Molecular Evidences for the Interactions of Auxin, Gibberellin, and Cytokinin in Bent Peduncle Phenomenon in Rose (Rosa sp.)

 , and
, and
Abstract
1. Introduction
2. Results
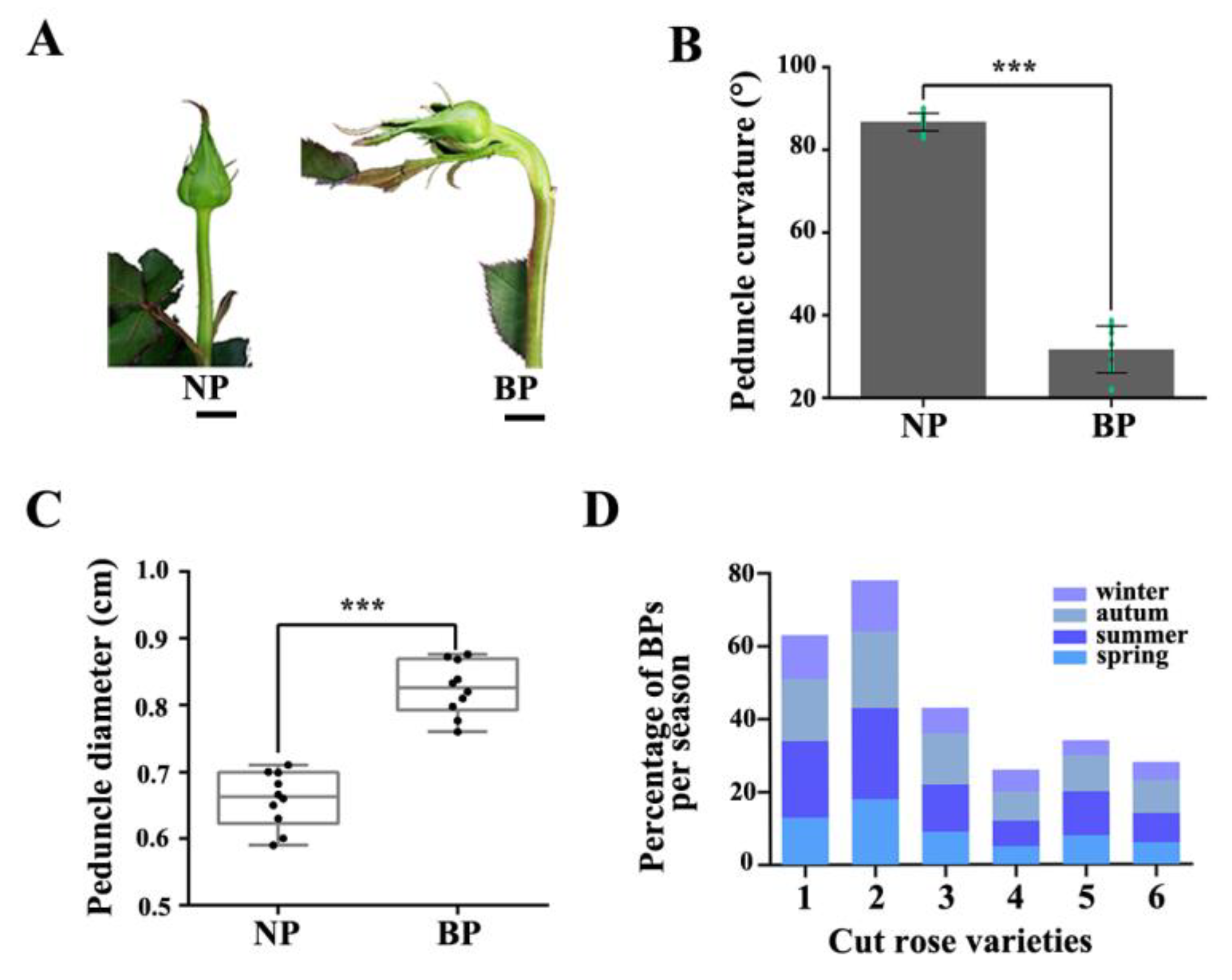
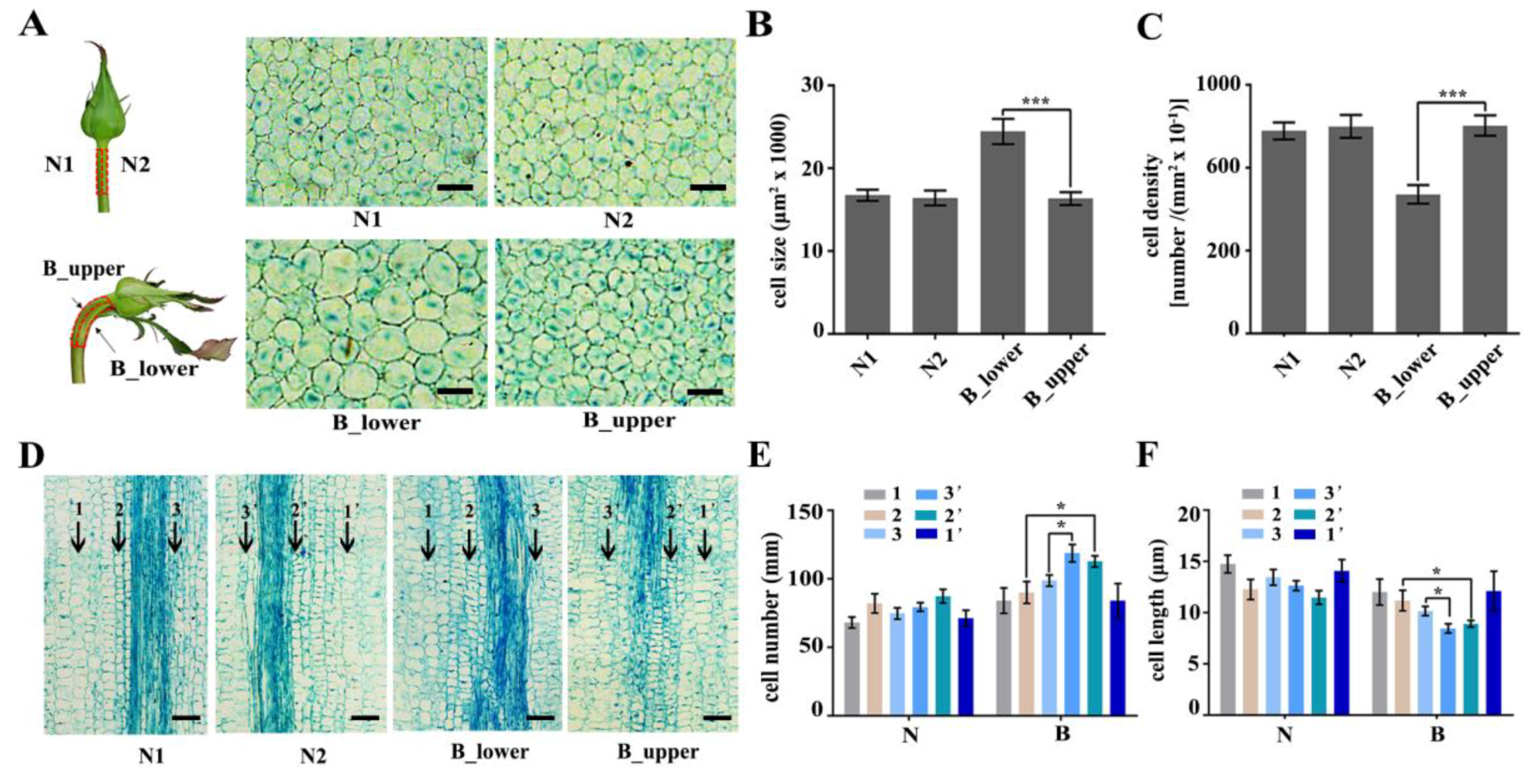
2.1. Morphological Characteristics and Cellular Aspects of BP in Rose
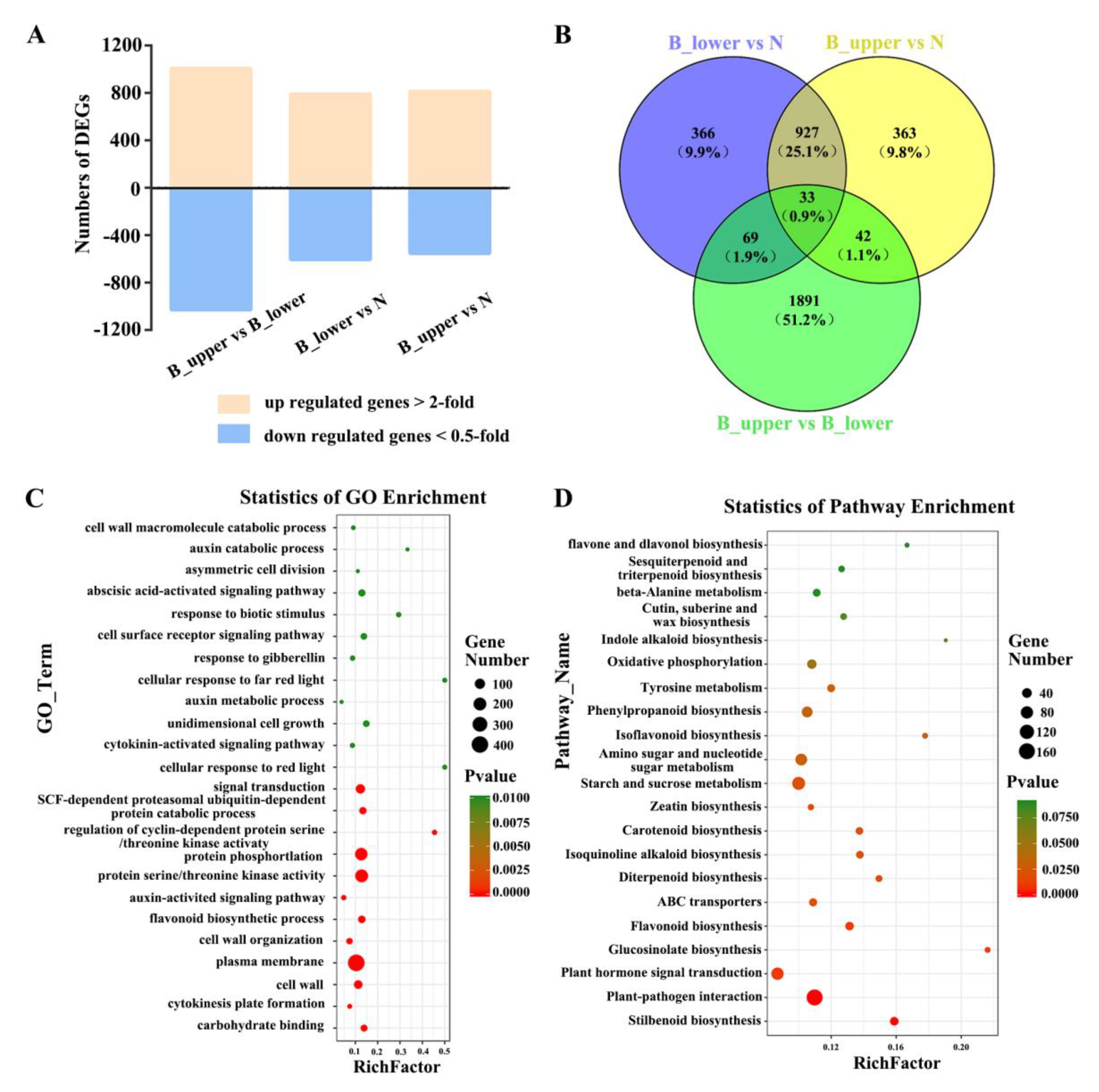
2.2. Transcriptome Sequencing of BP in Rose
2.3. Dynamic Transcriptome of BP
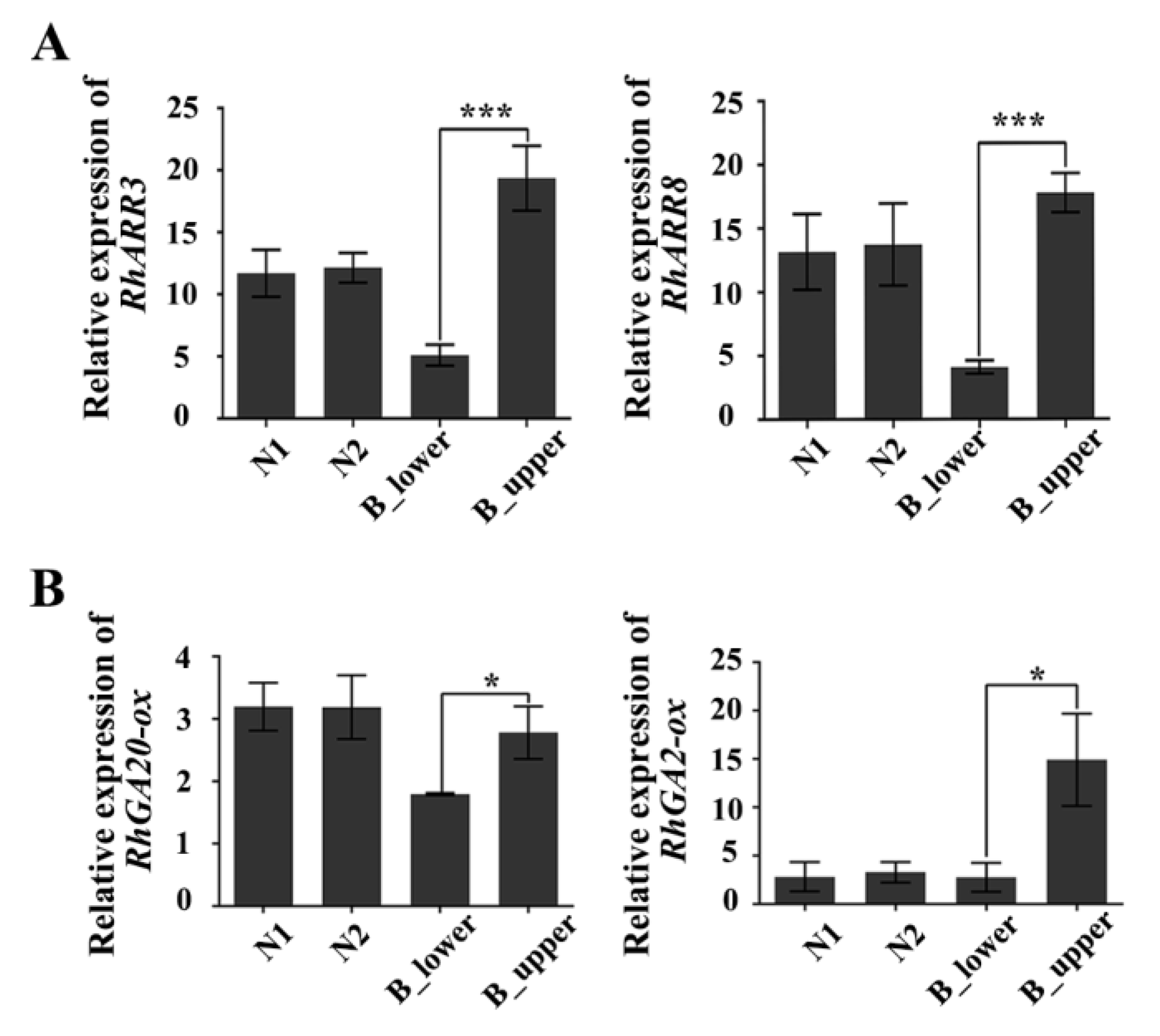
2.4. The Phytohormones Auxin, Cytokinin, and Gibberellin are Involved in BPP
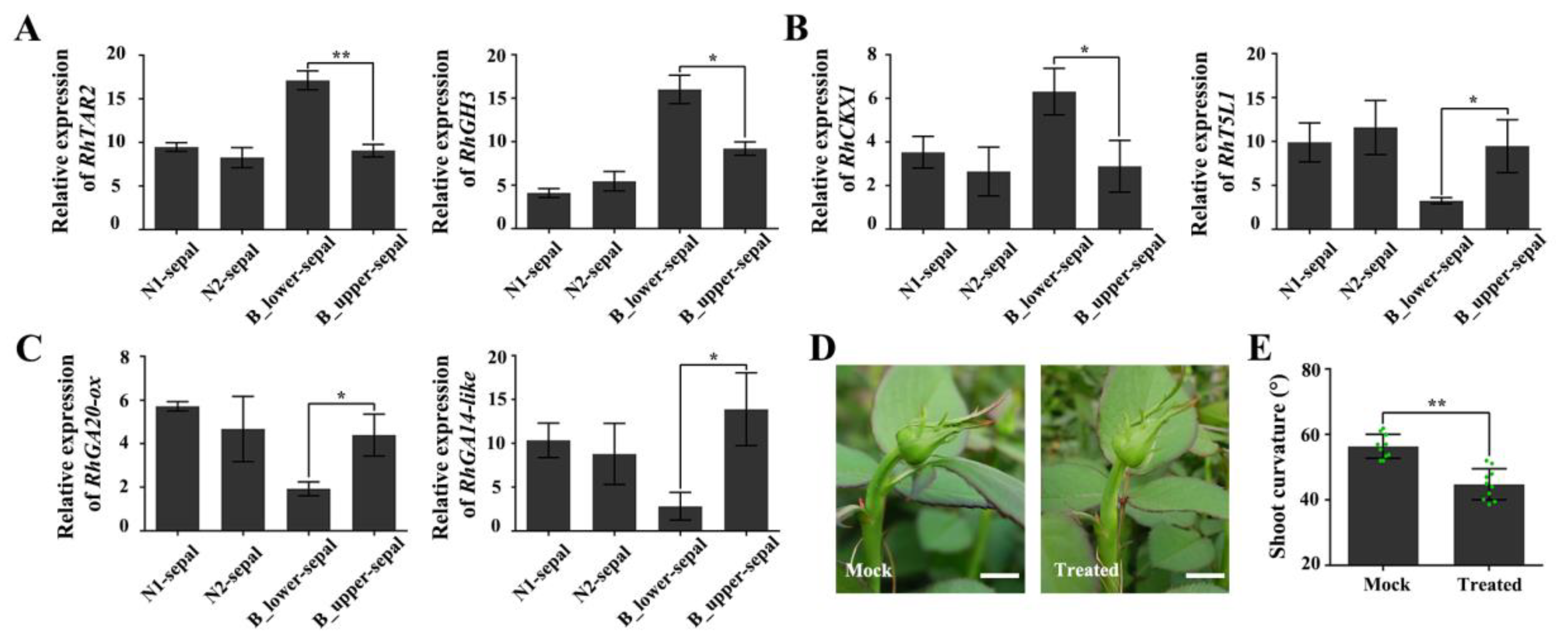
2.5. The Effect of Enlarged, Deformed Sepals on BPP
3. Discussion
3.1. BPP is a Physiological Disorder with High Frequency in Rose
3.2. Auxins, Cytokinins, and Gibberellins Regulate the Cell Division and Expansion of BP
3.3. The Deformed Sepal Affects BP Development
4. Materials and Methods
4.1. Plant Material
4.2. Total RNA Extraction and RNA-Seq Library Preparation
4.3. RNA-Seq Data Processing, Assembly, and Annotation
4.4. Paraffin Section
4.5. Quantitative RT-PCR
4.6. Removal of Deformed Sepal
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zaccai, M.; Ackerman, R.; Genis, O.; Riov, J.; Zik, M. The bent peduncle phenomenon in roses is a developmental process involving auxin. Plant Sci. 2009, 176, 736–743. [Google Scholar] [CrossRef]
- Seo, J.H.; Kim, W.S. Growth, Floral Morphology, and Phytohormone Levels of Flowering Shoots with Bent Peduncle in Greenhouse-grown Cut Rose ‘Beast’. Korean J. Horticult. Sci. Tech. 2013, 31, 714–719. [Google Scholar] [CrossRef][Green Version]
- Raz, V.; Koornneef, M. Cell division activity during apical hook development. Plant Physiol. 2001, 125, 219–226. [Google Scholar] [CrossRef]
- Powers, S.K.; Strader, L.C. Regulation of auxin transcriptional responses. Dev. Dyn. 2019, 139, 1–13. [Google Scholar]
- Choi, H.S.; Seo, M.; Cho, H.T. Two TPL-Binding Motifs of ARF2 Are Involved in Repression of Auxin Responses. Front Plant Sci. 2018, 9, 372. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, S.; Friml, J. Auxin: a trigger for change in plant development. Cell. 2009, 136, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.H.; Estelle, M. Diversity and specificity: auxin perception and signaling through the TIR1/AFB pathway. Curr. Opin. Plant Biol. 2014, 21, 51–58. [Google Scholar] [CrossRef]
- Nakazawa, M.; Yabe, N.; Ichikawa, T.; Yamamoto, Y.Y.; Yoshizumi, T.; Hasunuma, K.; Matsui, M. DFL1, an auxin-responsive GH3 gene homologue, negatively regulates shoot cell elongation and lateral root formation, and positively regulates the light response of hypocotyl length. Plant J. 2001, 25, 213–221. [Google Scholar] [CrossRef]
