TUBB Variants Underlying Different Phenotypes Result in Altered Vesicle Trafficking and Microtubule Dynamics

 , ,
, ,  , ,
, ,  , and
, and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
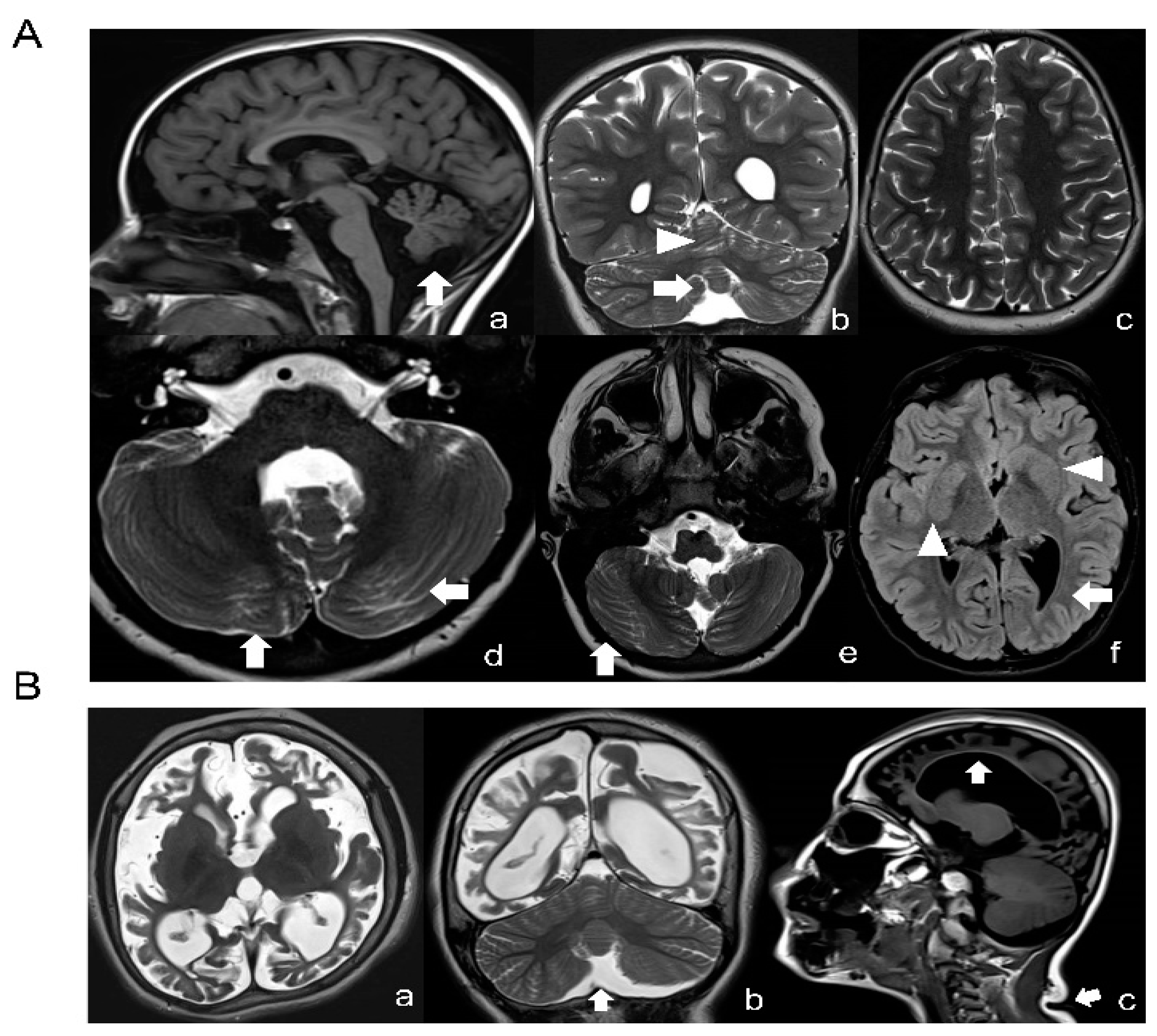
2.1. TUBB Gene Mutation Analysis
2.2. N52S and M73T Substitutions Affect Microtubule Dynamics In Vitro
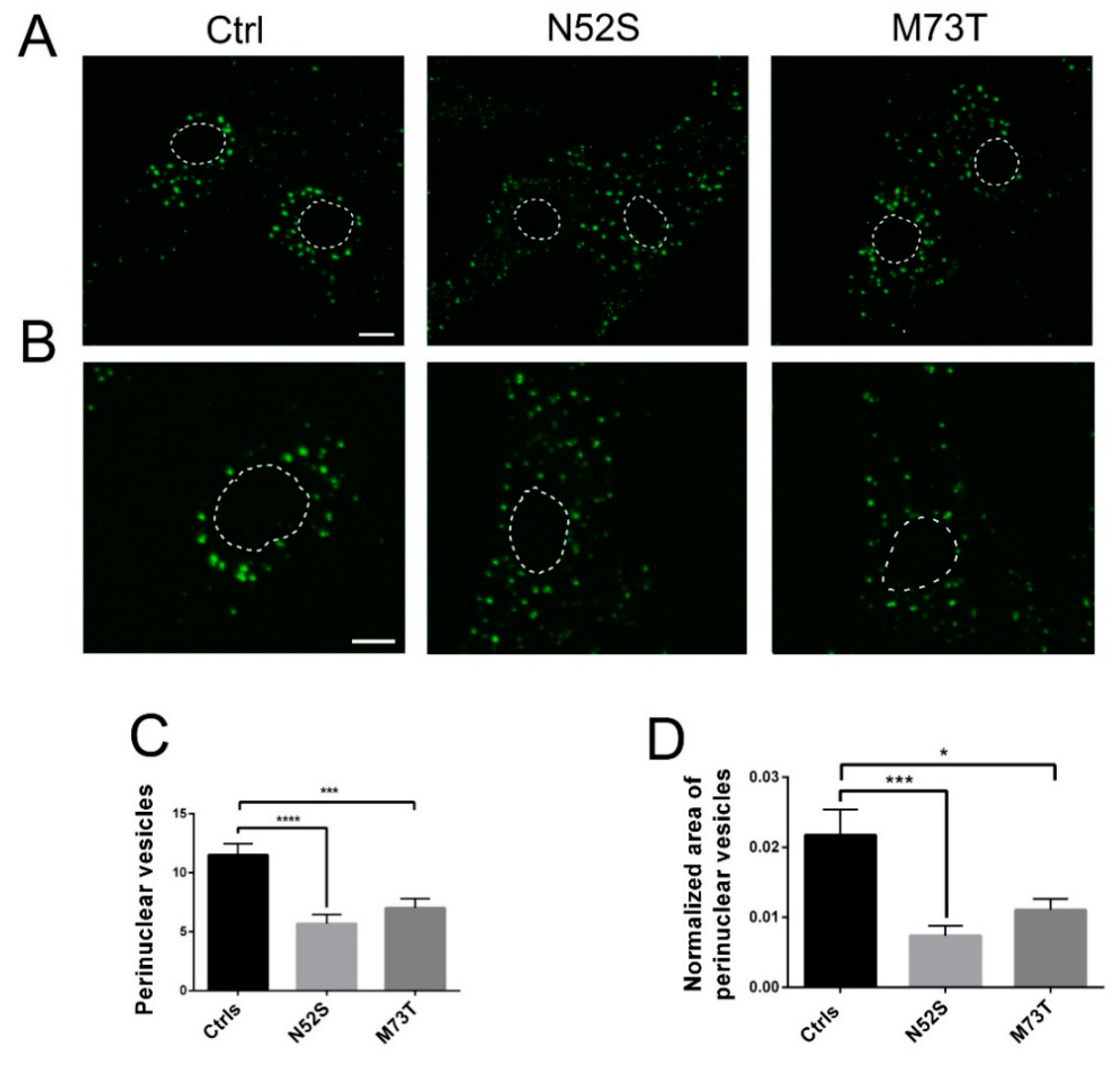
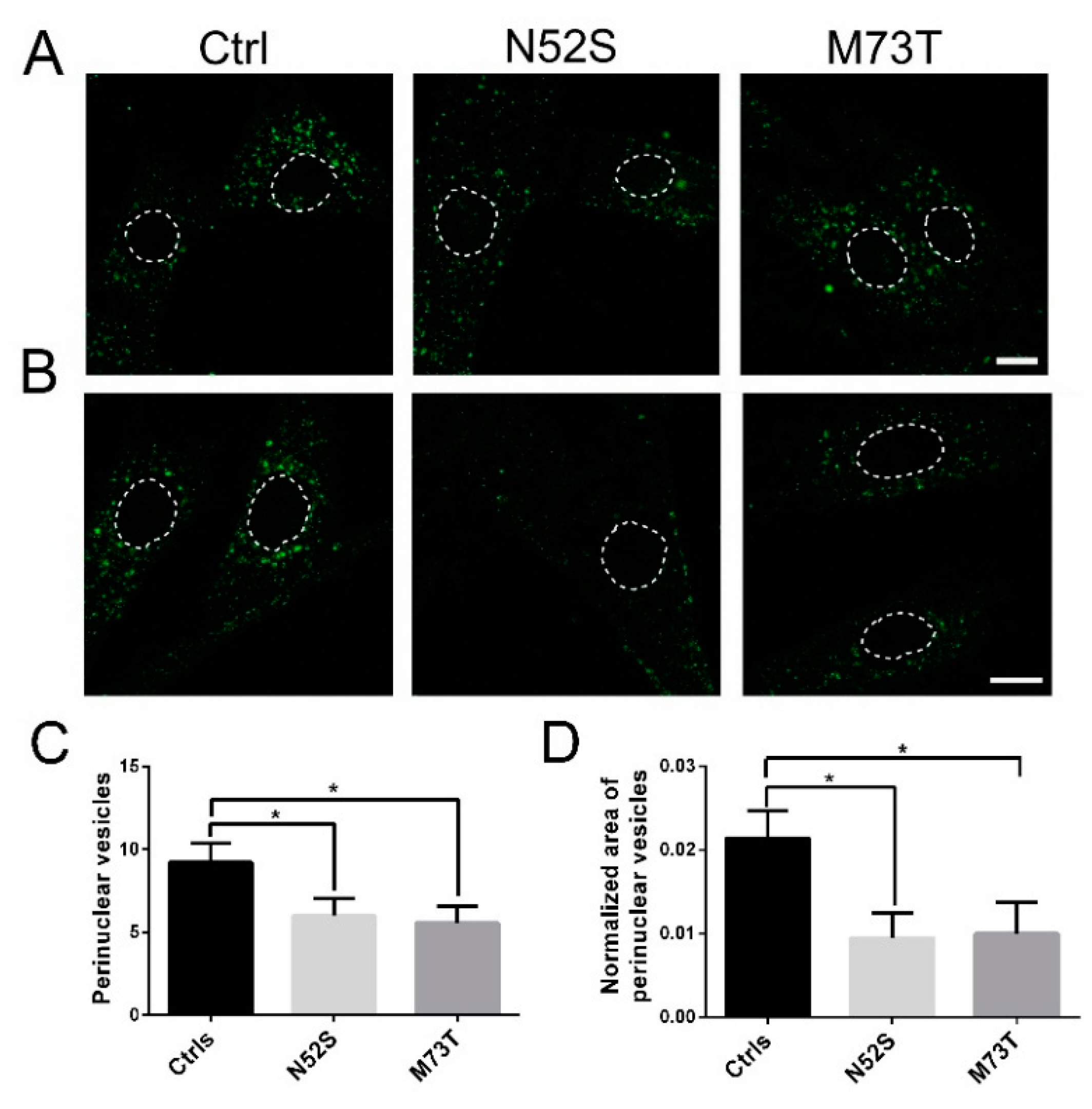
2.3. EGF and Tf Transport Are Impaired in M73T and N52S Mutant Fibroblasts
2.4. N52S and M73T Variants Affect Fibroblasts’ Migration
2.5. Structural Modeling of TUBB Variants
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Nocodazole Wash-out and Immunofluorescence Staining
4.3. Confocal Analysis
4.4. EGF and Tf Internalization Assay
4.5. Cell Migration Assay
4.6. Data analysis and Statistics
4.7. Homology Modeling of the Human Tubulin Beta Chain
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CSCKT | Circumferential skin creases Kunze type |
| MT | Microtubule |
| EGF | Epidermal growth factor |
| Tf | Transferrin |
| MAPs | Microtubule-associated proteins |
| PTMs | Post-translational modifications |
References
- Fallet-Bianco, C.; Laquerrière, A. Mutations in tubulin genes are frequent causes of various foetal malformations of cortical development including microlissencephaly. Acta Neuropathol. Commun. 2014, 25, 2–69. [Google Scholar] [CrossRef] [PubMed]
- Chang, B.S. Tubulinopathies and Their Brain Malformation Syndromes: Every TUB on Its Own Bottom. Epilepsy Curr. 2015, 15, 65–67. [Google Scholar] [CrossRef] [PubMed]
- Romaniello, R.; Arrigoni, F. Tubulin genes and malformations of cortical development. Eur. J. Med. Genet. 2018, 61, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Dent, E.W.; Gertler, F.B. Cytoskeletal Dynamics and Transport in Growth Cone Motility and Axon Guidance. Neuron. 2003, 40, 209–227. [Google Scholar] [CrossRef]
