Retrotransposons Manipulating Mammalian Skeletal Development in Chondrocytes



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
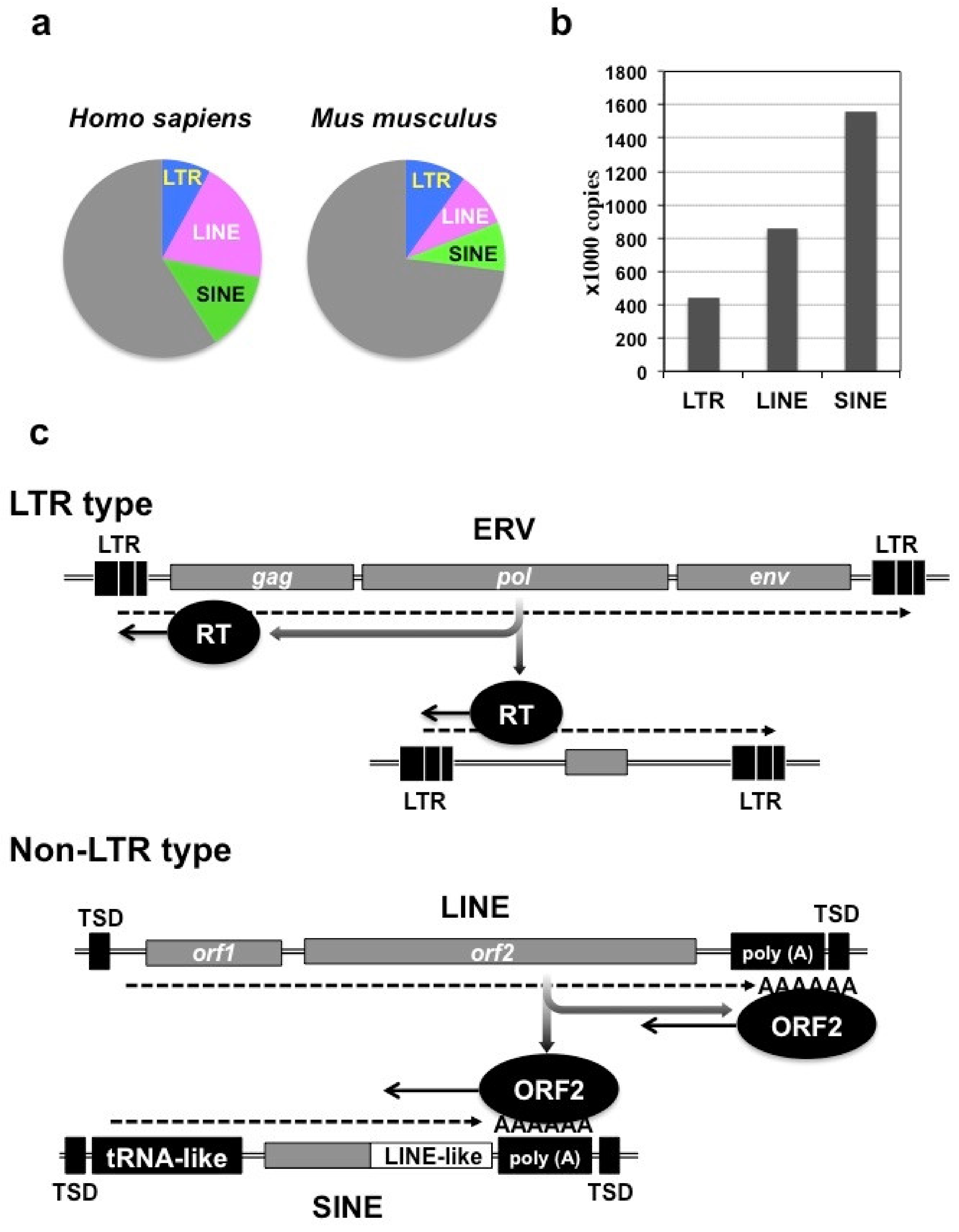
2. Retrotransposons in the Mammalian Genome
3. Creation and Modification of Genes by Retrotransposons
3.1. Modification of Pre-existing Host Genes by Retrotransposons
3.2. Creation of Novel Genes by Retrotransposons
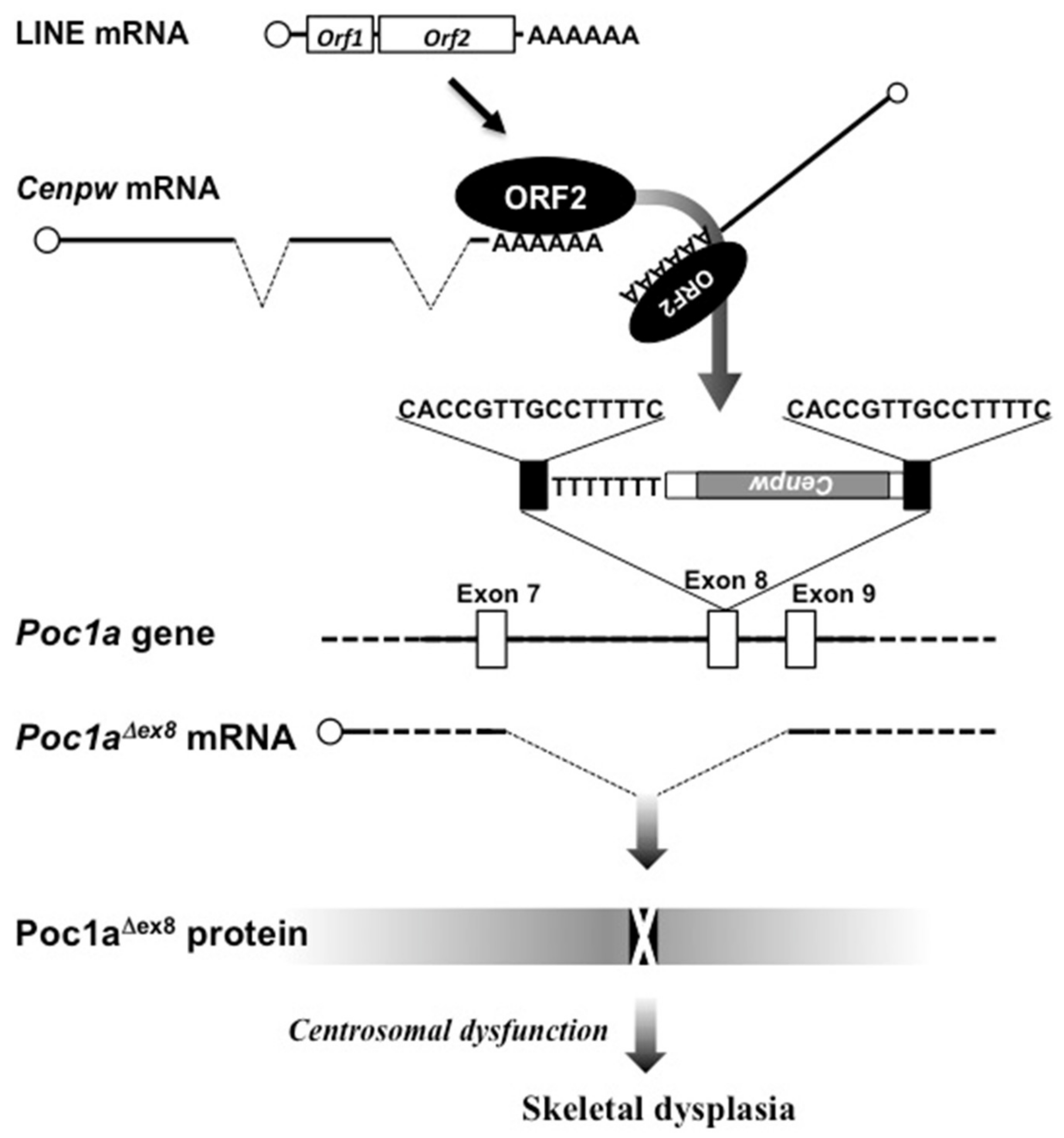
4. Gene Modification by a LINE Leading to Chondrodysplasia in Mice
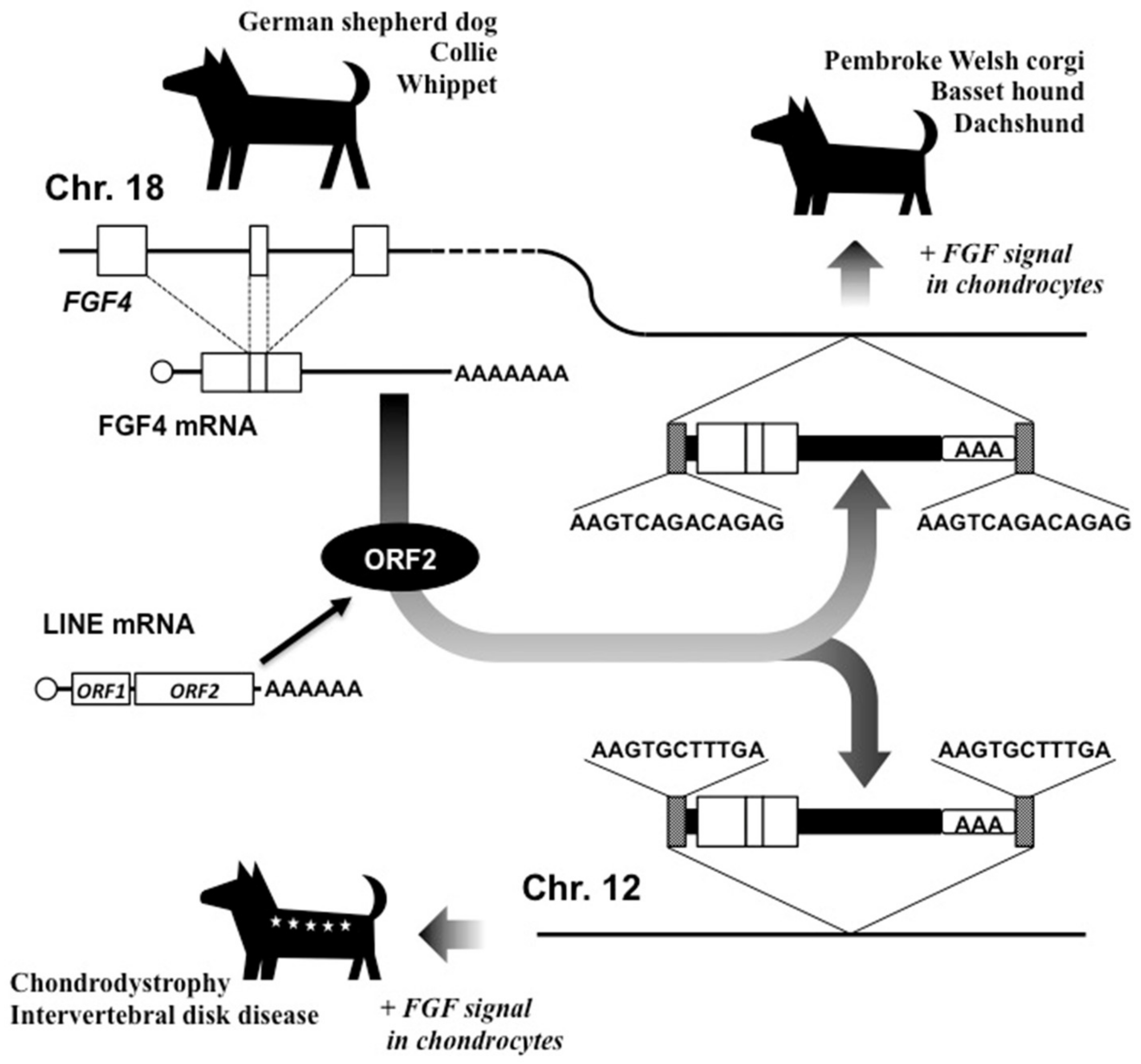
5. Creation of Short-legged Dogs: New FGF4 Gene Formation by LINE-1
6. Hallmarks of Human Genome Regulating Chondrogenesis and Skeletal Development
6.1. Height-related Gene Evolved along with Alu, a SINE Highly Specific to Primates
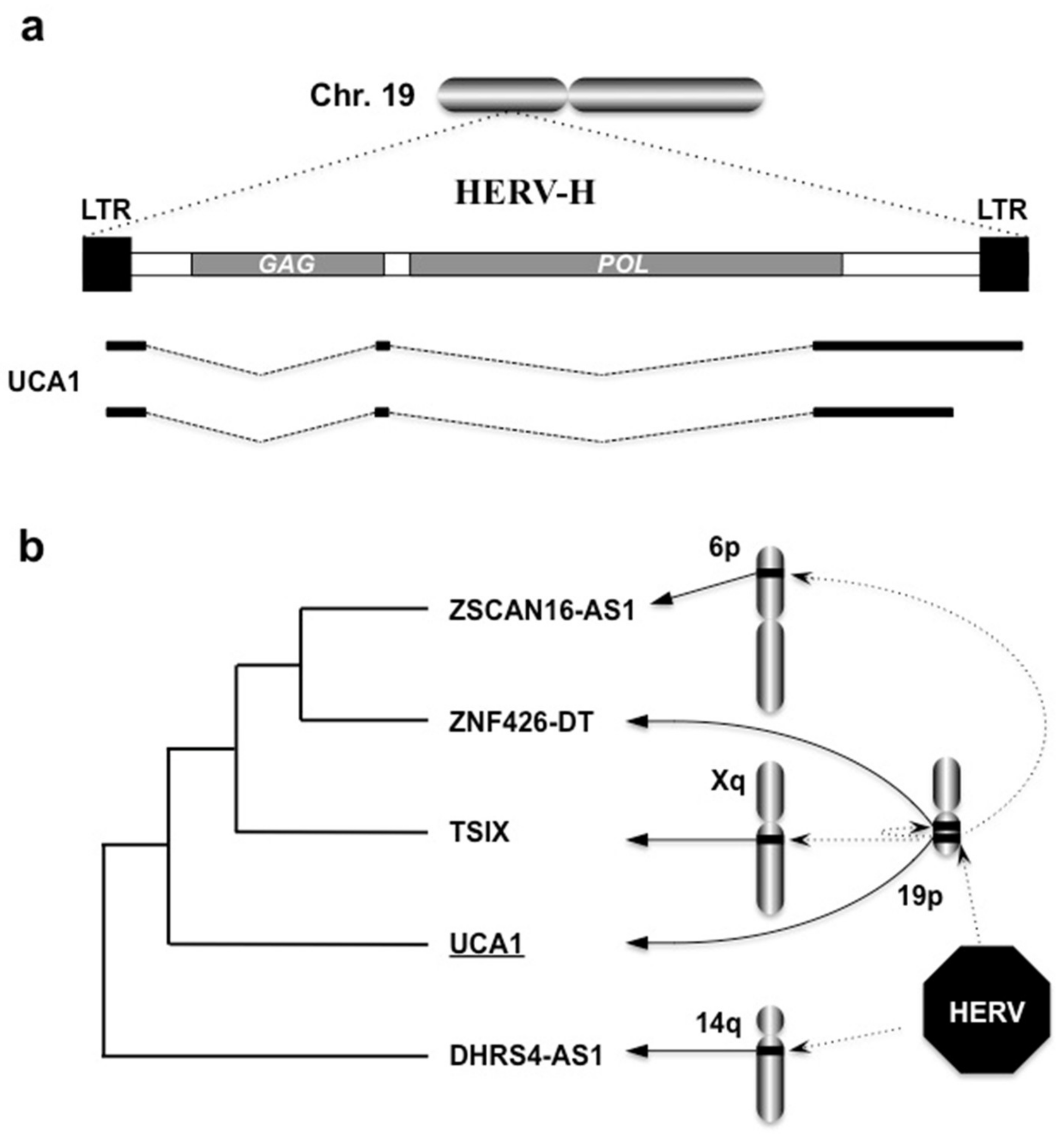
6.2. Human Endogenous Retroviruses
7. Retrotransposons in Human Joint Diseases
7.1. Osteoarthritis
7.2. Rheumatoid Arthritis
8. Conclusive Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ERV | Endogenous retrovirus |
| LINE | Long interspersed nuclear element |
| SINE | Short interspersed nuclear element |
References
- Moraes, F.; Góes, A. A decade of human genome project conclusion: Scientific diffusion about our genome knowledge. Biochem. Mol. Bio.l Educ. 2016, 44, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, S.; Svoboda, P. Retrotransposon-associated long non-coding RNAs in mice and men. Pflug. Arch. 2016, 468, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Powner, M.W.; Gerland, B.; Sutherland, J.D. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature 2009, 459, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Wochner, A.; Attwater, J.; Coulson, A.; Holliger, P. Ribozyme-catalyzed transcription of an active ribozyme. Science 2011, 332, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Kubiak, M.R.; Makałowska, I. Protein-coding genes’ retrocopies and their functions. Viruses 2017, 9, 80. [Google Scholar] [CrossRef]
- Temin, H.; Mizutani, S. RNA-dependent DNA Polymerase in Virions of Rous sarcoma virus. Nature 1972, 226, 1211–1213. [Google Scholar] [CrossRef]
- Deininger, P. Alu elements: Know the SINEs. Genome Biol. 2011, 12, 236. [Google Scholar] [CrossRef]
- Hughes, S.H. Reverse transcription of retroviruses and LTR retrotransposons. Microbiol. Spectr. 2015, 3, MDNA3-0027-2014. [Google Scholar]
- Ohnuki, M.; Tanabe, K.; Sutou, K.; Teramoto, I.; Sawamura, Y.; Narita, M.; Nakamura, M.; Tokunaga, Y.; Nakamura, M.; Watanabe, A.; et al. Dynamic regulation of human endogenous retroviruses mediates factor-induced reprogramming and differentiation potential. Proc. Natl. Acad. Sci. USA 2014, 111, 12426–12431. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Preissl, S.; Amaral, M.L.; Grinstein, J.D.; Farah, E.N.; Destici, E.; Qiu, Y.; Hu, R.; Lee, A.Y.; et al. Transcriptionally active HERV-H retrotransposons demarcate topologically associating domains in human pluripotent stem cells. Nat. Genet. 2019, 51, 1380–1388. [Google Scholar] [CrossRef]
- Svoboda, P.; Stein, P.; Anger, M.; Bernstein, E.; Hannon, G.J.; Schultz, R.M. RNAi and expression of retrotransposons MuERV-L and IAP in preimplantation mouse embryos. Dev. Biol. 2004, 269, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Bodak, M.; Yu, J.; Ciaudo, C. Regulation of LINE-1 in mammals. Biomol. Concepts 2014, 5, 409–428. [Google Scholar] [CrossRef] [PubMed]