- Takase, T.; Nakazawa, M.; Ishikawa, A.; Kawashima, M.; Ichikawa, T.; Takahashi, N.; Shimada, H.; Manabe, K.; Matsui, M. ydk1-D, an auxin-responsive GH3 mutant that is involved in hypocotyl and root elongation. Plant J. 2004, 37, 471–483. [Google Scholar] [CrossRef]
- Pelletier, S.; Van Orden, J.; Wolf, S.; Vissenberg, K.; Delacourt, J.; Ndong, Y.A.; Pelloux, J.; Bischoff, V.; Urbain, A.; Mouille, G.; et al. A role for pectin de-methylesterification in a developmentally regulated growth acceleration in dark-grown Arabidopsis hypocotyls. New Phytol. 2010, 188, 726–739. [Google Scholar] [CrossRef]
- Perrot-Rechenmann, C. Cellular Responses to Auxin: Division versus Expansion. Cold Spring Harb. Perspect. Biol. 2010, 2, a001446. [Google Scholar] [CrossRef] [PubMed]
- Esmon, C.A.; Tinsley, A.G.; Ljung, K.; Sandberg, G.; Hearne, L.B.; Liscum, E. A gradient of auxin and auxin-dependent transcription precedes tropic growth responses. Proc. Natl. Acad. Sci. USA 2006, 103, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Immanen, J.; Nieminen, K.; Smolander, O.P.; Kojima, M.; Alonso Serra, J.; Koskinen, P.; Zhang, J.; Elo, A.; Mahonen, A.P.; Street, N.; et al. Cytokinin and Auxin Display Distinct but Interconnected Distribution and Signaling Profiles to Stimulate Cambial Activity. Curr. Biol. 2016, 26, 1990–1997. [Google Scholar] [CrossRef] [PubMed]
- Kakimoto, T. Identification of plant cytokinin biosynthetic enzymes as dimethylallyl diphosphate: ATP/ADP isopentenyltransferases. Plant Cell Physiol. 2001, 42, 677–685. [Google Scholar] [CrossRef]
- Hwang, I.; Sheen, J.; Muller, B. Cytokinin Signaling Networks. Annu. Rev. Plant Biol. 2012, 63, 353–380. [Google Scholar] [CrossRef]
- Miyawaki, K.; Tarkowski, P.; Matsumoto-Kitano, M.; Kato, T.; Sato, S.; Tarkowska, D.; Tabata, S.; Sandberg, G.; Kakimoto, T. Roles of Arabidopsis ATP/ADP isopentenyltransferases and tRNA isopentenyltransferases in cytokinin biosynthesis. Proc. Natl. Acad. Sci. USA 2006, 103, 16598–16603. [Google Scholar] [CrossRef]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmulling, T. Cytokinin Regulates the Activity of Reproductive Meristems, Flower Organ Size, Ovule Formation, and Thus Seed Yield in Arabidopsis thaliana. Plant Cell. 2011, 23, 69–80. [Google Scholar] [CrossRef]
- D’agostino, I.B.; Deruere, J.; Kieber, J.J. Characterization of the response of the arabidopsis response regulator gene family to cytokinin. Plant Physiol. 2000, 124, 1706–1717. [Google Scholar] [CrossRef]
