The Binding of Aβ42 Peptide Monomers to Sphingomyelin/Cholesterol/Ganglioside Bilayers Assayed by Density Gradient Ultracentrifugation

 ,
,
Abstract
1. Introduction
2. Results
2.1. Binding Assessment through Flotation Assays
2.2. Thioflavin T Assays
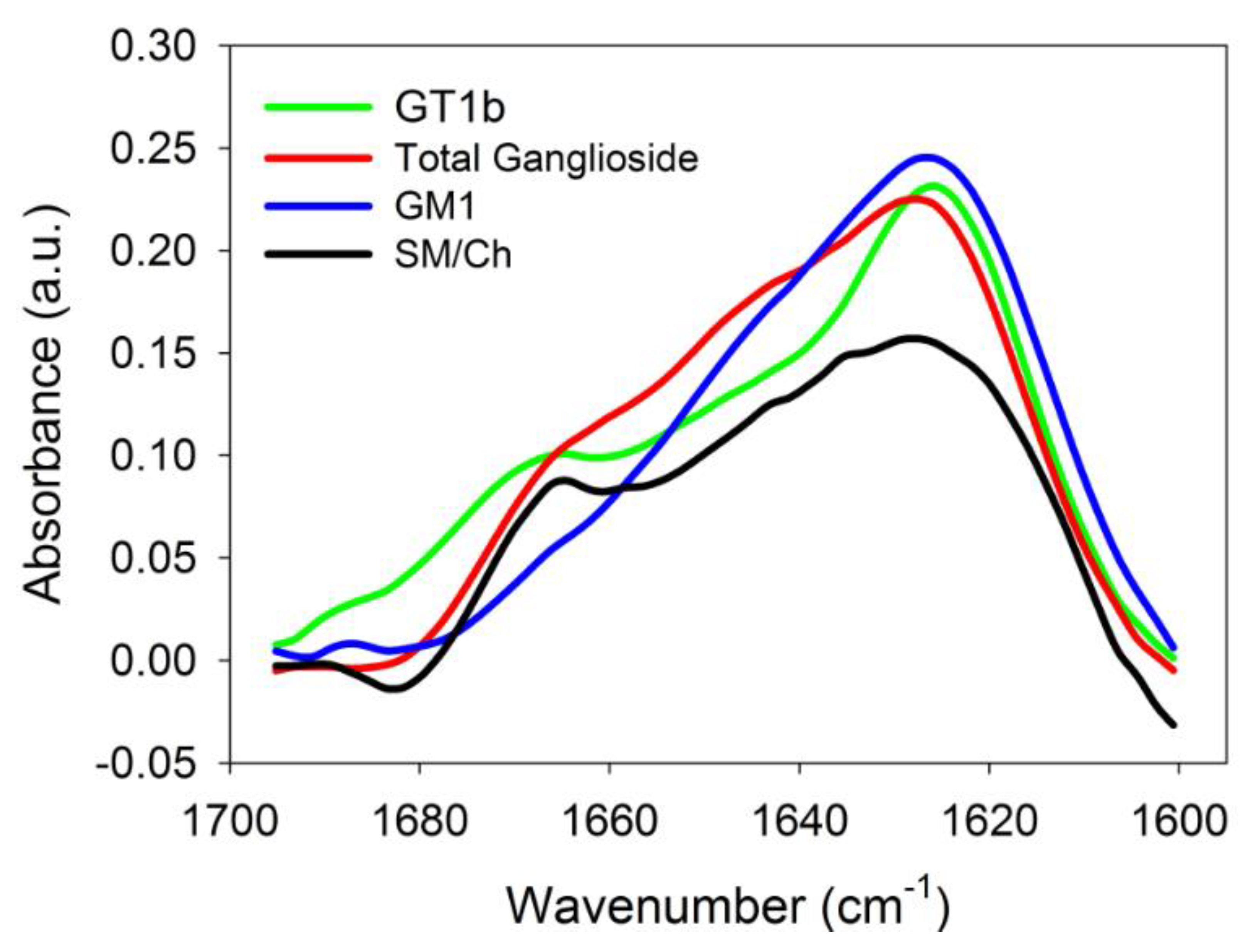
2.3. Infrared Spectroscopic Studies
2.4. Langmuir Balance Measurements
3. Materials and Methods
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Glenner, G.G.; Wong, C.W. Alzheimer’s disease: Initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem. Biophys. Res. Commun. 1984, 120, 885–890. [Google Scholar] [CrossRef]
- Huynh, T.V.; Holtzman, D.M. In Search of an Identity for Amyloid Plaques. Trends Neurosci. 2018, 41, 483–486. [Google Scholar] [CrossRef]
- Cline, E.N.; Bicca, M.A.; Viola, K.L.; Klein, W.L. The Amyloid-β Oligomer Hypothesis: Beginning of the Third Decade. J. Alzheimer’s Dis. 2018, 64, S567–S610. [Google Scholar] [CrossRef]
- Morgado, I.; Garvey, M. Lipids in Amyloid-β Processing, Aggregation and Toxicity. Adv. Exp. Med. Biol. 2015, 855, 67–94. [Google Scholar]
- Matsuzaki, K. How do membranes initiate Alzheimer’s Disease? Formation of toxic amyloid fibrils by the amyloid β-protein on ganglioside clusters. Acc. Chem Res. 2014, 47, 2397–2404. [Google Scholar] [CrossRef] [PubMed]
- Penke, B.; Paragi, G.; Gera, J.; Berkecz, R.; Kovács, Z.; Crul, T.; VÍgh, L. The Role of Lipids and Membranes in the Pathogenesis of Alzheimer’s Disease: A Comprehensive View. Curr. Alzheimer Res. 2018, 15, 1191–1212. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Zhang, Z.; Zhao, W.; Yang, J. Interactions between amyloid β peptide and lipid membranes. Biochim. Biophys. Acta Biomembr. 2018, 2736, 30114–30117. [Google Scholar] [CrossRef] [PubMed]
- Riddell, D.R.; Christie, G.; Hussain, I.; Dingwall, C. Compartmentalization of beta-secretase (Asp2) into low-buoyant density, noncaveolar lipid rafts. Curr. Biol. 2001, 11, 1288–1293. [Google Scholar] [CrossRef]
- Kakio, A.; Nishimoto, S.; Yanagisawa, K.; Kozutsumi, Y.; Matsuzaki, K. Interactions of amyloid beta-protein with various gangliosides in raft-like membranes: Importance of GM1 ganglioside-bound form as an endogenous seed for Alzheimer amyloid. Biochemistry 2002, 41, 7385–7390. [Google Scholar] [CrossRef]
- Goñi, F.M.; Alonso, A.; Bagatolli, L.A.; Brown, R.E.; Marsh, D.; Prieto, M.; Thewalt, J.L. Phase diagrams of lipid mixtures relevant to the study of membrane rafts. Biochim. Biophys. Acta 2008, 1781, 665–684. [Google Scholar] [CrossRef]