- Dewannieux, M.; Esnault, C.; Heidmann, T. LINE-mediated retrotransposition of marked Alu sequences. Nat. Genet. 2003, 35, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Teugels, E.; De Brakeleer, S.; Goelen, G.; Lissens, W.; Sermijn, E.; De Grève, J. De novo Alu element insertions targeted to a sequence common to the BRCA1 and BRCA2 genes. Hum. Mutat. 2005, 26, 284. [Google Scholar] [CrossRef] [PubMed]
- Cordaux, R.; Batzer, M.A. The impact of retrotransposons on human genome evolution. Nat. Rev. Genet. 2009, 10, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.B.; Fang, X.; Fushan, A.A.; Huang, Z.; Lobanov, A.V.; Han, L.; Marino, S.M.; Sun, X.; Turanov, A.A.; Yang, P.; et al. Genome sequencing reveals insights into physiology and longevity of the naked mole rat. Nature 2011, 479, 223–227. [Google Scholar] [CrossRef]
- Hon, C.C.; Ramilowski, J.A.; Harshbarger, J.; Bertin, N.; Rackham, O.J.; Gough, J.; Denisenko, E.; Schmeier, S.; Poulsen, T.M.; Severin, J.; et al. An atlas of human long non-coding RNAs with accurate 5’ ends. Nature 2017, 543, 199–204. [Google Scholar] [CrossRef]
- Vogt, P.K. Retroviral oncogenes: A historical primer. Nat. Rev. Cancer 2012, 12, 639–648. [Google Scholar] [CrossRef]
- Cha, K.B.; Karolyi, I.J.; Hunt, A.; Wenglikowski, A.M.; Wilkinson, E.; Dolan, D.F.; Dootz, G.; Finnegan, A.A.; Seasholtz, A.F.; Hankenson, K.D.; et al. Skeletal dysplasia and male infertility locus on mouse chromosome 9. Genomics 2004, 83, 951–960. [Google Scholar] [CrossRef]
- Geister, K.A.; Brinkmeier, M.L.; Cheung, L.Y.; Wendt, J.; Oatley, M.J.; Burgess, D.L.; Kozloff, K.M.; Cavalcoli, J.D.; Oatley, J.M.; Camper, S.A. LINE-1 mediated insertion into Poc1a (protein of centriole 1 a) causes growth insufficiency and male infertility in mice. Plos Genet. 2015, 11, e1005569. [Google Scholar] [CrossRef]
- Sarig, O.; Nahum, S.; Rapaport, D.; Ishida-Yamamoto, A.; Fuchs-Telem, D.; Qiaoli, L.; Cohen-Katsenelson, K.; Spiegel, R.; Nousbeck, J.; Israeli, S.; et al. Short stature, onychodysplasia, facial dysmorphism, and hypotrichosis syndrome is caused by a POC1A mutation. Am. J. Hum. Genet. 2012, 91, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Parker, H.G.; VonHoldt, B.M.; Quignon, P.; Margulies, E.H.; Shao, S.; Mosher, D.S.; Spady, T.C.; Elkahloun, A.; Cargill, M.; Jones, P.G.; et al. An expressed fgf4 retrogene is associated with breed-defining chondrodysplasia in domestic dogs. Science 2009, 325, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Ornitz, D.M.; Legeai-Mallet, L. Achondroplasia: Development, pathogenesis, and therapy. Dev. Dyn. 2017, 246, 291–309. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.A.; Dickinson, P.J.; Mansour, T.; Sturges, B.K.; Aguilar, M.; Young, A.E.; Korff, C.; Lind, J.; Ettinger, C.L.; Varon, S.; et al. FGF4 retrogene on CFA12 is responsible for chondrodystrophy and intervertebral disc disease in dogs. Proc. Natl. Acad. Sci. USA 2017, 114, 11476–11481. [Google Scholar] [CrossRef]
- Weedon, M.N.; Lango, H.; Lindgren, C.M.; Wallace, C.; Evans, D.M.; Mangino, M.; Freathy, R.M.; Perry, J.R.; Stevens, S.; Hall, A.S.; et al. Genome-wide association analysis identifies 20 loci that influence adult height. Nat. Genet. 2008, 40, 575–583. [Google Scholar] [CrossRef]
