The Central Role of Non-Structural Protein 1 (NS1) in Influenza Biology and Infection

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. The Influenza Virus
3. Non-Structural Protein 1
3.1. Structure and Function
3.2. Post-Translational Modifications: Phosphorylation, SUMOylation, and ISGylation
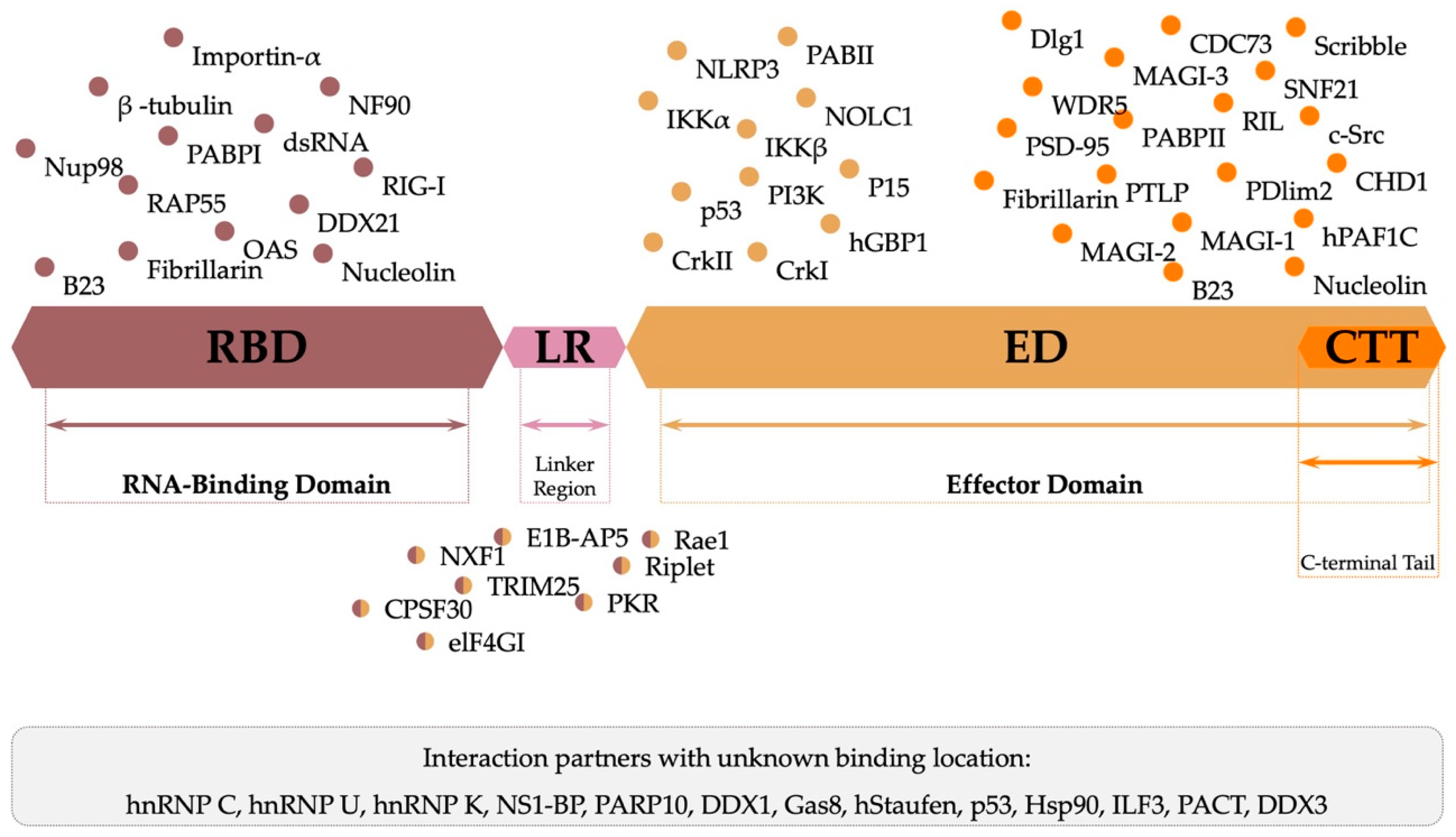
3.3. Function and Protein–Protein Interactions
3.4. NS1: Target to Manage Influenza Outbreaks
3.4.1. NS1-Modified Virus as Vaccines
3.4.2. Experimental and In Silico Approaches Towards a New Therapeutic Path
3.4.3. NS1’s Role in Influenza’s Host Tropism
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thompson, W.W.; Weintraub, E.; Dhankhar, P.; Cheng, P.-Y.; Brammer, L.; Meltzer, M.I.; Bresee, J.S.; Shay, D.K. Estimates of US Influenza-Associated Deaths Made Using Four Different Methods. Influenza Other Respir. Viruses 2009, 3, 37–49. [Google Scholar] [CrossRef]
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Estimates of Global Seasonal Influenza-Associated Respiratory Mortality: A Modelling Study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef]
- Molinari, N.-A.; Ortega-Sanchez, I.R.; Messonnier, M.L.; Thompson, W.W.; Wortley, P.M.; Weintraub, E.; Bridges, C.B. The Annual Impact of Seasonal Influenza in the US: Measuring Disease Burden and Costs. Vaccine 2007, 25, 5086–5096. [Google Scholar] [CrossRef]
- Webster, R.G.; Monto, A.S.; Braciale, T.J.; Lamb, R.A. Textbook of Influenza; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Zambon, M. Influenza and Other Emerging Respiratory Viruses. Medicine 2014, 42, 45–51. [Google Scholar] [CrossRef]
- Nicoll, A.; Ciancio, B.C.; Lopez Chavarrias, V.; Mølbak, K.; Pebody, R.; Pedzinski, B.; Penttinen, P.; van der Sande, M.; Snacken, R.; van Kerkhove, M.D. Influenza-Related Deaths. Available Methods for Estimating Numbers and Detecting Patterns for Seasonal and Pandemic Influenza in Europe. Eurosurveillance 2012, 17, 1–13. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bisen, P.S.; Raghuvanshi, R. Emerging Epidemics: Management and Control, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2013. [Google Scholar]
- Centers for Disease Control and Prevention. How Is Pandemic Flu Different from Seasonal Flu? Available online: https://www.cdc.gov/flu/pandemic-resources/basics/about.html (accessed on 15 June 2019).
- Mubareka, S.; Palese, P. Influenza Virus: The Biology of a Changing Virus. In Influenza Vaccines for the Future; Rappuoli, R., Del Giudice, G., Eds.; Springer Basel: Basel, Switzerland, 2011; pp. 3–26. [Google Scholar]
- Carrat, F.; Flahaul, A. Influenza Vaccine: The Challenge of Antigenic Drift. Vaccine 2007, 25, 6852–6862. [Google Scholar] [CrossRef] [PubMed]
- Paules, C.; Subbarao, K. Influenza. Lancet 2017, 390, 697–708. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Past Pandemics. Available online: https://www.cdc.gov/flu/pandemic-resources/basics/past-pandemics.html (accessed on 15 June 2019).