- Sani, M.A.; Gehman, J.D.; Separovic, F. Lipid matrix plays a role in Aβ fibril kinetics and morphology. FEBS Lett. 2011, 585, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Ray, I.; Chauhan, V.P.S. Interaction of amyloid β-protein with anionic phospholipids: Possible involvement of Lys-28 and C-terminus aliphatic amino acids. Neurochem. Res. 2000, 25, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Ahyayauch, H.; Raab, M.; Busto, J.V.; Andraka, N.; Arrondo, J.L.; Masserini, M.; Tvaroska, I.; Goñi, F.M. Binding of β-amyloid (1-42) peptide to negatively charged phospholipid membranes in the liquid-ordered state: Modeling and experimental studies. Biophys. J. 2012, 103, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, K.; Odaka, A.; Suzuki, N.; Ihara, Y. GM1 ganglioside-bound amyloid beta-protein (A beta): A possible form of preamyloid in Alzheimer’s disease. Nat. Med. 1995, 1, 1062–1066. [Google Scholar] [CrossRef]
- Goñi, F.M. “Rafts”: A nickname for putative transient nanodomains. Chem. Phys. Lipids 2019, 218, 34–39. [Google Scholar] [CrossRef]
- Vahed, M.; Neya, S.; Matsuzaki, K.; Hoshino, T. Analysis of Physicochemical Interaction of Aβ40 with a GM1 Ganglioside-Containing Lipid Membrane. J. Phys. Chem B. 2018, 22, 3771–3781. [Google Scholar] [CrossRef]
- Amaro, M.; Šachl, R.; Aydogan, G.; Mikhalyov, I.I.; Vácha, R.; Hof, M. GM1 Ganglioside Inhibits β-Amyloid Oligomerization Induced by Sphingomyelin. Angew. Chem. Int. Ed. Engl. 2016, 55, 9411–9415. [Google Scholar] [CrossRef]
- Manna, M.; Mukhopadhyay, C. Binding, conformational transition and dimerization of amyloid-β peptide on GM1-containing ternary membrane: Insights from molecular dynamics simulation. PLoS ONE 2013, 8, e71308. [Google Scholar] [CrossRef]
- Ewald, M.; Henry, S.; Lambert, E.; Feuillie, C.; Bobo, C.; Cullin, C.; Lecomte, S.; Molinari, M. High speed atomic force microscopy to investigate the interactions between toxic Aβ1-42 peptides and model membranes in real time: Impact of the membrane composition. Nanoscale 2019, 11, 7229–7238. [Google Scholar] [CrossRef]
- Okada, Y.; Okubo, K.; Ikeda, K.; Yano, Y.; Hoshino, M.; Hayashi, Y.; Kiso, Y.; Itoh-Watanabe, H.; Naito, A.; Matsuzaki, K. Toxic Amyloid Tape: A Novel Mixed Antiparallel/Parallel β-Sheet Structure Formed by Amyloid β-Protein on GM1 Clusters. ACS Chem. Neurosci. 2019, 10, 563–572. [Google Scholar] [CrossRef]
- Dai, Y.; Zhang, M.; Shi, X.; Wang, K.; Gao, G.; Shen, L.; Sun, T. Kinetic study of Aβ(1-42) amyloidosis in the presence of ganglioside-containing vesicles. Colloids Surf. B Biointerfaces 2020, 185, 110615. [Google Scholar] [CrossRef] [PubMed]
- Staneva, G.; Puff, N.; Stanimirov, S.; Tochev, T.; Angelova, M.I.; Seigneuret, M. The Alzheimer’s disease amyloid-β peptide affects the size-dynamics of raft-mimicking Lo domains in GM1-containing lipid bilayers. Soft Matter 2018, 14, 9609–9618. [Google Scholar] [CrossRef] [PubMed]
- Hirai, M.; Ajito, S.; Sato, S.; Ohta, N.; Igarashi, N.; Shimizu, N. Preferential Intercalation of Human Amyloid-β Peptide into Interbilayer Region of Lipid-Raft Membrane in Macromolecular Crowding Environment. J. Phys. Chem B. 2018, 122, 9482–9489. [Google Scholar] [CrossRef] [PubMed]
