EbARC1, an E3 Ubiquitin Ligase Gene in Erigeron breviscapus, Confers Self-Incompatibility in Transgenic Arabidopsis thaliana


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
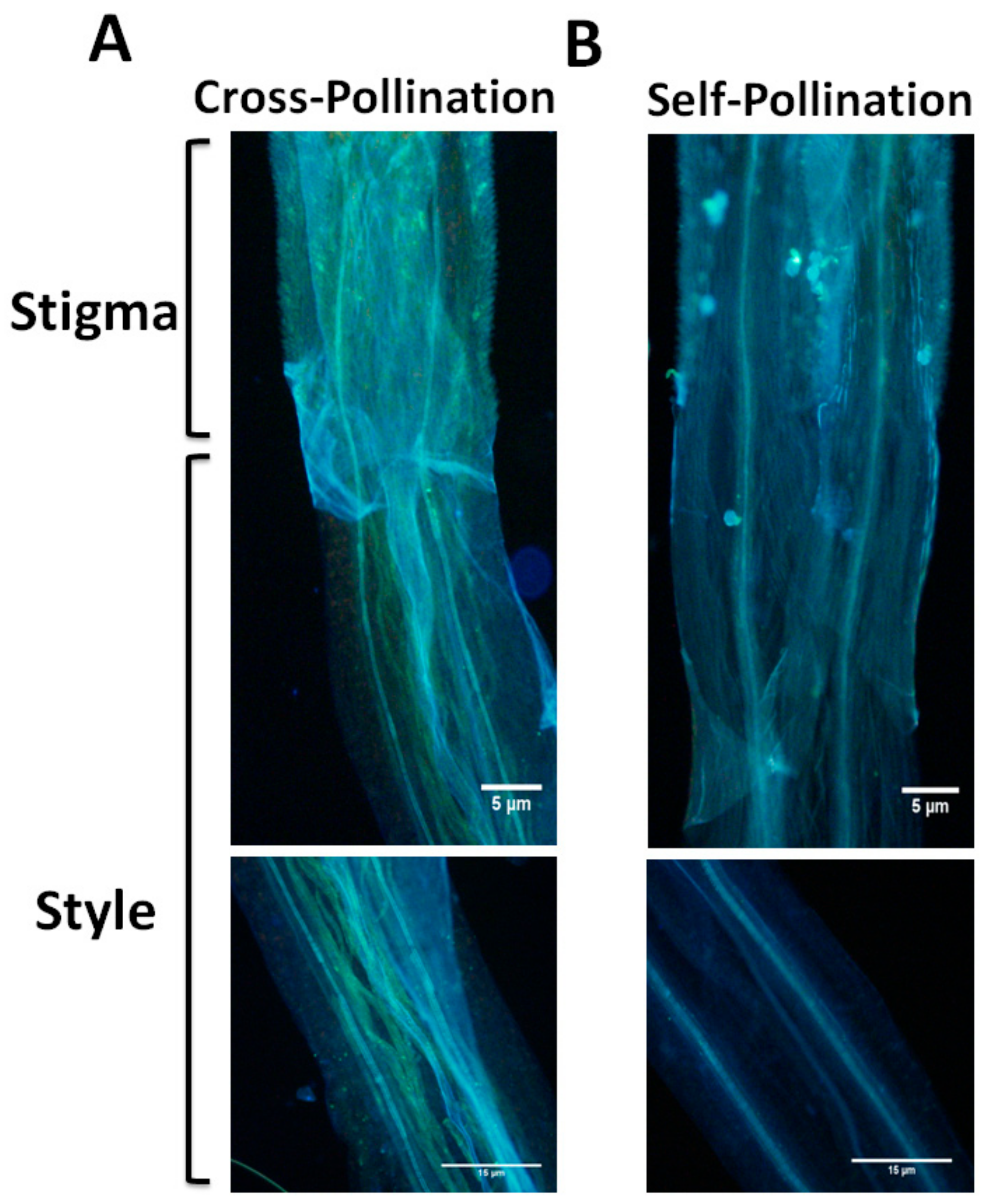
2.1. E. breviscapus has SSI Characteristics
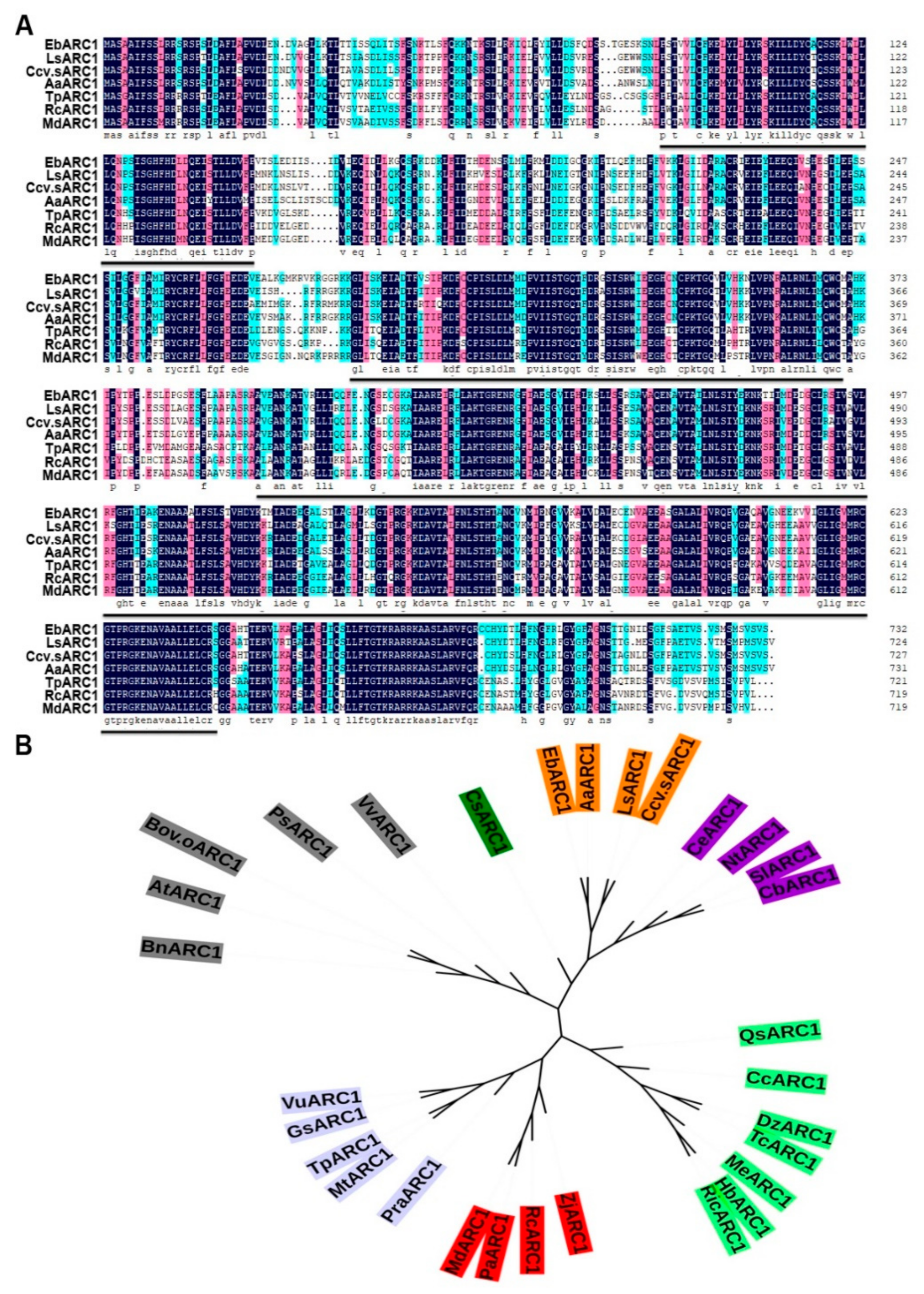
2.2. Molecular Characteristics and Expression Analysis of EbARC1
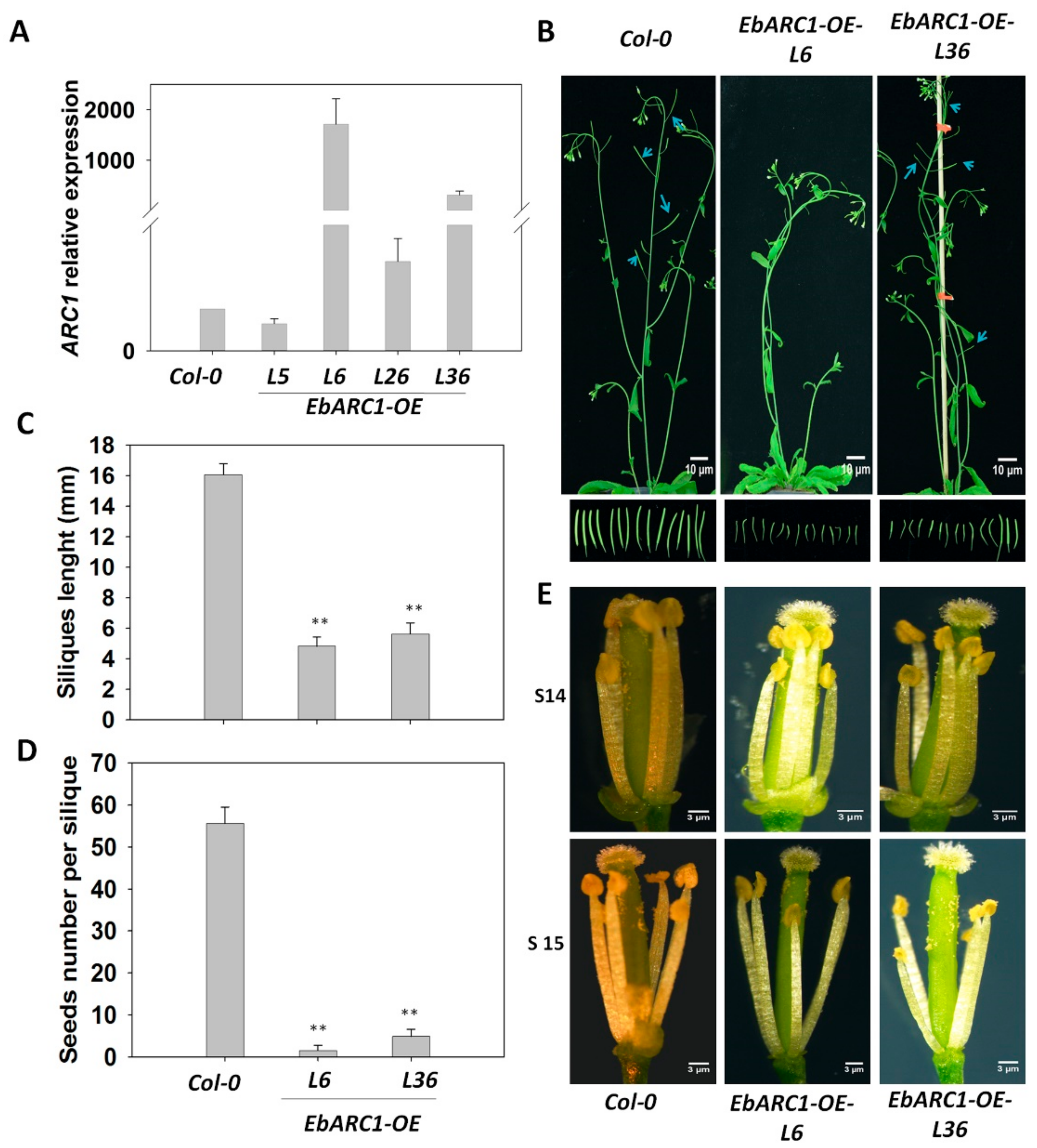
2.3. Overexpression of EbARC1 Causes Self-Sterility in A. thaliana
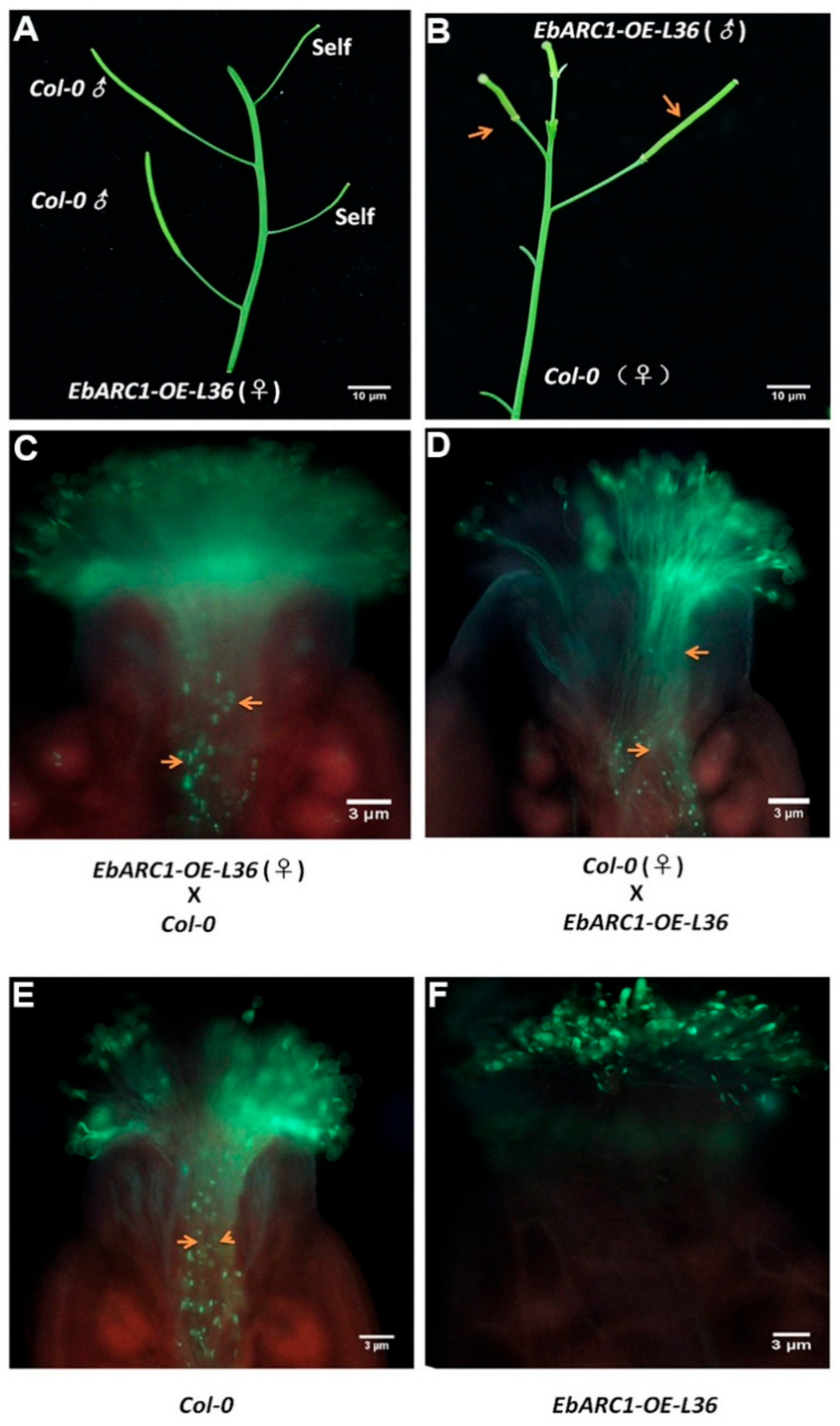
2.4. Transforming SC into SI through Overexpressing EbARC1 in A. thaliana
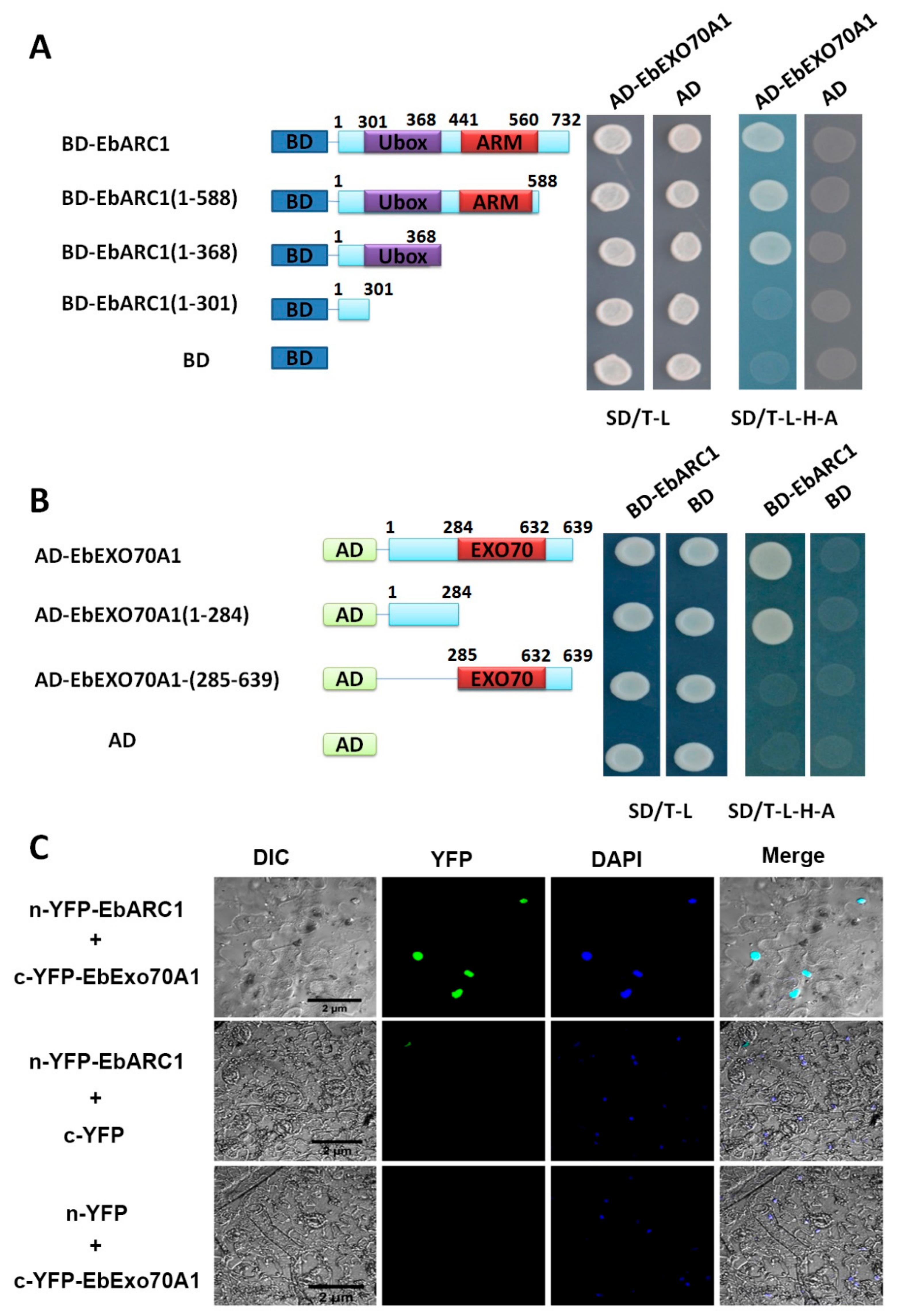
2.5. EbARC1 Protein Interacts with EbExo70A1
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Bioinformation Analysis
4.3. Vector Construction and Plasmid Transformation
4.4. Subcellular Localization Analysis
4.5. Pollination Assays
4.6. Aniline Blue Staining Assay
4.7. RNA Extraction and qRT-PCR
4.8. Yeast Two-Hybrid Assay
4.9. BiFC Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Barrett, S.C. Mating strategies in flowering plants: The outcrossing-selfing paradigm and beyond. Phil. Trans. R. Soc. Lond. B. 2003, 358, 991–1004. [Google Scholar] [CrossRef]
- Charlesworth, D. Evolution of plant breeding systems. Curr. Biol. 2006, 16, 726–735. [Google Scholar] [CrossRef]
- Webb, C.J.; Lloyd, D.G. The avoidance of interference between the presentation of pollen and stigmas in angiosperms ii. herkogamy. New Zeal J. Bot. 1986, 24, 163–178. [Google Scholar] [CrossRef]