- Hussain, M.; Galvin, H.; Haw, T.Y.; Nutsford, A.; Husain, M. Drug Resistance in Influenza A Virus: The Epidemiology and Management. Infect. Drug Resist. 2017, 10, 121–134. [Google Scholar] [CrossRef]
- Kilbourne, E.D. Influenza Pandemics of the 20th Century. Emerg. Infect. Dis. 2006, 12, 9–14. [Google Scholar] [CrossRef]
- Dawood, F.S.; Iuliano, A.D.; Reed, C.; Meltzer, M.I.; Shay, D.K.; Cheng, P.-Y.; Bandaranayake, D.; Breiman, R.F.; Brooks, W.A.; Buchy, P.; et al. Estimated Global Mortality Associated with the First 12 Months of 2009 Pandemic Influenza A H1N1 Virus Circulation: A Modelling Study. Lancet Infect. Dis. 2012, 12, 687–695. [Google Scholar] [CrossRef]
- Jiang, W.; Hou, G.; Li, J.; Peng, C.; Wang, S.; Liu, S.; Zhuang, Q.; Yuan, L.; Yu, X.; Li, Y.; et al. Antigenic Variant of Highly Pathogenic Avian Influenza A(H7N9) Virus, China, 2019. Emerg. Infect. Dis. 2020, 26, 379–380. [Google Scholar] [CrossRef] [PubMed]
- Aoki, F.Y.; Allen, U.D.; Stiver, H.G.; Evans, G.A. The Use of Antiviral Drugs for Influenza: Guidance for Practitioners 2012/2013. Can. J. Infect. Dis. Med. Microbiol. 2012, 23, e79–e92. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hsieh, H.-P.; Hsu, J. Strategies of Development of Antiviral Agents Directed Against Influenza Virus Replication. Curr. Pharm. Des. 2007, 13, 3531–3542. [Google Scholar] [CrossRef] [PubMed]
- Gasparini, R.; Amicizia, D.; Lai, P.L.; Bragazzi, N.L.; Panatto, D. Compounds with Anti-Influenza Activity: Present and Future of Strategies for the Optimal Treatment and Management of Influenza. Part II: Future Compounds against Influenza Virus. J. Prev. Med. Hyg. 2014, 55, 109–129. [Google Scholar]
- Das, K.; Aramini, J.M.; Ma, L.-C.; Krug, R.M.; Arnold, E. Structures of Influenza A Proteins and Insights into Antiviral Drug Targets. Nat. Struct. Mol. Biol. 2010, 17, 530–538. [Google Scholar] [CrossRef]
- Stadlbauer, D.; Zhu, X.; McMahon, M.; Turner, J.S.; Wohlbold, T.J.; Schmitz, A.J.; Strohmeier, S.; Yu, W.; Nachbagauer, R.; Mudd, P.A.; et al. Broadly Protective Human Antibodies That Target the Active Site of Influenza Virus Neuraminidase. Science 2019, 366, 499–504. [Google Scholar] [CrossRef]
- Hu, Y.; Sneyd, H.; Dekant, R.; Wang, J. Functional Viral Protein As a Hot Antiviral Drug Target. Curr. Top. Med. Chem. 2018, 17, 2271–2285. [Google Scholar]
- Beigel, J.H.; Hayden, F.G. Influenza Therapeutics in Clinical Practice—Challenges and Recent Advances. Cold Spring Harb. Perspect. Med. 2020, a038463. [Google Scholar] [CrossRef]
- Engel, D.A. The Influenza Virus NS1 Protein as a Therapeutic Target. Antivir. Res. 2013, 99, 409–416. [Google Scholar] [CrossRef]
- Talon, J.; Horvath, C.M.; Polley, R.; Basler, C.F.; Muster, T.; Palese, P.; Garcia-Sastre, A. Activation of Interferon Regulatory Factor 3 Is Inhibited by the Influenza A Virus NS1 Protein. J. Virol. 2000, 74, 7989–7996. [Google Scholar] [CrossRef]
- García-Sastre, A. Induction and Evasion of Type I Interferon Responses by Influenza Viruses. Virus Res. 2011, 162, 12–18. [Google Scholar] [CrossRef] [PubMed]
- García-Sastre, A. Inhibition of Interferon-Mediated Antiviral Responses by Influenza A Viruses and Other Negative-Strand RNA Viruses. Virology 2001, 279, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Gasparini, R.; Amicizia, D.; Lai, P.L.; Bragazzi, N.L.; Panatto, D. Compounds with Anti-Influenza Activity: Present and Future of Strategies for the Optimal Treatment and Management of Influenza. Part I: Influenza Life-Cycle and Currently Available Drugs. J. Prev. Med. Hyg. 2014, 55, 69–85. [Google Scholar] [PubMed]
- Rappuoli, R.; Del Giudice, G. Influenza Vaccines for the Future; Springer Science & Business Media: Berlin, Germany, 2010; p. 363. [Google Scholar] [CrossRef]
- CDC. Influenza, 13th ed.; Hamborsky, J., Kroger, A., Wolfe, S., Eds.; Public Health Foundation: Washington, DC, USA, 2015.
- Abbas, M.; Abidin, Z.U. Proteins of Influenza Virus: A Review. J. Infect. Mol. Biol. 2013, 1, 1–7. [Google Scholar]
- Samji, T. Influenza A: Understanding the Viral Life Cycle. Yale J. Biol. Med. 2009, 82, 153–159. [Google Scholar]
- Anastasina, M. Influenza A Virus-Host Interactions and Their Control by Viral Non-Structural Protein NS1. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2015. [Google Scholar]
- Eisfeld, A.J.; Neumann, G.; Kawaoka, Y. At the Centre: Influenza A Virus Ribonucleoproteins. Nat. Rev. Microbiol. 2015, 13, 28–41. [Google Scholar] [CrossRef]
- Xia, S. High Throughput Screening of Inhibitors for Influenza Protein NS1. Ph.D. Thesis, University of Texas at Austin, Austin, TX, USA, 2009. [Google Scholar]
- Du, J.; Cross, T.A.; Zhou, H.X. Recent Progress in Structure-Based Anti-Influenza Drug Design. Drug Discov. Today 2012, 17, 1111–1120. [Google Scholar] [CrossRef]
- Long, J.C.D.; Fodor, E. The PB2 Subunit of the Influenza A Virus RNA Polymerase Is Imported into the Mitochondrial Matrix. J. Virol. 2016, 90, 8729–8738. [Google Scholar] [CrossRef]
- Chen, W.; Calvo, P.A.; Malide, D.; Gibbs, J.; Schubert, U.; Bacik, I.; Basta, S.; O’Neill, R.; Schickli, J.; Palese, P.; et al. A Novel Influenza A Virus Mitochondrial Protein That Induces Cell Death. Nat. Med. 2001, 7, 1306–1312. [Google Scholar] [CrossRef]
- Hale, B.G.; Batty, I.H.; Downes, C.P.; Randall, R.E. Binding of Influenza A Virus NS1 Protein to the Inter-SH2 Domain of P85β Suggests a Novel Mechanism for Phosphoinositide 3-Kinase Activation. J. Biol. Chem. 2008, 283, 1372–1380. [Google Scholar] [CrossRef]
- Trigueiro-Louro, J.M.; Correia, V.; Santos, L.A.; Guedes, R.C.; Brito, R.M.M.; Rebelo-de-Andrade, H. To Hit or Not to Hit: Large-Scale Sequence Analysis and Structure Characterization of Influenza A NS1 Unlocks New Antiviral Target Potential. Virology 2019, 535, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, B.; Choi, J.-M.; Bornholdt, Z.A.; Sankaran, B.; Rice, A.P.; Prasad, B.V.V. The Influenza A Virus Protein NS1 Displays Structural Polymorphism. J. Virol. 2014, 88, 4113–4122. [Google Scholar] [CrossRef] [PubMed]
- Bornholdt, Z.A.; Prasad, B.V.V. X-Ray Structure of Influenza Virus NS1 Effector Domain. Nat. Struct. Mol. Biol. 2006, 13, 559–560. [Google Scholar] [CrossRef] [PubMed]
- Bornholdt, Z.A.; Prasad, B.V.V. X-Ray Structure of NS1 from a Highly Pathogenic H5N1 Influenza Virus. Nature 2008, 456, 985–988. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Robertus, J.D. X-Ray Structures of NS1 Effector Domain Mutants. Arch. Biochem. Biophys. 2010, 494, 198–204. [Google Scholar] [CrossRef]
- Yin, C.; Khan, J.A.; Swapna, G.V.T.; Ertekin, A.; Krug, R.M.; Tong, L.; Montelione, G.T. Conserved Surface Features Form the Double-Stranded RNA Binding Site of Non-Structural Protein 1 (NS1) from Influenza A and B Viruses. J. Biol. Chem. 2007, 282, 20584–20592. [Google Scholar] [CrossRef]
- Shen, Q.; Cho, J.-H. The Structure and Conformational Plasticity of the Nonstructural Protein 1 of the 1918 Influenza A Virus. Biochem. Biophys. Res. Commun. 2019, 518, 178–182. [Google Scholar] [CrossRef]
- Jureka, A.S.; Kleinpeter, A.B.; Cornilescu, G.; Cornilescu, C.C.; Petit, C.M. Structural Basis for a Novel Interaction between the NS1 Protein Derived from the 1918 Influenza Virus and RIG-I. Structure 2015, 23, 2001–2010. [Google Scholar] [CrossRef]
- Aramini, J.M.; Ma, L.; Lee, H.; Zhao, L.; Cunningham, K.; Ciccosanti, C.; Janjua, H.; Fang, Y.; Xiao, R.; Krug, R.M. Solution NMR Structure of the Monomeric W187R Mutant of A/Udorn NS1 Effector Domain. Northeast Struct. Genomics Target OR8C [W187R] J. 2009. [Google Scholar] [CrossRef]
