Elucidating the Possible Involvement of Maize Aquaporins in the Plant Boron Transport and Homeostasis Mediated by Rhizophagus irregularis under Drought Stress Conditions



Abstract
1. Introduction
2. Results
2.1. Plant Biomass and Symbiotic Development
2.2. Stomatal Conductance (gs)
2.3. Chlorophyll Content and Efficiency of Photosystem II
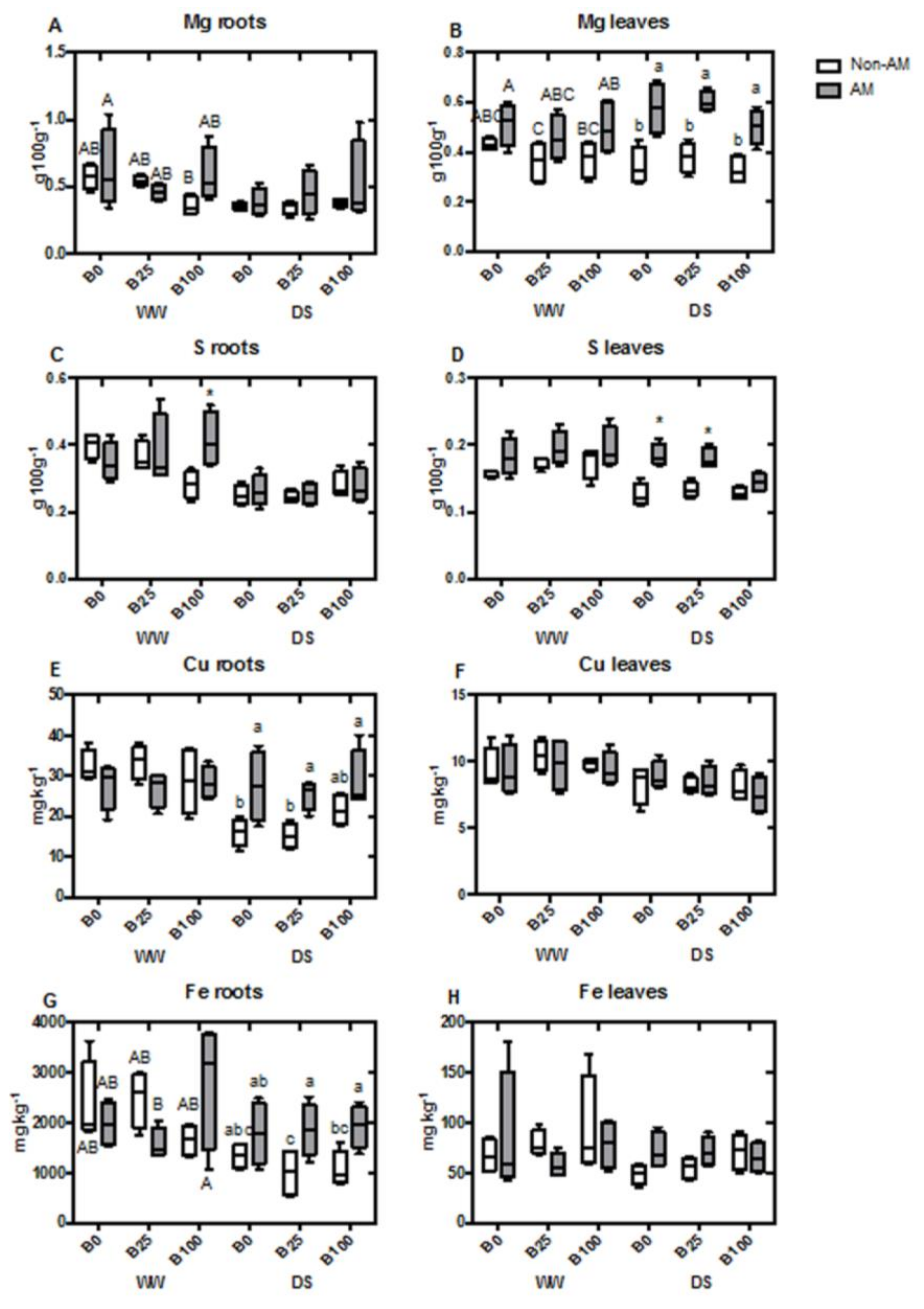
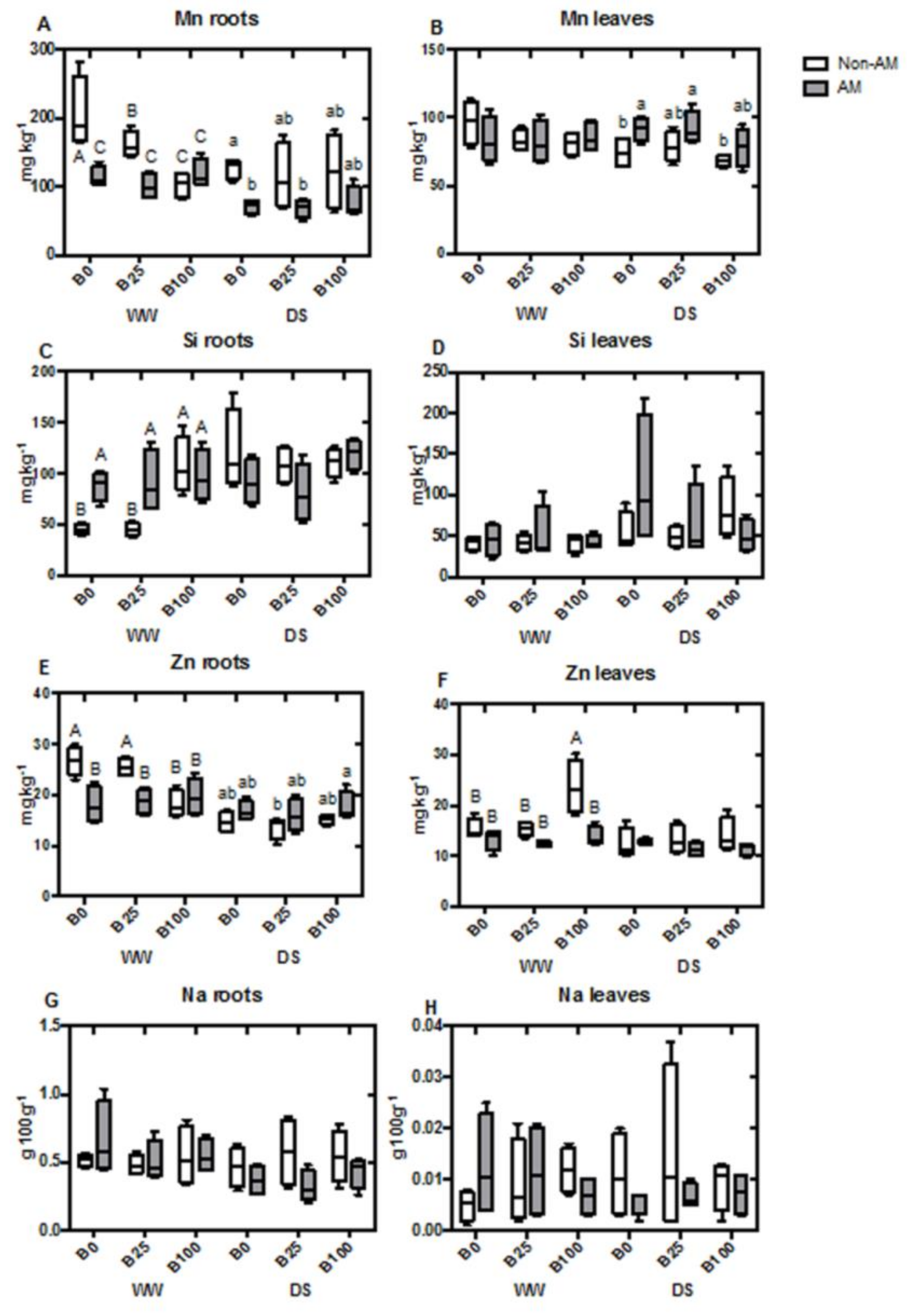
2.4. Mineral Content of Roots and Shoots
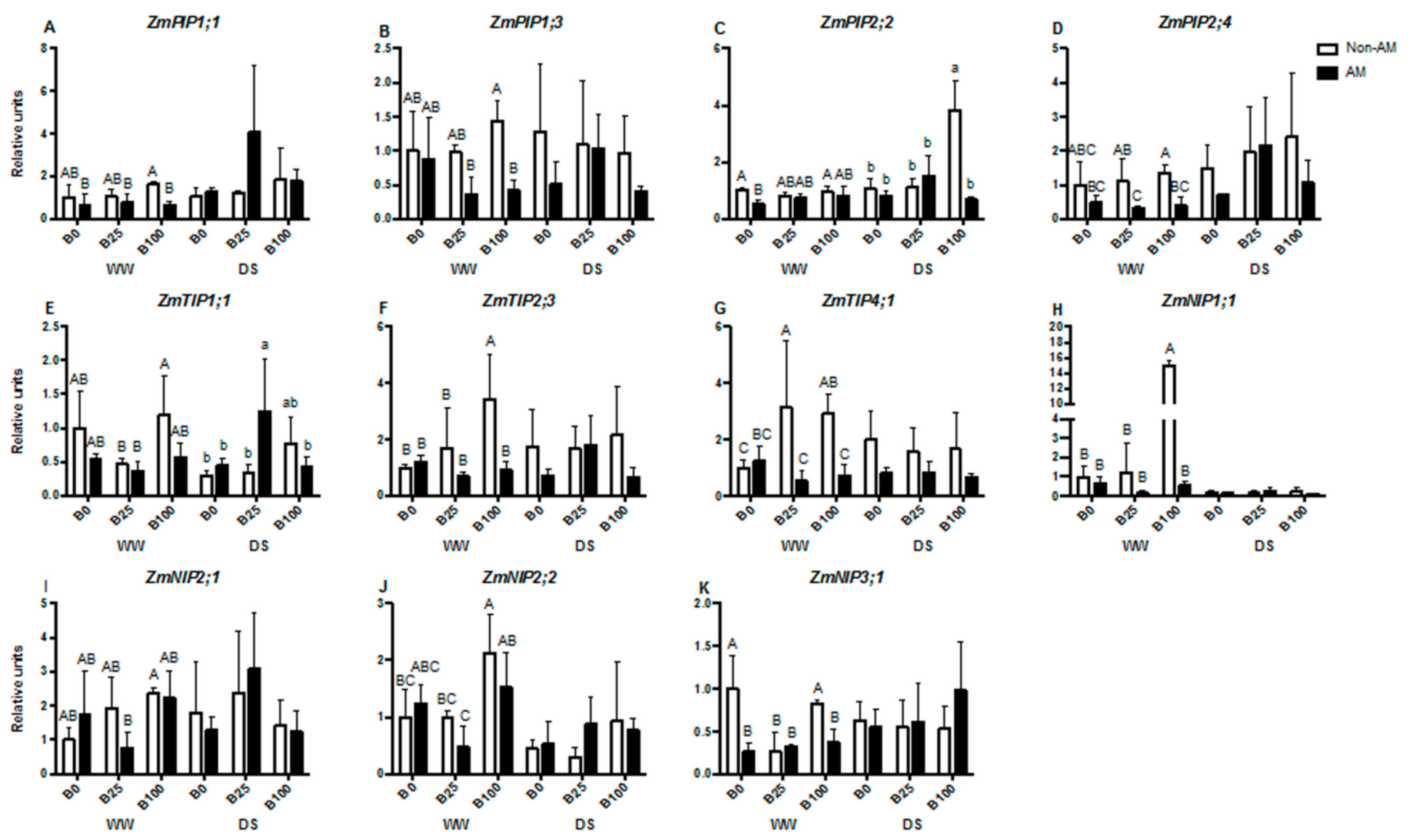
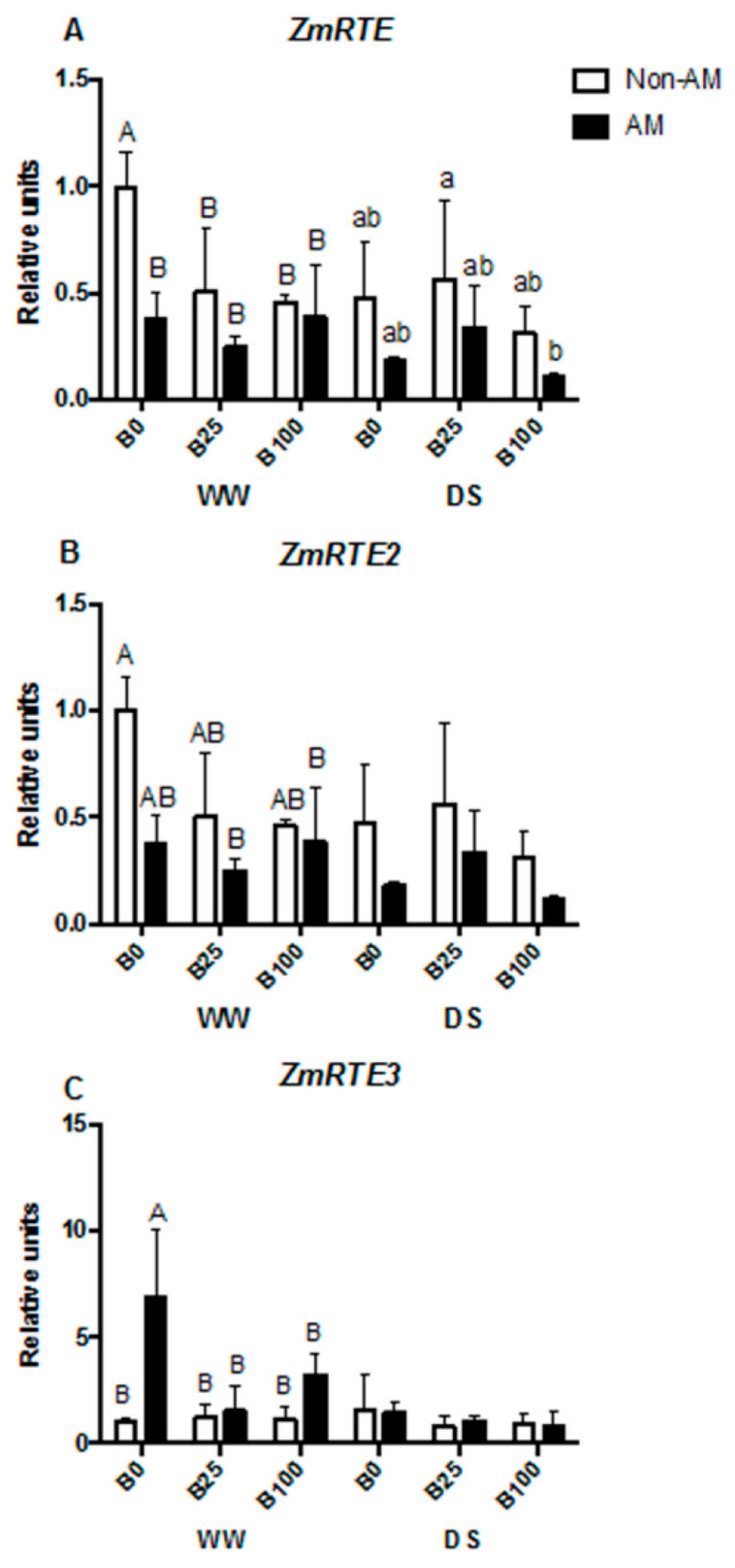
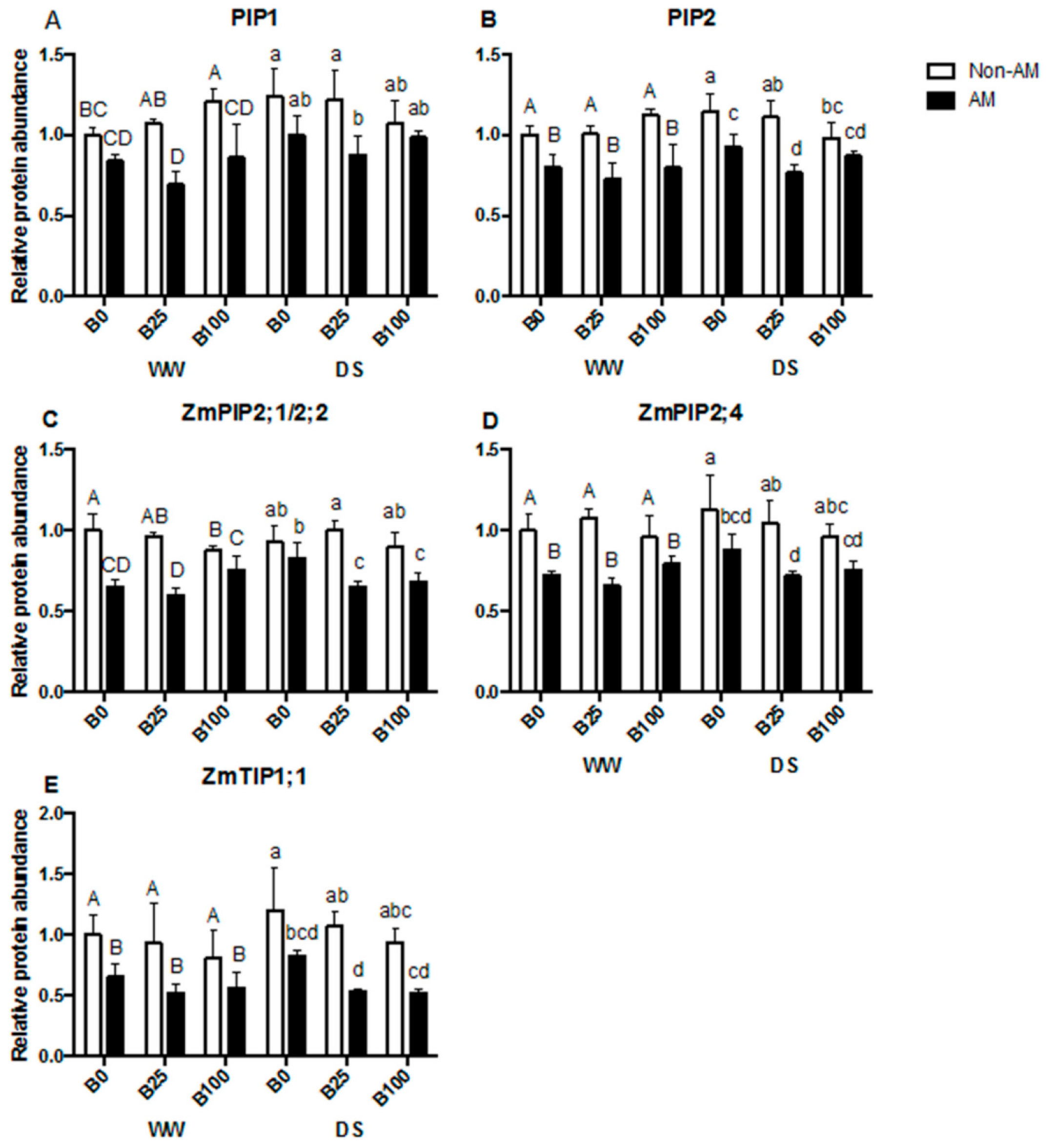
2.5. mRNA Relative Transcript Abundance of Aquaporins
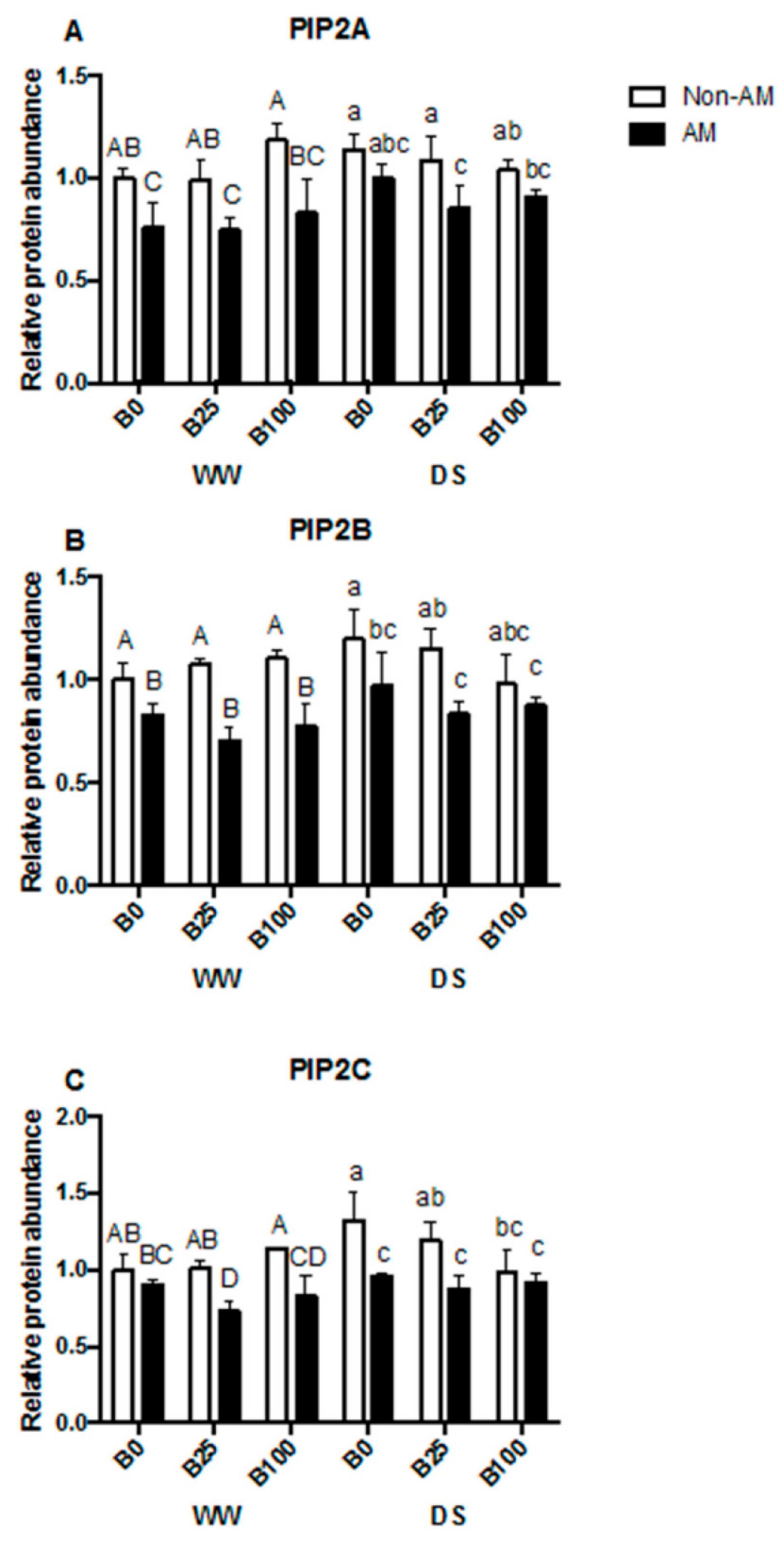
2.6. Aquaporin Protein Accumulation and PIP2s Phosphorylation Status
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Soil and Biological Materials
4.3. Growing Conditions
4.4. Parameters Measured
4.4.1. Biomass Production
4.4.2. Symbiotic Development
4.4.3. Stomatal Conductance
4.4.4. Leaf Chlorophyll Content
4.4.5. Photosynthetic Efficiency
4.4.6. Mineral Analysis
4.4.7. RT-qPCR
4.4.8. Aquaporins Abundance and PIP2s Phosphorylation Status
4.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kobayashi, M.; Matoh, T.; Azuma, J. Two Chains of Rhamnogalacturonan II Are Cross-Linked by Borate-Diol Ester Bonds in Higher Plant Cell Walls. Plant Physiol. 1996, 110, 1017–1020. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, M.A.; Ishii, T.; Albersheim, P.; Darvill, A.G. RHAMNOGALACTURONAN II: Structure and Function of a Borate Cross-Linked Cell Wall Pectic Polysaccharide. Annu. Rev. Plant Biol. 2004, 55, 109–139. [Google Scholar] [CrossRef]
- Voxeur, A.; Fry, S.C. Glycosylinositol phosphorylceramides from Rosa cell cultures are boron-bridged in the plasma membrane and form complexes with rhamnogalacturonan II. Plant J. 2014, 79, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Shireen, F.; Nawaz, M.A.; Chen, C.; Zhang, Q.; Zheng, Z.; Sohail, H.; Sun, J.; Cao, H.; Yuan, H.; Bie, Z. Boron: Functions and Approaches to Enhance Its Availability in Plants for Sustainable Agriculture. Int. J. Mol. Sci. 2018, 19, 1856. [Google Scholar] [CrossRef] [PubMed]
- Yoshinari, A.; Takano, J. Insights into the Mechanisms Underlying Boron Homeostasis in Plants. Front. Plant Sci. 2017, 8, 1951. [Google Scholar] [CrossRef] [PubMed]
- Nable, R.O.; Bañuelos, G.S.; Paull, J.G. Boron toxicity. Plant Soil 1997, 193, 181–198. [Google Scholar] [CrossRef]