- Iwano, M.; Takayama, S. Self/non-self discrimination in angiosperm self-incompatibility. Curr. Opin. Plant Biol. 2012, 15, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Indriolo, E.; Safavian, D.; Goring, D.R. The ARC1 E3 ligase promotes two different self-pollen avoidance traits in Arabidopsis. Plant Cell 2014, 26, 1525–1543. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Widmer, A. Herkogamy and its effects on mating patterns in Arabidopsis thaliana. PLoS ONE 2013, 8, e57902. [Google Scholar] [CrossRef] [PubMed]
- Hiscock, S.J. Self-incompatibility in Senecio squalidus L. (Asteraceae). Ann. Botany. 2000, 85, 181–190. [Google Scholar] [CrossRef]
- Takasaki, T.; Hatakeyama, K.; Suzuki, G.; Watanabe, M.; Isogai, A.; Hinata, K. The S receptor kinase determines self-incompatibility in Brassica stigma. Nature 2000, 403, 913–916. [Google Scholar] [CrossRef]
- Takayama, S.; Shimosato, H.; Shiba, H.; Funato, M.; Che, F.S.; Watanabe, M.; Iwano, M.; Isogai, A. Direct ligand–receptor complex interaction controls Brassica self-incompatibility. Nature 2001, 413, 534–538. [Google Scholar] [CrossRef]
- Iwano, M.; Shiba, H.; Funato, M.; Shimosato, H.; Takayama, S.; Isogai, A. Immunohistochemical studies on translocation of pollen S-haplotype determinant in self-incompatibility of Brassica rapa. Plant Cell Physiol. 2003, 44, 428–436. [Google Scholar] [CrossRef]
- Gu, T.; Mazzurco, M.; Sulaman, W.; Matias, D.D.; Goring, D.R. Binding of an ARM repeat containing protein to the kinase domain of the S-locus receptor kinase. Proc. Natl. Acad. Sci. USA 1998, 95, 382–387. [Google Scholar] [CrossRef]