- Cho, Y.S.; Go, M.J.; Kim, Y.J.; Heo, J.Y.; Oh, J.H.; Ban, H.J.; Yoon, D.; Lee, M.H.; Kim, D.J.; Park, M.; et al. A large-scale genome-wide association study of Asian populations uncovers genetic factors influencing eight quantitative traits. Nat. Genet. 2009, 41, 527–534. [Google Scholar] [CrossRef]
- Hong, K.W.; Shin, Y.B.; Jin, H.S.; Lim, J.E.; Choi, J.Y.; Chang, K.T.; Kim, H.S.; Oh, B. Alternative splicing of human height-related zinc finger and BTB domain-containing 38 gene through Alu exonization. Biochem. Genet. 2011, 49, 283–291. [Google Scholar] [CrossRef]
- Wang, F.; Li, X.; Xie, X.; Zhao, L.; Chen, W. UCA1, a non-protein-coding RNA up-regulated in bladder carcinoma and embryo, influencing cell growth and promoting invasion. FEBS Lett. 2008, 582, 1919–1927. [Google Scholar] [CrossRef]
- Xue, M.; Li, X.; Wu, W.; Zhang, S.; Wu, S.; Li, Z.; Chen, W. Upregulation of long non-coding RNA urothelial carcinoma associated 1 by CCAAT/enhancer binding protein α contributes to bladder cancer cell growth and reduced apoptosis. Oncol. Rep. 2014, 31, 1993–2000. [Google Scholar] [CrossRef]
- Eissa, S.; Matboli, M.; Essawy, N.O.; Shehta, M.; Kotb, Y.M. Rapid detection of urinary long non-coding RNA urothelial carcinoma associated one using a PCR-free nanoparticle-based assay. Biomarkers 2015, 20, 212–217. [Google Scholar] [CrossRef]
- Li, Z.; Yu, D.; Li, H.; Lv, Y.; Li, S. Long non-coding RNA UCA1 confers tamoxifen resistance in breast cancer endocrinotherapy through regulation of the EZH2/p21 axis and the PI3K/AKT signaling pathway. Int. J. Oncol. 2019, 54, 1033–1042. [Google Scholar] [PubMed]
- Liu, C.; Jin, J.; Shi, J.; Wang, L.; Gao, Z.; Guo, T.; He, Y. Long noncoding RNA UCA1 as a novel biomarker of lymph node metastasis and prognosis in human cancer: A meta-analysis. Biosci. Rep. 2019, 39, BSR20180995. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Taheri, M. UCA1 long non-coding RNA: An update on its roles in malignant behavior of cancers. Biomed. Pharm. 2019, 120, 109459. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.S.; Zhang, Z.; Wang, H.C.; Cai, J.L.; Xu, Q.W.; Li, M.Q.; Chen, Y.C.; Qian, X.P.; Lu, T.J.; Yu, L.Z.; et al. Rapid identification of UCA1 as a very sensitive and specific unique marker for human bladder carcinoma. Clin. Cancer Res. 2006, 12, 4851–4858. [Google Scholar] [CrossRef] [PubMed]
- Mammalian Gene Collection (MGC) Program Team. Generation and initial analysis of more than 15,000 full-length human and mouse cDNA sequences. Proc. Natl. Acad. Sci. USA 2002, 99, 16899–16903. [Google Scholar] [CrossRef] [PubMed]
- Mlynarczyk, S.K.; Panning, B. X inactivation: Tsix and Xist as yin and yang. Curr Biol 2000, 10, R899–R903. [Google Scholar] [CrossRef]
- Wang, C.; Wang, G.; Zhang, Z.; Wang, Z.; Ren, M.; Wang, X.; Li, H.; Yu, Y.; Liu, J.; Cai, L.; et al. The downregulated long noncoding RNA DHRS4-AS1 is protumoral and associated with the prognosis of clear cell renal cell carcinoma. Onco Targets 2018, 11, 5631–5646. [Google Scholar] [CrossRef]
- Yang, Y.; Su, Z.; Song, X.; Liang, B.; Zeng, F.; Chang, X.; Huang, D. Enhancer RNA-driven looping enhances the transcription of the long noncoding RNA DHRS4-AS1, a controller of the DHRS4 gene cluster. Sci. Rep. 2016, 6, 20961. [Google Scholar] [CrossRef]