- Ren, B.; Liang, Y.; Deng, Y.; Chen, Q.G.; Zhang, J.; Yang, X.H.; Zuo, J.R. Genome-wide comparative analysis of type-A Arabidopsis response regulator genes by overexpression studies reveals their diverse roles and regulatory mechanisms in cytokinin signaling. Cell Res. 2009, 19, 1178–1190. [Google Scholar] [CrossRef]
- Roberts, A.V.; Blake, P.S.; Lewis, R.; Taylor, J.M.; Dunstan, D.I. The Effect of Gibberellins on Flowering in Roses. J. Plant Growth Regul. 1999, 18, 113–119. [Google Scholar] [CrossRef]
- Belay, T.A.; Volker, M.; Tatjana, P.; Čož-Rakovac, R.; Jocelyn, A.; OzgaDennis, M.R.; Lewis, N.M.; Yuji, K.; Shinjiro, Y.; Branka, S. Endogenous Gibberellin Profile During Christmas Rose (Helleborus niger L.) Flower and Fruit Development. J. Plant Growth Regul. 2010, 29, 194–209. [Google Scholar]
- Ganelevin, R.; Zieslin, N. Contribution of sepals and gibberellin treatments to growth and development of rose (Rosa hybrida) flowers. Plant Growth Regul. 2002, 37, 255–261. [Google Scholar] [CrossRef]
- Zieslin, N.; Starkman, F.; Zamski, E. Growth of rose flower peduncles and effects of applied plant growth regulators. Plant Growth Regul 1989, 8, 65–76. [Google Scholar]
- Ibanes, M.; Fàbregas, N.; Chory, J.; Caño-Delgado, A.I. Brassinosteroid signaling and auxin transport are required to establish the periodic pattern of Arabidopsis shoot vascular bundles. Proc. Natl. Acad. Sci. USA 2009, 106, 13630–13635. [Google Scholar] [CrossRef]
- Daviere, J.M.; De Lucas, M.; Prat, S. Transcriptional factor interaction: a central step in DELLA function. Curr. Opin. Genet. Dev. 2008, 18, 295–303. [Google Scholar] [CrossRef]
- Dayan, J.; Voronin, N.; Gong, F.; Sun, T.P.; Hedden, P.; Fromm, H.; Aloni, A. Leaf-Induced Gibberellin Signaling Is Essential for Internode Elongation, Cambial Activity, and Fiber Differentiation in Tobacco Stems. Plant Cell. 2012, 24, 66–79. [Google Scholar] [CrossRef]
- Sun, T.P.; Gubler, F. Molecular mechanism of gibberellin signaling in plants. Annu Rev Plant Biol. 2004, 55, 197–223. [Google Scholar] [CrossRef]
- Sachs, R.M.; Bretz, C.F.; Lang, A. Shoot Histogenesis - the Early Effects of Gibberellin Upon Stem Elongation in 2 Rosette Plants. Am. J. Bot. 1959, 46, 376–384. [Google Scholar] [CrossRef]
- Zhu, Y.Y.; Nomura, T.; Xu, Y.H.; Zhang, Y.Y.; Peng, Y.; Mao, B.Z.; Hanada, A.; Zhou, H.C.; Wang, R.X.; Li, P.J.; et al. ELONGATED UPPERMOST INTERNODE encodes a cytochrome P450 monooxygenase that epoxidizes gibberellins in a novel deactivation reaction in rice. Plant Cell. 2006, 18, 442–456. [Google Scholar] [CrossRef]
