Carboxy-Terminal Cementum Protein 1-Derived Peptide 4 (cemp1-p4) Promotes Mineralization through wnt/β-catenin Signaling in Human Oral Mucosa Stem Cells

 , ,
, ,
Abstract
:
1. Introduction
2. Results
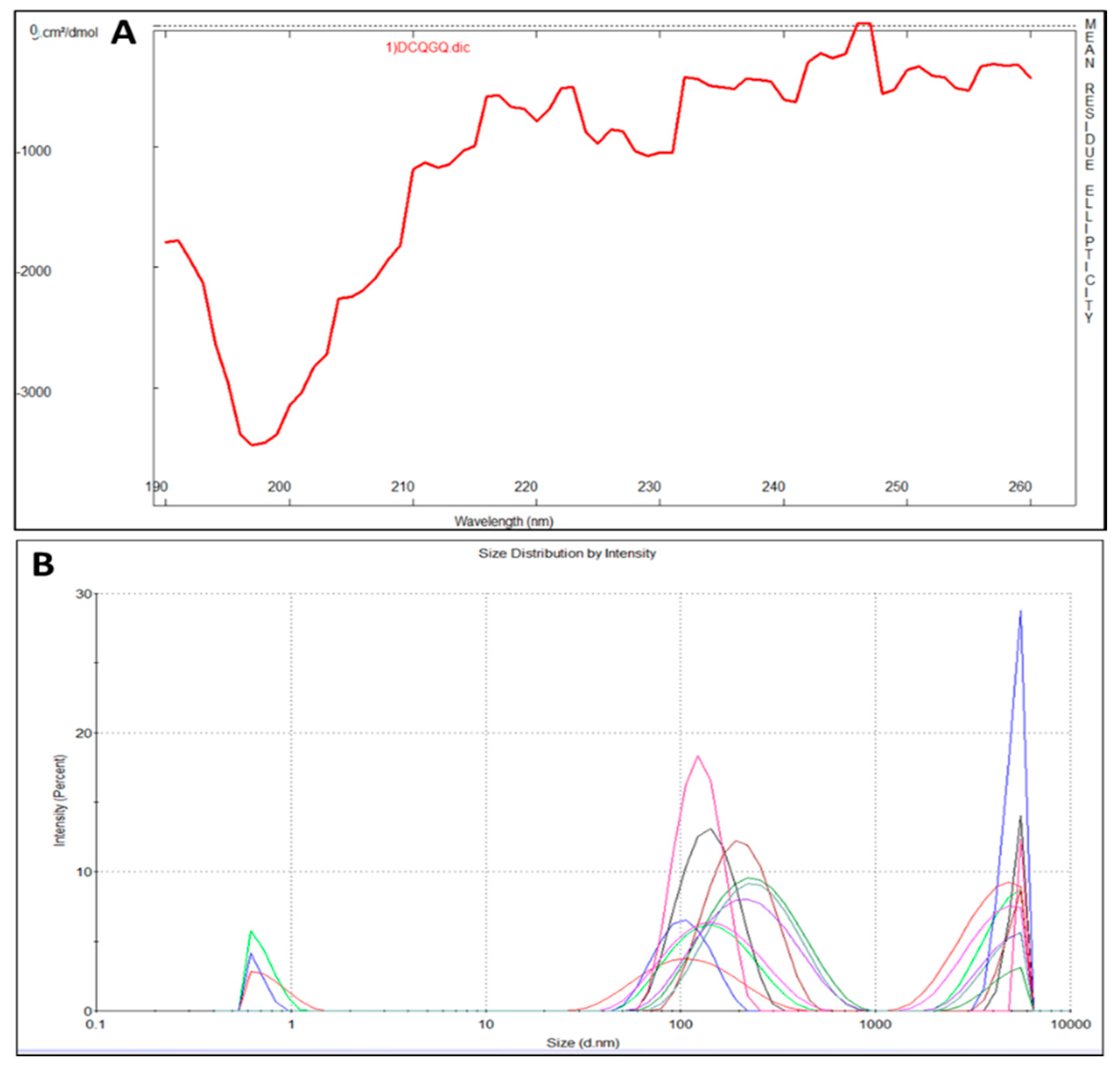
2.1. CEMP1-p4 Physicochemical Characteristics
2.2. CEMP1-p4 Induces Mineralization in a Cell-Free System
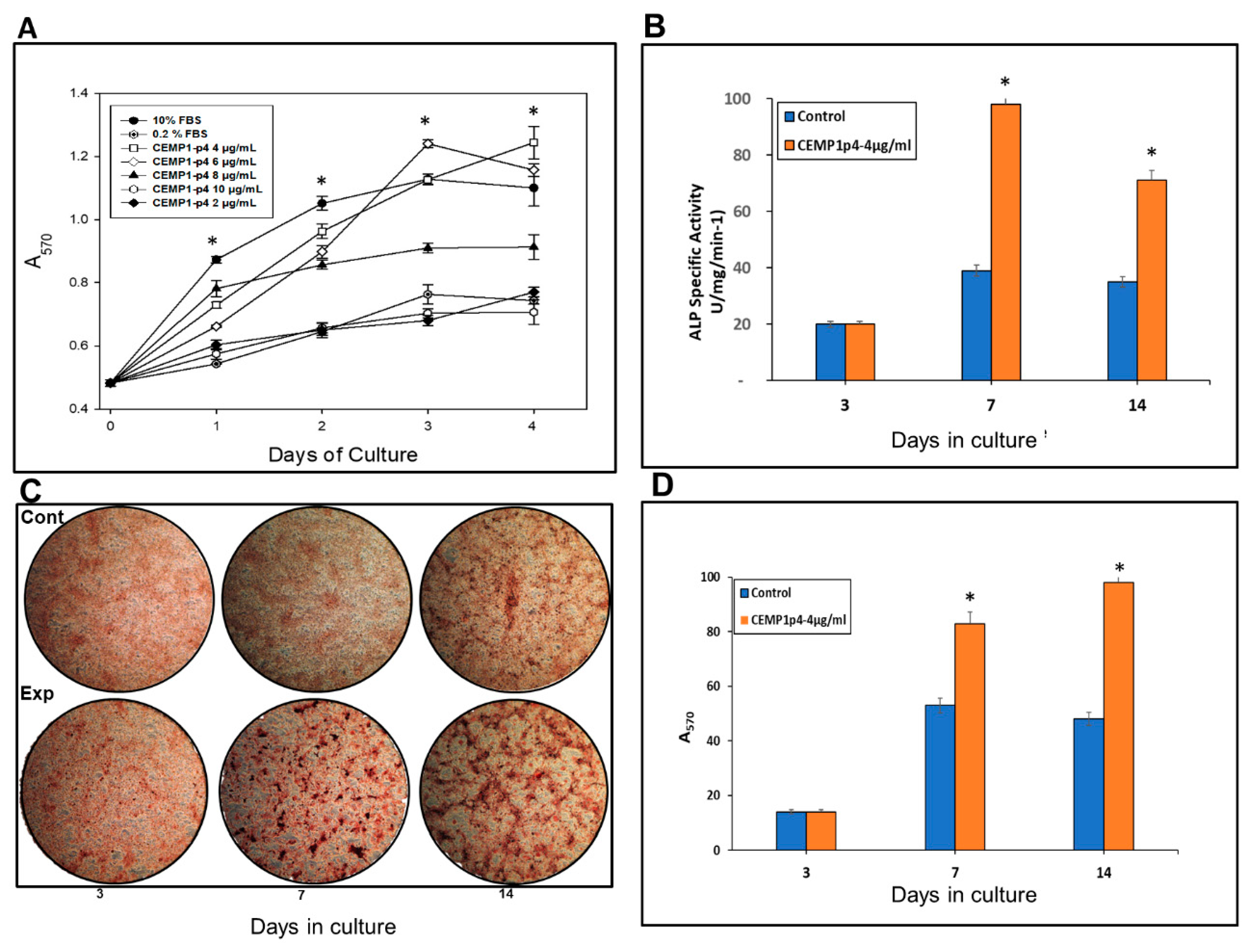
2.3. CEMP1-p4 Promotes Cell Proliferation
2.4. CEMP1-p4 Stimulated ALP Specific Activity by HOMSCs
2.5. CEMP1-p4 induced HOMSCS’s Mineralization
2.6. Effect of CEMP1-p4 on Cell Signaling Pathway
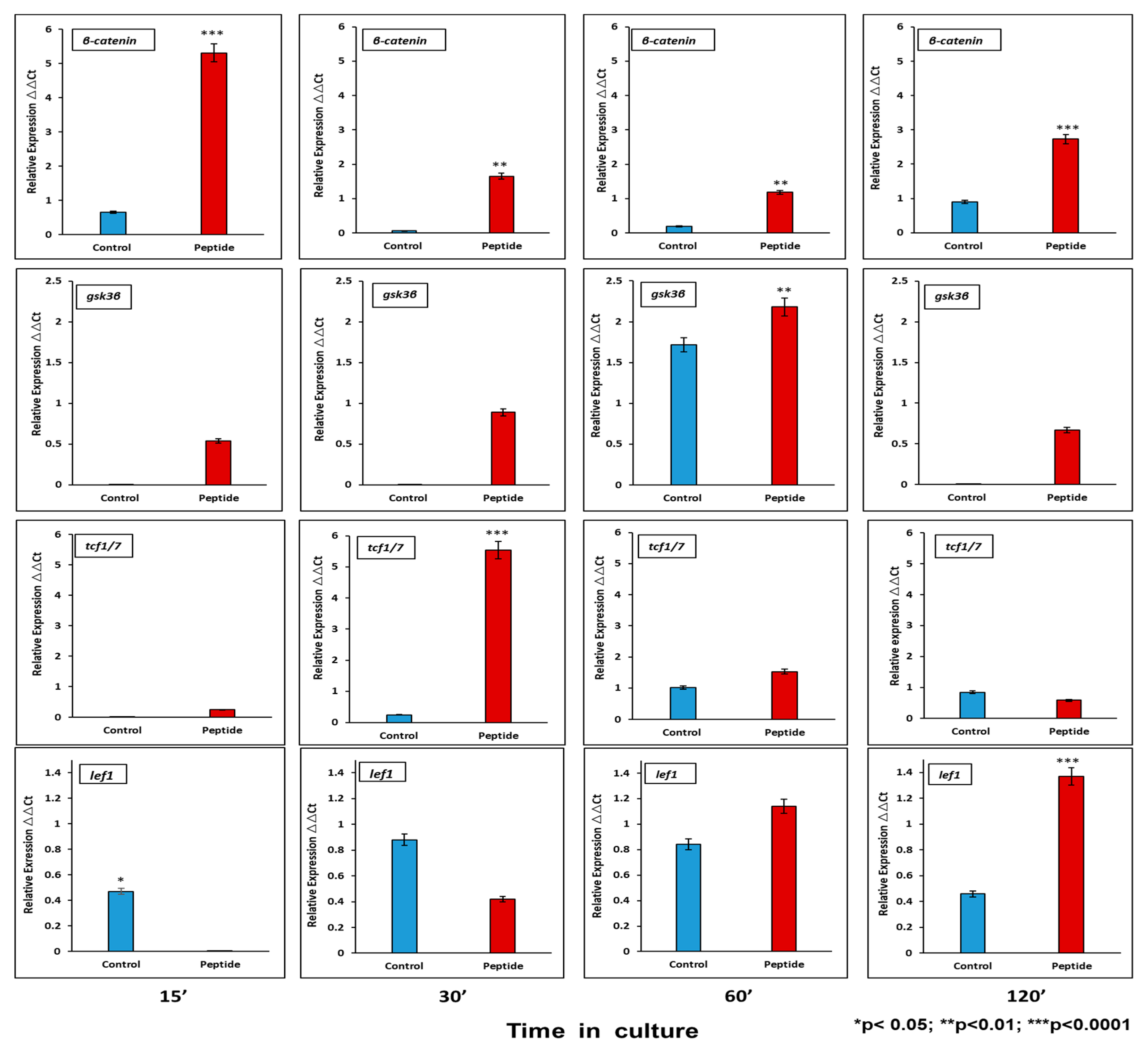
2.6.1. CEMP1-p4 Induced mRNA Transcription of Wnt Signaling Pathway-Related Proteins
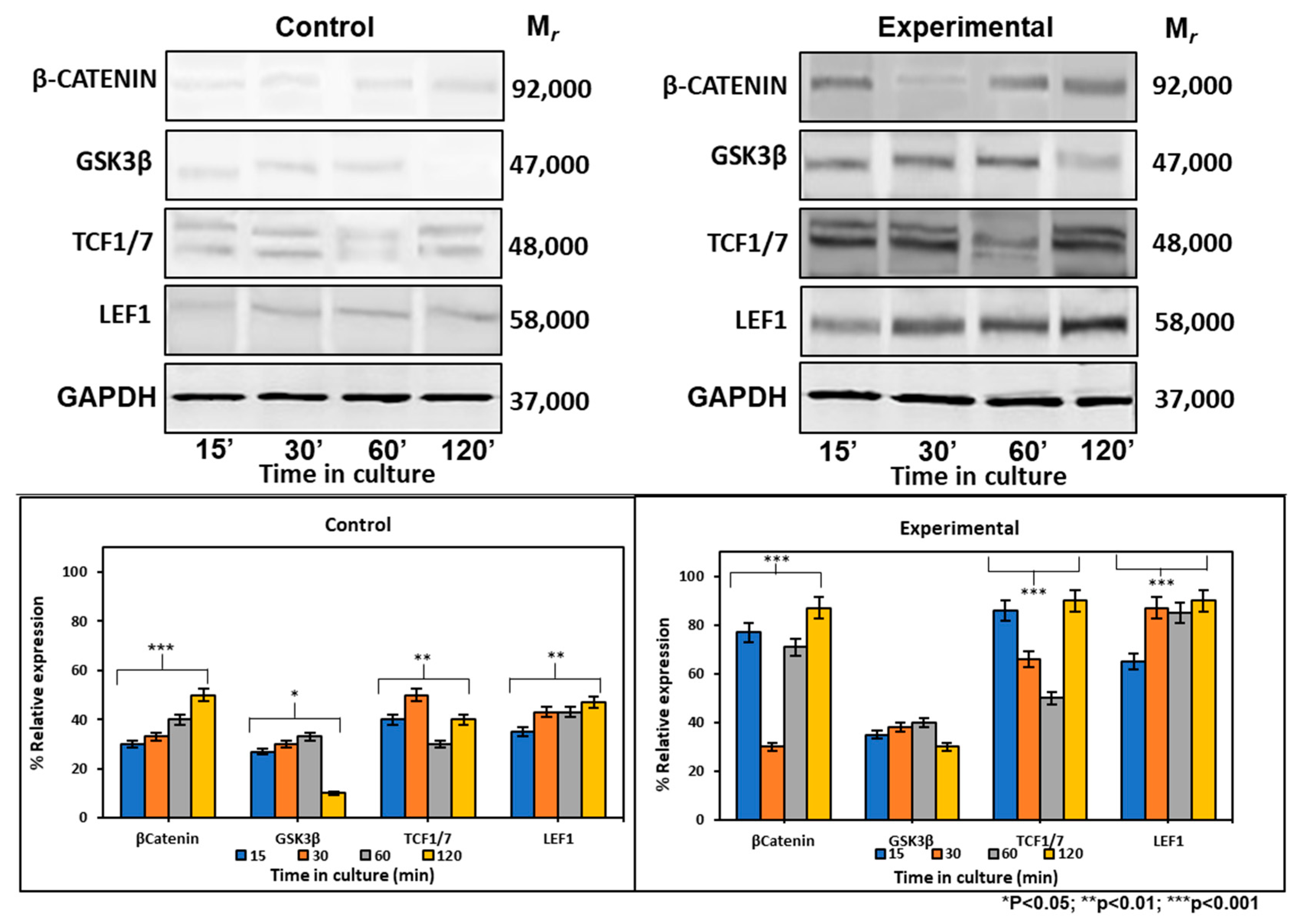
2.6.2. CEMP1-p4 Affected Protein Levels of Wnt Signaling-Related Proteins
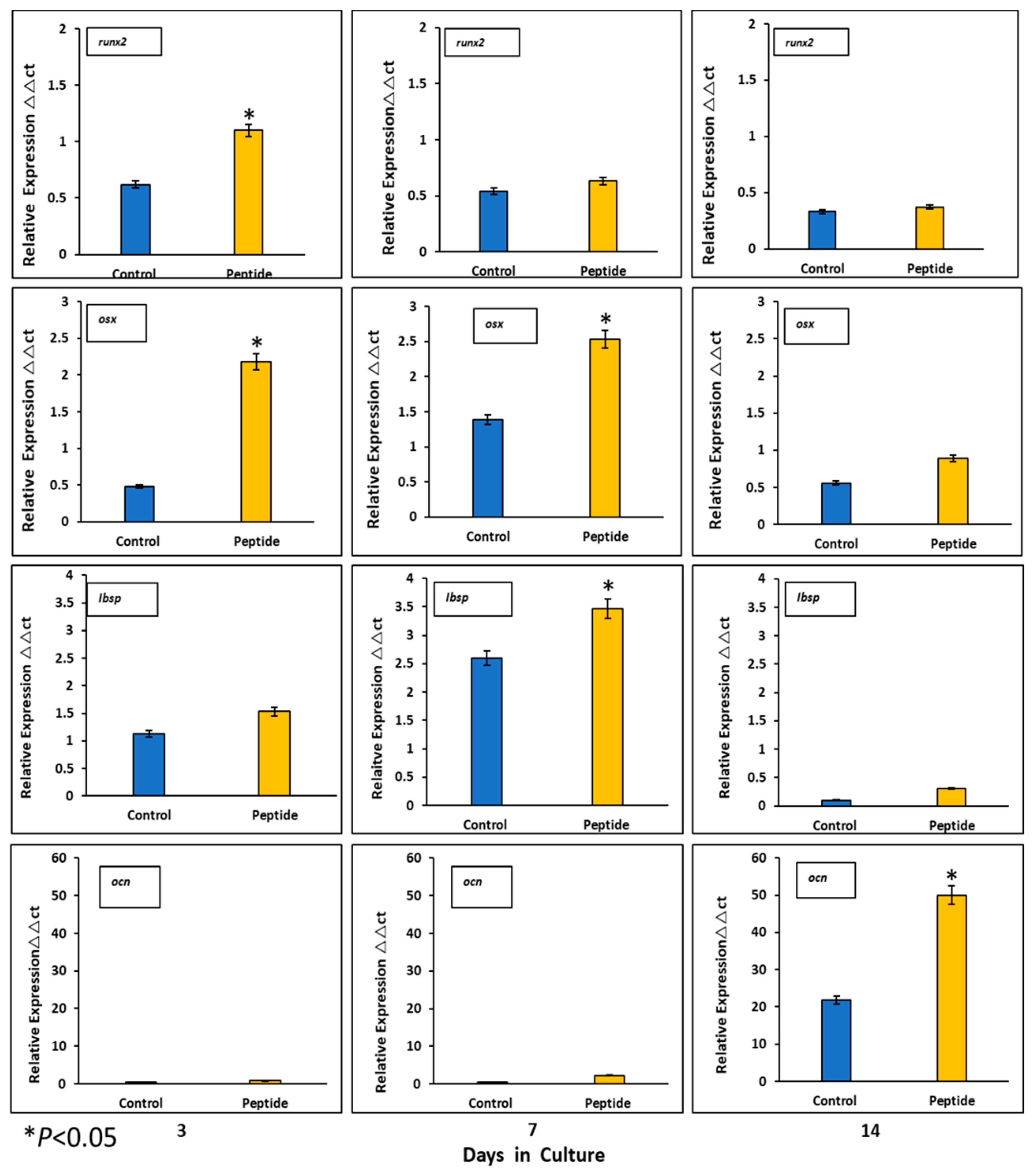
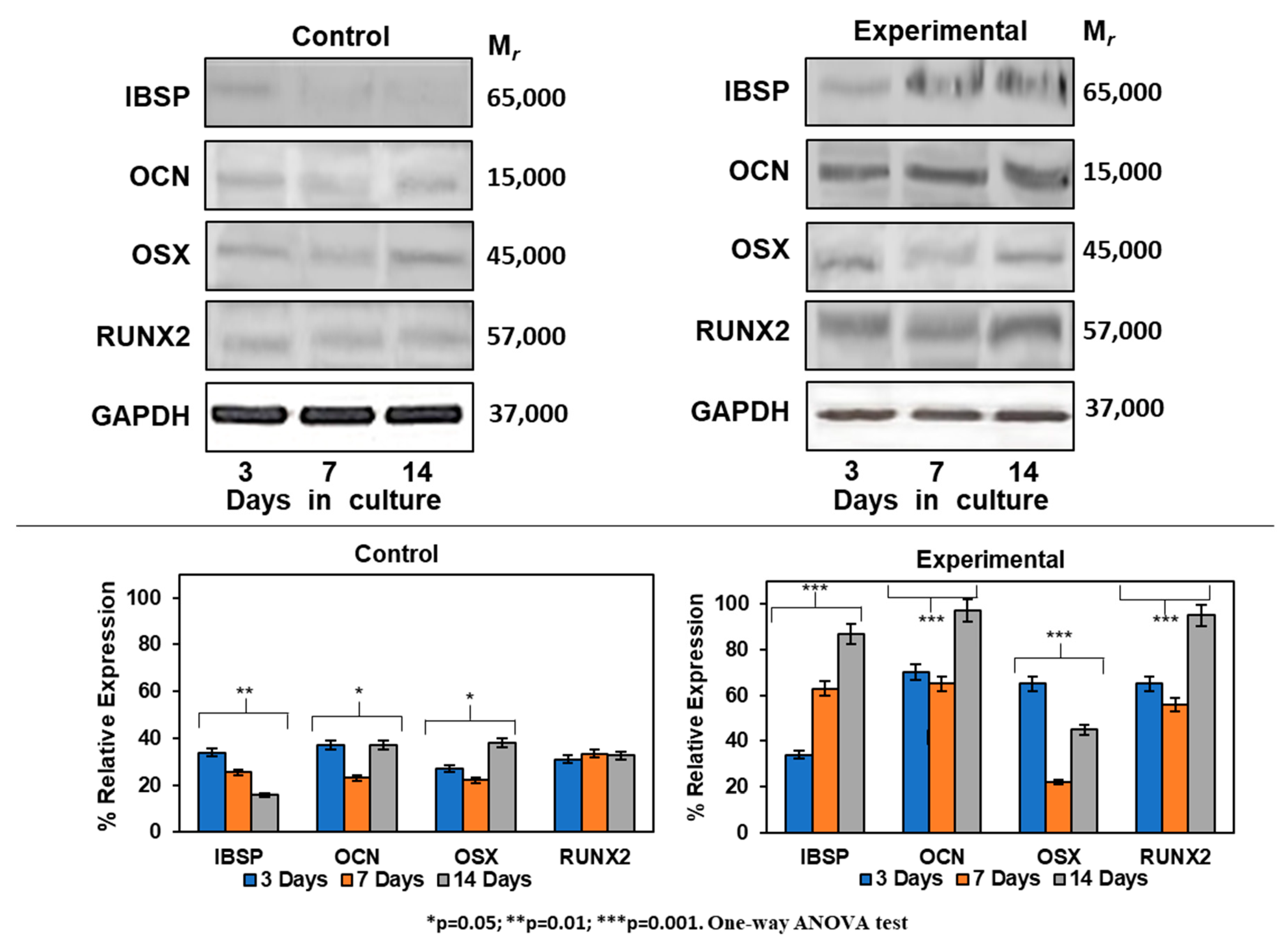
2.7. Expression of Mineralization-Related Markers at the mRNA and Protein Levels
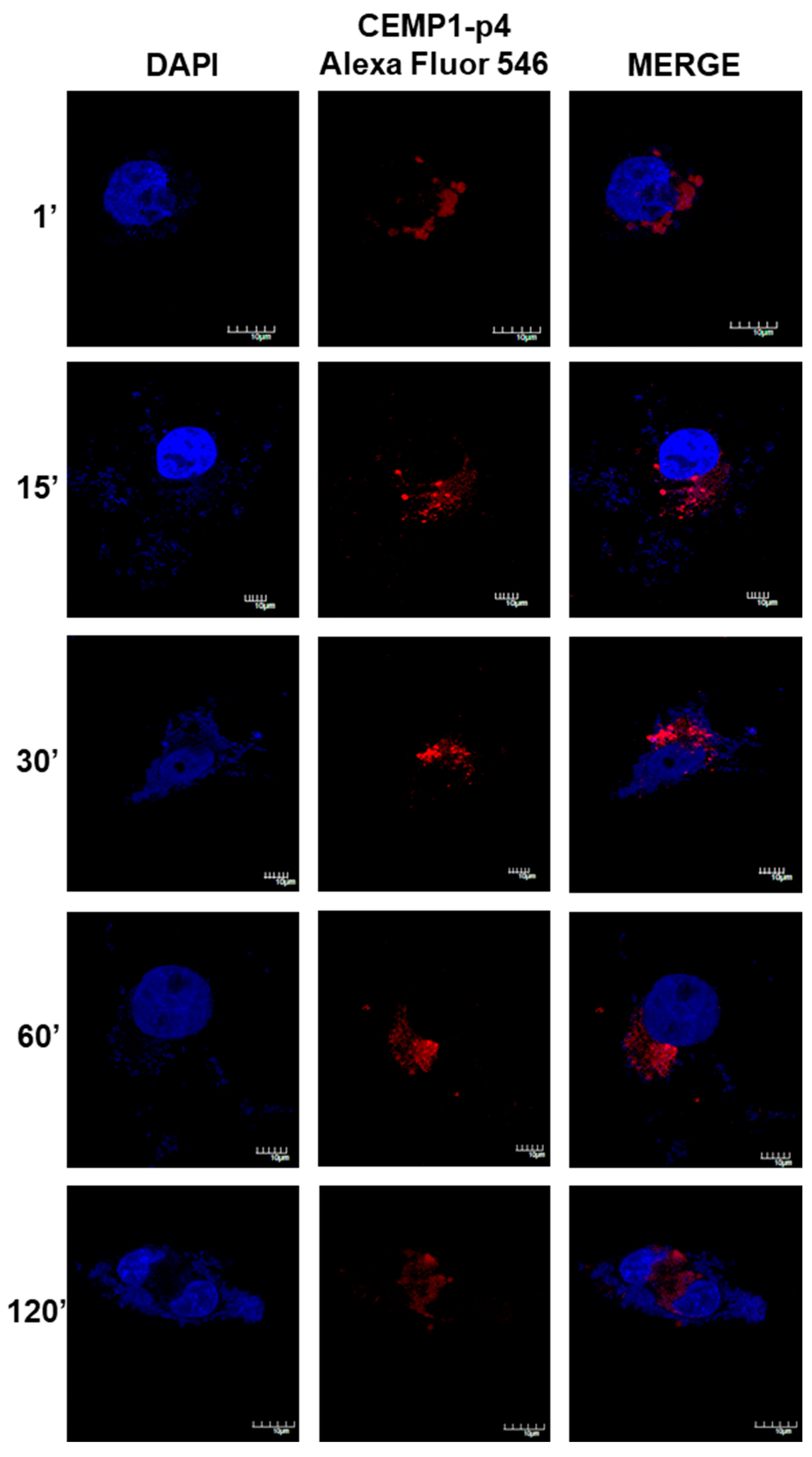
2.8. CEMP1-p4 was Internalized by HOMSCs
2.9. β-Catenin Translocation by CEMP1-p4
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis
4.2. Characterization of CEMP1-p4 Synthetic Peptide
4.3. Circular Dichroism
4.4. Dynamic Light Scattering
4.5. Mineralization in a Cell-Free System
4.6. Cell Culture
4.7. Cell Proliferation Assay
4.8. Time Course Experiments
4.9. HOMSC Mineralization
4.10. Alkaline Phosphatase Activity
4.11. Alizarin Red Staining and Quantification of Mineralization
4.12. Real-Time Quantitative RT-PCR
4.13. Western Blot Analysis
4.14. Confocal Microscopy Analysis
4.15. Immunofluorescence
5. Conclusions
Author Contributions
Acknowledgements
Conflicts of Interest
Abbreviations
| CAP | Cementum Attachment Protein |
| CEMP1 | Cementum Protein 1 |
| CEMP1-p4 | Cementum Protein1-Peptide 4 |
| CD | Circular Dichroism |
| CPC | Cetylpyridinium Chloride |
| CPPs | Cell Penetrating Peptides |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DLS | Dynamic Light Scattering |
| EDS | Energy Dispersive X ray spectroscopy |
| FBS | Fetal Bovine Serum |
| Fmoc | Fluorenylmethyloxycarbonyl |
| GAPDH | Glyceraldehyde-3-Phosphate Dehydrogenase |
| GRAVY | Grand Average of Hydropathicity |
| GSK3β | Glycogen synthase kinase 3 beta |
| HA | Hydroxyapatite |
| HEPES | 2-[4-(2-hydroxyethyl) piperazin-1-yl] ethanesulfonic acid |
| HOMSCs | Human Oral Mucosa Stem Cells |
| IBSP | Integrin Bone Sialoprotein |
| IDP | Intrinsically Disordered Protein |
| LEF1 | Lymphoid Enhancer binding Factor 1 |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| OCN | Osteocalcin |
| OSX | Osterix |
| RUNX2 | Core-Binding Factor Subunit Alpha-1 |
| RPLC | Reverse-Phase Liquid Chromatography |
| SEM | Scanning Electron Microscopy |