- Simón-Grao, S.; Nieves, M.; Martínez-Nicolás, J.J.; Alfosea-Simón, M.; Cámara-Zapata, J.M.; Fernández-Zapata, J.C.; García-Sánchez, F. Arbuscular mycorrhizal symbiosis improves tolerance of Carrizo citrange to excess boron supply by reducing leaf B concentration and toxicity in the leaves and roots. Ecotoxicol. Environ. Saf. 2019, 173, 322–330. [Google Scholar] [CrossRef]
- Miwa, K.; Kamiya, T.; Fujiwara, T. Homeostasis of the structurally important micronutrients, B and Si. Curr. Opin. Plant Biol. 2009, 12, 307–311. [Google Scholar] [CrossRef]
- Stangoulis, J.C.R.; Reid, R.J.; Brown, P.H.; Graham, R.D. Kinetic analysis of boron transport in Chara. Planta 2001, 213, 142–146. [Google Scholar] [CrossRef]
- Takano, J.; Wada, M.; Ludewig, U.; Schaaf, G.; Von Wirén, N.; Fujiwara, T. The Arabidopsis Major Intrinsic Protein NIP5;1 Is Essential for Efficient Boron Uptake and Plant Development under Boron Limitation. Plant Cell 2006, 18, 1498–1509. [Google Scholar] [CrossRef]
- Dannel, F.; Pfeffer, H.; Römheld, V. Characterisation of root boron pools, boron uptake and translocation in sunflower using the stable isotopes 10B and 11B. Aust. J. Plant Physiol. 2000, 27, 397–405. [Google Scholar]
- Maurel, C.; Verdoucq, L.; Luu, D.-T.; Santoni, V. Plant aquaporins: Membrane channels with multiple integrated functions. Annu. Rev. Plant Biol. 2008, 59, 595–624. [Google Scholar] [CrossRef] [PubMed]
- Fox, A.R.; Maistriaux, L.C.; Chaumont, F. Toward understanding of the high number of plant aquaporin isoforms and multiple regulation mechanisms. Plant Sci. 2017, 264, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Takano, J.; Noguchi, K.; Yasomuri, M.; Kobayashi, M.; Gajdos, Z.; Miwa, K.; Hayashi, H.; Yoneyama, T.; Fujiwara, T. Arabidopsis boron transporter for xylem loading. Nature 2002, 420, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Shimotohno, A.; Sotta, N.; Sato, T.; De Ruvo, M.; Marée, A.F.M.; Grieneisen, V.A.; Fujiwara, T. Mathematical modeling and experimental validation of the spatial distribution of boron in the root of Arabidopsis thaliana identify high boron accumulation in the tip and predict a distinct root tip uptake function. Plant Cell Physiol. 2015, 56, 620–630. [Google Scholar] [CrossRef]
- Hanaoka, H.; Uraguchi, S.; Takano, J.; Tanaka, M.; Fujiwara, T. OsNIP3;1, a rice boric acid channel, regulates boron distribution and is essential for growth under boron-deficient conditions. Plant J. 2014, 78, 890–902. [Google Scholar] [CrossRef]