- Tamada, T.; Kitadokoro, K.; Higuchi, Y.; Inaka, K.; Yasui, A.; de Ruiter, P.; Eker, A.P.M.; Miki, K.; Chien, C.Y.; Tejero, R.; et al. A Novel RNA-Binding Motif in Influenza A Virus Non-Structural Protein 1. Nat. Struct. Biol. 1997, 4, 891–895. [Google Scholar]
- Chien, C.; Xu, Y.; Xiao, R.; Aramini, J.M.; Sahasrabudhe, P.V.; Krug, R.M.; Montelione, G.T. Biophysical Characterization of the Complex between Double-Stranded RNA and the N-Terminal Domain of the NS1 Protein from Influenza A Virus: Evidence for a Novel RNA-Binding Mode. Biochemistry 2004, 43, 1950–1962. [Google Scholar] [CrossRef] [PubMed]
- Darapaneni, V.; Prabhaker, V.K.; Kukol, A. Large-Scale Analysis of Influenza A Virus Sequences Reveals Potential Drug Target Sites of Non-Structural Proteins. J. Gen. Virol. 2009, 90, 2124–2133. [Google Scholar] [CrossRef] [PubMed]
- Nemeroff, M.E.; Qian, X.-Y.; Krug, R.M. The Influenza Virus NS1 Protein Forms Multimers in Vitro and in Vivo. Virology 1995, 212, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Kumar, D.; Hu, L.; Sankaran, B.; Moosa, M.M.; Rice, A.P.; Ferreon, J.C.; Ferreon, A.C.M.; Prasad, B.V.V. Influenza A Virus Protein NS1 Exhibits Strain-Independent Conformational Plasticity. J. Virol. 2019. [Google Scholar] [CrossRef]
- Klemm, C.; Boergeling, Y.; Ludwig, S.; Ehrhardt, C. Immunomodulatory Nonstructural Proteins of Influenza A Viruses. Trends Microbiol. 2018, 26, 624–636. [Google Scholar] [CrossRef]
- Krug, R.M.; García-Sastre, A. The NS1 Protein: A Master Regulator of Host and Viral Functions. In Textbook of Influenza; Webster, R.G., Monto, A.S., Braciale, T.J., Lamb, R.A., Eds.; John Wiley & Sons, Ltd.: Oxford, UK, 2013. [Google Scholar]
- Hale, B.G.; Knebel, A.; Botting, C.H.; Galloway, C.S.; Precious, B.L.; Jackson, D.; Elliott, R.M.; Randall, R.E. CDK/ERK-Mediated Phosphorylation of the Human Influenza A Virus NS1 Protein at Threonine-215. Virology 2009, 383, 6–11. [Google Scholar] [CrossRef]
- Hsiang, T.-Y.; Zhou, L.; Krug, R.M. Roles of the Phosphorylation of Specific Serines and Threonines in the NS1 Protein of Human Influenza A. Viruses. J. Virol. 2012, 86, 10370–10376. [Google Scholar] [CrossRef]
- Kathum, O.A.; Schräder, T.; Anhlan, D.; Nordhoff, C.; Liedmann, S.; Pande, A.; Mellmann, A.; Ehrhardt, C.; Wixler, V.; Ludwig, S. Phosphorylation of Influenza A Virus NS1 Protein at Threonine 49 Suppresses Its Interferon Antagonistic Activity. Cell. Microbiol. 2016, 18, 784–791. [Google Scholar] [CrossRef]
- Zheng, W.; Cao, S.; Chen, C.; Li, J.; Zhang, S.; Jiang, J.; Niu, Y.; Fan, W.; Li, Y.; Bi, Y.; et al. Threonine 80 Phosphorylation of Non-Structural Protein 1 Regulates the Replication of Influenza A Virus by Reducing the Binding Affinity with RIG-I. Cell. Microbiol. 2017, 19, e12643. [Google Scholar] [CrossRef]
- Hale, B.G.; Randall, R.E.; Ortin, J.; Jackson, D. The Multifunctional NS1 Protein of Influenza A Viruses. J. Gen. Virol. 2008, 89, 2359–2376. [Google Scholar] [CrossRef]
- Hay, R.T. SUMO. Mol. Cell 2005, 18, 1–12. [Google Scholar] [CrossRef]
- Xu, K.; Klenk, C.; Liu, B.; Keiner, B.; Cheng, J.; Zheng, B.-J.; Li, L.; Han, Q.; Wang, C.; Li, T.; et al. Modification of Nonstructural Protein 1 of Influenza A Virus by SUMO1. J. Virol. 2011, 85, 1086–1098. [Google Scholar] [CrossRef]
- Santos, A.; Pal, S.; Chacon, J.; Meraz, K.; Gonzalez, J.; Prieto, K.; Rosas-Acosta, G. SUMOylation Affects the Interferon Blocking Activity of the Influenza A Nonstructural Protein NS1 without Affecting Its Stability or Cellular Localization. J. Virol. 2013, 87, 5602–5620. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Marc, D. Influenza Virus Non-Structural Protein NS1: Interferon Antagonism and Beyond. J. Gen. Virol. 2014, 95, 2594–2611. [Google Scholar] [CrossRef] [PubMed]
- Melen, K.; Kinnunen, L.; Fagerlund, R.; Ikonen, N.; Twu, K.Y.; Krug, R.M.; Julkunen, I. Nuclear and Nucleolar Targeting of Influenza A Virus NS1 Protein: Striking Differences between Different Virus Subtypes. J. Virol. 2007, 81, 5995–6006. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Hsiang, T.-Y.; Kuo, R.-L.; Krug, R.M. ISG15 Conjugation System Targets the Viral NS1 Protein in Influenza A Virus–Infected Cells. Proc. Natl. Acad. Sci. USA 2010, 107, 2253–2258. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhong, G.; Zhu, L.; Liu, X.; Shan, Y.; Feng, H.; Bu, Z.; Chen, H.; Wang, C. Herc5 Attenuates Influenza A Virus by Catalyzing ISGylation of Viral NS1 Protein. J. Immunol. 2010, 184, 5777–5790. [Google Scholar] [CrossRef]
- Yuan, W. Influenza B Virus NS1 Protein Inhibits Conjugation of the Interferon (IFN)-Induced Ubiquitin-like ISG15 Protein. EMBO J. 2001, 20, 362–371. [Google Scholar] [CrossRef]
- Greenspan, D.; Palese, P.; Krystal, M. Two Nuclear Location Signals in the Influenza Virus NS1 Nonstructural Protein. J. Virol. 1988, 62, 3020–3026. [Google Scholar] [CrossRef]
- Melén, K.; Tynell, J.; Fagerlund, R.; Roussel, P.; Hernandez-Verdun, D.; Julkunen, I. Influenza A H3N2 Subtype Virus NS1 Protein Targets into the Nucleus and Binds Primarily via Its C-Terminal NLS2/NoLS to Nucleolin and Fibrillarin. Virol. J. 2012, 9, 167. [Google Scholar] [CrossRef]
- Li, Y.; Yamakita, Y.; Krug, R.M. Regulation of a Nuclear Export Signal by an Adjacent Inhibitory Sequence: The Effector Domain of the Influenza Virus NS1 Protein. Proc. Natl. Acad. Sci. USA 1998, 95, 4864–4869. [Google Scholar] [CrossRef] [PubMed]
- Pichlmair, A.; Schulz, O.; Tan, C.P.; Naslund, T.I.; Liljestrom, P.; Weber, F.; Reis e Sousa, C. RIG-I-Mediated Antiviral Responses to Single-Stranded RNA Bearing 5′-Phosphates. Science 2006, 314, 997–1001. [Google Scholar] [CrossRef] [PubMed]
- Jahan, A.S.; Biquand, E.; Muñoz-Moreno, R.; Le Quang, A.; Mok, C.K.-P.; Wong, H.H.; Teo, Q.W.; Valkenburg, S.A.; Chin, A.W.H.; Man Poon, L.L.; et al. OTUB1 Is a Key Regulator of RIG-I-Dependent Immune Signaling and Is Targeted for Proteasomal Degradation by Influenza A NS1. Cell Rep. 2020, 30, 1570–1584. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Riedel, K.; Lynch, P.; Chien, C.; Montelione, G.; Krug, R.M. RNA Binding by the Novel Helical Domain of the Influenza Virus NS1 Protein Requires Its Dimer Structure and a Small Number of Specific Basic Amino Acids. RNA 1999, 5, S1355838299981621. [Google Scholar] [CrossRef]
- Jureka, A.S.; Kleinpeter, A.B.; Tipper, J.L.; Harrod, K.S.; Petit, C.M. The Influenza NS1 Protein Modulates RIG-I Activation via a Strain-Specific Direct Interaction with the Second CARD of RIG-I. J. Biol. Chem. 2020, 295, 1153–1164. [Google Scholar] [CrossRef]
- Cheng, A.; Wong, S.M.; Yuan, Y.A. Structural Basis for DsRNA Recognition by NS1 Protein of Influenza A Virus. Cell Res. 2009, 19, 187–195. [Google Scholar] [CrossRef]
- Gack, M.U.; Albrecht, R.A.; Urano, T.; Inn, K.-S.; Huang, I.-C.; Carnero, E.; Farzan, M.; Inoue, S.; Jung, J.U.; García-Sastre, A. Influenza A Virus NS1 Targets the Ubiquitin Ligase TRIM25 to Evade Recognition by the Host Viral RNA Sensor RIG-I. Cell Host Microbe 2009, 5, 439–449. [Google Scholar] [CrossRef]
- Oshiumi, H.; Miyashita, M.; Inoue, N.; Okabe, M.; Matsumoto, M.; Seya, T. The Ubiquitin Ligase Riplet Is Essential for RIG-I-Dependent Innate Immune Responses to RNA Virus Infection. Cell Host Microbe 2010, 8, 496–509. [Google Scholar] [CrossRef]
- Martín-Vicente, M.; Medrano, L.M.; Resino, S.; García-Sastre, A.; Martínez, I. TRIM25 in the Regulation of the Antiviral Innate Immunity. Front. Immunol. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Koliopoulos, M.G.; Lethier, M.; van der Veen, A.G.; Haubrich, K.; Hennig, J.; Kowalinski, E.; Stevens, R.V.; Martin, S.R.; Reis e Sousa, C.; Cusack, S.; et al. Molecular Mechanism of Influenza A NS1-Mediated TRIM25 Recognition and Inhibition. Nat. Commun. 2018, 9, 1820. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, L.; Zhang, R.; Chi, X.; Yang, Z.; Xie, Y.; Shu, S.; Liao, Y.; Chen, J.L. Identification of Two Residues within the NS1 of H7N9 Influenza A Virus That Critically Affect the Protein Stability and Function 11 Medical and Health Sciences 1108 Medical Microbiology 06 Biological Sciences 0601 Biochemistry and Cell Biology. Vet. Res. 2018, 49, 1–14. [Google Scholar]