- Mazzurco, M.; Sulaman, W.; Elina, H.; Cock, J.M.; Goring, D.R. Further analysis of the interactions between the Brassica S receptor kinase and three interacting proteins (ARC1, THL1 and THL2) in the yeast two-hybrid system. Plant Mol. Biol. 2001, 45, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Anderson, E.M.; Mullen, R.T.; Goring, D.R. ARC1 is an E3 ubiquitin ligase and promotes the ubiquitination of proteins during the rejection of self-incompatible Brassica pollen. Plant Cell 2003, 15, 885–898. [Google Scholar] [CrossRef] [PubMed]
- Indriolo, E.; Goring, D.R. A conserved role for the ARC1 E3 ligase in Brassicaceae self-incompatibility. Front. Plant Sci. 2014, 5, 181. [Google Scholar] [CrossRef] [PubMed]
- Indriolo, E.; Goring, D.R. Yeast two-hybrid interactions between Arabidopsis lyrata S receptor kinase and the ARC1 E3 ligase. Plant Signal. Behav. 2016, 11. [Google Scholar] [CrossRef][Green Version]
- Samuel, M.A.; Chong, Y.T.; Haasen, K.E.; Aldea-Brydges, M.G.; Stone, S.L.; Goring, D.R. Cellular pathways regulating responses to compatible and self-incompatible pollen in Brassica and Arabidopsis stigmas intersect at Exo70A1, a putative component of the exocyst complex. Plant Cell 2009, 21, 2655–2671. [Google Scholar] [CrossRef]
- Sankaranarayanan, S.; Jamshed, M.; Samuel, M.A. Degradation of glyoxalase I in Brassica napus stigma leads to self-incompatibility response. Nat. Plants 2015, 1, 15185. [Google Scholar] [CrossRef]
- Scandola, S.; Samuel, M.A. A flower-specific phospholipase D Is a stigmatic compatibility factor targeted by the self-incompatibility response in Brassica napus. Curr. Biol. 2019, 29, 506–512. [Google Scholar] [CrossRef]