| SMS | Sodium Metasilicate |
| TCF | T-cell transcription Factor |
| Wnt | Wingless-related integration site |
References
- Tonetti, M.S.; Jepsen, S.; Jin, L.; Otomo-Corgel, J. Impact of the global burden of periodontal diseases on health, nutrition and wellbeing of mankind: A call for global action. J. Clin. Periodontol. 2017, 44, 456–462. [Google Scholar] [CrossRef] [Green Version]
- Arzate, H.; Zeichner-David, M.; Mercado-Celis, G. Cementum proteins: role in cementogenesis, biomineralization, periodontium formation and regeneration. Periodontol 2000 2015, 67, 211–233. [Google Scholar] [CrossRef]
- King, G.N. New regenerative technologies: Rationale and potential for periodontal regeneration: 1. New advances in established regenerative strategies. Dent. Update 2017, 28, 7–12. [Google Scholar] [CrossRef] [PubMed]
- King, G.N. New regenerative technologies: Rationale and potential for periodontal regeneration: 2. Growth factors. Dent. Update 2017, 28, 60–65. [Google Scholar] [CrossRef] [PubMed]
- King, G.N.; Cochran, D.L. Factors that modulate the effects of bone morphogenetic protein-induced periodontal Cementum proteins 229 regeneration: a critical review. J. Periodontol. 2002, 73, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Treves-Manusevitz, S.; Hoz, L.; Rachima, H.; Montoya, G.; Tzur, E.; Vardimon, A.; Narayanan, A.S.; Amar, S.; Arzate, H.; Pitaru, S. Stem cells of the lamina propria of human oral mucosa and gingiva develop into mineralized tissues in vivo. J. Clin. Periodontol. 2013, 40, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.; Narayanan, S.; Zeichner-David, M.; Carmona, B.; Arzate, H. Molecular cloning, expression and immunolocalization of a novel human cementum-derived protein (CP-23). Bone 2006, 38, 409–419. [Google Scholar] [CrossRef]
- Valdés De Hoyos, A.; Hoz-Rodríguez, L.; Arzate, H.; Narayanan, A.S. Isolation of protein-tyrosine phosphatase-like member-a variant from cementum. J. Dent. Res. 2012, 91, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Hoz, L.; Romo, E.; Zeichner-David, M.; Sanz, M.; Nuñez, J.; Gaitán, L.; Mercado, G.; Arzate, H. Cementum protein 1 (CEMP1) induces differentiation by human periodontal ligament cells under three-dimensional culture conditions. Cell Biol. Int. 2012, 36, 129–136. [Google Scholar] [CrossRef]
- Nuñez, J.; Sanz-Blasco, S.; Vignoletti, F.; Muñoz, F.; Arzate, H.; Villalobos, C.; Nuñez, L.; Caffesse, R.G.; Sanz, M. Periodontal regeneration following implantation of cementum and periodontal ligament-derived cells. J. Periodontal Res. 2012, 47, 33–44. [Google Scholar] [CrossRef]