- Sutton, T.; Baumann, U.; Hayes, J.; Collins, N.C.; Shi, B.; Schnurbusch, T.; Hay, A.; Mayo, G.; Pallotta, M.; Tester, M.; et al. Boron-Toxicity Tolerance in Transporter Amplification. Science 2007, 318, 1446–1449. [Google Scholar] [CrossRef]
- Schnurbusch, T.; Hayes, J.; Hrmova, M.; Baumann, U.; Ramesh, S.A.; Tyerman, S.D.; Langridge, P.; Sutton, T. Boron Toxicity Tolerance in Barley through Reduced Expression of the Multifunctional Aquaporin HvNIP2;1. Plant Physiol. 2010, 153, 1706–1715. [Google Scholar] [CrossRef]
- Leaungthitikanchana, S.; Fujibe, T.; Tanaka, M.; Wang, S.; Sotta, N.; Takano, J.; Fujiwara, T. Differential expression of three BOR1 genes corresponding to different genomes in response to boron conditions in hexaploid wheat (Triticum aestivum L.). Plant Cell Physiol. 2013, 54, 1056–1063. [Google Scholar] [CrossRef]
- Porcel, R.; Bustamante, A.; Ros, R.; Serrano, R.; Mulet Salort, J.M. BvCOLD1: A novel aquaporin from sugar beet (Beta vulgaris L.) involved in boron homeostasis and abiotic stress. Plant Cell Environ. 2018, 2014, 1–14. [Google Scholar] [CrossRef]
- Chatterjee, M.; Liu, Q.; Menello, C.; Galli, M.; Gallavotti, A. The combined action of duplicated boron transporters is required for maize growth in boron-deficient conditions. Genetics 2017, 206, 2041–2051. [Google Scholar] [CrossRef] [PubMed]
- Lordkaew, S.; Dell, B.; Jamjod, S.; Rerkasem, B. Boron deficiency in maize. Plant Soil 2011, 342, 207–220. [Google Scholar] [CrossRef]
- Blevins, D.G.; Lukaszewski, K.M. Boron in Plant Structure and Function. Annual Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 481–500. [Google Scholar] [CrossRef] [PubMed]
- Durbak, A.R.; Phillips, K.A.; Pike, S.; O’Neill, M.A.; Mares, J.; Gallavotti, A.; Malcomber, S.T.; Gassmann, W.; McSteen, P. Transport of Boron by the tassel-less1 Aquaporin Is Critical for Vegetative and Reproductive Development in Maize. Plant Cell 2014, 26, 2978–2995. [Google Scholar] [CrossRef]
- Leonard, A.; Holloway, B.; Guo, M.; Rupe, M.; Yu, G.; Beatty, M.; Zastrow-Hayes, G.; Meeley, R.; Llaca, V.; Butler, K.; et al. tassel-less1 encodes a boron channel protein required for inflorescence development in maize. Plant Cell Physiol. 2014, 55, 1044–1054. [Google Scholar] [CrossRef]
- Bárzana, G.; Aroca, R.; Bienert, G.P.; Chaumont, F.; Ruiz-Lozano, J.M. New insights into the regulation of aquaporins by the arbuscular mycorrhizal symbiosis in maize plants under drought stress and possible Implications for plant performance. Mol. Plant-Microbe Interact. 2014, 27, 349–363. [Google Scholar] [CrossRef]
