The Role of Th17-Related Cytokines in Atopic Dermatitis


Abstract
1. Introduction
2. Th17 Cells in Pathogenesis of Psoriasis
3. Involvement of Th17-Related Cytokines in AD
3.1. Th17-Related Cytokine Expression in AD
3.2. AD Mouse Models and Th17-Related Cytokines
4. Expression of Th17 Cytokines in Other Skin Diseases
5. Clinical Trial for AD Targeting Th17 Cytokines
5.1. Clinical Effect of Ustekinumab on AD
5.2. Other Biologics and AD
6. Th2 Cytokines Are Key Players in AD
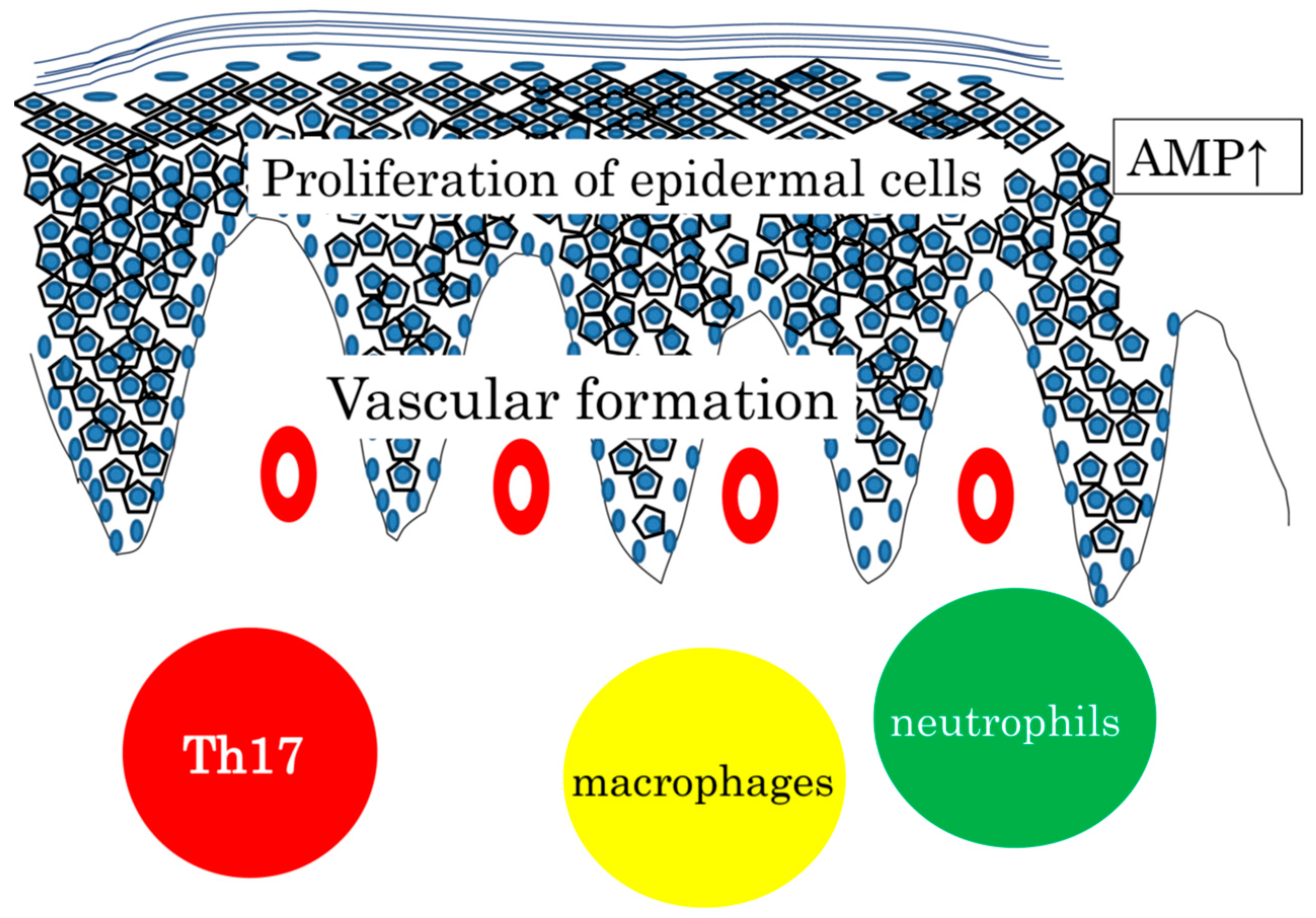
7. The Importance of Th17 Cytokines in the Skin
8. Ideal Biologics for Psoriasis, AD, and CTCL
9. Conclusions
Funding
Conflicts of Interest
Abbreviations
| AD | atopic dermatitis |
| CTCL | cutaneous T-cell lymphoma |
| TNF | tumor necrosis factor |
| Th | T helper |
| IL | interleukin |
| IFN | interferon |
| AMPs | antimicrobial peptides |
| TARC | thymus and activation-regulated chemokine |
| SCORAD | SCORing AD |
| TWEAK | TNF-like weak inducer of apoptosis |
| FGF | fibroblast growth factor |
| MF | mycosis fungoides |
| SS | Sezary syndrome |
| TSLP | thymic stromal lymphopoietin |
| MCP | monocyte chemotactic protein |
References
- Saeki, H.; Nakahara, T.; Tanaka, A.; Kabashima, K.; Sugaya, M.; Murota, H.; Ebihara, T.; Kataoka, Y.; Aihara, M.; Etoh, T.; et al. Clinical practice guidelines for the management of atopic dermatitis. J. Dermatol. 2016, 43, 1117–1145. [Google Scholar] [CrossRef] [PubMed]
- Saeki, H.; Oiso, N.; Honma, M.; Iizuka, H.; Kawada, A.; Tamaki, K. Prevalence of atopic dermatitis in Japanese adults and community validation of the U.K. diagnostic criteria. J. Dermatol. Sci. 2009, 55, 140–141. [Google Scholar] [CrossRef] [PubMed]
- Williams, H.C.; Strachan, D.P. The natural history of childhood eczema: Observations from the British 1958 birth cohort study. Br. J. Dermatol. 1998, 139, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Fukiwake, N.; Furusyo, N.; Kubo, N.; Takeoka, H.; Toyoda, K.; Morita, K.; Shibata, S.; Nakahara, T.; Kido, M.; Hayashida, S.; et al. Incidence of atopic dermatitis in nursery school children—a follow-up study from 2001 to 2004, Kyushu University Ishigaki Atopic Dermatitis Study (KIDS). Eur. J. Dermatol. 2006, 16, 416–419. [Google Scholar] [PubMed]
- Zackheim, H.S.; Koo, J.; LeBoit, P.E.; McCalmont, T.H.; Bowman, P.H.; Kashani-Sabet, M.; Jones, C.; Zehnder, J. Psoriasiform mycosis fungoides with fatal outcome after treatment with cyclosporine. J. Am. Acad. Dermatol. 2002, 47, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Miyagaki, T.; Sugaya, M. Erythrodermic cutaneous T-cell lymphoma: How to differentiate this rare disease from atopic dermatitis. J. Dermatol. Sci. 2011, 64, 1–6. [Google Scholar] [CrossRef]
- Suga, H.; Sugaya, M.; Toyama, T.; Sumida, H.; Fujita, H.; Kogure, A.; Igarashi, A.; Sato, S. A case of mycosis fungoides with large cell transformation associated with infliximab treatment. Acta. Derm. Venereol. 2014, 94, 233–234. [Google Scholar] [CrossRef]