- Komaki, M.; Iwasaki, K.; Arzate, H.; Narayanan, A.S.; Izumi, Y.; Morita, I. Cementum protein 1 (CEMP1) induces a cementoblastic phenotype and reduces osteoblastic differentiation in periodontal ligament cells. J. Cell Physiol. 2012, 227, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Correa, R.; Arenas, J.; Montoya, G.; Hoz, L.; López, S.; Salgado, F.; Arroyo, R.; Salmeron, N.; Romo, E.; Zeichner-David, M.; et al. Synthetic cementum protein 1-derived peptide regulates mineralization in vitro and promotes bone regeneration in vivo. FASEB J. 2019, 33, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.D.; Nusse, R. Wnt signaling: Multiple pathways, multiple receptors, and multiple transcription factors. J. Biol. Chem. 2006, 281, 22429–22433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, R.T.; Kohn, A.D.; De Ferrari, G.V.; Kaykas, A. WNT and beta-catenin signalling: Diseases and therapies. Nat. Rev. Genet. 2004, 5, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Stadeli, R.; Hoffmans, R.; Basler, K. Transcription under the control of nuclear Arm/beta-catenin. Curr. Biol. 2006, 16, R378–R385. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Srivastava, A.S.; Mishra, R.; Carrie, E.A. Regulatory role of Wnt signaling pathway in the hematopoietic differentiation of murine embryonic stem cells. Biochem. Biophys. Res. Commun. 2004, 324, 1333–1339. [Google Scholar] [CrossRef]
- Sato, N.; Meijer, L.; Skaltsounis, L.; Greengard, P.; Brivanlou, A.H. Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nat. Med. 2004, 10, 55–63. [Google Scholar] [CrossRef]
- Liu, Z.; Habener, J.F. Glucagon-like Peptide-1 Activation of TCF7L2-dependent Wnt Signaling Enhances Pancreatic Beta Cell Proliferation. J Biol Chem. 2008, 283, 8723–8735. [Google Scholar] [CrossRef] [Green Version]
- Willert, K.; Jones, K.A. Wnt signaling: Is the party in the nucleus? Genes Dev. 2006, 20, 1394–1404. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.S. “Tuning in” to mollusk shell nacre- and prismaticassociated protein terminal sequences. Implications for biomineralization and the construction of high-performance inorganic-organic composites. Chem Rev. 2008, 108, 4455–4462. [Google Scholar] [CrossRef]
- Nudelman, F.; Chen, H.H.; Goldberg, H.A.; Weiner, S.; Addadi, L. Spiers Memorial Lecture Lessons from biomineralization: comparing the growth strategies of mollusc shell prismatic and nacreous layers in Atrina rigida. Faraday Discuss. 2007, 136, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of osteoblast differentiation by transcription factors. J. Cell Biochem. 2006, 99, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Jeong, W.S.; Yang, S.J.; Park, E.J.; Oh, T.S.; Koh, K.S. Appropriate and effective dosage of BMP-2 for the ideal regeneration of calvarial bone defects in beagles. Plast Reconstr. Surg. 2016, 138, 64e–72e. [Google Scholar] [CrossRef] [PubMed]