- An, F.; Zhang, X.; Zhu, Z.; Ji, Y.; He, W.; Jiang, Z.; Li, M.; Guo, H. Coordinated regulation of apical hook development by gibberellins and ethylene in etiolated Arabidopsis seedlings. Cell Res. 2012, 22, 915–927. [Google Scholar] [CrossRef]
- Kaneko, M.; Itoh, H.; Ueguchi-Tanaka, M.; Ashikari, M.; Matsuoka, M. The alpha-amylase induction in endosperm during rice seed germination is caused by gibberellin synthesized in epithelium. Plant Physiol. 2002, 128, 1264–1270. [Google Scholar] [CrossRef] [PubMed]
- Sakai, M.; Sakamoto, T.; Saito, T.; Matsuoka, M.; Tanaka, H.; Kobayashi, M. Expression of novel rice gibberellin 2-oxidase gene is under homeostatic regulation by biologically active gibberellins. J Plant Res. 2003, 116, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Zhang, F.; Yang, Y.; Ma, Y.; Liu, Y.X.; Li, H.; Dai, H.Y.; Zhang, Z.H. Modification of Plant Height via RNAi Suppression of MdGA20-ox Gene Expression in Apple. J. Am. Soc. Hortic. Sci. 2016, 141, 242–248. [Google Scholar] [CrossRef]
- Schrager-Lavelle, A.; Gath, N.N.; Devisetty, U.K.; Carrera, E.; Lopez-Diaz, I.; Blazquez, M.A.; Maloof, J.N. The role of a class III gibberellin 2-oxidase in tomato internode elongation. Plant J. 2019, 97, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Kepinski, S.; Leyser, O. The Arabidopsis F-box protein TIR1 is an auxin receptor. Nat. 2005, 435, 446–451. [Google Scholar] [CrossRef]
- Cao, M.; Chen, R.; Li, P.; Yu, Y.; Zheng, R.; Ge, D.; Zheng, W.; Wang, X.; Gu, Y.; Gelova, Z.; et al. TMK1-mediated auxin signalling regulates differential growth of the apical hook. Nat. 2019, 568, 240–243. [Google Scholar] [CrossRef]
- Wu, L.; Ma, N.; Jia, Y.C.; Zhang, Y.; Feng, M.; Jiang, C.Z.; Ma, C.; Gao, J.P. An Ethylene-Induced Regulatory Module Delays Flower Senescence by Regulating Cytokinin Content. Plant Physiol. 2017, 173, 853–862. [Google Scholar] [CrossRef]
- Muday, G.K. Auxins and tropisms. J Plant Growth Regul. 2001, 20, 226–243. [Google Scholar] [CrossRef]
- Philosoph-Hadas, S.; Friedman, H.; Meir, S. Gravitropic bending and plant hormones. Plant Horm. 2005, 72, 31–78. [Google Scholar]
- Wisniewska, J.; Xu, J.; Seifertova, D.; Brewer, P.B.; Ruzicka, K.; Blilou, I.; Rouquie, D.; Benkova, E.; Scheres, B.; Friml, J. Polar PIN localization directs auxin flow in plants. Science 2006, 312, 883. [Google Scholar] [CrossRef]
- Werner, T.; Motyka, V.; Strnad, M.; Schmulling, T. Regulation of plant growth by cytokinin. Proc. Natl. Acad. Sci. USA 2001, 98, 10487–10492. [Google Scholar] [CrossRef]
- Mahonen, A.P.; Bishopp, A.; Higuchi, M.; Nieminen, K.M.; Kinoshita, K.; Tormakangas, K.; Ikeda, Y.; Oka, A.; Kakimoto, T.; Helariutta, Y. Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science 2006, 311, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef] [PubMed]