- Antón, Z.; Landajuela, A.; Hervás, J.H.; Montes, L.R.; Hernández-Tiedra, S.; Velasco, G.; Goñi, F.M.; Alonso, A. Human Atg8-cardiolipin interactions in mitophagy: Specific properties of LC3B, GABARAPL2 and GABARAP. Autophagy 2016, 12, 2386–2403. [Google Scholar] [CrossRef]
- Rujas, E.; Caaveiro, J.M.; Insausti, S.; García-Porras, M.; Tsumoto, K.; Nieva, J.L. Peripheral Membrane Interactions Boost the Engagement by an Anti-HIV-1 Broadly Neutralizing Antibody. J. Biol Chem. 2017, 292, 5571–5583. [Google Scholar] [CrossRef]
- Gobbi, M.; Re, F.; Canovi, M.; Beeg, M.; Gregori, M.; Sesana, S.; Sonnino, S.; Brogioli, D.; Musicanti, C.; Gasco, P.; et al. Lipid-based nanoparticles with high binding affinity for amyloid-beta1-42 peptide. Biomaterials 2010, 31, 6519–6529. [Google Scholar] [CrossRef]
- Nicastro, M.C.; Spigolon, D.; Librizzi, F.; Moran, O.; Ortore, M.G.; Bulone, D.; Biagio, P.L.; Carrotta, R. Amyloid β-peptide insertion in liposomes containing GM1-cholesterol domains. Biophys. Chem. 2016, 208, 9–16. [Google Scholar] [CrossRef]
- Arrondo, J.L.; Muga, A.; Castresana, J.; Goñi, F.M. Quantitative studies of the structure of proteins in solution by Fourier-transform infrared spectroscopy. Prog. Biophys. Mol. Biol. 1993, 59, 23–56. [Google Scholar] [CrossRef]
- Sánchez-Magraner, L.; Cortajarena, A.L.; Ostolaza, H. Membrane insertion of Escherichia coli alpha-hemolysin is independent from membrane lysis. J. Biol. Chem. 2006, 281, 5461–5467. [Google Scholar] [CrossRef]
- Nilsson, M.R. Techniques to study amyloid fibril formation in vitro. Methods 2004, 34, 151–160. [Google Scholar] [CrossRef]
- Mayer, L.D.; Hope, M.J.; Cullis, P.R. Vesicles of variable sizes produced by a rapid extrusion procedure. Biochim. Biophys. Acta 1986, 858, 161–168. [Google Scholar] [CrossRef]
- Andraka, N.; Sánchez-Magraner, L.; García-Pacios, M.; Goñi, F.M.; Arrondo, J.L. The conformation of human phospholipid scramblase 1, as studied by infrared spectroscopy. Effects of calcium and detergent. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Vaz, W.L. Model systems, lipid rafts, and cell membranes. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 269–295. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, L.; Simons, K. Lipid rafts and membrane dynamics. J. Cell Sci. 2005, 118, 1099–1102. [Google Scholar] [CrossRef] [PubMed]
- Goni, F.M. Membrane Nanodomains; eLS: Washington, DC, USA, 2020; in press. [Google Scholar]
- Christensen, M.; Schiøtt, B. Revealing a Dual Role of Ganglioside Lipids in the Aggregation of Membrane-Associated Islet Amyloid Polypeptide. J. Membr. Biol. 2019, 252, 343–356. [Google Scholar] [CrossRef]
- Cebecauer, M.; Hof, M.; Amaro, M. Impact of GM1 on Membrane-Mediated Aggregation/Oligomerization of β-Amyloid: A Unifying View. Biophys J. 2017, 113, 1194–1199. [Google Scholar] [CrossRef]