- Chatterjee, M.; Tabi, Z.; Galli, M.; Malcomber, S.; Buck, A.; Muszynski, M.; Gallavotti, A. The Boron Efflux Transporter ROTTEN EAR Is Required for Maize Inflorescence Development and Fertility. Plant Cell 2014, 26, 2962–2977. [Google Scholar] [CrossRef]
- Kumar, A.; Verma, J.P. Does plant—Microbe interaction confer stress tolerance in plants: A review? Microbiol. Res. 2018, 207, 41–52. [Google Scholar] [CrossRef]
- Augé, R.M. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 2001, 11, 3–42. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Porcel, R.; Bárzana, G.; Azcón, R.; Aroca, R. Contribution of arbuscular mycorrhizal symbiosis to plant drought tolerance: State of the art. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Aroca, R., Ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2012; pp. 335–362. ISBN 978-3-642-32652-3. [Google Scholar]
- Maurel, C.; Plassard, C. Aquaporins: For more than water at the plant – fungus interface? New Phytol. 2011, 190, 815–817. [Google Scholar] [CrossRef] [PubMed]
- Uehlein, N.; Fileschi, K.; Eckert, M.; Bienert, G.P.; Bertl, A.; Kaldenhoff, R. Arbuscular mycorrhizal symbiosis and plant aquaporin expression. Phytochemistry 2007, 68, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Filho, J.A.C.; Sobrinho, R.R.; Pascholati, S.F. Arbuscular mycorrhizal symbiosis and its role in plant nutrition in sustainable agriculture. In Agriculturally Important Microbes for Sustainable Agriculture; Meena, V.S., Ed.; Springer Nature: Singapore, 2017; pp. 129–164. ISBN 9789811053436. [Google Scholar]
- Shorrocks, V.M. The occurrence and correction of boron deficiency. Plant Soil 1997, 193, 121–148. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Aroca, R.; Chaumont, F.; Ruiz-Lozano, J.M. Enhanced drought stress tolerance by the arbuscular mycorrhizal symbiosis in a drought-sensitive maize cultivar Is related to a broader and differential regulation of host plant aquaporins than in a drought-tolerant cultivar. Front. Plant Sci. 2017, 8, 1056. [Google Scholar] [CrossRef] [PubMed]
- Matthes, M.S.; Robil, J.M.; Tran, T.; Kimble, A.; McSteen, P. Increased transpiration is correlated with reduced boron deficiency symptoms in the maize tassel-less1 mutant. Physiol. Plant. 2018, 163, 344–355. [Google Scholar] [CrossRef]