- Xu, X.; van Galen, L.S.; Koh, M.J.A.; Bajpai, R.; Thng, S.; Yew, Y.W.; Ho, V.P.Y.; Alagappan, U.; Järbrink, K.S.A.; Car, J. Factors influencing quality of life in children with atopic dermatitis and their caregivers: A cross-sectional study. Sci. Rep. 2019, 9, 15990. [Google Scholar] [CrossRef]
- Ražnatović Đurović, M.; Janković, J.; Tomić Spirić, V.; Relić, M.; Sojević Timotijević, Z.; Ćirković, A.; Đurić, S.; Janković, S. Does age influence the quality of life in children with atopic dermatitis? PLoS ONE 2019, 14, e0224618. [Google Scholar] [CrossRef]
- Kabashima, K. New concept of the pathogenesis of atopic dermatitis: Interplay among the barrier, allergy, and pruritus as a trinity. J. Dermatol. Sci. 2013, 70, 3–11. [Google Scholar] [CrossRef]
- Salimi, M.; Barlow, J.L.; Saunders, S.P.; Xue, L.; Gutowska-Owsiak, D.; Wang, X.; Huang, L.C.; Johnson, D.; Scanlon, S.T.; McKenzie, A.N.; et al. A role for IL-25 and IL-33-driven type-2 innate lymphoid cells in atopic dermatitis. J. Exp. Med. 2013, 210, 2939–2950. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Yasuda, K.; Sakaguchi, Y.; Haneda, T.; Mizutani, H.; Yoshimoto, T.; Nakanishi, K.; Yamanishi, K. Skin-specific expression of IL-33 activates group 2 innate lymphoid cells and elicits atopic dermatitis-like inflammation in mice. Proc. Natl. Acad. Sci. USA 2013, 110, 13921–13926. [Google Scholar] [CrossRef] [PubMed]
- Van de Veen, W.; Akdis, M. The use of biologics for immune modulation in allergic disease. J. Clin. Invest. 2019, 130, 1452–1462. [Google Scholar] [CrossRef] [PubMed]
- Grewe, M.; Gyufko, K.; Schöpf, E.; Krutmann, J. Lesional expression of interferon-gamma in atopic eczema. Lancet 1994, 343, 25–26. [Google Scholar] [CrossRef]
- Czarnowicki, T.; He, H.; Canter, T.; Han, J.; Lefferdink, R.; Erickson, T.; Rangel, S.; Kameyama, N.; Kim, H.J.; Pavel, A.B.; et al. Evolution of pathologic T-cell subsets in atopic dermatitis from infancy to adulthood. J. Allergy Clin. Immunol. 2020, 145, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Koga, C.; Kabashima, K.; Shiraishi, N.; Kobayashi, M.; Tokura, Y. Possible pathogenic role of Th17 cells for atopic dermatitis. J. Invest. Dermatol. 2008, 128, 2625–2630. [Google Scholar] [CrossRef] [PubMed]
- Eyerich, K.; Pennino, D.; Scarponi, C.; Foerster, S.; Nasorri, F.; Behrendt, H.; Ring, J.; Traidl-Hoffmann, C.; Albanesi, C.; Cavani, A. IL-17 in atopic eczema: Linking allergen-specific adaptive and microbial-triggered innate immune response. J. Allergy. Clin. Immunol. 2009, 123, 59–66. [Google Scholar] [CrossRef]
- Nograles, K.E.; Zaba, L.C.; Shemer, A.; Fuentes-Duculan, J.; Cardinale, I.; Kikuchi, T.; Ramon, M.; Bergman, R.; Krueger, J.G.; Guttman-Yassky, E. IL-22-producing “T22” T cells account for upregulated IL-22 in atopic dermatitis despite reduced IL-17-producing TH17 T cells. J. Allergy. Clin. Immunol. 2009, 123, 1244–1252. [Google Scholar] [CrossRef]