- Montoya, G.; Correa, R.; Arenas, J.; Hoz, L.; Romo, E.; Arroyo, R.; Zeichner-David, M.; Arzate, H. Cementum protein 1-derived peptide (CEMP 1-p1) modulates hydroxyapatite crystal formation in vitro. J. Pept. Sci. 2019, 25, e3211. [Google Scholar] [CrossRef]
- Carstanjen, B.; Desbois, C.; Hekmati, M.; Behr, L. Successful engraftment of cultured autologous mesenchymal stem cells in a surgically repaired soft palate defect in an adult horse. Can. J. Vet. Res. 2006, 70, 143–147. [Google Scholar]
- Bergman, R.J.; Gazit, D.; Kahn, A.J.; Gruber, H.; McDougall, S.; Hahn, T.J. Age-related changes in osteogenic stem cells in mice. J. Bone Miner. Res. 1996, 11, 568–577. [Google Scholar] [CrossRef]
- Grigoriadis, A.E.; Heersche, J.N.; Aubin, J.E. Differentiation of muscle, fat, cartilage, and bone from progenitor cells present in a bone-derived clonal cell population: effect of dexamethasone. J. Cell Biol. 1988, 106, 2139–2151. [Google Scholar] [CrossRef]
- Johnstone, B.; Hering, T.M.; Caplan, A.I.; Goldberg, V.M.; Yoo, J.U. In vitro chondrogenesis of bone marrow-derived mesenchymal progenitor cells. Exp. Cell Res. 1998, 238, 265–272. [Google Scholar] [CrossRef]
- Kadiyala, S.; Young, R.G.; Thiede, M.A.; Bruder, S.P. Culture expanded canine mesenchymal stem cells possess osteochondrogenic potential in vivo and in vitro. Cell Transplant. 1997, 6, 125–134. [Google Scholar] [CrossRef]
- Zhang, H.; Li, H. Tricin enhances osteoblastogenesis through the regulation of Wnt/β-catenin signaling in human mesenchymal stem cells. Mech Dev. 2018, 152, 38–43. [Google Scholar] [CrossRef]
- De Boer, J.; Wang, H.J.; Van Blitterswijk, C. Effects of Wnt signaling on proliferation and differentiation of human mesenchymal stem cells. Tissue Eng. 2004, 10, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xie, R.L.; Croce, C.M.; Stein, J.L.; Lian, J.B.; van Wijnen, A.J.; Stein, G.S. A program of microRNAs controls osteogenic lineage progression by targeting transcription factor Runx2. Proc. Natl. Acad. Sci. USA. 2011, 108, 9863–9868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Wang, Z.; Hao, Y. Osterix overexpression enhances osteoblast differentiation of muscle satellite cells in vitro. Int. J. Oral Maxillofac. Surg. 2008, 37, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.K.; Shapiro, H.S.; Wrana, J.L.; Reimers, S.; Heersche, J.N.; Sodek, J. Localization of bone sialoprotein (BSP) expression to sites of mineralized tissue formation in fetal rat tissues by in situ hybridization. Matrix 1991, 11, 133–143. [Google Scholar] [CrossRef]
- Hunter, G.K.; Goldberg, H.A. Nucleation of hydroxyapatite by bone sialoprotein. Proc. Natl. Acad. Sci. USA. 1993, 90, 8562–8565. [Google Scholar] [CrossRef] [Green Version]