- Wimmer, M.A.; Eichert, T. Review: Mechanisms for boron deficiency-mediated changes in plant water relations. Plant Sci. 2013, 203–204, 25–32. [Google Scholar] [CrossRef]
- Calderan-Rodrigues, M.J.; Fonseca, J.G.; De Moraes, F.E.; Setem, L.V.; Begossi, A.C.; Labate, C.A. Plant cell wall proteomics: A focus on monocot species, Brachypodium distachyon, Saccharum spp. and Oryza sativa. Int. J. Mol. Sci. 2019, 20, 1975. [Google Scholar] [CrossRef]
- Pallotta, M.; Schnurbusch, T.; Hayes, J.; Hay, A.; Baumann, U.; Paull, J.; Langridge, P.; Sutton, T. Molecular basis of adaptation to high soil boron in wheat landraces and elite cultivars. Nature 2014, 514, 88–91. [Google Scholar] [CrossRef]
- Pommerrenig, B.; Junker, A.; Abreu, I.; Bieber, A.; Fuge, J.; Willner, A.; Bienert, M.D.; Altmann, T.; Bienert, G.P. Identification of Rapeseed (Brassica napus) cultivars with a high tolerance to boron-deficient conditions. Front. Plant Sci. 2018, 9, 1142. [Google Scholar] [CrossRef]
- Lewis, D.H. Boron: The essential element for vascular plants that never was. New Phytol. 2019, 221, 1685–1690. [Google Scholar] [CrossRef]
- Dannel, F.; Pfeffer, H.; Römheld, V. Update on boron in higher plants - Uptake, primary translocation and compartmentation. Plant Biol. 2002, 4, 193–204. [Google Scholar] [CrossRef]
- Hrmova, M.; Gilliham, M. Plants fighting back: To transport or not to transport, this is a structural question. Curr. Opin. Plant Biol. 2018, 46, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Ruuhola, T.; Lehto, T. Do ectomycorrhizas affect boron uptake in Betula pendula ? Can. J. For. Res. 2014, 44, 1013–1019. [Google Scholar] [CrossRef]
- Sonmez, O.; Aydemir, S.; Kaya, C. Mitigation effects of mycorrhiza on boron toxicity in wheat (Triticum durum) plants. New Zealand J. Crop Hortic. Sci. 2009, 37, 99–104. [Google Scholar] [CrossRef]
- Ferrol, N.; Tamayo, E.; Vargas, P. The heavy metal paradox in arbuscular mycorrhizas: From mechanisms to biotechnological applications. J. Exp. Bot. 2016, 67, 6253–6565. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhao, H.; Zou, C.; Li, Y.; Chen, Y.; Wang, Z.; Jiang, Y.; Liu, A.; Zhao, P.; Wang, M.; et al. Combined inoculation with multiple arbuscular mycorrhizal fungi improves growth, nutrient uptake and photosynthesis in cucumber seedlings. Front. Microbiol. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Essahibi, A.; Benhiba, L.; Babram, M.A.; Ghoulam, C.; Waddoury, A. Influence of arbuscular mycorrhizal fungi on the functional mechanisms associated with drought tolerance in carob ( Ceratonia siliqua L.). Trees 2017, 32, 87–97. [Google Scholar] [CrossRef]