- Noda, S.; Suárez-Fariñas, M.; Ungar, B.; Kim, S.J.; de Guzman Strong, C.; Xu, H.; Peng, X.; Estrada, Y.D.; Nakajima, S.; Honda, T.; et al. The Asian atopic dermatitis phenotype combines features of atopic dermatitis and psoriasis with increased TH17 polarization. J. Allergy. Clin. Immunol. 2015, 136, 1254–1264. [Google Scholar] [CrossRef]
- Esaki, H.; Brunner, P.M.; Renert-Yuval, Y.; Czarnowicki, T.; Huynh, T.; Tran, G.; Lyon, S.; Rodriguez, G.; Immaneni, S.; Johnson, D.B.; et al. Early-onset pediatric atopic dermatitis is Th2 but also Th17 polarized in skin. J. Allergy. Clin. Immunol. 2016, 138, 1639–1651. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, U.; Romano, P.; Mulcahy, L.D.; Dooley, L.T.; Baker, D.G.; Gottlieb, A.B. Efficacy and safety of infliximab monotherapy for plaque-type psoriasis: A randomised trial. Lancet 2001, 357, 1842–1847. [Google Scholar] [CrossRef]
- Leonardi, C.L.; Kimball, A.B.; Papp, K.A.; Yeilding, N.; Guzzo, C.; Wang, Y.; Li, S.; Dooley, L.T.; Gordon, K.B.; PHOENIX 1 study investigators. Efficacy and safety of ustekinumab, a human interleukin-12/23 monoclonal antibody, in patients with psoriasis: 76-week results from a randomised, double-blind, placebo-controlled trial (PHOENIX 1). Lancet 2008, 371, 1665–1674. [Google Scholar] [CrossRef]
- Leonardi, C.; Matheson, R.; Zachariae, C.; Cameron, G.; Li, L.; Edson-Heredia, E.; Braun, D.; Banerjee, S. Anti-interleukin-17 monoclonal antibody ixekizumab in chronic plaque psoriasis. N. Engl. J. Med. 2012, 366, 1190–1199. [Google Scholar] [CrossRef] [PubMed]
- Gordon, K.B.; Duffin, K.C.; Bissonnette, R.; Prinz, J.C.; Wasfi, Y.; Li, S.; Shen, Y.K.; Szapary, P.; Randazzo, B.; Reich, K. A phase 2 trial of guselkumab versus adalimumab for plaque psoriasis. N. Engl. J. Med. 2015, 373, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Jovanovic, D.V.; Di Battista, J.A.; Martel-Pelletier, J.; Jolicoeur, F.C.; He, Y.; Zhang, M.; Mineau, F.; Pelletier, J.P. IL-17 stimulates the production and expression of proinflammatory cytokines, IL-beta and TNF-alpha, by human macrophages. J. Immunol. 1998, 160, 3513–3521. [Google Scholar] [PubMed]
- Shibata, M.; Shintaku, Y.; Matsuzaki, K.; Uematsu, S. The effect of IL-17 on the production of proinflammatory cytokines and matrix metalloproteinase-1 by human periodontal ligament fibroblasts. Orthod. Craniofac. Res. 2014, 17, 60–68. [Google Scholar] [CrossRef]
- Roussel, L.; Houle, F.; Chan, C.; Yao, Y.; Bérubé, J.; Olivenstein, R.; Martin, J.G.; Huot, J.; Hamid, Q.; Ferri, L.; et al. IL-17 promotes p38 MAPK-dependent endothelial activation enhancing neutrophil recruitment to sites of inflammation. J. Immunol. 2010, 184, 4531–4537. [Google Scholar] [CrossRef]