- Hunter, G.K.; Hauschka, P.V.; Pooole, R.A.; Rosenberg, L.C.; Golderg, H.A. Nucleation and inhibition of Hydroxyapatite formation by mineralized tissue proteins. Biochem. J. 1996, 317, 59–64. [Google Scholar] [CrossRef]
- Hough, T.A.; Polewski, M.; Johnson, K.; Cheeseman, M.; Nolan, P.M.; Vizor, L.; Rastan, S.; Boyde, A.; Pritzker, K.; Hunter, A.J.; et al. Novel mouse model of autosomal semi dominant adult hypophosphatasia has a splice site mutation in the tissue nonspecific alkaline phosphatase gene Akp2. J. Bone Miner. Res. 2007, 22, 1397–1407. [Google Scholar] [CrossRef]
- Thomas, G.P.; Baker, S.U.; Eisman, J.A.; Gardiner, E.M. Changing RANKL/OPG mRNA expression in differentiating murine primary osteoblasts. J. Endocrinol. 2002, 170, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Tang, N.; Song, W.X.; Luo, J.; Luo, X.; Chen, J.; Sharff, K.A.; Bi, Y.; He, B.C.; Huang, J.Y.; Zhu, G.H.; et al. BMP-9 induced osteogenic differentiation of mesenchymal progenitors requires functional canonical Wnt/beta-catenin signalling. J. Cell Mol. Med. 2009, 13, 2448–2464. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.L.; Sharff, K.A.; Tang, N.; Song, W.X.; Luo, J.; Luo, X.; Chen, X.; Bennett, E.; Reid, R.; Manning, D.; et al. Regulation of osteogenic differentiation during skeletal development. Front. Biosci. 2008, 13, 2001–2021. [Google Scholar] [CrossRef] [Green Version]
- Karsenty, G.; Wagner, E.F. Reaching a genetic and molecular understanding of skeletal development. Dev. Cell 2002, 2, 389–406. [Google Scholar] [CrossRef] [Green Version]
- Molofsky, A.V.; Pardal, R.; Morrison, S.J. Diverse mechanisms regulate stem cell self-renewal. Curr. Opin. Cell Biol. 2004, 16, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wynshaw-Boris, A. The canonical Wnt pathway in early mammalian embryogenesis and stem cell maintenance/ differentiation. Curr. Opin. Genet. Dev. 2004, 14, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Kleber, M.; Sommer, L. Wnt signaling and the regulation of stem cell function. Curr. Opin. Cell Biol. 2004, 16, 681–687. [Google Scholar] [CrossRef]
- Reya, T.; Clevers, H. Wnt signalling in stem cells and cancer. Nature 2005, 434, 843–850. [Google Scholar] [CrossRef]
- Luo, J.; Chen, J.; Deng, Z.L.; Luo, X.; Song, W.X.; Sharff, K.A.; Tang, N.; Haydon, R.C.; Luu, H.H.; He, T.C. Wnt signaling and human diseases: What are the therapeutic implications? Lab. Invest. 2007, 87, 97–103. [Google Scholar] [CrossRef]
- Wang, H.; Charles, P.C.; Wu, Y.; Ren, R.; Pi, X.; Moser, M.; Barshishat-Kupper, M.; Rubin, J.S.; Perou, C.; Bautch, V.; et al. Gene expression profile signatures indicate a role for Wnt signaling in endothelial commitment from embryonic stem cells. Circ. Res. 2006, 98, 1331–1339. [Google Scholar] [CrossRef]