- Ishikawa, T.; Nishida, T.; Ono, M.; Takarada, T.; Nguyen, H.T.; Kurihara, S.; Furumatsu, T.; Murase, Y.; Takigawa, M.; Oohashi, T.; et al. Physiological role of urothelial cancer-associated one long noncoding RNA in human skeletogenic cell differentiation. J. Cell Physiol. 2018, 233, 4825–4840. [Google Scholar] [CrossRef]
- Shu, T.; He, L.; Wang, X.; Pang, M.; Yang, B.; Feng, F.; Wu, Z.; Liu, C.; Zhang, S.; Liu, B.; et al. Long noncoding RNA UCA1 promotes chondrogenic differentiation of human bone marrow mesenchymal stem cells via miRNA-145-5p/SMAD5 and miRNA-124-3p/SMAD4 axis. Biochem. Biophys. Res. Commun. 2019, 514, 316–322. [Google Scholar] [CrossRef]
- Kumar, P.P.; Emechebe, U.; Smith, R.; Franklin, S.; Moore, B.; Yandell, M.; Lessnick, S.L.; Moon, A.M. Coordinated control of senescence by lncRNA and a novel T-box3 co-repressor complex. eLife 2014, 3, e02805. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Feng, W.; Gu, S.; Wang, H.; Zhang, Y.; Chen, W.; Xu, W.; Lin, C.; Gong, A.; Xu, M. The UCA1/KRAS axis promotes human pancreatic ductal adenocarcinoma stem cell properties and tumor growth. Am. J. Cancer Res. 2019, 9, 496–510. [Google Scholar] [PubMed]
- Wang, Z.Q.; He, C.Y.; Hu, L.; Shi, H.P.; Li, J.F.; Gu, Q.L.; Su, L.P.; Liu, B.Y.; Li, C.; Zhu, Z. Long noncoding RNA UCA1 promotes tumour metastasis by inducing GRK2 degradation in gastric cancer. Cancer Lett. 2017, 408, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Jin, L.; Wang, S.; Zhou, D.; Wang, J.; Tang, Z.; Quan, Z. Long non-coding RNA UCA1 promotes gallbladder cancer progression by epigenetically repressing p21 and E-cadherin expression. Oncotarget 2017, 8, 47957–47968. [Google Scholar] [CrossRef]
- Wang, X.; Gong, Y.; Jin, B.; Wu, C.; Yang, J.; Wang, L.; Zhang, Z.; Mao, Z. Long non-coding RNA urothelial carcinoma associated 1 induces cell replication by inhibiting BRG1 in 5637 cells. Oncol. Rep. 2014, 32, 1281–1290. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, N.; Watabe, K.; Lu, Z.; Wu, F.; Xu, M.; XMo, Y.-Y. Long non-coding RNA UCA1 promotes breast tumor growth by suppression of p27 (Kip1). Cell Death Dis. 2014, 5, e1008. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Cai, Q.; Hu, L.; He, C.Y.; Li, J.F.; Quan, Z.W.; Liu, B.Y.; Li, C.; Zhu, Z.G. Long noncoding RNA UCA1 induced by SP1 promotes cell proliferation via recruiting EZH2 and activating AKT pathway in gastric cancer. Cell Death Dis. 2017, 8, e2839. [Google Scholar] [CrossRef]
- Lin, X.; Spindler, T.J.; de Souza Fonseca, M.A.; Corona, R.I.; Seo, J.H.; Dezem, F.S.; Li, L.; Lee, J.M.; Long, H.W.; Sellers, T.A.; et al. Super-enhancer-associated lncRNA UCA1 interacts directly with AMOT to activate YAP target genes in epithelial ovarian cancer. iScience 2019, 17, 242–255. [Google Scholar] [CrossRef]
- Yang, Z.; Shi, X.; Li, C.; Wang, X.; Hou, K.; Li, Z.; Zhang, X.; Fan, Y.; Qu, X.; Che, X.; et al. Long non-coding RNA UCA1 upregulation promotes the migration of hypoxia-resistant gastric cancer cells through the miR-7-5p/EGFR axis. Exp. Cell Res. 2018, 368, 194–201. [Google Scholar] [CrossRef]
- Li, D.; Hao, S.; Zhang, J. Long non-coding RNA UCA1 exerts growth modulation by miR-15a in human thyroid cancer TPC-1 cells. Artif Cells Nanomed. Biotechnol. 2019, 47, 1815–1822. [Google Scholar] [CrossRef]