- Jaglin, X.H.; Chelly, J. Tubulin-related cortical dysgeneses: Microtubule dysfunction underlying neuronal migration defects. Trend in Genetics. 2009, 25, 555–566. [Google Scholar] [CrossRef]
- Liu, G.; Dwyer, T. Microtubule dynamics in axon guidance. Neurosci Bull. 2014, 30, 569–583. [Google Scholar] [CrossRef]
- Wu, Q.; Liu, J. The dynamics of neuronal migration. Adv. Exp. Med. Biol. 2014, 800, 25–36. [Google Scholar]
- Kapitein, L.C.; Hoogenraad, C.C. Building the Neuronal Microtubule Cytoskeleton. Neuron 2015, 87, 492–506. [Google Scholar] [CrossRef]
- Compagnucci, C.; Piermarini, E. Cytoskeletal dynamics during in vitro neurogenesis of induced pluripotent stem cells (iPSCs). Mol. Cell Neurosci. 2016, 77, 113–124. [Google Scholar] [CrossRef]
- Breuss, M.W.; Leca, I. Tubulins and brain development - The origins of functional specification. Mol. Cell Neurosci. 2017, 84, 58–67. [Google Scholar] [CrossRef]
- Poirier, K.; Keays, D.A. Large spectrum of lissencephaly and pachygyria phenotypes resulting from de novo missense mutations in tubulin alpha 1A (TUBA1A). Hum. Mut. 2007, 28, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Jaglin, X.H.; Poirier, K. Mutations in the beta-tubulin gene TUBB2B result in asymmetrical polymicrogyria. Nat. Genet. 2009, 6, 746–752. [Google Scholar] [CrossRef]
- Bahi-Buisson, N.; Poirier, K. The wide spectrum of tubulinopathies: What are the key features for the diagnosis? Brain 2014, 137, 1676–1700. [Google Scholar] [CrossRef] [PubMed]
- Poirier, K.; Saillour, Y. Mutations in the neuronal β-tubulin subunit TUBB3 result in malformation of cortical development and neuronal migration defects. Hum. Mol. Genet. 2010, 19, 4462–4473. [Google Scholar] [CrossRef] [PubMed]
- Kawauchi, T. Tubulin isotype specificity in neuronal migration: Tuba8 can’t fill in for Tuba1a. J. Cell Biol. 2017, 216, 2247–2249. [Google Scholar] [CrossRef]