- Paling, N.R.; Wheadon, H.; Bone, H.K.; Welham, M.J. Regulation of embryonic stem cell self-renewal by phosphoinositide 3-kinase-dependent signaling. J. Biol. Chem. 2004, 279, 48063–48070. [Google Scholar] [CrossRef] [Green Version]
- Cadigan, K.M.; Nusse, R. Wnt signaling: A common theme in animal development. Genes Dev. 1997, 11, 3286–3305. [Google Scholar] [CrossRef] [Green Version]
- Glass, D.A.; Karsenty, G. In vivo analysis of Wnt signaling in bone. Endocrinology. 2007, 148, 2630–2634. [Google Scholar] [CrossRef] [Green Version]
- Bergwitz, C.; Wendlandt, T.; Kispert, A.; Brabanta, G. Wnts differentially regulate colony growth and differentiation of chondrogenic rat calvaria cells. Biochim. Biophys Acta. 2001, 1538, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Fischer, L.; Boland, G.; Tuan, R.S. Wnt signaling during BMP-2 stimulation of mesenchymal chondrogenesis. J. Cell Biochem. 2002, 84, 816–831. [Google Scholar] [CrossRef] [PubMed]
- Glass, D.A.; Karsenty, G. Molecular bases of the regulation of bone remodeling by the canonical Wnt signaling pathway. Curr. Top. Dev. Biol. 2006, 73, 43–84. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [Green Version]
- He, T.C.; Sparks, A.B.; Rago, C.; Hermeking, H.; Zawel, L.; da Costa, L.T.; Morin, P.J.; Vogelstein, B.; Kinzler, K.W. Identification of c-MYC as a target of the APC pathway. Science 1998, 281, 1509–1512. [Google Scholar] [CrossRef]
- Tetsu, O.; McCormick, F. Beta-catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature 1999, 398, 422–426. [Google Scholar] [CrossRef]
- He, T.C.; Chan, T.A.; Vogelstein, B.; Kinzler, K.W. PPAR delta is an APC-regulated target of nonsteroidal anti-inflammatory drugs. Cell 1999, 99, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Qu, Z.H.; Zhang, X.L.; Tang, T.T.; Dai, K.R. Promotion of osteogenesis through beta-catenin signaling by desferrioxamine. Biochem. Biophys Res. Commun. 2008, 370, 332–337. [Google Scholar] [CrossRef]
- Bain, G.; Muller, T.; Wang, X.; Papkoff, J. Activated beta-catenin induces osteoblast differentiation of C3H10T1/2 cells and participates in BMP2 mediated signal transduction. Biochem. Biophys. Res. Commun. 2003, 301, 84–91. [Google Scholar] [CrossRef]
- Jackson, A.; Vayssiere, B.; Garcia, T.; Newell, W.; Baron, R.; Roman-Roman, S.; Rawadi, G. Gene array analysis of Wnt-regulated genes in C3H10T1/2 cells. Bone. 2005, 36, 585–598. [Google Scholar] [CrossRef]