- Stone, S.L.; Arnoldo, M.; Goring, D.R. A breakdown of Brassica self-incompatibility in ARC1 antisense transgenic plants. Science 1999, 286, 1729–1731. [Google Scholar] [CrossRef]
- Indriolo, E.; Tharmapalan, P.; Wright, S.I.; Goring, D.R. The ARC1 E3 ligase gene is frequently deleted in self-compatible Brassicaceae species and has a conserved role in Arabidopsis lyrata self-pollen rejection. Plant Cell 2012, 24, 4607–4620. [Google Scholar] [CrossRef]
- Wikström, N.; Savolainen, V.; Chase, M.W. Evolution of the angiosperms: Calibrating the family tree. P Roy. Soc. B-Biol. Sci. 2001, 268, 2211–2220. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.M.; Good-Avila, S.V. Macrophylogenetic analyses of the gain and loss of self-incompatibility in the asteraceae. New Phytol. 2007, 173, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Hiscock, S.J.; Tabah, D.A. The different mechanisms of sporophytic self-incompatibility in flowering plants. Philos. Trans. R. Soc. 2003, 358, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Mcinnis, S.M.; Costa, L.M.; Gutiérrez-Marcos, J.F.; Henderson, C.A.; Hiscock, S.J. Isolation and characterization of a polymorphic stigma-specific class III peroxidase gene from senecio squalidus L. (Asteraceae). Plant Mol. Biol. 2005, 57, 659–677. [Google Scholar] [CrossRef] [PubMed]
- McInnis, S.; Emery, D.; Porter, R.; Desikan, R.; Hancock, J.T.; Hiscock, S.J. The role of stigma peroxidases in flowering plants: Insights from further characterization of a stigma-specific peroxidase (SSP) from Senecio squalidus (Asteraceae). J. Exp. Bot. 2006, 57, 1835–1846. [Google Scholar] [CrossRef]