- Nogales, A.; Martinez-Sobrido, L.; Topham, D.J.; Dediego, M.L. Modulation of Innate Immune Responses by the Influenza a NS1 and PA-X Proteins. Viruses 2018, 10, 708. [Google Scholar] [CrossRef] [PubMed]
- Ayllon, J.; García-Sastre, A. The NS1 Protein: A Multitasking Virulence Factor. Curr. Top. Microbiol. Immunol. 2015, 386, 73–107. [Google Scholar] [PubMed]
- Robertson, J.S.; Schubert, M.; Lazzarini, R.A. Polyadenylation Sites for Influenza Virus MRNA. J. Virol. 1981, 38, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Fortes, P.; Beloso, A.; Ortín, J. Influenza Virus NS1 Protein Inhibits Pre-MRNA Splicing and Blocks MRNA Nucleocytoplasmic Transport. EMBO J. 1994, 13, 704–712. [Google Scholar] [CrossRef]
- Nemeroff, M.E.; Barabino, S.M.L.; Li, Y.; Keller, W.; Krug, R.M. Influenza Virus NS1 Protein Interacts with the Cellular 30 KDa Subunit of CPSF and Inhibits 3′ End Formation of Cellular Pre-MRNAs. Mol. Cell 1998, 1, 991–1000. [Google Scholar] [CrossRef]
- Das, K.; Ma, L.-C.; Xiao, R.; Radvansky, B.; Aramini, J.; Zhao, L.; Marklund, J.; Kuo, R.-L.; Twu, K.Y.; Arnold, E.; et al. Structural Basis for Suppression of a Host Antiviral Response by Influenza A Virus. Proc. Natl. Acad. Sci. USA 2008, 105, 13093–13098. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Z.-Y.; Wang, W.; Baker, C.C.; Krug, R.M. The 3′-End-Processing Factor CPSF Is Required for the Splicing of Single-Intron Pre-MRNAs in Vivo. RNA 2001, 7, S1355838201010226. [Google Scholar] [CrossRef]
- Chen, Z. Influenza A Virus NS1 Protein Targets Poly(A)-Binding Protein II of the Cellular 3′-End Processing Machinery. EMBO J. 2002, 18, 2273–2283. [Google Scholar] [CrossRef]
- Satterly, N.; Tsai, P.-L.; van Deursen, J.; Nussenzveig, D.R.; Wang, Y.; Faria, P.A.; Levay, A.; Levy, D.E.; Fontoura, B.M.A. Influenza Virus Targets the MRNA Export Machinery and the Nuclear Pore Complex. Proc. Natl. Acad. Sci. USA 2007, 104, 1853–1858. [Google Scholar] [CrossRef]
- Zhang, K.; Xie, Y.; Muñoz-Moreno, R.; Wang, J.; Zhang, L.; Esparza, M.; García-Sastre, A.; Fontoura, B.M.A.; Ren, Y. Structural Basis for Influenza Virus NS1 Protein Block of MRNA Nuclear Export. Nat. Microbiol. 2019, 4, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- Min, J.-Y.; Li, S.; Sen, G.C.; Krug, R.M. A Site on the Influenza A Virus NS1 Protein Mediates Both Inhibition of PKR Activation and Temporal Regulation of Viral RNA Synthesis. Virology 2007, 363, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Min, J.-Y.; Krug, R.M.; Sen, G.C. Binding of the Influenza A Virus NS1 Protein to PKR Mediates the Inhibition of Its Activation by Either PACT or Double-Stranded RNA. Virology 2006, 349, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Schierhorn, K.L.; Jolmes, F.; Bespalowa, J.; Saenger, S.; Peteranderl, C.; Dzieciolowski, J.; Mielke, M.; Budt, M.; Pleschka, S.; Herrmann, A.; et al. Influenza A Virus Virulence Depends on Two Amino Acids in the N-Terminal Domain of Its NS1 Protein To Facilitate Inhibition of the RNA-Dependent Protein Kinase PKR. J. Virol. 2017, 91, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Min, J.-Y.; Krug, R.M. The Primary Function of RNA Binding by the Influenza A Virus NS1 Protein in Infected Cells: Inhibiting the 2′-5′ Oligo (A) Synthetase/RNase L Pathway. Proc. Natl. Acad. Sci. USA 2006, 103, 7100–7105. [Google Scholar] [CrossRef]
- Ehrhardt, C.; Marjuki, H.; Wolff, T.; Nurnberg, B.; Planz, O.; Pleschka, S.; Ludwig, S. Bivalent Role of the Phosphatidylinositol-3-Kinase (PI3K) during Influenza Virus Infection and Host Cell Defence. Cell. Microbiol. 2006, 8, 1336–1348. [Google Scholar] [CrossRef]
- Hale, B.G.; Kerry, P.S.; Jackson, D.; Precious, B.L.; Gray, A.; Killip, M.J.; Randall, R.E.; Russell, R.J. Structural Insights into Phosphoinositide 3-Kinase Activation by the Influenza A Virus NS1 Protein. Proc. Natl. Acad. Sci. USA 2010, 107, 1954–1959. [Google Scholar] [CrossRef]
- Chen, G.; Liu, C.-H.; Zhou, L.; Krug, R.M. Cellular DDX21 RNA Helicase Inhibits Influenza A Virus Replication but Is Counteracted by the Viral NS1 Protein. Cell Host Microbe 2014, 15, 484–493. [Google Scholar] [CrossRef]
- Carrasco, L.; de la Luna, S.; Ortin, J.; Aragon, T.; Nieto, A.; Novoa, I. Eukaryotic Translation Initiation Factor 4GI Is a Cellular Target for NS1 Protein, a Translational Activator of Influenza Virus. Mol. Cell. Biol. 2002, 20, 6259–6268. [Google Scholar]
- Burgui, I.; Aragón, T.; Ortín, J.; Nieto, A. PABP1 and EIF4GI Associate with Influenza Virus NS1 Protein in Viral MRNA Translation Initiation Complexes. J. Gen. Virol. 2003, 84, 3263–3274. [Google Scholar] [CrossRef]
- Falcon, A.M.; Fortes, P.; Marion, R.M.; Beloso, A.; Ortin, J. Interaction of Influenza Virus NS1 Protein and the Human Homologue of Staufen in Vivo and in Vitro. Nucleic Acids Res. 1999, 27, 2241–2247. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Zheng, F.; Zhu, J.; Liu, M.; Liu, N.; Li, X.; Zhang, L.; Deng, Z.; Zhao, Q.; Liu, H. The Interaction between NOLC1 and IAV NS1 Protein Promotes Host Cell Apoptosis and Reduces Virus Replication. Oncotarget 2017, 8, 94519–94527. [Google Scholar] [CrossRef] [PubMed]
- Shapira, S.D.; Gat-Viks, I.; Shum, B.O.V.; Dricot, A.; Marciela, M.; Liguo, W.; Gupta, P.B.; Hao, T.; Silver, S.J.; David, E.; et al. A Physical and Regulatory Map of Host-Influenza Interactions Reveals Pathways in H1N1 Infection. Cell 2009, 139, 1255–1267. [Google Scholar] [CrossRef] [PubMed]
- De Chassey, B.; Aublin-Gex, A.; Ruggieri, A.; Meyniel-Schicklin, L.; Pradezynski, F.; Davoust, N.; Chantier, T.; Tafforeau, L.; Mangeot, P.-E.; Ciancia, C.; et al. The Interactomes of Influenza Virus NS1 and NS2 Proteins Identify New Host Factors and Provide Insights for ADAR1 Playing a Supportive Role in Virus Replication. PLoS Pathog. 2013, 9, e1003440. [Google Scholar] [CrossRef] [PubMed]
- Raman, S.N.T.; Zhou, Y.; Raman, S.N.T.; Thulasi Raman, S.N.; Zhou, Y. Networks of Host Factors That Interact with NS1 Protein of Influenza A Virus. Front. Microbiol. 2016, 7, 654. [Google Scholar]
- Kuo, R.L.; Li, Z.H.; Li, L.H.; Lee, K.M.; Tam, E.H.; Liu, H.M.; Liu, H.P.; Shih, S.R.; Wu, C.C. Interactome Analysis of the NS1 Protein Encoded by Influenza A H1N1 Virus Reveals a Positive Regulatory Role of Host Protein PRP19 in Viral Replication. J. Proteome Res. 2016, 15, 1639–1648. [Google Scholar] [CrossRef]
- Ylösmäki, L.; Fagerlund, R.; Kuisma, I.; Julkunen, I.; Saksela, K. Nuclear Translocation of Crk Adaptor Proteins by the Influenza A Virus NS1 Protein. Viruses 2016, 8, 101. [Google Scholar] [CrossRef]
- Marazzi, I.; Ho, J.S.Y.; Kim, J.; Manicassamy, B.; Dewell, S.; Albrecht, R.A.; Seibert, C.W.; Schaefer, U.; Jeffrey, K.L.; Prinjha, R.K.; et al. Suppression of the Antiviral Response by an Influenza Histone Mimic. Nature 2012, 483, 428–433. [Google Scholar] [CrossRef]
- Qin, S.; Liu, Y.; Tempel, W.; Eram, M.S.; Bian, C.; Liu, K.; Senisterra, G.; Crombet, L.; Vedadi, M.; Min, J. Structural Basis for Histone Mimicry and Hijacking of Host Proteins by Influenza Virus Protein NS1. Nat. Commun. 2014, 5, 3952. [Google Scholar] [CrossRef]
- Han, X.; Li, Z.; Chen, H.; Wang, H.; Mei, L.; Wu, S.; Zhang, T.; Liu, B.; Lin, X. Influenza Virus A/Beijing/501/2009(H1N1) NS1 Interacts with β-Tubulin and Induces Disruption of the Microtubule Network and Apoptosis on A549 Cells. PLoS ONE 2012, 7, e48340. [Google Scholar] [CrossRef]