- Xiao, Y.; Jiao, C.; Lin, Y.; Chen, M.; Zhang, J.; Wang, J.; Zhang, Z. lncRNA UCA1 Contributes to Imatinib Resistance by Acting as a ceRNA Against miR-16 in Chronic Myeloid Leukemia Cells. DNA Cell Biol. 2017, 36, 8–25. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.Y.; Bai, W.D.; Ye, X.M.; Yang, A.G.; Jia, L.T. Long non-coding RNA UCA1 desensitizes breast cancer cells to trastuzumab by impeding miR-18a repression of Yes-associated protein 1. Biochem. Biophys. Res. Commun. 2018, 496, 1308–1313. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Yuan, Y.; Li, Z.; Gao, M.; Lu, Y.; Gao, H. LncRNA UCA1 sponges miR-26a to regulate the migration and proliferation of vascular smooth muscle cells. Gene 2018, 673, 159–166. [Google Scholar] [CrossRef]
- Fang, Q.; Chen, X.; Zhi, X. Long non-coding RNA (LncRNA) urothelial carcinoma associated 1 (UCA1) increases multi-drug resistance of gastric cancer via downregulating miR-27b. Med. Sci. Monit. 2016, 22, 3506–3513. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Chen, M.; Shen, Z.; Wang, R.; Fang, X.; Song, B. LncRNA-UCA1 modulates progression of colon cancer through regulating the miR-28-5p/HOXB3 axis. J. Cell Biochem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, Y.; Ding, W.; Hua, Z.; Wang, L.; Zhu, Y.; Qian, H.; Dai, T. LncRNA UCA1 impacts cell proliferation, invasion, and migration of pancreatic cancer through regulating miR-96/FOXO3. Iubmb Life 2018, 70, 276–290. [Google Scholar] [CrossRef]
- Gong, J.; Lu, X.; Xu, J.; Xiong, W.; Zhang, H.; Yu, X. Coexpression of UCA1 and ITGA2 in pancreatic cancer cells target the expression of miR-107 through focal adhesion pathway. J. Cell Physiol. 2019, 234, 12884–12896. [Google Scholar] [CrossRef]
- Zhou, Y.; Meng, X.; Chen, S.; Li, W.; Li, D.; Singer, R.; Gu, W. IMP1 regulates UCA1-mediated cell invasion through facilitating UCA1 decay and decreasing the sponge effect of UCA1 for miR-122-5p. Breast Cancer Res. 2018, 20, 32. [Google Scholar] [CrossRef]
- Zhang, T.H.; Liang, L.Z.; Liu, X.L.; Wu, J.N.; Su, K.; Chen, J.Y.; Zheng, Q.Y. LncRNA UCA1/miR-124 axis modulates TGFβ1-induced epithelial-mesenchymal transition and invasion of tongue cancer cells through JAG1/Notch signaling. J. Cell Biochem. 2019, 120, 10495–10504. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Xu, X. Knockdown of LncRNA-UCA1 suppresses chemoresistance of pediatric AML by inhibiting glycolysis through the microRNA-125a/hexokinase 2 pathway. J. Cell Biochem. 2018, 119, 6296–6308. [Google Scholar] [CrossRef]
- Sun, M.D.; Zheng, Y.Q.; Wang, L.P.; Zhao, H.T.; Yang, S. Long noncoding RNA UCA1 promotes cell proliferation, migration and invasion of human leukemia cells via sponging miR-126. Eur. Rev. Med. Pharm. Sci. 2018, 22, 2233–2245. [Google Scholar]
- Liu, Q.; Li, Y.; Lv, W.; Zhang, G.; Tian, X.; Li, X.; Cheng, H.; Zhu, C. UCA1 promotes cell proliferation and invasion and inhibits apoptosis through regulation of the miR129-SOX4 pathway in renal cell carcinoma. Onco. Targets 2018, 11, 2475–2487. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gao, F.; Zhou, L.; Wang, H.; Shi, G.; Tan, X. UCA1 Regulates the Growth and Metastasis of Pancreatic Cancer by Sponging miR-135a. Oncol. Res. 2017, 25, 1529–1541. [Google Scholar] [CrossRef] [PubMed]
- Tuo, Y.L.; Li, X.M.; Luo, J. Long noncoding RNA UCA1 modulates breast cancer cell growth and apoptosis through decreasing tumor suppressive miR-143. Eur. Rev. Med. Pharm. Sci. 2015, 19, 3403–3411. [Google Scholar]