- Tabah, D.A.; Mcinnis, S.M.; Hiscock, S.J. Members of the S-receptor kinase multigene family in Senecio squalidus L. (Asteraceae), a species with sporophytic self-incompatibility. Sex. Plant Repord. 2004, 17, 131–140. [Google Scholar] [CrossRef]
- Lin, R.; Chen, Y.L.; Shi, Z. Asteraceae. In Flora of China; China Science and Technology Press: Beijing, China, 1985; Volume 74, pp. 297–308. [Google Scholar]
- Liu, X.N.; Cheng, J.; Zhang, G.H.; Ding, W.T.; Duan, L.J.; Yang, J.; Kui, L.; Cheng, X.Z.; Ruan, J.X.; Fan, W.; et al. Engineering yeast for the production of breviscapine by genomic analysis and synthetic biology approaches. Nat. Commun. 2018, 9, 448. [Google Scholar] [CrossRef]
- Zhang, W.; Wei, X.; Meng, H.L.; Ma, C.H.; Jiang, N.H.; Zhang, G.H.; Yang, S.C. Transcriptomic comparison of the self-pollinated and cross-pollinated flowers of Erigeron breviscapus to analyze candidate self-incompatibility-associated genes. BMC Plant Biol. 2015, 15, 248. [Google Scholar] [CrossRef]
- Allen, A.M.; Thorogood, C.J.; Hegarty, M.J.; Lexer, C.; Hiscock, S.J. Pollen-pistil interactions and self-incompatibility in the Asteraceae: New insights from studies of senecio squalidus (oxford ragwort). Ann. Botany. 2011, 108, 687–698. [Google Scholar] [CrossRef]
- Koseva, B.; Crawford, D.J.; Brown, K.E.; Mort, M.E.; Kelly, J.K. The genetic breakdown of sporophytic self-incompatibility in tolpis coronopifolia (Asteraceae). New Phytologist. 2017, 216, 1256–1267. [Google Scholar] [CrossRef]