- Mok, B.W.-Y.; Song, W.; Wang, P.; Tai, H.; Chen, Y.; Zheng, M.; Wen, X.; Lau, S.-Y.; Wu, W.L.; Matsumoto, K.; et al. The NS1 Protein of Influenza A Virus Interacts with Cellular Processing Bodies and Stress Granules through RNA-Associated Protein 55 (RAP55) during Virus Infection. J. Virol. 2012, 86, 12695–12707. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Li, X.; Zhu, W.; Wang, H.; Mei, L.; Wu, S.; Lin, X.; Han, X. NF90 Is a Novel Influenza A Virus NS1-interacting Protein That Antagonizes the Inhibitory Role of NS1 on PKR Phosphorylation. FEBS Lett. 2016, 590, 2797–2810. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, Y.; Zhou, X.; Yang, Z.; Liu, X.; Cao, Z.; Song, H.; He, Y.; Huang, P. The NS1 Protein of Influenza a Virus Interacts with Heat Shock Protein Hsp90 in Human Alveolar Basal Epithelial Cells: Implication for Virus-Induced Apoptosis. Virol. J. 2011, 8, 181. [Google Scholar] [CrossRef] [PubMed]
- Cheong, W.C.; Kang, H.R.; Yoon, H.; Kang, S.J.; Ting, J.P.Y.; Song, M.J. Influenza A Virus NS1 Protein Inhibits the NLRP3 Inflammasome. PLoS ONE 2015, 10, e0126456. [Google Scholar] [CrossRef]
- Gao, S.; Song, L.; Li, J.; Zhang, Z.; Peng, H.; Jiang, W.; Wang, Q.; Kang, T.; Chen, S.; Huang, W. Influenza A Virus-Encoded NS1 Virulence Factor Protein Inhibits Innate Immune Response by Targeting IKK. Cell. Microbiol. 2012, 14, 1849–1866. [Google Scholar] [CrossRef]
- Zhu, C.; Zheng, F.; Sun, T.; Duan, Y.; Cao, J.; Feng, H.; Shang, L.; Zhu, Y.; Liu, H. Interaction of Avian Influenza Virus NS1 Protein and Nucleolar and Coiled-Body Phosphoprotein 1. Virus Genes 2013, 46, 287–292. [Google Scholar] [CrossRef]
- Dubois, J.; Traversier, A.; Julien, T.; Padey, B.; Lina, B.; Bourdon, J.; Marcel, V.; Bolvin, G.; Rosa-Calatrava, M.; Terrier, O. The Nonstructural NS1 Protein of Influenza Viruses Modulates TP53 Splicing through Host Factor CPSF4. J. Virol. 2019, 93, e02168-18. [Google Scholar] [CrossRef]
- Yu, J.; Li, X.; Wang, Y.; Li, B.; Li, H.; Li, Y.; Zhou, W.; Zhang, C.; Wang, Y.; Rao, Z.; et al. PDlim2 Selectively Interacts with the PDZ Binding Motif of Highly Pathogenic Avian H5N1 Influenza A Virus NS1. PLoS ONE 2011, 6, e19511. [Google Scholar] [CrossRef]
- Liu, H.; Golebiewski, L.; Dow, E.C.; Krug, R.M.; Javier, R.T.; Rice, A.P. The ESEV PDZ-Binding Motif of the Avian Influenza A Virus NS1 Protein Protects Infected Cells from Apoptosis by Directly Targeting Scribble. J. Virol. 2010, 84, 11164–11174. [Google Scholar] [CrossRef]
- Kumar, M.; Liu, H.; Rice, A.P. Regulation of Interferon-β by MAGI-1 and Its Interaction with Influenza A Virus NS1 Protein with ESEV PBM. PLoS ONE 2012, 7, e41251. [Google Scholar] [CrossRef]
- Bavagnoli, L.; Dundon, W.G.; Garbelli, A.; Zecchin, B.; Milani, A.; Parakkal, G.; Baldanti, F.; Paolucci, S.; Volmer, R.; Tu, Y.; et al. The PDZ-Ligand and Src-Homology Type 3 Domains of Epidemic Avian Influenza Virus NS1 Protein Modulate Human Src Kinase Activity during Viral Infection. PLoS ONE 2011, 6, e27789. [Google Scholar] [CrossRef] [PubMed]
- Richt, J.A.; García-Sastre, A.; Garcia-Sastre, A. Attenuated Influenza Virus Vaccines with Modified NS1 Proteins. Curr. Top. Microbiol. Immunol. 2009, 333, 177–195. [Google Scholar]
- Wacheck, V.; Egorov, A.; Groiss, F.; Pfeiffer, A.; Fuereder, T.; Hoeflmayer, D.; Kundi, M.; Popow-Kraupp, T.; Redlberger-Fritz, M.; Mueller, C.A.; et al. A Novel Type of Influenza Vaccine: Safety and Immunogenicity of Replication-Deficient Influenza Virus Created by Deletion of the Interferon Antagonist NS1. J. Infect. Dis. 2010, 201, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Morokutti, A.; Muster, T.; Ferko, B. Intranasal Vaccination with a Replication-Deficient Influenza Virus Induces Heterosubtypic Neutralising Mucosal IgA Antibodies in Humans. Vaccine 2014, 32, 1897–1900. [Google Scholar] [CrossRef] [PubMed]
- Nicolodi, C.; Groiss, F.; Kiselev, O.; Wolschek, M.; Seipelt, J.; Muster, T. Safety and Immunogenicity of a Replication-Deficient H5N1 Influenza Virus Vaccine Lacking NS1. Vaccine 2019, 37, 3722–3729. [Google Scholar] [CrossRef]
- Mossler, C.; Groiss, F.; Wolzt, M.; Wolschek, M.; Seipelt, J.; Muster, T. Phase I/II Trial of a Replication-Deficient Trivalent Influenza Virus Vaccine Lacking NS1. Vaccine 2013, 31, 6194–6200. [Google Scholar] [CrossRef]
- Eyer, L.; Hruska, K. Antiviral Agents Targeting the Influenza Virus: A Review and Publication Analysis. Vet. Med. 2013, 58, 113–185. [Google Scholar] [CrossRef]
- Ortigoza, M.B.; Dibben, O.; Maamary, J.; Martinez-Gil, L.; Leyva-Grado, V.H.; Abreu, P.; Ayllon, J.; Palese, P.; Shaw, M.L. A Novel Small Molecule Inhibitor of Influenza A Viruses That Targets Polymerase Function and Indirectly Induces Interferon. PLoS Pathog. 2012, 8, e1002668. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, G.; Li, Y.; Jin, Y.; Dale, R.; Sun, L.-Q.; Wang, M. Inhibition of Highly Pathogenic Avian H5N1 Influenza Virus Replication by RNA Oligonucleotides Targeting NS1 Gene. Biochem. Biophys. Res. Commun. 2008, 365, 369–374. [Google Scholar] [CrossRef]
- Rajput, R.; Khanna, M.; Kumar, P.; Kumar, B.; Sharma, S.; Gupta, N.; Saxena, L. Small Interfering RNA Targeting the Nonstructural Gene 1 Transcript Inhibits Influenza A Virus Replication in Experimental Mice. Nucleic Acid Ther. 2012, 22, 414–422. [Google Scholar] [CrossRef]
- Wen, X.; Sun, J.; Wang, X.; Bao, H.; Zhao, Y.; Zeng, X.; Xu, X.; Ma, Y.; Gu, L.; Chen, H. Identification of a Novel Linear Epitope on the NS1 Protein of Avian Influenza Virus. BMC Microbiol. 2015, 15, 168. [Google Scholar] [CrossRef] [PubMed]
- He, J.-L.; Hsieh, M.-S.; Chiu, Y.-C.; Juang, R.-H.; Wang, C.-H. Preparation of Monoclonal Antibodies against Poor Immunogenic Avian Influenza Virus Proteins. J. Immunol. Methods 2013, 387, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Bhatia, S.; Malik, Y.P.S.; Dubey, S.C. Production and Characterization of Monoclonal Antibodies Against NS1 Protein of H5N1 Avian Influenza Virus. Hybridoma 2010, 29, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Lou, K.; Wang, W. New Small-Molecule Drug Design Strategies for Fi Ghting Resistant in Fl Uenza A. Acta Pharm. Sin. B 2015, 5, 419–430. [Google Scholar] [CrossRef]
- Basu, D.; Walkiewicz, M.P.; Frieman, M.; Baric, R.S.; Auble, D.T.; Engel, D.A. Novel Influenza Virus NS1 Antagonists Block Replication and Restore Innate Immune Function. J. Virol. 2009, 83, 1881–1891. [Google Scholar] [CrossRef]
- Beyleveld, G.; White, K.M.; Ayllon, J.; Shaw, M.L. New-Generation Screening Assays for the Detection of Anti-Influenza Compounds Targeting Viral and Host Functions. Antivir. Res. 2013, 100, 120–132. [Google Scholar] [CrossRef]
- Walkiewicz, M.P.; Basu, D.; Jablonski, J.J.; Geysen, H.M.; Engel, D.A. Novel Inhibitor of Influenza Non-Structural Protein 1 Blocks Multi-Cycle Replication in an RNase L-Dependent Manner. J. Gen. Virol. 2011, 92, 60–70. [Google Scholar] [CrossRef]
- Jablonski, J.J.; Basu, D.; Engel, D.A.; Geysen, H.M. Design, Synthesis, and Evaluation of Novel Small Molecule Inhibitors of the Influenza Virus Protein NS1. Bioorg. Med. Chem. 2012, 20, 487–497. [Google Scholar] [CrossRef][Green Version]
- Kleinpeter, A.B.; Jureka, A.S.; Falahat, S.M.; Green, T.J.; Petit, C.M. Structural Analyses Reveal the Mechanism of Inhibition of Influenza Virus NS1 by Two Antiviral Compounds. J. Biol. Chem. 2018, 293, 14659–14668. [Google Scholar] [CrossRef]
- Patnaik, S.; Basu, D.; Dehdashti, S.; Zheng, W.; Ferrer, M.; Southall, N.; Taylor, M.; Engel, D.A.; Marugan, J.J. Discovery of Small Molecule Influenza Virus NS1 Antagonist; U.S. National Library of Medicine: Bethesda, MD, USA, 2012.