- Li, D.; Li, H.; Yang, Y.; Kang, L. Long Noncoding RNA Urothelial Carcinoma-Associated 1 Promotes the Proliferation and Metastasis of Human Lung Tumor Cells by Regulating MicroRNA-144. Oncol. Res. 2018, 26, 537–546. [Google Scholar] [CrossRef]
- Xue, M.; Pang, H.; Li, X.; Li, H.; Pan, J.; Chen, W. Long non-coding RNA urothelial cancer-associated 1 promotes bladder cancer cell migration and invasion by way of the hsa-miR-145-ZEB1/2-FSCN1 pathway. Cancer Sci. 2016, 107, 18–27. [Google Scholar] [CrossRef]
- He, Z.; You, C.; Zhao, D. Long non-coding RNA UCA1/miR-182/PFKFB2 axis modulates glioblastoma-associated stromal cells-mediated glycolysis and invasion of glioma cells. Biochem. Biophys. Res. Commun. 2018, 500, 569–576. [Google Scholar] [CrossRef]
- Zhou, G.; Li, C.; Feng, J.; Zhang, J.; Fang, Y. lncRNA UCA1 Is a Novel Regulator in Cardiomyocyte Hypertrophy through Targeting the miR-184/HOXA9 Axis. Cardiorenal Med. 2018, 8, 130–139. [Google Scholar] [CrossRef]
- Wu, H.; Zhou, C. Long non-coding RNA UCA1 promotes lung cancer cell proliferation and migration via microRNA-193a/HMGB1 axis. Biochem. Biophys. Res. Commun. 2018, 496, 738–745. [Google Scholar] [CrossRef]
- Li, H.J.; Sun, X.M.; Li, Z.K.; Yin, Q.W.; Pang, H.; Pan, J.J.; Li, X.; Chen, W. LncRNA UCA1 promotes mitochondrial function of bladder cancer via the miR-195/ARL2 signaling pathway. Cell Physiol. Biochem. 2017, 43, 2548–2561. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, C. Silence of long non-coding RNA UCA1 inhibits hemangioma cells growth, migration and invasion by up-regulation of miR-200c. Life Sci. 2019, 226, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.N.; Yan, T.H.; Yu, R.M.; Gao, Y.; Zeng, W.L.; Lu, S.W.; Que, H.X.; Liu, Z.P.; Jiang, J.H. Long non-coding RNA UCA1 regulates the expression of Snail2 by miR-203 to promote hepatocellular carcinoma progression. J. Cancer Res. Clin. Oncol. 2017, 143, 981–990. [Google Scholar] [CrossRef]
- Bian, Z.; Jin, L.; Zhang, J.; Yin, Y.; Quan, C.; Hu, Y.; Feng, Y.; Liu, H.; Fei, B.; Mao, Y.; et al. LncRNA-UCA1 enhances cell proliferation and 5-fluorouracil resistance in colorectal cancer by inhibiting miR-204-5p. Sci. Rep. 2016, 6, 23892. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Tian, Y.; Hao, F. Downregulation of lncRNA UCA1 inhibits proliferation and invasion of cervical cancer cells through miR-206 expression. Oncol. Res. 2018. [CrossRef] [PubMed]
- Wang, C.J.; Zhu, C.C.; Xu, J.; Wang, M.; Zhao, W.Y.; Liu, Q.; Zhao, G.; Zhang, Z.Z. The lncRNA UCA1 promotes proliferation, migration, immune escape and inhibits apoptosis in gastric cancer by sponging anti-tumor miRNAs. Mol. Cancer 2019, 18, 115. [Google Scholar] [CrossRef]
- Wang, F.; Ying, H.Q.; He, B.S.; Pan, Y.Q.; Deng, Q.W.; Sun, H.L.; Chen, J.; Liu, X.; Wang, S.K. Upregulated lncRNA-UCA1 contributes to progression of hepatocellular carcinoma through inhibition of miR-216b and activation of FGFR1/ERK signaling pathway. Oncotarget 2015, 6, 7899–7917. [Google Scholar] [CrossRef]
- Guo, N.; Sun, Q.; Fu, D.; Zhang, Y. Long non-coding RNA UCA1 promoted the growth of adrenocortical cancer cells via modulating the miR-298-CDK6 axis. Gene 2019, 703, 26–34. [Google Scholar] [CrossRef]