- Liu, C.; Dai, Z.; Cui, M.; Lu, W.; Sun, H. Arbuscular mycorrhizal fungi alleviate boron toxicity in Puccinellia tenuiflora under the combined stresses of salt and drought. Environ. Pollut. 2018, 240, 557–565. [Google Scholar] [CrossRef]
- Macho-Rivero, M.A.; Herrera-Rodríguez, M.B.; Brejcha, R.; Schäffner, A.R.; Tanaka, N.; Fujiwara, T.; González-Fontes, A.; Camacho-Cristóbal, J.J.; Herrera-Rodrguez, M.B.; Brejcha, R.; et al. Boron Toxicity Reduces Water Transport from Root to Shoot in Arabidopsis Plants. Evidence for a Reduced Transpiration Rate and Expression of Major PIP Aquaporin Genes. Plant Cell Physiol. 2018, 59, 836–844. [Google Scholar] [CrossRef]
- Moshelion, M.; Hachez, C.; Ye, Q.; Cavez, D.; Bajji, M.; Jung, R.; Chaumont, F. Membrane water permeability and aquaporin expression increase during growth of maize suspension cultured cells. Plant Cell Environ. 2009, 32, 1334–1345. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Ding, L.; Chaumont, F.; Aroca, R.; Ruiz-Lozano, J.M. The arbuscular mycorrhizal symbiosis regulates aquaporins activity and improves root cell water permeability in maize plants subjected to water stress. Plant Cell Environ. 2019, 42, 1–17. [Google Scholar] [CrossRef]
- Kaya, C.; Tuna, A.L.; Dikilitas, M.; Ashraf, M.; Koskeroglu, S.; Guneri, M. Supplementary phosphorus can alleviate boron toxicity in tomato. Scientia Hortic. 2009, 121, 284–288. [Google Scholar] [CrossRef]
- Zhen, M.; Cui, M.; Xia, J.; Ma, C.; Liu, C. Effect of nitrogen and phosphorus on alleviation of boron toxicity in Puccinellia tenuiflora under the combined stresses of salt and drought. J. Plant Nutr. 2019, 42, 1594–1604. [Google Scholar] [CrossRef]
- Gunes, A.; Soylemezoglu, G.; Inal, A.; Bagci, E.G.; Coban, S.; Sahin, O. Antioxidant and stomatal responses of grapevine (Vitis vinifera L.) to boron toxicity. Scientia Hortic. 2006, 110, 279–284. [Google Scholar] [CrossRef]
- Gupta, U.; Verma, P.; Solanki, H. Impact of high boron concentration on plants. Biolife 2014, 2, 610–614. [Google Scholar]
- Bansal, A.; Sankararamakrishnan, R. Homology modeling of major intrinsic proteins in rice, maize and Arabidopsis: Comparative analysis of transmembrane helix association and aromatic/arginine selectivity filters. BMC Struct. Biol. 2007, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Wallace, I.S.; Roberts, D.M. Distinct Transport Selectivity of Two Structural Subclasses of the Nodulin-like Intrinsic Protein Family of Plant Aquaglyceroporin Channels. Biochemistry 2005, 44, 16826–16834. [Google Scholar] [CrossRef] [PubMed]