- Vemu, A.; Atherton, J.A. Tubulin isoform composition tunes microtubule dynamics. Mol. Biol. Cell 2017, 28, 3564–3572. [Google Scholar] [CrossRef]
- Park, J.H.; Roll-Mecak, A. The tubulin code in neuronal polarity. Curr Opin Neurobiol. 2018, 51, 95–102. [Google Scholar] [CrossRef]
- Wawro, M.E.; Sobierajska, K. Tubulin beta 3 and 4 are involved in the generation of early fibrotic stages. Cell Signal. 2017, 38, 26–38. [Google Scholar] [CrossRef]
- Breuss, M.; Heng, J.I. Mutations in the β-tubulin gene TUBB5 cause microcephaly with structural brain abnormalities. Cell Rep. 2012, 2, 1554–1562. [Google Scholar] [CrossRef]
- Sferra, A.; Fattori, F. Defective kinesin binding of TUBB2A causes progressive spastic ataxia syndrome resembling sacsinopathy. Hum. Mol. Genet. 2018, 27, 1892–1904. [Google Scholar] [CrossRef]
- Madrigal, I.; Rabionet, R. Spectrum of clinical heterogeneity of β-tubulin TUBB5 gene mutations. Gene 2019, 695, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Wouters, L.; Rodriguez Rodriguez, C.M. Circumferential skin creases, cleft palate, typical face, intellectual disability and growth delay: “circumferential skin creases Kunze type”. Eur. J. Med. Genet. 2011, 54, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Ngo, L.; Haas, M. TUBB5 and its disease-associated mutations influence the terminal differentiation and dendritic spine densities of cerebral cortical neurons. Hum. Mol. Genet. 2014, 23, 5147–5158. [Google Scholar] [CrossRef] [PubMed]
- Isrie, M.; Breuss, M. Mutations in Either TUBB or MAPRE2 Cause Circumferential Skin Creases Kunze Type. Am. J. Hum. Genet. 2015, 97, 790–800. [Google Scholar] [CrossRef] [PubMed]
- Dentici, M.L.; Terracciano, A. Intrafamiliar clinical variability of circumferential skin creases Kunze type caused by a novel heterozygous mutation of N-terminal TUBB gene. Clin. Genet. 2018, 93, 1223–1228. [Google Scholar] [CrossRef]