- Kong, J.; Shen, J.; Huang, Y.; Ruan, R.; Xiang, B.; Zheng, X.; Cheng, K.; Wang, W. Development of a Yeast Two-Hybrid Screen for Selection of A/H1N1 Influenza NS1 Non-Structural Protein and Human CPSF30 Protein Interaction Inhibitors. Acta Pharm. Sin. 2010, 45, 388–394. [Google Scholar]
- Mata, M.A.; Satterly, N.; Versteeg, G.A.; Frantz, D.; Wei, S.; Williams, N.; Schmolke, M.; Peña-Llopis, S.; Brugarolas, J.; Forst, C.V.; et al. Chemical Inhibition of RNA Viruses Reveals REDD1 as a Host Defense Factor. Nat. Chem. Biol. 2011, 7, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.J.; Xia, S.; Ma, L.-C.; Robertus, J.; Krug, R.M.; Anslyn, E.V.; Montelione, G.T.; Ellington, A.D. Identification of Influenza Virus Inhibitors Targeting NS1A Utilizing Fluorescence Polarization–Based High-Throughput Assay. J. Biomol. Screen. 2012, 17, 448–459. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Cho, E.J.; Leavitt, J.; Ma, L.-C.; Montelione, G.T.; Anslyn, E.V.; Krug, R.M.; Ellington, A.; Robertus, J.D. Synthesis and Evaluation of Quinoxaline Derivatives as Potential Influenza NS1A Protein Inhibitors. Bioorg. Med. Chem. Lett. 2011, 21, 3007–3011. [Google Scholar] [CrossRef]
- Maroto, M.; Fernandez, Y.; Ortin, J.; Pelaez, F.; Cabello, M.A. Development of an HTS Assay for the Search of Anti-Influenza Agents Targeting the Interaction of Viral RNA with the NS1 Protein. J. Biomol. Screen. 2008, 13, 581–590. [Google Scholar] [CrossRef]
- Kuss-Duerkop, S.K.; Wang, J.; Mena, I.; White, K.; Metreveli, G.; Sakthivel, R.; Mata, M.A.; Muñoz-Moreno, R.; Chen, X.; Krammer, F.; et al. Influenza Virus Differentially Activates MTORC1 and MTORC2 Signaling to Maximize Late Stage Replication. PLoS Pathog. 2017, 13, e1006635. [Google Scholar] [CrossRef]
- Martínez-Gil, L.; Ayllon, J.; Ortigoza, M.B.; García-Sastre, A.; Shaw, M.L.; Palese, P. Identification of Small Molecules with Type I Interferon Inducing Properties by High-Throughput Screening. PLoS ONE 2012, 7, e49049. [Google Scholar] [CrossRef]
- Shimberg, G.D.; Michalek, J.L.; Oluyadi, A.A.; Rodrigues, A.V.; Zucconi, B.E.; Neu, H.M.; Ghosh, S.; Sureschandra, K.; Wilson, G.M.; Stemmler, T.L.; et al. Cleavage and Polyadenylation Specificity Factor 30: An RNA-Binding Zinc-Finger Protein with an Unexpected 2Fe–2S Cluster. Proc. Natl. Acad. Sci. USA 2016, 113, 4700–4705. [Google Scholar] [CrossRef]
- Ai, H.; Zhang, L.; Chang, A.K.; Wei, H.; Che, Y.; Liu, H. Virtual Screening of Potential Inhibitors from TCM for the CPSF30 Binding Site on the NS1A Protein of Influenza A Virus. J. Mol. Model. 2014, 20, 2142. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Ai, H.; Zheng, F.; Zhu, C.; Sun, T.; Zhang, L.; Liu, X.; Li, X.; Zhu, G.; Liu, H. Discovery of Novel Influenza Inhibitors Targeting the Interaction of DsRNA with the NS1 Protein by Structure-Based Virtual Screening. Int. J. Bioinform. Res. Appl. 2010, 6, 449. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Li, L.; Hsu, W.-L.; Pilcher, M.N.; Uversky, V.; Zhou, Y.; Dunker, A.K.; Meroueh, S.O. Exploring the Molecular Design of Protein Interaction Sites with Molecular Dynamics Simulations and Free Energy Calculations. Biochemistry 2009, 48, 399–414. [Google Scholar] [CrossRef] [PubMed]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA Methods to Estimate Ligand-Binding Affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Tieleman, D.P.; Berendsen, H.J.C. Molecular Dynamics Simulations of a Fully Hydrated Dipalmitoylphosphatidylcholine Bilayer with Different Macroscopic Boundary Conditions and Parameters. J. Chem. Phys. 1996, 105, 4871–4880. [Google Scholar] [CrossRef]
- Hutchinson, E.C.; Charles, P.D.; Hester, S.S.; Thomas, B.; Trudgian, D.; Martínez-Alonso, M.; Fodor, E. Conserved and Host-Specific Features of Influenza Virion Architecture. Nat. Commun. 2014, 5, 4816. [Google Scholar] [CrossRef]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable Molecular Dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef]
- Iwasaki, A.; Pillai, P.S. Innate Immunity to Influenza Virus Infection. Nat. Rev. Immunol. 2014, 14, 315–328. [Google Scholar] [CrossRef]
- Barnwal, B.; Mok, C.-K.; Wu, J.; Diwakar, M.K.; Gupta, G.; Zeng, Q.; Chow, V.T.K.; Song, J.; Yuan, Y.A.; Tan, Y.-J. A Monoclonal Antibody Binds to Threonine 49 in the Non-Structural 1 Protein of Influenza A Virus and Interferes with Its Ability to Modulate Viral Replication. Antivir. Res. 2015, 116, 55–61. [Google Scholar] [CrossRef]