- Jones, C.E.; Chan, K. Interleukin-17stimulates the expression of interleukin-8, growth-related oncogene-alpha, and granulocyte-colony-stimulating factor by human airway epithelial cells. Am. J. Respir. Cell. Mol. Biol. 2002, 26, 748–753. [Google Scholar] [CrossRef]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef]
- Sonnenberg, G.F.; Fouser, L.A.; Artis, D. Border patrol: Regulation of immunity, inflammation and tissue homeostasis at barrier surfaces by IL-22. Nat. Immunol. 2011, 12, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Gallo, R.L.; Nakatsuji, T. Microbial symbiosis with the innate immune defense system of the skin. J. Invest. Dermatol. 2011, 131, 1974–1980. [Google Scholar] [CrossRef] [PubMed]
- Boniface, K.; Bernard, F.X.; Garcia, M.; Gurney, A.L.; Lecron, J.C.; Morel, F. IL-22 inhibits epidermal differentiation and induces proinflammatory gene expression and migration of human keratinocytes. J. Immunol. 2005, 174, 3695–3702. [Google Scholar] [CrossRef] [PubMed]
- Dang, A.T.; Teles, R.M.; Weiss, D.I.; Parvatiyar, K.; Sarno, E.N.; Ochoa, M.T.; Cheng, G.; Gilliet, M.; Bloom, B.R.; Modlin, R.L. IL-26 contributes to host defense against intracellular bacteria. J. Clin. Investg. 2019, 129, 1926–1939. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, S.; Uchi, H.; Moroi, Y.; Furue, M. Decrease in circulating Th17 cells correlates with increased levels of CCL17, IgE and eosinophils in atopic dermatitis. J. Dermatol. Sci. 2011, 61, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Czarnowicki, T.; Gonzalez, J.; Shemer, A.; Malajian, D.; Xu, H.; Zheng, X.; Khattri, S.; Gilleaudeau, P.; Sullivan-Whalen, M.; Suárez-Fariñas, M.; et al. Severe atopic dermatitis is characterized by selective expansion of circulating TH2/TC2 and TH22/TC22, but not TH17/TC17, cells within the skin-homing T-cell population. J. Allergy. Clin. Immunol. 2015, 136, 104–115. [Google Scholar] [CrossRef]
- Romanet-Manent, S.; Charpin, D.; Magnan, A.; Lanteaume, A.; Vervloet, D.; EGEA Cooperative Group. Allergic vs nonallergic asthma: What makes the difference? Allergy 2002, 57, 607–613. [Google Scholar] [CrossRef]
- Tokura, Y. Extrinsic and intrinsic types of atopic dermatitis. J. Dermatol. Sci. 2010, 58, 1–7. [Google Scholar] [CrossRef]
- Suárez-Fariñas, M.; Dhingra, N.; Gittler, J.; Shemer, A.; Cardinale, I.; de Guzman Strong, C.; Krueger, J.G.; Guttman-Yassky, E. Intrinsic atopic dermatitis shows similar TH2 and higher TH17 immune activation compared with extrinsic atopic dermatitis. J. Allergy. Clin. Immunol. 2013, 132, 361–370. [Google Scholar] [CrossRef]
- Kamijo, H.; Miyagaki, T.; Hayashi, Y.; Akatsuka, T.; Watanabe-Otobe, S.; Oka, T.; Shishido-Takahashi, N.; Suga, H.; Sugaya, M.; Sato, S. Increased IL-26 expression promotes T helper type 17- and T helper type 2-associated cytokine production by keratinocytes in atopic dermatitis. J. Invest. Dermatol. 2019, in press. [Google Scholar] [CrossRef]