- Granger, E.; McNee, G. The role of the cytoskeleton and molecular motors in endosomal dynamics. Semin. Cell Dev. Biol. 2014, 31, 20–29. [Google Scholar] [CrossRef]
- Yu, Y.; Gaillard, S. Inhibition of Spleen Tyrosine Kinase Potentiates Paclitaxel-Induced Cytotoxicity in Ovarian Cancer Cells by Stabilizing Microtubules. Cancer Cell. 2015, 28, 82–96. [Google Scholar] [CrossRef]
- Ganguly, A.; Yang, H.J. The role of microtubules and their dynamics in cell migration. Biol. Chem. 2012, 287, 43359–43369. [Google Scholar] [CrossRef]
- Allan, V.J.; Schroer, T.A. Membrane motors. Curr. Opin. Cell Biol. 1999, 11, 476–482. [Google Scholar] [CrossRef]
- Cole, N.B.; Lippincott-Schwartz, J. Organization of organelles and membrane traffic by microtubules. Curr. Opin. Cell Biol. 1995, 7, 55–64. [Google Scholar] [CrossRef]
- Matteoni, R.; Kreis, T.E. Translocation and clustering of endosomes and lysosomes depends on microtubules. J. Cell Biol. 1987, 105, 1253–1265. [Google Scholar] [CrossRef] [PubMed]
- Gruenberg, J.; Griffiths, G. Characterization of the early endosome and putative endocytic carrier vesicles in vivo and with an assay of vesicle fusion in vitro. J. Cell Biol. 1989, 108, 1301–1316. [Google Scholar] [CrossRef] [PubMed]
- Bayer, N.; Schober, D. Effect of bafilomycin A1 and nocodazole on endocytic transport in HeLa cells: Implications for viral uncoating and infection. J. Virol. 1998, 72, 9645–9655. [Google Scholar] [CrossRef] [PubMed]
- Baravalle, G.; Schober, D. Transferrin recycling and dextran transport to lysosomes is differentially affected by bafilomycin, nocodazole, and low temperature. Cell Tissue Res. 2005, 32, 99–113. [Google Scholar] [CrossRef]
- Elkjaer, M.L.; Birn, H. Effects of microtubule disruption on endocytosis, membrane recycling and polarized distribution of Aquaporin-1 and gp330 in proximal tubule cells. Eur. J. Cell Biol. 1995, 67, 57–72. [Google Scholar]