- Meyer, V.G. Flower Abnormalities. Bot. Rev. 1966, 32, 165–218. [Google Scholar] [CrossRef]
- Aloni, R.; Aloni, E.; Langhans, M.; Ullrich, C.I. Role of cytokinin and auxin in shaping root architecture: regulating vascular differentiation, lateral root initiation, root apical dominance and root gravitropism. Ann Bot. 2006, 97, 883–893. [Google Scholar] [CrossRef]
- Ma, C.; Wang, H.; Macnish, A.J.; Estrada-Melo, A.C.; Lin, J.; Chang, Y.H.; Reid, M.S.; Jiang, C.Z. Transcriptomic analysis reveals numerous diverse protein kinases and transcription factors involved in desiccation tolerance in the resurrection plant Myrothamnus flabellifolia. Hort. Res. 2015, 2, 47. [Google Scholar] [CrossRef]
- Meng, Y.L.; Li, N.; Tian, J.; Gao, J.P.; Zhang, C.Q. Identification and validation of reference genes for gene expression studies in postharvest rose flower (Rosa hybrida). Sci. Hortic. 2013, 158, 16–21. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, Q.C.; Zeng, D.X.; Xu, J.H.; Zhou, H.G.; Wang, F.L.; Ma, N.; Li, Y.H. RhMYB108, an R2R3-MYB transcription factor, is involved in ethylene- and JA-induced petal senescence in rose plants. Hortic. Res. 2019, 6, 131. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Total |
|---|---|
| No. of reads | 398,886,448 |
| No. of valid reads | 395,462,558 |
| No. of mapped reads | 334,973,525 |
| No. of assembled transcripts | 260,541 |
| Average length of transcripts | 2021 bp |
| Total length of transcripts | 526,553,361 bp |
| Gene ID | FC | Log2(FC) | Up/Down Regulation | Annotation |
|---|---|---|---|---|
| Response to auxin | ||||
| RchiOBHmChr1g0342931 | 3.94 | 1.98 | up | Carotenoid cleavage dioxygenase 7 |
| RchiOBHmChr3g0478151 | 4.25 | 2.09 | up | Ubiquitin-like protein Nedd8 |
| RchiOBHmChr3g0478171 | 2.96 | 1.57 | up | Ubiquitin-like protein Nedd8 |
| RchiOBHmChr4g0392981 | 2.03 | 1.02 | up | Transcription factor MYB15 |
| RchiOBHmChr4g0424331 | 2.23 | 1.16 | up | Transcription factor MYB12 |
| RchiOBHmChr5g0056751 | 3.13 | 1.65 | up | Auxin-induced protein 15A-like |
| RchiOBHmChr6g0268541 | 26.95 | 4.75 | up | Auxin-responsive protein SAUR50-like |
| RchiOBHmChr6g0305471 | 10.96 | 3.45 | up | Auxin-responsive protein SAUR71-like |
| RchiOBHmChr7g0210641 | 2.20 | 1.14 | up | Auxin-responsive protein SAUR14 |
| RchiOBHmChr7g0210711 | 40.28 | 5.33 | up | Auxin-responsive protein SAUR3 |
| RchiOBHmChr7g0210731 | 40.28 | 5.33 | up | Auxin-responsive protein SAUR20 |
| RchiOBHmChr7g0210801 | 1.40 | 0.48 | up | Auxin-responsive protein SAUR23 |