- Chang, J.; Sonoyama, W.; Wang, Z.; Jin, Q.; Zhang, C.; Krebsbach, P.H.; Giannobile, W.; Shi, S.; Wang, C.Y. Noncanonical Wnt-4 signaling enhances bone regeneration of mesenchymal stem cells in craniofacial defects through activation of p38 MAPK. J. Biol. Chem. 2007, 282, 30938–30948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaur, T.; Lengner, C.J.; Hovhannisyan, H.; Bhat, R.A.; Bodine, P.V.; Komm, B.S.; Javed, A.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; et al. Canonical WNT signaling promotes osteogenesis by directly stimulating Runx2 gene expression. J. Biol. Chem. 2005, 280, 33132–33140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, T.F.; Guo, X.; Garrett-Beal, L.; Yang, Y. Wnt/beta-catenin signaling in mesenchymal progenitors controls osteoblast and chondrocyte differentiation during vertebrate skeletogenesis. Dev. Cell 2005, 8, 739–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, L.; Rathmer, B.; Ewan, K.; Bange, T.; Heinrichs, S.; Dale, T.C.; Schade, D.; Grossmann, T.N. Cell Permeable Stapled Peptide Inhibitor of Wnt Signaling that Targets β-Catenin Protein-Protein Interactions. Cell Chem. Biol. 2017, 24, 958–968. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, A. Thermodynamic studies and binding mechanisms of cell-penetrating peptides with lipids and glycosaminoglycans. Adv. Drug Deliv. Rev. 2008, 60, 580–594. [Google Scholar] [CrossRef]
- Gallivan, J.P.; Dougherty, D.A. Cation-pi interactions in structural biology. Proc. Natl. Acad. Sci. USA. 1999, 96, 9459–9464. [Google Scholar] [CrossRef] [Green Version]
- Persson, S.; Killian, J.A.; Lindblom, G. Molecular ordering of interfacially localized tryptophan analogs in ester- and ether-lipid bilayers studied by 2H-NMR. Biophys. J. 1998, 75, 1365–1371. [Google Scholar] [CrossRef] [Green Version]
- Qian, Z.; Martyna, A.; Hard, R.L.; Wang, J.; Appiah-Kubi, G.; Coss, C.; Phelps, M.A.; Rossman, J.S.; Pei, D. Discovery and mechanism of highly efficient cyclic cell-penetrating peptides. Biochemistry 2016, 55, 2601–2612. [Google Scholar] [CrossRef]
- Brock, R. The uptake of arginine-rich cell-penetrating peptides: putting the puzzle together. Bioconjug. Chem. 2014, 25, 863–868. [Google Scholar] [CrossRef]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: Components, mechanisms and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [Green Version]
- Peterson-Nedry, W.; Erdeniz, N.; Kremer, S.; Yu, J.; Baig-Lewis, S.; Wehrli, M.; Baig-Lewis, S.; Wehrli, M. Unexpectedly robust assembly of the Axin destruction complex regulates Wnt/Wg signaling in Drosophila as revealed by analysis in vivo. Dev. Biol. 2008, 320, 226–241. [Google Scholar] [CrossRef] [PubMed]
- Salins, P.; Shawesh, S.; He, Y.; Dibrov, A.; Kashour, T.; Arthur, G.; Amara, F. Lovastatin protects human neurons against Abeta-induced toxicity and causes activation of beta-catenin-TCF/LEF signaling. Neurosci. Lett. 2007, 412, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.C. Examination of effects of GSK3beta phosphorylation, beta-catenin phosphorylation and beta-catenin degradation on kinetics of Wnt signaling pathway using computational method. Theor. Biol. Med. Model. 2009, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provencher, S.W.; Glöckner, J. Estimation of globular protein secondary structure from circular dichroism. Biochemistry 1981, 20, 33–37. [Google Scholar] [CrossRef]