- Pooja, S.; Pushpanathan, M. Endocytosis‒Mediated Invasion and Pathogenicity of Streptococcus agalactiae in Rat Cardiomyocyte (H9C2). PLoS ONE 2015, 10, e0139733. [Google Scholar] [CrossRef]
- Jeon, H.; Go, Y. Functional selection of phagocytosis-promoting genes: Cell sorting-based selection. J. Biomol. Screen. 2010, 15, 949–955. [Google Scholar] [CrossRef]
- Wilson, B.J.; Allen, J.L. Vesicle trafficking pathways that direct cell migration in 3D matrices and in vivo. Traffic 2018, 19, 899–909. [Google Scholar] [CrossRef]
- Fletcher, S.J.; Rappoport, J.Z. The role of vesicle trafficking in epithelial cell motility. Biochem Soc Trans. 2009, 37, 1072–1076. [Google Scholar] [CrossRef]
- Miettinen, P.J.; Huotari, M. Impaired migration and delayed differentiation of pancreatic islet cells in mice lacking EGF-receptors. Development. 2000, 127, 2617–2627. [Google Scholar]
- Shirakata, Y.; Kimura, R. Heparin-binding EGF-like growth factor accelerates keratinocyte migration and skin wound healing. J. Cell Sci. 2005, 118, 2363–2370. [Google Scholar] [CrossRef] [PubMed]
- Etienne-Manneville, S. Actin and Microtubules in Cell Motility: Which One is in Control? Traffic 2004, 5, 470–477. [Google Scholar] [CrossRef]
- Akhshi, T.K.; Wernike, D. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton (Hoboken) 2014, 71, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Gupton, S.L.; Anderson, K.L. Cell migration without a lamellipodium: Translation of actin dynamics into cell movement mediated by tropomyosin. J. Cell Biol. 2015, 168, 619–631. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, R.; Ehrlicher, A. The forces behind cell movement. International. J. Biol. Sci. 2007, 3, 303–317. [Google Scholar]
- O’Rourke, N.A.; Chenn, A. Postmitotic neurons migrate tangentially in the cortical ventricular zone. Development. 1997, 124, 997–1005. [Google Scholar]
- Hu, N.; Strobl-Mazzulla, P.H. Epigenetic regulation in neural crest development. Dev. Biol. 2014, 396, 159–168. [Google Scholar] [CrossRef]