- Zhu, G.; Liu, X.; Su, Y.; Kong, F.; Hong, X.; Lin, Z. Knockdown of urothelial carcinoma-associated 1 suppressed cell growth and migration through regulating miR-301a and CXCR4 in osteosarcoma MHCC97 cells. Oncol. Res. 2018, 27, 55–64. [Google Scholar] [CrossRef]
- Yang, Y.; Jiang, Y.; Wan, Y.; Zhang, L.; Qiu, J.; Zhou, S.; Cheng, W. UCA1 functions as a competing endogenous RNA to suppress epithelial ovarian cancer metastasis. Tumour Biol. 2016, 37, 10633–10641. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, W.G.; Lu, J.H.; Liu, Z.J.; Li, H.B.; Liu, G.J.; She, H.Y.; Li, G.Y.; Shi, X.H. LncRNA UCA1 promotes renal cell carcinoma proliferation through epigenetically repressing p21 expression and negatively regulating miR-495. Tumour Biol. 2017, 39, 1010428317701632. [Google Scholar] [CrossRef]
- Wang, P.; Liu, X.; Han, G.; Dai, S.; Ni, Q.; Xiao, S.; Huang, J. Downregulated lncRNA UCA1 acts as ceRNA to adsorb microRNA-498 to repress proliferation, invasion and epithelial mesenchymal transition of esophageal cancer cells by decreasing ZEB2 expression. Cell Cycle 2019, 18, 2359–2376. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Yang, P.; Jiang, X.; Li, X.; Wang, Y.; Zhang, X.; Sun, B.; Zhang, Y.; Jia, Y. Genetic and epigenetic silencing of mircoRNA-506-3p enhances COTL1 oncogene expression to foster non-small lung cancer progression. Oncotarget 2017, 8, 644–657. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Sun, Q.; Zhao, L.; Wu, J.; Chen, X.; Wang, Y.; Zang, W.; Zhao, G. LncRNA UCA1-miR-507-FOXM1 axis is involved in cell proliferation, invasion and G0/G1 cell cycle arrest in melanoma. Med. Oncol. 2016, 33, 88. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, W.; Ning, J.; Yu, W.; Rao, T.; Cheng, F. Long noncoding RNA UCA1 targets miR-582-5p and contributes to the progression and drug resistance of bladder cancer cells through ATG7-mediated autophagy inhibition. Onco. Targets 2019, 12, 495–508. [Google Scholar] [CrossRef]
- Gu, L.; Lu, L.S.; Zhou, D.L.; Liu, Z.C. UCA1 promotes cell proliferation and invasion of gastric cancer by targeting CREB1 sponging to miR-590-3p. Cancer Med. 2018, 7, 1253–1263. [Google Scholar] [CrossRef]
- Razmara, E.; Bitaraf, A.; Yousefi, H.; Nguyen, T.H.; Garshasbi, M.; Cho, W.C.-S.; Babashah, S. Non-coding RNAs in cartilage development: An updated review. Int. J. Mol. Sci. 2019, 20, 18. [Google Scholar] [CrossRef]
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J.P. Osteoarthritis. Nat. Rev. Dis. Primers 2016, 2, 16072. [Google Scholar] [CrossRef]
- Rollín, R.; Alvarez-Lafuente, R.; Marco, F.; López-Durán, L.; Hoyas, J.A.; Jover, J.A.; Fernández-Gutiérrez, B. The ubiquitin-proteasome pathway and viral infections in articular cartilage of patients with osteoarthritis. Rheumatol. Int. 2009, 29, 969–972. [Google Scholar] [CrossRef]
- Bendiksen, S.; Martinez-Zubiavrra, I.; Tümmler, C.; Knutsen, G.; Elvenes, J.; Olsen, E.; Olsen, R.; Moens, U. Human endogenous retrovirus W activity in cartilage of osteoarthritis patients. Biomed. Res. Int. 2014, 2014, 698609. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, X.; Dai, L.; Hu, X.; Zhu, J.; Li, L.; Zhou, C.; Ao, Y. Long noncoding RNA related to cartilage injury promotes chondrocyte extracellular matrix degradation in osteoarthritis. Arthritis Rheumatol 2015, 66, 969–978. [Google Scholar] [CrossRef]