- Moreno, A.; Mas-Oliva, J.; Soriano-García, M.; Salvador-Oliver, S.; Bolaños-García, V.M. Turbidity as a useful optical parameter to predict protein crystallization by dynamic light scattering. J. Mol. Struct. 2000, 19, 243–256. [Google Scholar] [CrossRef]
- Lowry, O.H.; Roberts, N.R.; Wu, M.-L.; Hixon, W.S.; Crawford, E.J. The quantitative histochemistry of brain, II: enzyme measurements. J. Biol Chem. 1954, 207, 19–37. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence 5´-3´ | |
|---|---|---|
| Forward | Reverse | |
| β-CATENIN | GGTGCTGACTATCCAGTTG | GGCAGAGTAAAGTATTCACCC |
| GSK3β | GACTTTGGAAGTGCAAAGC | AGGAAATATTGGTTGTCCTAGC |
| LEF1 | GAAGAGGAGGGCGACTTAG | CTTTCCTTCATCAGGGTGTTC |
| TCF1 | CCAACATTCTCAGGTCGC | GAGCAAGCCAGGTGTTC |
| IBSP | AACGAAGAAAGCGAAGCAGAA | TCTGCCTCTGTGCTGTTGGT |
| OCN | GTTGCAGGCTCAATCCATTT | CCATCCTCATACCTGCACCT |
| OSX | GCCAGAAGCTGTGAAACCTC | GCTGCAAGCTCTCCATAACC |
| RUNX2 | ACCCAGAAGGCACAGACAGAAG | AGGAATCGCCCCTAAATCACT |
| GAPDH | CAACGGATTTGGTCGTATTGG | GCAACAATATCCACTTTACCAGAGTTAA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arroyo, R.; López, S.; Romo, E.; Montoya, G.; Hoz, L.; Pedraza, C.; Garfias, Y.; Arzate, H. Carboxy-Terminal Cementum Protein 1-Derived Peptide 4 (cemp1-p4) Promotes Mineralization through wnt/β-catenin Signaling in Human Oral Mucosa Stem Cells. Int. J. Mol. Sci. 2020, 21, 1307. https://doi.org/10.3390/ijms21041307
Arroyo R, López S, Romo E, Montoya G, Hoz L, Pedraza C, Garfias Y, Arzate H. Carboxy-Terminal Cementum Protein 1-Derived Peptide 4 (cemp1-p4) Promotes Mineralization through wnt/β-catenin Signaling in Human Oral Mucosa Stem Cells. International Journal of Molecular Sciences. 2020; 21(4):1307. https://doi.org/10.3390/ijms21041307
Chicago/Turabian StyleArroyo, Rita, Sonia López, Enrique Romo, Gonzalo Montoya, Lía Hoz, Claudia Pedraza, Yonathan Garfias, and Higinio Arzate. 2020. "Carboxy-Terminal Cementum Protein 1-Derived Peptide 4 (cemp1-p4) Promotes Mineralization through wnt/β-catenin Signaling in Human Oral Mucosa Stem Cells" International Journal of Molecular Sciences 21, no. 4: 1307. https://doi.org/10.3390/ijms21041307
APA StyleArroyo, R., López, S., Romo, E., Montoya, G., Hoz, L., Pedraza, C., Garfias, Y., & Arzate, H. (2020). Carboxy-Terminal Cementum Protein 1-Derived Peptide 4 (cemp1-p4) Promotes Mineralization through wnt/β-catenin Signaling in Human Oral Mucosa Stem Cells. International Journal of Molecular Sciences, 21(4), 1307. https://doi.org/10.3390/ijms21041307