- Moffat, J.J.; Ka, M. Genes and brain malformations associated with abnormal neuron positioning. Mol. Brain. 2015, 8, 72. [Google Scholar] [CrossRef]
- Sun, X.Z.; Takahashi, S. Normal and abnormal neuronal migration in the developing cerebral cortex. J. Med. Invest. 2002, 49, 97–110. [Google Scholar]
- Nasrallah, I.M.; Golden, J.A. Brain malformations associated with cell migration. Pediatr Dev. Pathol. 2006, 9, 89–97. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sferra, A.; Petrini, S.; Bellacchio, E.; Nicita, F.; Scibelli, F.; Dentici, M.L.; Alfieri, P.; Cestra, G.; Bertini, E.S.; Zanni, G. TUBB Variants Underlying Different Phenotypes Result in Altered Vesicle Trafficking and Microtubule Dynamics. Int. J. Mol. Sci. 2020, 21, 1385. https://doi.org/10.3390/ijms21041385
Sferra A, Petrini S, Bellacchio E, Nicita F, Scibelli F, Dentici ML, Alfieri P, Cestra G, Bertini ES, Zanni G. TUBB Variants Underlying Different Phenotypes Result in Altered Vesicle Trafficking and Microtubule Dynamics. International Journal of Molecular Sciences. 2020; 21(4):1385. https://doi.org/10.3390/ijms21041385
Chicago/Turabian StyleSferra, Antonella, Stefania Petrini, Emanuele Bellacchio, Francesco Nicita, Francesco Scibelli, Maria Lisa Dentici, Paolo Alfieri, Gianluca Cestra, Enrico Silvio Bertini, and Ginevra Zanni. 2020. "TUBB Variants Underlying Different Phenotypes Result in Altered Vesicle Trafficking and Microtubule Dynamics" International Journal of Molecular Sciences 21, no. 4: 1385. https://doi.org/10.3390/ijms21041385
APA StyleSferra, A., Petrini, S., Bellacchio, E., Nicita, F., Scibelli, F., Dentici, M. L., Alfieri, P., Cestra, G., Bertini, E. S., & Zanni, G. (2020). TUBB Variants Underlying Different Phenotypes Result in Altered Vesicle Trafficking and Microtubule Dynamics. International Journal of Molecular Sciences, 21(4), 1385. https://doi.org/10.3390/ijms21041385