- Nakajima, S.; Kitoh, A.; Egawa, G.; Natsuaki, Y.; Nakamizo, S.; Moniaga, C.S.; Otsuka, A.; Honda, T.; Hanakawa, S.; Amano, W.; et al. IL-17A as an inducer for Th2 immune responses in murine atopic dermatitis models. J. Invest. Dermatol. 2014, 134, 2122–2130. [Google Scholar] [CrossRef] [PubMed]
- Milovanovic, M.; Drozdenko, G.; Weise, C.; Babina, M.; Worm, M. Interleukin-17A promotes IgE production in human B cells. J. Invest. Dermatol. 2010, 130, 2621–2628. [Google Scholar] [CrossRef] [PubMed]
- Sparber, F.; De Gregorio, C.; Steckholzer, S.; Ferreira, F.M.; Dolowschiak, T.; Ruchti, F.; Kirchner, F.R.; Mertens, S.; Prinz, I.; Joller, N.; et al. The skin commensal yeast Malassezia triggers a type 17 response that coordinates anti-fungal immunity and exacerbates skin inflammation. Cell. Host. Microbe. 2019, 25, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, H.; Wang, X.; Lu, M.; Tan, X.; Peng, L.; Tan, F.; Xiao, T.; Xiao, S.; Xia, Y. Experimental atopic dermatitis is dependent on the TWEAK/Fn14 signaling pathway. Clin. Exp. Immunol. 2020, 199, 56–67. [Google Scholar] [CrossRef]
- Kim, M.H.; Jin, S.P.; Jang, S.; Choi, J.Y.; Chung, D.H.; Lee, D.H.; Kim, K.H.; Kim, H.Y. IL-17A-producing innate lymphoid cells promote skin inflammation by inducing IL-33-driven type 2 immune responses. J. Investg. Dermatol. 2019, in press. [Google Scholar] [CrossRef]
- Shen, H.H.; Fan, Y.; Wang, Y.N.; Zhao, C.N.; Zhang, Z.K.; Pan, H.F.; Wu, G.C. Elevated Circulating Interleukin-17 levels in patients with systemic lupus erythematosus: A meta-analysis. Immunol. Invest. 2019, in press. [Google Scholar] [CrossRef]
- Nakashima, T.; Jinnin, M.; Yamane, K.; Honda, N.; Kajihara, I.; Makino, T.; Masuguchi, S.; Fukushima, S.; Okamoto, Y.; Hasegawa, M.; et al. Impaired IL-17 signaling pathway contributes to the increased collagen expression in scleroderma fibroblasts. J. Immunol. 2012, 188, 3573–3583. [Google Scholar] [CrossRef]
- Ahmed, S.; Misra, D.P.; Agarwal, V. Interleukin-17 pathways in systemic sclerosis-associated fibrosis. Rheumatol. Int. 2019, 39, 1135–1143. [Google Scholar] [CrossRef]
- Miyagaki, T.; Sugaya, M.; Suga, H.; Kamata, M.; Ohmatsu, H.; Fujita, H.; Asano, Y.; Tada, Y.; Kadono, T.; Sato, S. IL-22, but not IL-17, dominant environment in cutaneous T-cell lymphoma. Clin. Cancer Res. 2011, 17, 7529–7538. [Google Scholar] [CrossRef]
- Fernández-Antón Martínez, M.C.; Alfageme Roldán, F.; Ciudad Blanco, C.; Suárez Fernández, R. Ustekinumab in the treatment of severe atopic dermatitis: A preliminary report of our experience with 4 patients. Actas Dermosifiliogr. 2014, 105, 312–313. [Google Scholar] [CrossRef]
- Agusti-Mejias, A.; Messeguer, F.; García, R.; Febrer, I. Severe refractory atopic dermatitis in an adolescent patient successfully treated with ustekinumab. Ann. Dermatol. 2013, 25, 368–370. [Google Scholar] [CrossRef] [PubMed]