- Chaumont, F.; Tyerman, S.D. Aquaporins: Highly Regulated Channels Controlling Plant Water Relations. Plant Physiol. 2014, 164, 1600–1618. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Aroca, R.; Chaumont, F.; Ruiz-Lozano, J.M. Contribution of the arbuscular mycorrhizal symbiosis to the regulation of radial root water transport in maize plants under water deficit. Environ. Exp. Bot. 2019, 167, 103821. [Google Scholar] [CrossRef]
- Kapilan, R.; Vaziri, M.; Zwiazek, J.J. Regulation of aquaporins in plants under stress. Biol. Res. 2018, 51, 4. [Google Scholar] [CrossRef]
- Chaumont, F.; Moshelion, M.; Daniels, M.J. Regulation of plant aquaporin activity. Biol. Cell 2005, 97, 749–764. [Google Scholar] [CrossRef]
- Luu, D.T.; Maurel, C. Aquaporin Trafficking in Plant Cells: An Emerging Membrane-Protein Model. Traffic 2013, 14, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Santoni, V. Plant Aquaporin Posttranslational Regulation. In Plant Aquaporins, Signaling and Communication in Plants; Chaumont, F., Tyerman, S.D., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 83–105. [Google Scholar]
- Quiroga, G.; Erice, G.; Aroca, R.; Zamarreño, Á.M.; García-Mina, J.M.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal symbiosis and salicylic acid regulate aquaporins and root hydraulic properties in maize plants subjected to drought. Agric. Water Manag. 2018, 202, 271–284. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 1–32. [Google Scholar]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2^-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hachez, C.; Moshelion, M.; Zelazny, E.; Cavez, D.; Chaumont, F. Localization and quantification of plasma membrane aquaporin expression in maize primary root: A clue to understanding their role as cellular plumbers. Plant Mol. Biol. 2006, 62, 305–323. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Molina, S.; Zamarreño, A.M.; García-Mina, J.M.; Aroca, R. The symbiosis with the arbuscular mycorrhizal fungus Rhizophagus irregularis drives root water transport in flooded tomato plants. Plant Cell Physiol. 2014, 55, 1017–1029. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycorrhization (%) | Plant DW (g plant−1) | Gs (mmol H2O m−2s−1) | SPAD | ∆Fv/Fm’ | |||