| RchiOBHmChr1g0324921 | 0.21 | −2.25 | down | Auxin-induced protein X15 |
| RchiOBHmChr1g0362991 | 0.30 | −1.75 | down | Indole-3-acetic acid-induced protein ARG7-like |
| RchiOBHmChr4g0422041 | 0.36 | −1.49 | down | Flowering-promoting factor 1-like protein 3 |
| RchiOBHmChr5g0048841 | 0.36 | −1.47 | down | Vacuolar-related NAC-domain 6 |
| RchiOBHmChr7g0202161 | 0.40 | −1.32 | down | Auxin-responsive protein SAUR36-like |
| RchiOBHmChr7g0209651 | 0.21 | −2.27 | down | Auxin-responsive protein SAUR40 |
| RchiOBHmChr7g0188911 | 0.60 | −0.75 | down | Auxin-responsive factor 2 ARF2 |
| RchiOBHmChr2g0095551 | 0.72 | −0.48 | down | Auxin-responsive factor 7 ARF7 |
| RchiOBHmChr6g0254401 | 0.67 | −0.58 | down | Auxin-responsive GH3 family protein GH3 |
| Auxin-activated signaling pathway | ||||
| RchiOBHmChr4g0428791 | 2.35 | 1.23 | up | Auxin-responsive protein 31 IAA31 |
| RchiOBHmChr6g0311541 | 2.37 | 1.25 | up | BIG GRAIN 1-like B |
| RchiOBHmChr7g0224691 | 2.24 | 1.16 | up | Arabidopsis SKP1 homologue 1 ASK1 |
| RchiOBHmChr3g0451081 | 0.49 | −1.03 | down | Indole-3-acetic acid inducible 4 like IAA4-like |
| RchiOBHmChr4g0389611 | 0.57 | −0.80 | down | Indole-3-acetic acid inducible 5 IAA5 |
| RchiOBHmChr2g0137311 | 0.52 | −0.94 | down | Indole-3-acetic acid inducible 6 IAA6 |
| RchiOBHmChr4g0434391 | 0.55 | −0.87 | down | Indole-3-acetic acid inducible 32 IAA32 |
| Auxin homeostasis | ||||
| RchiOBHmChr2g0132571 | 2.92 | 1.55 | up | Shikimate O-hydroxycinnamoyl transferase-like |
| RchiOBHmChr2g0132651 | 3.35 | 1.74 | up | Shikimate O-hydroxycinnamoyl transferase-like |
| RchiOBHmChr5g0066541 | 0.35 | −1.52 | down | Shikimate O-hydroxycinnamoyltransferase-like |
| RchiOBHmChr6g0302041 | 0.20 | −2.30 | down | Transcription factor SHI RELATED SEQUENCE 1-like |
| Auxin biosynthetic process | ||||
| RchiOBHmChr6g0266901 | 0.45 | −1.16 | down | Transcription factor SHI RELATED SEQUENCE 3-like |
| RchiOBHmChr7g0192941 | 0.25 | −2.01 | down | Trytophan aminotransferase related 2 TAR2 |
| RchiOBHmChr6g0292661 | 0.56 | −0.84 | down | Transport inhibitor response 1 TIR1 |
| RchiOBHmChr3g0492431 | 0.57 | −0.80 | down | Receptor-like transmembrane kinase I TMK1 |
| Cellular response to auxin stimulus | ||||
| RchiOBHmChr1g0318281 | 0.32 | −1.62 | down | Transcription factor agamous-like MADS-box AGL15 |
| RchiOBHmChr4g0389411 | 0.01 | −7.00 | down | Enhancer of mRNA-decapping 4-like |
| Auxin catabolic process | ||||
| RchiOBHmChr2g0096111 | 10.10 | 3.34 | up | 2-Oxoglutarate-dependent dioxygenase DAOlike |
| Auxin metabolic process | ||||
| RchiOBHmChr4g0444701 | 2.33 | 1.22 | up | IAA-amino acid hydrolase ILR1-like 4 |