- Gao, J.L.; Chen, G.; He, H.Q.; Liu, C.; Xiong, X.J.; Li, J.; Wang, J. Therapeutic Effects of Breviscapine in Cardiovascular Diseases: A Review. Front. Pharmacol. 2017, 8, 289. [Google Scholar] [CrossRef]
- Sherman-Broyles, S.; Nathan, B.; Agnes, F.; Pei, L.; Julia, V.; Nasrallah, M.E.; Nasrallah, J.B. S Locus Genes and the Evolution of Self-Fertility in Arabidopsis thaliana. Plant Cell 2007, 19, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Tsuchimatsu, T.; Suwabe, K.; Shimizu-Inatsugi, R.; Isokawa, S.; Pavlidis, P.; Stadler, T.; Suzuki, G.; Takayama, S.; Watanabe, M.; Kentaro, K.; et al. Evolution of self-compatibility in Arabidopsis by a mutation in the male specificity gene. Nature 2010, 464, 1342–1346. [Google Scholar] [CrossRef] [PubMed]
- Nasrallah, M.E.; Liu, P.; Sherman-Broyles, S.; Boggs, N.A.; Nasrallah, J.B. Natural variation in expression of self-incompatibility in Arabidopsis thaliana: Implications for the evolution of selfing. Proc. Natl. Acad Sci. USA 2004, 101, 16070–16074. [Google Scholar] [CrossRef] [PubMed]
- Boggs, N.A.; Dwyer, K.G.; Shah, P.; McCulloch, A.A.; Bechsgaard, J.; Schierup, M.H.; Nasrallah, M.E.; Nasrallah, J.B. Expression of distinct self-incompatibility specificities in Arabidopsis thaliana. Genetics 2009, 182, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhou, G.; Goring, D.R.; Liang, X.; Macgregor, S.; Dai, C.; Wen, J.; Yi, B.; Shen, J.; Tu, J.; et al. Generation of transgenic self-incompatible Arabidopsis thaliana shows a genus-specific preference for self-incompatibility genes. Plants 2019, 8, 570. [Google Scholar] [CrossRef] [PubMed]