- Zhang, H. Tissue and Host Tropism of Influenza Viruses: Importance of Quantitative Analysis. Sci. China Ser. C Life Sci. 2009, 52, 1101–1110. [Google Scholar] [CrossRef]
- Eng, C.L.; Tong, J.; Tan, T. Predicting Host Tropism of Influenza A Virus Proteins Using Random Forest. BMC Med. Genom. 2014, 7, S1. [Google Scholar] [CrossRef]
- Pavlin, B.I.; Schloegel, L.M.; Daszak, P. Risk of Importing Zoonotic Diseases through Wildlife Trade, United States. Emerg. Infect. Dis. 2009, 15, 1721–1726. [Google Scholar] [CrossRef] [PubMed]
- Squires, R.B.; Noronha, J.; Hunt, V.; García-Sastre, A.; Macken, C.; Baumgarth, N.; Suarez, D.; Pickett, B.E.; Zhang, Y.; Larsen, C.N.; et al. Influenza Research Database: An Integrated Bioinformatics Resource for Influenza Research and Surveillance. Influenza Other Respir. Viruses 2012, 6, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Rajao, D.S.; Anderson, T.K.; Gauger, P.C.; Vincent, A.L. Evolution and Ecology of Influenza A Viruses. In Current Topics in Microbiology and Immunology Influenza Pathogenesis and Control-Volume I; Springer: Cham, Switzerland, 2014; Volume 385, pp. 307–321. [Google Scholar]
- Kargarfard, F.; Sami, A.; Mohammadi-Dehcheshmeh, M.; Ebrahimie, E. Novel Approach for Identification of Influenza Virus Host Range and Zoonotic Transmissible Sequences by Determination of Host-Related Associative Positions in Viral Genome Segments. BMC Genom. 2016, 17, 925. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.-L. Current and Novel Antiviral Strategies for Influenza Infection. Curr. Opin. Virol. 2016, 18, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Eng, C.L.P.; Tong, J.C.; Tan, T.W. Distinct Host Tropism Protein Signatures to Identify Possible Zoonotic Influenza A Viruses. PLoS ONE 2016, 11, e0150173. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RNA Segment | Protein(s) Coded | Function [20,31,35] | Structural Data [20,31,35,36] |
|---|---|---|---|
| 1 | PB2 759 aas | Located in the nucleus of infected cells; Signals the viral polymerase passage to the host’s nucleus; Enhances the formation of the cap structures necessary for viral messenger RNA (mRNA) transcription; Located in the mitochondria of infected cells [37]; Inhibits Interferon-β; Helps determine host range. | The three proteins, PB2 (polymerase basic protein 2), PB1 (polymerase basic protein 1) and PA (polymerase acidic protein), form the viral RNA polymerase, responsible for viral RNA transcription and replication. |
| 2 | PB1 757 aas | Responsible for the elongation of the primed nascent viral mRNA; Located in the nucleus of infected cells; Enhances the association of the 3 subunits of the RNA polymerase complex. | |
| 3 | PA 716 aas | Functions still unknown, but evidence points to helicase-like functions; Important for viral transcription; Assembly of the polymerase complex. | |
| 4 | HA 550 aas | Attaches the virions to the sialic acid (SA) moieties of the host’s receptors; Around 30% variation between subtypes. | Hemagglutinin (HA) is a homotrimeric integral cylinder-like membrane glycoprotein on the virus surface; 4 antigenic sites with direct impact on virulence and pathogenicity of the virus. |
| 5 | NP 498 aas | Binds non-specifically to single-stranded RNA (ssRNA); Encapsidates viral RNA; Helps recruiting RNA polymerase for synthesis of viral positive-sense RNA (cRNA); Related to host range. | Nucleoprotein (NP) is a 56 kDa basic protein; RNA-binding protein; Structural unit of RNPs; Forms oligomers stabilized by vRNA. |
| 6 | NA 470 aas | Unnecessary for virus replication; Required for budding of newly formed viral particles from surface of infected cells; Facilitates virus movement to the target cell by cleavage of sialic acids from respiratory tract mucins; Helps the release of virions from infected cells. | Neuraminidase (NA) is a homotetrameric integral membrane glycoprotein with 4 structural domains; Antigenic sites help circumvent the immune responses aiding on the virulence and pathogenicity of the virus. |
| 7 | M1 252 aas | Membrane-binding and RNA-binding protein; Forms a coat inside the viral envelope; Determines virion’s shape; Interacts with vRNP and other cytoplasmic domains of integral membrane proteins; Increases vRNPs export and decreases import; Helps assembly and budding of virions. | Matrix protein (M1) formed by a globular N-terminal domain and a flexible C-terminal tail; Oligomerization state and binding to lipid bilayer are highly dependent on pH. |