| Auxin transport | ||||
| RchiOBHmChr6g0305141 | 0.37 | −1.45 | down | UDP-glucose 4,6-dehydratase |
| RchiOBHmChr4g0441311 | 0.63 | −0.67 | down | PIN-FORMED 3 PIN3 |
| RchiOBHmChr3g0470311 | 0.70 | −0.52 | down | ATP-binding cassette B1 ABCB1 |
| RchiOBHmChr2g0168761 | 0.59 | −0.77 | down | ATP-binding cassette B19 ABCB19 |
| RchiOBHmChr6g0291961 | 0.68 | −0.57 | down | Auxin binding protein 1 ABP1 |
| RchiOBHmChr2g0162401 | 2.26 | 1.17 | up | Serine/threonine kinase |
| Gene ID | FC | Log2(FC) | Up/Down Regulation | Annotation |
|---|---|---|---|---|
| Gibberellic acid-mediated signaling pathway | ||||
| RchiOBHmChr2g0159111 | 2.88 | 1.53 | up | Transcription factor MYB101 |
| RchiOBHmChr6g0309121 | 36.45 | 5.19 | up | Gibberellic acid stimulated Arabidopsis 10 |
| RchiOBHmChr2g0156431 | 0.44 | −1.20 | down | F-box protein SNE |
| RchiOBHmChr2g0170521 | 0.34 | −1.57 | down | Gibberellin-regulated 14-like GA14-like |
| Response to gibberellin | ||||
| RchiOBHmChr1g0378621 | 2.27 | 1.18 | up | Transcription factor WRKY 27 |
| RchiOBHmChr4g0444681 | 5.42 | 2.44 | up | Gibberellin-regulated 3-like |
| RchiOBHmChr5g0062851 | 2.18 | 1.12 | up | Transcription factor MYB48-like |
| Gibberellin biosynthetic process | ||||
| RchiOBHmChr1g0353791 | 3.41 | 1.77 | up | Gibberellin 20 oxidase GA20-ox |
| RchiOBHmChr2g0089721 | 0.17 | –2.58 | down | Gibberellin 3-beta-dioxygenase 1-like |
| Gibberellin catabolic process | ||||
| RchiOBHmChr3g0486971 | 2.30 | 1.20 | up | Gibberellin 2-beta-dioxygenase GA2-ox |
| RchiOBHmChr1g0318281 | 0.32 | –1.62 | down | Agamous-like MADS-box AGL15 |
| Cellular response to gibberellin stimulus | ||||
| RchiOBHmChr7g0202321 | 2.07 | 1.05 | up | Acid beta-fructofuranosidase 2, vacuolar |
| Regulation of gibberellic acid-mediated signaling pathway | ||||
| RchiOBHmChr6g0302041 | 0.20 | –2.30 | down | SHI RELATED SEQUENCE 1-like |
| Gibberellin metabolic process | ||||
| RchiOBHmChr5g0003991 | 3.35 | 1.75 | up | Gibberellin 2-beta-dioxygenase 8 GA2-ox8 |
| Gene ID | FC | Log2(FC) | Up/down regulation | Annotation |
|---|---|---|---|---|
| Response to cytokinin | ||||
| RchiOBHmChr7g0238791 | 12.27 | 3.62 | up | Inosine-5 -monophosphate dehydrogenase 2-like |
| RchiOBHmChr4g0415601 | 0.16 | −2.63 | down | BAHD acyltransferase At5g47980-like |
| RchiOBHmChr4g0415621 | 0.16 | −2.66 | down | BAHD acyltransferase At5g47980-like |
| RchiOBHmChr4g0431311 | 0.24 | −2.08 | down | Deacetylvindoline O-acetyltransferase |
| RchiOBHmChr5g0048841 | 0.36 | −1.47 | down | NAC transcription factor 29-like |
| RchiOBHmChr6g0284071 | 0.41 | −1.27 | down | Wound-induced basic protein |