- Wang, G.; Bu, X.; Zhang, Y.; Zhao, X.; Kong, Y.; Ma, L.; Niu, S.; Wu, B.; Meng, C. LncRNA-UCA1 enhances MMP-13 expression by inhibiting miR- 204-5p in human chondrocytes. Oncotarget 2017, 8, 91281–91290. [Google Scholar] [CrossRef] [PubMed]
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef]
- Iwakura, Y.; Saijo, S.; Kioka, Y.; Nakayama-Yamada, J.; Itagaki, K.; Tosu, M.; Asano, M.; Kanai, Y.; Kakimoto, K. Autoimmunity induction by human T cell leukemia virus type 1 in transgenic mice that develop chronic inflammatory arthropathy resembling rheumatoid arthritis in humans. J. Immunol. 1995, 155, 1588–1598. [Google Scholar] [PubMed]
- Seidl, C.; Donner, H.; Petershofen, E.; Usadel, K.H.; Seifried, E.; Kaltwasser, J.P.; Badenhoop, K. An endogenous retroviral long terminal repeat at the HLA-DQB1 gene locus confers susceptibility to rheumatoid arthritis. Hum. Immunol. 1999, 60, 63–68. [Google Scholar] [CrossRef]
- Stransky, G.; Vernon, J.; Aicher, W.K.; Moreland, L.W.; Gay, R.E.; Gay, S. Virus-like particles in synovial fluids from patients with rheuma- toid arthritis. Br. J. Rheumatol. 1993, 32, 1044–1048. [Google Scholar] [CrossRef]
- Neidhart, M.; Rethage, J.; Kuchen, S.; Künzler, P.; Crowl, R.M.; Billingham, M.E.; Gay, R.E.; Gay, S. Retrotransposable L1 elements expressed in rheumatoid arthritis synovial tissue: Association with genomic DNA hypomethylation and influence on gene expression. Arthritis Rheum. 2000, 43, 2634–2647. [Google Scholar] [CrossRef]
- Piednoël, M.; Gonçalves, I.R.; Higuet, D.; Bonnivard, E. Eukaryote DIRS1-like retrotransposons: An overview. BMC Genom. 2011, 12, 621. [Google Scholar]
- Nefedova, L.N.; Kim, A.I. Mechanisms of LTR-retroelement transposition: Lessons from Drosophila melanogaster. Viruses 2017, 9, 81. [Google Scholar] [CrossRef]
- Kim, V.N.; Han, J.; Siomi, M.C. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 2009, 10, 126–139. [Google Scholar] [CrossRef]
- Iwasaki, Y.W.; Siomi, M.C.; Siomi, H. PIWI-Interacting RNA: Its Biogenesis and Functions. Annu. Rev. Biochem. 2015, 84, 405–433. [Google Scholar] [CrossRef]
- Cardona-Maya, W.; Velilla, P.A.; Montoya, C.J.; Cadavid, Á.; Rugeles, M.T. In vitro human immunodeficiency virus and sperm cell interaction mediated by the mannose receptor. J. Reprod. Immunol. 2011, 92, 1–7. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubota, S.; Ishikawa, T.; Kawata, K.; Hattori, T.; Nishida, T. Retrotransposons Manipulating Mammalian Skeletal Development in Chondrocytes. Int. J. Mol. Sci. 2020, 21, 1564. https://doi.org/10.3390/ijms21051564
Kubota S, Ishikawa T, Kawata K, Hattori T, Nishida T. Retrotransposons Manipulating Mammalian Skeletal Development in Chondrocytes. International Journal of Molecular Sciences. 2020; 21(5):1564. https://doi.org/10.3390/ijms21051564
Chicago/Turabian StyleKubota, Satoshi, Takanori Ishikawa, Kazumi Kawata, Takako Hattori, and Takashi Nishida. 2020. "Retrotransposons Manipulating Mammalian Skeletal Development in Chondrocytes" International Journal of Molecular Sciences 21, no. 5: 1564. https://doi.org/10.3390/ijms21051564
APA StyleKubota, S., Ishikawa, T., Kawata, K., Hattori, T., & Nishida, T. (2020). Retrotransposons Manipulating Mammalian Skeletal Development in Chondrocytes. International Journal of Molecular Sciences, 21(5), 1564. https://doi.org/10.3390/ijms21051564