- Franzke, A.; Lysak, M.A.; Al-Shehbaz, I.A.; Koch, M.A.; Mummenhoff, K. Cabbage family affairs: The evolutionary history of Brassicaceae. Trends Plant Sci. 2011, 16, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.M.; Gao, Q.G.; Zeng, J.; Liu, X.H.; Pu, Q.M.; Liu, G.X.; Zhang, H.C.; Yang, X.H.; Zhu, L.Q. N-terminal domains of ARC1 are essential for interaction with the N-terminal region of Exo70A1 in transducing self-incompatibility of Brassica oleracea. Acta Bioch. Bioph. Sin. 2016, 48, 777–787. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, H.C.; Lian, X.P.; Converse, R.; Zhu, L.Q. Identification of Interacting Motifs Between ARM repeatadillo Repeat Containing 1 (ARC1) and Exocyst 70 A1 (Exo70A1) Proteins in Brassica oleracea. Protein J. 2016, 35, 34–43. [Google Scholar] [CrossRef]
- Zárský, V.; Kulich, I.; Fendrych, M.; Peèenková, T. Exocyst complexes multiple functions in plant cells secretory pathways. Curr. Opin. Plant. Biol. 2013, 16, 726–733. [Google Scholar] [CrossRef]
- Li, S.P.; Chen, M.; Yu, D.L.; Ren, S.C.; Sun, S.F.; Liu, L.D.; Ketelaar, T.; Emons, A.M.; Liu, C.M. EXO70A1-Mediated Vesicle Trafficking Is Critical for Tracheary Element Development in Arabidopsis. Plant Cell 2013, 25, 1774–1786. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.Q.; Zhang, Y.T.; Tu, J.X.; Shen, J.X.; Yi, B.; Fu, T.D.; Dai, C.; Ma, C.Z. The Brassica napus GATA transcription factor BnA5.ZML1 is a stigma compatibility factor. J. Integr. Plant Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Yang, Y.; Li, B.; Liu, Z.Q.; Fawad, K.; Zhang, T.; Zhou, G.L.; Tu, J.X.; Shen, J.X.; Yi, B.; et al. Functional analysis of M-locus protein kinase revealed a novel regulatory mechanism of Self-Incompatibility in Brassica napus L. Int. J. Mol. Sci. 2019, 20, 3303. