Google Scholar] [CrossRef]
- Christensen, H.E.; Ramachandran, S.; Tan, C.T.; Surana, U.; Dong, C.H.; Chua, N.H. Arabidopsis profilins are functionally similar to yeast profilins. identification of a vascular bundle-specific profilin and a pollen-specific profilin. Plant J. 1996, 10, 269–279. [Google Scholar] [CrossRef]
- Huang, S.; McDowell, J.M.; Weise, M.J.; Meagher, R.B. The Arabidopsis profilin gene family. Evidence for an ancient split between constitutive and pollen-specific profilin genes. Plant Physiol. 1996, 111, 115–126. [Google Scholar] [CrossRef]
- Kandasamy, M.K.; McKinney, E.C.; Meagher, R.B. The late pollen-specific actins in angiosperms. Plant J. 1999, 18, 681–691. [Google Scholar] [CrossRef]
- Zhai, L.; Sun, W.; Zhang, K.; Jia, H.; Liu, L.; Liu, Z.; Teng, F.; Zhang, Z. Identification and characterization of Argonaute gene family and meiosis-enriched Argonaute during sporogenesis in maize. J. Integr. Plant Biol. 2014, 56, 1042–1052. [Google Scholar] [CrossRef]
- Sun, W.; Xiang, X.; Zhai, L.; Zhang, D.; Cao, Z.; Liu, L.; Zhang, Z. AGO18b negatively regulates determinacy of spikelet meristems on the tassel central spike in maize. J. Integr. Plant Biol. 2018, 60, 65–78. [Google Scholar] [CrossRef]
- Zhai, L.; Teng, F.; Zheng, K.; Xiao, J.; Deng, W.; Sun, W. Expression analysis of Argonaute genes in maize (Zea mays L.) in response to abiotic stress. Hereditas 2019, 156, 27. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Guo, Y.; Wu, H.; Li, X.; Li, Q.; Zhao, X.; Duan, X.; An, Y.; Lv, W.; An, H. Identification and expression of GRAS family genes in maize (Zea mays L.). PLoS ONE 2017, 12, e0185418. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆Ct Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Location | Accession Number | Open Reading Frame Length (bp) | Amino Acids (aa) | Isoelectric Point | Molecular Weight (KD)(Kda) | GRAVY |
|---|---|---|---|---|---|---|---|
| ZmADF1 | 7:148879558..148880837(F) | GRMZM2G117603 | 432 | 144 | 6.32 | 16.54 | −0.586 |
| ZmADF2 | 2:199484159..199485220(R) | GRMZM2G097122 | 417 | 139 | 5.57 | 16.08 | −0.642 |
| ZmADF3 | 1:293162401..293166106(F) | GRMZM2G060702 | 414 | 138 | 12.55 | 15.63 | −1.231 |
| ZmADF4 | 6:35896131..35897803(R) | GRMZM2G037140 | 417 | 139 | 7.66 | 15.86 | −0.369 |
| ZmADF5 | 1:32867955..32871278(R) | GRMZM2G077942 | 429 | 143 | 8.41 | 16.41 | −0.287 |
| ZmADF6 | 5:5245812..5249187(F) | GRMZM2G130678 | 435 | 145 | 6.15 | 16.83 | −0.471 |
| ZmADF7 | 4:153268732..153270773(R) | GRMZM2G463471 | 417 | 139 | 6.31 | 15.86 | −0.316 |
| ZmADF8 | 9:143174940..143177192(F) | GRMZM2G147775 | 516 | 172 | 9.51 | 20.04 | −0.505 |
| ZmADF9 | 1:284216333..284217771(F) | GRMZM2G108807 | 420 | 140 | 7.78 | 16.41 | −0.469 |
| ZmADF10 | 5:2362323..2364777(R) | GRMZM2G002825 | 417 | 139 | 5.47 | 15.91 | −0.485 |
| ZmADF11 | 1:80054390..80058323(R) | GRMZM2G064875 | 369 | 123 | 5.6 | 14.38 | −0.667 |
| ZmADF12 | 2:18815692..18816950(F) | GRMZM2G071327 | 417 | 139 | 5.27 | 15.98 | −0.570 |
| ZmADF13 | 5:193084737..193087687(R) | GRMZM2G015127 | 417 | 139 | 7.56 | 15.89 | −0.271 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.; Sun, W.; Ren, J.; Yang, R.; Fan, J.; Li, Y.; Wang, X.; Joseph, S.; Deng, W.; Zhai, L. Genome-Wide Identification and Characterization of Actin-Depolymerizing Factor (ADF) Family Genes and Expression Analysis of Responses to Various Stresses in Zea Mays L. Int. J. Mol. Sci. 2020, 21, 1751. https://doi.org/10.3390/ijms21051751
Huang J, Sun W, Ren J, Yang R, Fan J, Li Y, Wang X, Joseph S, Deng W, Zhai L. Genome-Wide Identification and Characterization of Actin-Depolymerizing Factor (ADF) Family Genes and Expression Analysis of Responses to Various Stresses in Zea Mays L. International Journal of Molecular Sciences. 2020; 21(5):1751. https://doi.org/10.3390/ijms21051751
Chicago/Turabian StyleHuang, Jun, Wei Sun, Jiaxin Ren, Ruichun Yang, Jingsheng Fan, Yunfeng Li, Xin Wang, Shija Joseph, Wenbin Deng, and Lihong Zhai. 2020. "Genome-Wide Identification and Characterization of Actin-Depolymerizing Factor (ADF) Family Genes and Expression Analysis of Responses to Various Stresses in Zea Mays L." International Journal of Molecular Sciences 21, no. 5: 1751. https://doi.org/10.3390/ijms21051751
APA StyleHuang, J., Sun, W., Ren, J., Yang, R., Fan, J., Li, Y., Wang, X., Joseph, S., Deng, W., & Zhai, L. (2020). Genome-Wide Identification and Characterization of Actin-Depolymerizing Factor (ADF) Family Genes and Expression Analysis of Responses to Various Stresses in Zea Mays L. International Journal of Molecular Sciences, 21(5), 1751. https://doi.org/10.3390/ijms21051751