| M2 97 aas | Vital for viral replication; Forms proton channel in virus envelope; Lowers the pH inside the viral particle to promote uncoating of RNPs; Modulates Golgi’s pH; Helps to stabilize HA’s native conformation during virus assembly. | Matrix-2 protein (M2) is a 97-residue single-pass membrane protein; Three segments: N-terminal outward segment, transmembrane (TM) helix, and C-terminal inward segment; TM helices from 4 subunits pack to form proton-channel; Highly conserved His37 and Trp41 residues. | |
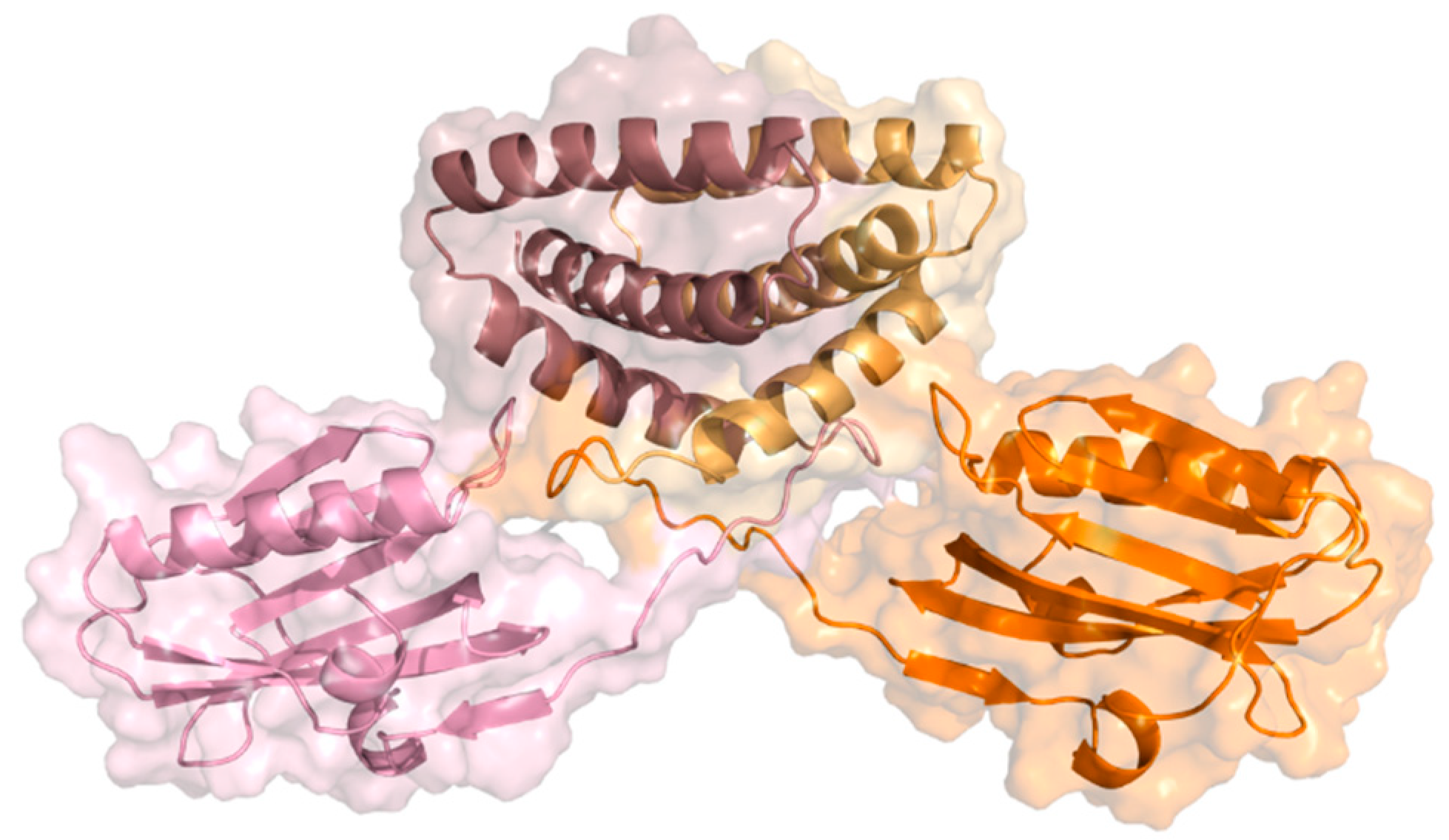
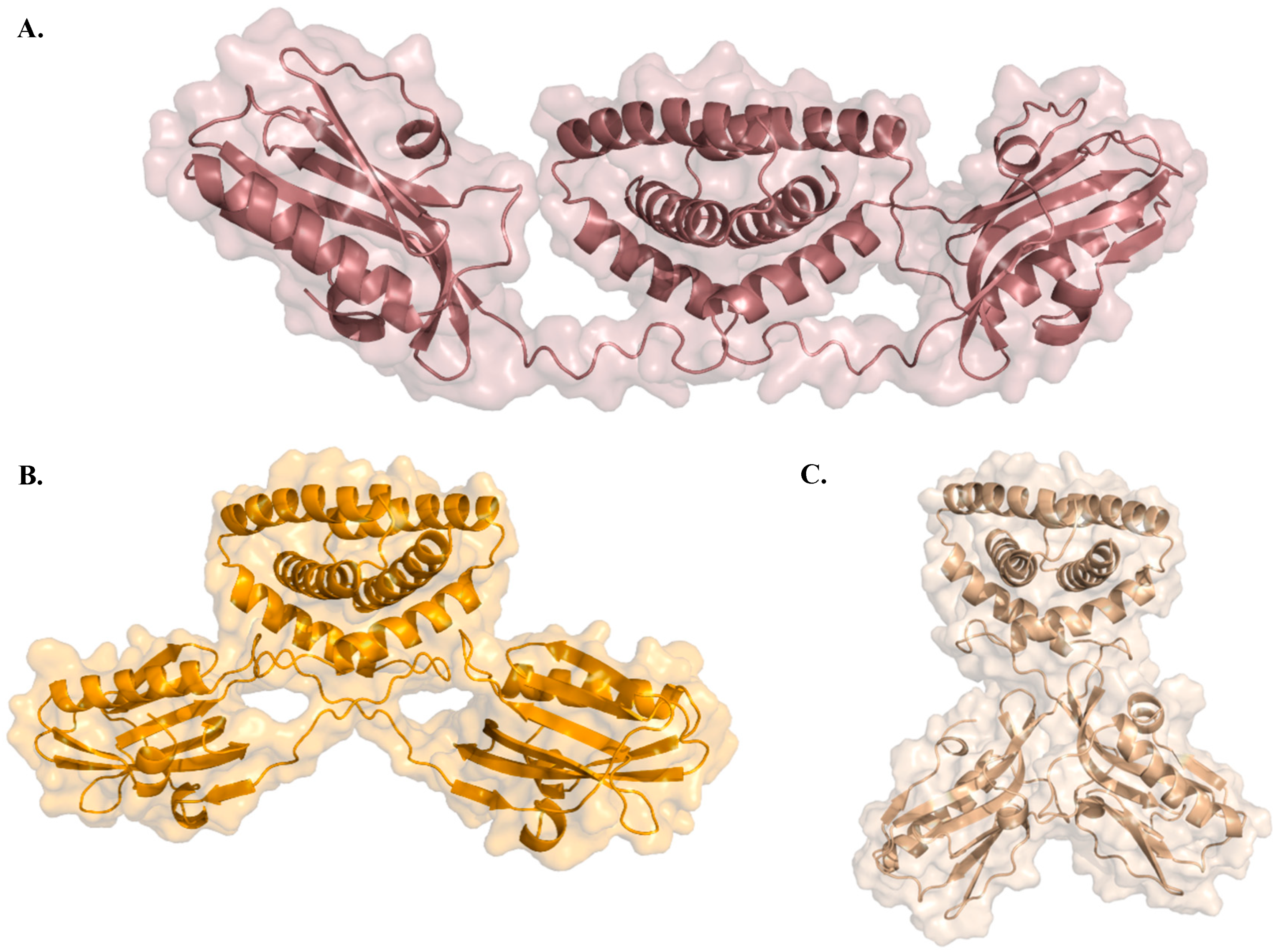
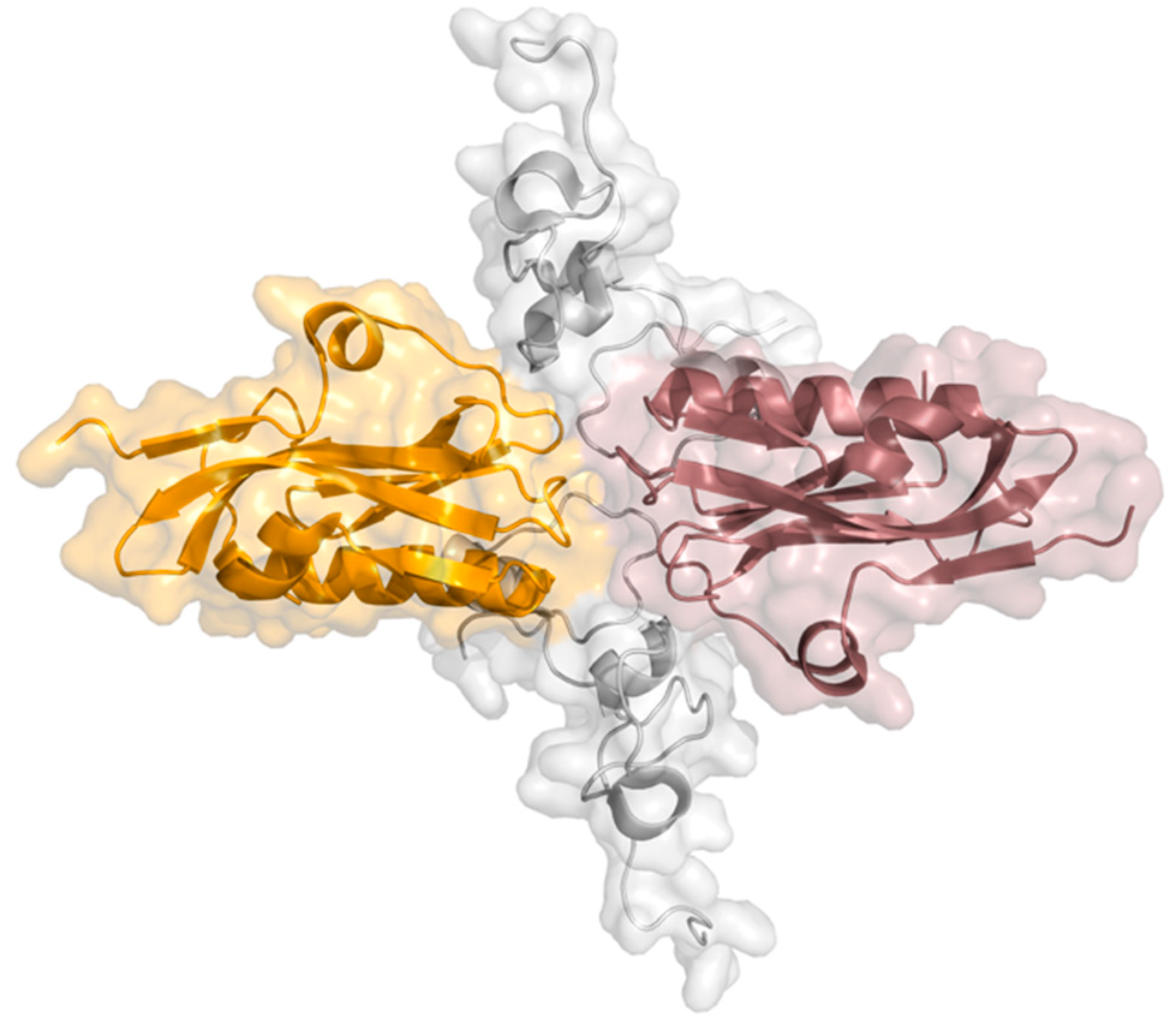
| 8 | NS1 230 aas | NS1 acts as a promoter of viral replication and an inhibitor of the host’s immune response; Present in the cytoplasm and nucleus of the host cell. | Non-structural protein 1 (NS1) has two structural domains—RNA-binding domain (RBD) and the effector domain (ED)—connected by a short linker (LR), and a disordered C-terminal tail (CTT). |
| NEP/NS2 121 aas | Promotes viral RNA replication; Regulates vRNP’s export from the nucleus to the cytoplasm; RNA nuclear export; Interacts with the viral matrix M1 protein. | Nuclear Export Protein (NEP) has a protease-sensitive N-terminal domain (residues 1–53) and a protease-resistant C-terminal domain (residues 54–121) mostly formed by a helical hairpin. |
| Ligand | Activity | Method | Mode of Action | Ref |
|---|---|---|---|---|
| NSC128164 NSC109834 NSC95676 | Blocks NS1’s action on IFN-related pathways | Yeast-based assay | Attenuates NS1 expression | [136] |
| NSC125044 | Yeast-based assay | Induction of IFN-β and decrease of viral proteins expression | ||
| JJ3297 | NS1 antagonist | Yeast-based assay | Restores IFN antiviral state and inhibits virus replication | [138] |
| ML303 | NS1 antagonist | Yeast-based assay | Restores IFN signaling pathway | [141] |
| “Shuanghualian” | NS1 antagonist | Yeast-based assay | Inhibits CPSF30 function by binding to NS1A binding site | [142] |
| Naphthalimide family of compounds | NS1 antagonist | Fluorescence polarization-based high-throughput assay | Up-regulation of REDD1 and mTOR1 inhibition | [143] |
| A9 (JJ3297) and A22 | NS1 antagonist | Molecular docking, NMR, X-ray crystallography | Inhibit CPSF30 function by binding to NS1A binding site | [140] |
| 30256 and 31674 (see paper) | NS1 inhibitor | Molecular Docking, Molecular Dynamics, MM-PBSA | Inhibit CPSF30 function by binding to NS1A binding site | [151,157,158] |
| (no name, see paper) | Blocks NS1 | Molecular Docking | Inhibits dsRNA binding | [151,152] |
| Epigallocatechin gallate (EGCG) | Proposed binding to NS1’s Arg38 | Fluorescence polarization-based high-throughput assay | Inhibits dsRNA binding | [144] |
| Quinoxaline family of compounds | NS1 inhibitor | Fluorescence polarization-based high-throughput assay | Inhibit dsRNA binding | [145] |
| (no name, see paper for three compounds) | Block NS1 | Radiolabeled RNA | Inhibit NS1 binding to RNA | [146] |
| C3 | Counteracts IFN blockage by NS1 | Caspase 3-based high-throughput assay | IFN-induction | [148] |
| mAb | Binds selectively to NS1 of avian influenza virus | Antigen screening using recombinant peptides | Recognition of NS1’s presence | [132] |
| mAb | Binds to Thr49 region of NS1 of avian influenza virus | Enzyme-Linked Immunosorbent Assay (ELISA), surface plasmon resonance | Interferes with the virus ability to replicate | [159] |
| mAb D9 | Binds to linear epitope in NS1 | Western blot assay | Recognition of NS1’s presence | [131] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosário-Ferreira, N.; Preto, A.J.; Melo, R.; Moreira, I.S.; Brito, R.M.M. The Central Role of Non-Structural Protein 1 (NS1) in Influenza Biology and Infection. Int. J. Mol. Sci. 2020, 21, 1511. https://doi.org/10.3390/ijms21041511
Rosário-Ferreira N, Preto AJ, Melo R, Moreira IS, Brito RMM. The Central Role of Non-Structural Protein 1 (NS1) in Influenza Biology and Infection. International Journal of Molecular Sciences. 2020; 21(4):1511. https://doi.org/10.3390/ijms21041511
Chicago/Turabian StyleRosário-Ferreira, Nícia, António J. Preto, Rita Melo, Irina S. Moreira, and Rui M. M. Brito. 2020. "The Central Role of Non-Structural Protein 1 (NS1) in Influenza Biology and Infection" International Journal of Molecular Sciences 21, no. 4: 1511. https://doi.org/10.3390/ijms21041511
APA StyleRosário-Ferreira, N., Preto, A. J., Melo, R., Moreira, I. S., & Brito, R. M. M. (2020). The Central Role of Non-Structural Protein 1 (NS1) in Influenza Biology and Infection. International Journal of Molecular Sciences, 21(4), 1511. https://doi.org/10.3390/ijms21041511