- Lis-Święty, A.; Skrzypek-Salamon, A.; Arasiewicz, H.; Brzezińska-Wcisło, L. Atopic dermatitis exacerbated with ustekinumab in a psoriatic patient with childhood history of atopy. Allergol. Int. 2015, 64, 382–383. [Google Scholar] [CrossRef] [PubMed]
- Samorano, L.P.; Hanifin, J.M.; Simpson, E.L.; Leshem, Y.A. Inadequate response to ustekinumab in atopic dermatitis - a report of two patients. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 522–523. [Google Scholar] [CrossRef] [PubMed]
- Ishiuji, Y.; Umezawa, Y.; Asahina, A.; Fukuta, H.; Aizawa, N.; Yanaba, K.; Nakagawa, H. Exacerbation of atopic dermatitis symptoms by ustekinumab in psoriatic patients with elevated serum immunoglobulin E levels: Report of two cases. J. Dermatol. 2018, 45, 732–734. [Google Scholar] [CrossRef] [PubMed]
- Saeki, H.; Kabashima, K.; Tokura, Y.; Murata, Y.; Shiraishi, A.; Tamamura, R.; Randazzo, B.; Imanaka, K. Efficacy and safety of ustekinumab in Japanese patients with severe atopic dermatitis: A randomized, double-blind, placebo-controlled, phase II study. Br. J. Dermatol. 2017, 177, 419–427. [Google Scholar] [CrossRef]
- Khattri, S.; Brunner, P.M.; Garcet, S.; Finney, R.; Cohen, S.R.; Oliva, M.; Dutt, R.; Fuentes-Duculan, J.; Zheng, X.; Li, X.; et al. Efficacy and safety of ustekinumab treatment in adults with moderate-to-severe atopic dermatitis. Exp. Dermatol. 2017, 26, 28–35. [Google Scholar] [CrossRef]
- Munera-Campos, M.; Ballesca, F.; Richarz, N.; Ferrandiz, C.; Carrascosa, J.M. Paradoxical eczematous reaction to ixekizumab. J. Eur. Acad. Dermatol. Venereol. 2019, 33, e40–e42. [Google Scholar]
- Burlando, M.; Cozzani, E.; Russo, R.; Parodi, A. Atopic-like dermatitis after secukinumab injection: A case report. Dermatol. Ther. 2019, 32, e12751. [Google Scholar] [CrossRef]
- Napolitano, M.; Gallo, L.; Patruno, C.; Fabbrocini, G.; Megna, M. Eczematous reaction to ixekizumab successfully treated with dupilumab. Dermatol. Ther. 2020, in press. [Google Scholar] [CrossRef]
- Guttman-Yassky, E.; Brunner, P.M.; Neumann, A.U.; Khattri, S.; Pavel, A.B.; Malik, K.; Singer, G.K.; Baum, D.; Gilleaudeau, P.; Sullivan-Whalen, M.; et al. Efficacy and safety of fezakinumab (an IL-22 monoclonal antibody) in adults with moderate-to-severe atopic dermatitis inadequately controlled by conventional treatments: A randomized, double-blind, phase 2a trial. J. Am. Acad. Dermatol. 2018, 78, 872–881. [Google Scholar] [CrossRef]
- Brunner, P.M.; Pavel, A.B.; Khattri, S.; Leonard, A.; Malik, K.; Rose, S.; Jim On, S.; Vekaria, A.S.; Traidl-Hoffmann, C.; Singer, G.K.; et al. Baseline IL-22 expression in patients with atopic dermatitis stratifies tissue responses to fezakinumab. J. Allergy. Clin. Immunol. 2019, 143, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Ulzii, D.; Vu, Y.H.; Tsuji, G.; Kido-Nakahara, M.; Nakahara, T. Pathogenesis of atopic dermatitis: CurrentpParadigm. Iran. J. Immunol. 2019, 16, 97–107. [Google Scholar] [PubMed]