- Mori, K.; Mahmood, M.I.; Neya, S.; Matsuzaki, K.; Hoshino, T. Formation of GM1 ganglioside clusters on the lipid membrane containing sphingomyeline and cholesterol. J. Phys. Chem B. 2012, 116, 5111–5121. [Google Scholar] [CrossRef]
- Brown, A.M.; Bevan, D.R. Influence of sequence and lipid type on membrane perturbation by human and rat amyloid β-peptide (1-42). Arch. Biochem. Biophys. 2017, 614, 1–13. [Google Scholar] [CrossRef]
- Seghezza, S.; Diaspro, A.; Canale, C.; Dante, S. Cholesterol drives aβ(1-42) interaction with lipid rafts in model membranes. Langmuir 2014, 30, 13934–13941. [Google Scholar] [CrossRef]
- Lockhart, C.; Klimov, D.K. Cholesterol Changes the Mechanisms of Aβ Peptide Binding to the DMPC Bilayer. J. Chem. Inf. Model 2017, 57, 2554–2565. [Google Scholar] [CrossRef] [PubMed]
- Xiang, N.; Lyu, Y.; Zhu, X.; Narsimhan, G. Investigation of the interaction of amyloid β peptide (11-42) oligomers with a 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) membrane using molecular dynamics simulation. Phys. Chem. Phys. 2018, 20, 6817–6829. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Fractions | |||||
|---|---|---|---|---|---|
| F1 | F2 | F3 | F4 | L | |
| Bilayer composition | |||||
| SM/Chol (1:1 mol ratio) | n.d. | n.d. | 28 ± 2.7 | 41 ± 3.3 | 30 ± 2.2 |
| SM/Chol + 5 mol% total gangliosides | n.d. | n.d. | 4.9 ± 0.1 | 84 ± 3.9 | 11 ± 1.4 |
| SM/Chol + 5 mol% GM1 | n.d. | n.d. | 8.2 ± 0.3 | 91 ± 1.7 | n.d. |
| SM/Chol + 5 mol% GT1b | n.d. | n.d. | 10 ± 1.1 | 89 ± 1.3 | n.d. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahyayauch, H.; de la Arada, I.; Masserini, M.E.; Arrondo, J.L.R.; Goñi, F.M.; Alonso, A. The Binding of Aβ42 Peptide Monomers to Sphingomyelin/Cholesterol/Ganglioside Bilayers Assayed by Density Gradient Ultracentrifugation. Int. J. Mol. Sci. 2020, 21, 1674. https://doi.org/10.3390/ijms21051674
Ahyayauch H, de la Arada I, Masserini ME, Arrondo JLR, Goñi FM, Alonso A. The Binding of Aβ42 Peptide Monomers to Sphingomyelin/Cholesterol/Ganglioside Bilayers Assayed by Density Gradient Ultracentrifugation. International Journal of Molecular Sciences. 2020; 21(5):1674. https://doi.org/10.3390/ijms21051674
Chicago/Turabian StyleAhyayauch, Hasna, Igor de la Arada, Massimo E. Masserini, José L. R. Arrondo, Félix M. Goñi, and Alicia Alonso. 2020. "The Binding of Aβ42 Peptide Monomers to Sphingomyelin/Cholesterol/Ganglioside Bilayers Assayed by Density Gradient Ultracentrifugation" International Journal of Molecular Sciences 21, no. 5: 1674. https://doi.org/10.3390/ijms21051674
APA StyleAhyayauch, H., de la Arada, I., Masserini, M. E., Arrondo, J. L. R., Goñi, F. M., & Alonso, A. (2020). The Binding of Aβ42 Peptide Monomers to Sphingomyelin/Cholesterol/Ganglioside Bilayers Assayed by Density Gradient Ultracentrifugation. International Journal of Molecular Sciences, 21(5), 1674. https://doi.org/10.3390/ijms21051674