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, M.; Fan, W.; Hao, B.; Zhang, W.; Yan, M.; Zhao, Y.; Liang, Y.; Liu, G.; Lu, Y.; Zhang, G.; et al. EbARC1, an E3 Ubiquitin Ligase Gene in Erigeron breviscapus, Confers Self-Incompatibility in Transgenic Arabidopsis thaliana. Int. J. Mol. Sci. 2020, 21, 1458. https://doi.org/10.3390/ijms21041458
Chen M, Fan W, Hao B, Zhang W, Yan M, Zhao Y, Liang Y, Liu G, Lu Y, Zhang G, et al. EbARC1, an E3 Ubiquitin Ligase Gene in Erigeron breviscapus, Confers Self-Incompatibility in Transgenic Arabidopsis thaliana. International Journal of Molecular Sciences. 2020; 21(4):1458. https://doi.org/10.3390/ijms21041458
Chicago/Turabian StyleChen, Mo, Wei Fan, Bing Hao, Wei Zhang, Mi Yan, Yan Zhao, Yanli Liang, Guanze Liu, Yingchun Lu, Guanghui Zhang, and et al. 2020. "EbARC1, an E3 Ubiquitin Ligase Gene in Erigeron breviscapus, Confers Self-Incompatibility in Transgenic Arabidopsis thaliana" International Journal of Molecular Sciences 21, no. 4: 1458. https://doi.org/10.3390/ijms21041458
APA StyleChen, M., Fan, W., Hao, B., Zhang, W., Yan, M., Zhao, Y., Liang, Y., Liu, G., Lu, Y., Zhang, G., Zhao, Z., Hu, Y., & Yang, S. (2020). EbARC1, an E3 Ubiquitin Ligase Gene in Erigeron breviscapus, Confers Self-Incompatibility in Transgenic Arabidopsis thaliana. International Journal of Molecular Sciences, 21(4), 1458. https://doi.org/10.3390/ijms21041458