- Kakinuma, T.; Nakamura, K.; Wakugawa, M.; Mitsui, H.; Tada, Y.; Saeki, H.; Torii, H.; Asahina, A.; Onai, N.; Matsushima, K.; et al. Thymus and activation-regulated chemokine in atopic dermatitis: Serum thymus and activation-regulated chemokine level is closely related with disease activity. J. Allergy. Clin. Immunol. 2001, 107, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Soumelis, V.; Reche, P.A.; Kanzler, H.; Yuan, W.; Edward, G.; Homey, B.; Gilliet, M.; Ho, S.; Antonenko, S.; Lauerma, A.; et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat. Immunol. 2002, 3, 673–680. [Google Scholar] [CrossRef]
- Masuoka, M.; Shiraishi, H.; Ohta, S.; Suzuki, S.; Arima, K.; Aoki, S.; Toda, S.; Inagaki, N.; Kurihara, Y.; Hayashida, S.; et al. Periostin promotes chronic allergic inflammation in response to Th2 cytokines. J. Clin. Invest. 2012, 122, 2590–2600. [Google Scholar] [CrossRef]
- Tatsuno, K.; Fujiyama, T.; Yamaguchi, H.; Waki, M.; Tokura, Y. TSLP directly interacts with skin-homing Th2 Cells highly expressing its receptor to enhance IL-4 production in atopic dermatitis. J. Investg. Dermatol. 2015, 135, 3017–3024. [Google Scholar] [CrossRef]
- Takahashi, N.; Sugaya, M.; Suga, H.; Oka, T.; Kawaguchi, M.; Miyagaki, T.; Fujita, H.; Sato, S. Thymic stromal chemokine TSLP acts through Th2 cytokine production to induce cutaneous T-cell lymphoma. Cancer Res. 2016, 76, 6241–6252. [Google Scholar] [CrossRef]
- Blauvelt, A.; de Bruin-Weller, M.; Gooderham, M.; Cather, J.C.; Weisman, J.; Pariser, D.; Simpson, E.L.; Papp, K.A.; Hong, H.C.; Rubel, D.; et al. Long-term management of moderate-to-severe atopic dermatitis with dupilumab and concomitant topical corticosteroids (LIBERTY AD CHRONOS): A 1-year, randomised, double-blinded, placebo-controlled, phase 3 trial. Lancet 2017, 389, 2287–2303. [Google Scholar] [CrossRef]
- Thewes, M.; Stadler, R.; Korge, B.; Mischke, D. Normal psoriatic epidermis expression of hyperproliferation-associated keratins. Arch. Derm. Res. 1991, 283, 465–471. [Google Scholar] [CrossRef]
- Zhang, X.; Yin, M.; Zhang, L.J. Keratin 6, 16 and 17-Critical Barrier Alarmin Molecules in Skin Wounds and Psoriasis. Cells 2019, 8, E807. [Google Scholar] [CrossRef]
- Egberts, F.; Heinrich, M.; Jensen, J.M.; Winoto-Morbach, S.; Pfeiffer, S.; Wickel, M.; Schunck, M.; Steude, J.; Saftig, P.; Proksch, E.; et al. Cathepsin D is involved in the regulation of transglutaminase 1 and epidermal differentiation. J. Cell. Sci. 2004, 117, 2295–2307. [Google Scholar] [CrossRef] [PubMed]
- Brockmann, L.; Giannou, A.D.; Gagliani, N.; Huber, S. Regulation of Th17 cells and associated cytokines in wound healing, tissue regeneration, and carcinogenesis. Int. J. Mol. Sci. 2017, 18, E1033. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Cai, G.; Liu, C.; Zhao, J.; Gu, C.; Wu, L.; Hamilton, T.A.; Zhang, C.J.; Ko, J.; Zhu, L.; et al. IL-17R-EGFR axis links wound healing to tumorigenesis in Lrig1+ stem cells. J. Exp. Med. 2019, 216, 195–214. [Google Scholar]