| RchiOBHmChr6g0304241 | 0.36 | −1.47 | down | Vinorine synthase-like |
| RchiOBHmChr4g0444831 | 1.79 | 0.84 | up | A-type response regulator 3 ARR3 |
| RchiOBHmChr2g0125351 | 1.56 | 0.64 | up | A-type response regulator 8 ARR8 |
| RchiOBHmChr1g0370561 | 1.67 | 0.74 | up | TMO5-LIKE1 T5L1 |
| Cytokinin-activated signaling pathway | ||||
| RchiOBHmChr1g0358681 | 2.82 | 1.5 | up | Ethylene-responsive transcription factor CRF4-like |
| RchiOBHmChr5g0061201 | 0.19 | −2.40 | down | Histidine-containing phosphotransfer 1-like |
| RchiOBHmChr6g0250591 | 0.37 | −1.42 | down | Histidine kinase CKI1-like |
| RchiOBHmChr6g0250601 | 0.27 | −1.89 | down | LYSM-containing receptor-like kinase 1 |
| Positive regulation of cytokinesis | ||||
| RchiOBHmChr4g0410231 | 2.26 | 1.17 | up | NEDD1 protein |
| Cytokinin catabolic process | ||||
| RchiOBHmChr1g0319331 | 0.61 | −0.72 | down | Cytokinin oxidase/dehydrogenase 1 CKX1 |
| Cytokinesis by cell plate formation | ||||
| RchiOBHmChr1g0341961 | 2.11 | 1.08 | up | Tubulin binding cofactor A |
| RchiOBHmChr1g0361251 | 3.10 | 1.63 | up | HAUS augmin-like complex subunit 2 |
| RchiOBHmChr3g0495401 | 2.02 | 1.02 | up | Cytoskeleton-associated protein 5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, W.; Zhang, S.; Fan, Y.; Deng, Y.; Wang, C.; Lu, J.; Sun, X.; Ma, N.; Shahid, M.O.; Li, Y.; et al. Molecular Evidences for the Interactions of Auxin, Gibberellin, and Cytokinin in Bent Peduncle Phenomenon in Rose (Rosa sp.). Int. J. Mol. Sci. 2020, 21, 1360. https://doi.org/10.3390/ijms21041360
Jing W, Zhang S, Fan Y, Deng Y, Wang C, Lu J, Sun X, Ma N, Shahid MO, Li Y, et al. Molecular Evidences for the Interactions of Auxin, Gibberellin, and Cytokinin in Bent Peduncle Phenomenon in Rose (Rosa sp.). International Journal of Molecular Sciences. 2020; 21(4):1360. https://doi.org/10.3390/ijms21041360
Chicago/Turabian StyleJing, Weikun, Shuai Zhang, Youwei Fan, Yinglong Deng, Chengpeng Wang, Jingyun Lu, Xiaoming Sun, Nan Ma, Muhammad Owais Shahid, Yonghong Li, and et al. 2020. "Molecular Evidences for the Interactions of Auxin, Gibberellin, and Cytokinin in Bent Peduncle Phenomenon in Rose (Rosa sp.)" International Journal of Molecular Sciences 21, no. 4: 1360. https://doi.org/10.3390/ijms21041360
APA StyleJing, W., Zhang, S., Fan, Y., Deng, Y., Wang, C., Lu, J., Sun, X., Ma, N., Shahid, M. O., Li, Y., & Zhou, X. (2020). Molecular Evidences for the Interactions of Auxin, Gibberellin, and Cytokinin in Bent Peduncle Phenomenon in Rose (Rosa sp.). International Journal of Molecular Sciences, 21(4), 1360. https://doi.org/10.3390/ijms21041360