|---|---|---|---|---|---|---|---|
| WW | B0 | NON-AM | n.d. | 3.98±0.18 AB | 40.3±8.05 C | 10.6±0.66 B | 0.65±0.01A |
| AM | 54.3 ± 4.48 abc | 4.06±0.20 AB | 148.4±33.0 A | 10.5 ±0.54 B | 0.66±0.01 A | ||
| B25 | NON-AM | n.d. | 3.71±0.11 B | 53.8±10.3 C | 10.6 ±0.43 B | 0.63 ±0.01 AB | |
| AM | 47.7±2.60 bc | 4.21±0.13 A | 105.4±16.6 AB | 10.6 ±0.64 B | 0.64 ±0.01 AB | ||
| B100 | NON-AM | n.d. | 3.13±0.16 C | 63.9±6.36 BC | 10.7 ±0.55 B | 0.66 ±0.01 A | |
| AM | 44.3±1.20 c | 4.33±0.18 A | 117.6±23.4 A | 12.6 ±0.65 A | 0.62±0.01 B | ||
| DS | B0 | NON-AM | n.d. | 2.86±0.08 b | 44.3 ±4.01 ab | 5.66 ±0.53 b | 0.52 ±0.04 b |
| AM | 46.3±2.33 bc | 3.49±0.13 a | 16.9±3.42 c | 8.98 ±0.29 a | 0.61 ±0.04 ab | ||
| B25 | NON-AM | n.d. | 2.72±0.09 b | 47.6±10.6 a | 5.38 ±0.41 b | 0.58±0.04 ab | |
| AM | 58.7±6.77 ab | 3.63±0.22 a | 25.5±5.11c | 8.39 ±0.27 a | 0.61±0.03 ab | ||
| B100 | NON-AM | n.d. | 2.72±0.11 b | 30.3±3.96 bc | 4.92 ±0.18 b | 0.62 ±0.04 ab | |
| AM | 64.7±2.90 a | 3.71±0.09 a | 27.2±2.35 c | 8.97 ±1.02 a | 0.63 ±0.02 a | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiroga, G.; Erice, G.; Aroca, R.; Ruiz-Lozano, J.M. Elucidating the Possible Involvement of Maize Aquaporins in the Plant Boron Transport and Homeostasis Mediated by Rhizophagus irregularis under Drought Stress Conditions. Int. J. Mol. Sci. 2020, 21, 1748. https://doi.org/10.3390/ijms21051748
Quiroga G, Erice G, Aroca R, Ruiz-Lozano JM. Elucidating the Possible Involvement of Maize Aquaporins in the Plant Boron Transport and Homeostasis Mediated by Rhizophagus irregularis under Drought Stress Conditions. International Journal of Molecular Sciences. 2020; 21(5):1748. https://doi.org/10.3390/ijms21051748
Chicago/Turabian StyleQuiroga, Gabriela, Gorka Erice, Ricardo Aroca, and Juan Manuel Ruiz-Lozano. 2020. "Elucidating the Possible Involvement of Maize Aquaporins in the Plant Boron Transport and Homeostasis Mediated by Rhizophagus irregularis under Drought Stress Conditions" International Journal of Molecular Sciences 21, no. 5: 1748. https://doi.org/10.3390/ijms21051748
APA StyleQuiroga, G., Erice, G., Aroca, R., & Ruiz-Lozano, J. M. (2020). Elucidating the Possible Involvement of Maize Aquaporins in the Plant Boron Transport and Homeostasis Mediated by Rhizophagus irregularis under Drought Stress Conditions. International Journal of Molecular Sciences, 21(5), 1748. https://doi.org/10.3390/ijms21051748