- Song, X.; Dai, D.; He, X.; Zhu, S.; Yao, Y.; Gao, H.; Wang, J.; Qu, F.; Qiu, J.; Wang, H.; et al. Growth factor FGF2 cooperates with interleukin-17 to repair intestinal epithelial damage. Immunity 2015, 43, 488–501. [Google Scholar] [CrossRef] [PubMed]
- Khader, S.A.; Gaffen, S.L.; Kolls, J.K. Th17 cells at the crossroads of innate and adaptive immunity against infectious diseases at the mucosa. Mucosal. Immunol. 2009, 2, 403–411. [Google Scholar]
- Zhang, J.; Qiao, Q.; Liu, M.; He, T.; Shi, J.; Bai, X.; Zhang, Y.; Li, Y.; Cai, W.; Han, S.; et al. IL-17 promotes scar formation by inducing macrophage infiltration. Am. J. Pathol. 2018, 188, 1693–1702. [Google Scholar] [CrossRef] [PubMed]
- Suga, H.; Sugaya, M.; Miyagaki, T.; Ohmatsu, H.; Kawaguchi, M.; Takahashi, N.; Fujita, H.; Asano, Y.; Tada, Y.; Kadono, T.; et al. Skin barrier dysfunction and low antimicrobial peptide expression in cutaneous T-cell lymphoma. Clin. Cancer Res. 2014, 20, 4339–4348. [Google Scholar] [CrossRef]
- Miyagaki, T.; Sugaya, M. Recent advances in atopic dermatitis and psoriasis: Genetic background, barrier function, and therapeutic targets. J. Dermatol. Sci. 2015, 78, 89–94. [Google Scholar] [CrossRef]
- Prince, H.M.; Kim, Y.H.; Horwitz, S.M.; Dummer, R.; Scarisbrick, J.; Quaglino, P.; Zinzani, P.L.; Wolter, P.; Sanches, J.A.; Ortiz-Romero, P.L.; et al. ALCANZA study group. Brentuximab vedotin or physician’s choice in CD30-positive cutaneous T-cell lymphoma (ALCANZA): An international, open-label, randomised, phase 3, multicentre trial. Lancet 2017, 390, 555–566. [Google Scholar] [CrossRef]
- Kim, Y.H.; Bagot, M.; Pinter-Brown, L.; Rook, A.H.; Porcu, P.; Horwitz, S.M.; Whittaker, S.; Tokura, Y.; Vermeer, M.; Zinzani, P.L.; et al. MAVORIC Investigators. Mogamulizumab versus vorinostat in previously treated cutaneous T-cell lymphoma (MAVORIC): An international, open-label, randomised, controlled phase 3 trial. Lancet Oncol. 2018, 19, 1192–1204. [Google Scholar] [CrossRef]


{kind=link}
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugaya, M. The Role of Th17-Related Cytokines in Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 1314. https://doi.org/10.3390/ijms21041314
Sugaya M. The Role of Th17-Related Cytokines in Atopic Dermatitis. International Journal of Molecular Sciences. 2020; 21(4):1314. https://doi.org/10.3390/ijms21041314
Chicago/Turabian StyleSugaya, Makoto. 2020. "The Role of Th17-Related Cytokines in Atopic Dermatitis" International Journal of Molecular Sciences 21, no. 4: 1314. https://doi.org/10.3390/ijms21041314
APA StyleSugaya, M. (2020). The Role of Th17-Related Cytokines in Atopic Dermatitis. International Journal of Molecular Sciences, 21(4), 1314. https://doi.org/10.3390/ijms21041314