Omega-3 Versus Omega-6 Polyunsaturated Fatty Acids in the Prevention and Treatment of Inflammatory Skin Diseases

 ,
,
Abstract
1. Introduction
2. Sources of PUFAs
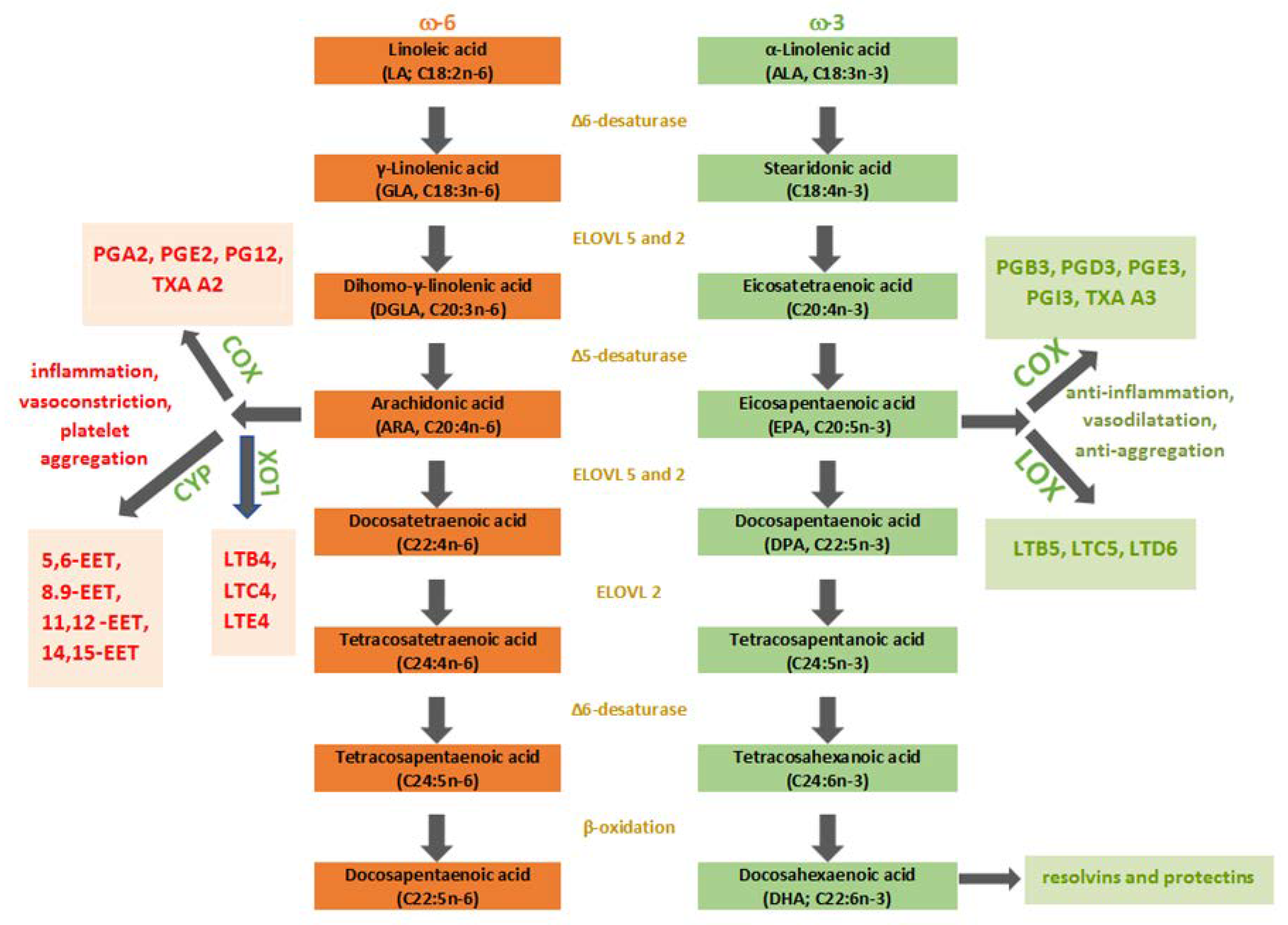
3. Metabolism of PUFAs
4. PUFAs as Gene Expression Regulators
5. Toll-Like Receptors and PUFAs—Modulating Immune Response
6. Skin and PUFAs
7. Atopic Dermatitis and PUFAs
7.1. Supplementation with ω-3PUFAs in Atopic Dermatitis
7.2. Supplementation with ω-6PUFAs in Atopic Dermatitis
8. Psoriasis and PUFAs
9. Acne and PUFAs
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Burr, G.O.; Burr, M.M. A new deficiency disease produced by the rigid exclusion of fat from the diet. J. Biol. Chem. 1929, 82, 345–367. [Google Scholar] [CrossRef] [PubMed]
- Burr, G.O.; Burr, M.M. On the nature and role of the fatty acids essential in nutrition. J. Biol. Chem. 1930, 86, 587–621. [Google Scholar] [CrossRef]
- Kaur, N.; Chugh, V.; Gupta, A.K. Essential fatty acids as functional components of foods—A review. J. Food Sci. Technol. 2014, 51, 2289–2303. [Google Scholar] [CrossRef] [PubMed]
- Sampath, H.; Ntambi, J.M. Polyunsaturated Fatty Acid Regulation of Gene Expression. Nutr. Rev. 2004, 62, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, C.; Chatgilialoglu, C. Lipidomic Profiles and Intervention Strategies in Prevention and Diseases. In Membrane Lipidomics for Personalized Health; Ferreri, C., Chatgilialoglu, C., Eds.; Wiley Online Books: Hoboken, NJ, USA, 2015; pp. 135–155. [Google Scholar]
- Moghadasian, M.H. Advances in dietary enrichment with n-3 fatty acids. Crit. Rev. Food Sci. Nutr. 2008, 48, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Shetty, N.P.; Giridhar, P. GC-FID/MS Analysis of Fatty Acids in Indian Cultivars of Moringa oleifera: Potential Sources of PUFA. J. Am. Oil Chem. Soc. 2014, 91, 1029–1034. [Google Scholar] [CrossRef]
- Saini, R.K.; Shang, X.M.; Ko, E.Y.; Choi, J.H.; Kim, D.; Keum, Y.-S. Characterization of nutritionally important phytoconstituents in minimally processed ready-to-eat baby-leaf vegetables using HPLC–DAD and GC–MS. J. Food Meas. Charact. 2016, 10, 341–349. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006, 60, 502–507. [Google Scholar] [CrossRef]
- Harwood, J.L. Algae: Critical sources of very long-chain polyunsaturated fatty acids. Biomolecules 2019, 9, 708. [Google Scholar] [CrossRef]
- Parmentier, M.; Mahmoud, C.A.S.; Linder, M.; Fanni, J. Polar lipids: N-3 PUFA carriers for membranes and brain: Nutritional interest and emerging processes. Oléagineux Corps gras Lipides 2007, 14, 224–229. [Google Scholar] [CrossRef]
- Hernandez, E.M. 4-Specialty Oils: Functional and Nutraceutical Properties. In Functional Dietary Lipids: Food Formulation, Consumer Issues and Innovation for Health; Sanders, T., Ed.; Woodhead Publishing-Elsevier: Cambridge, UK, 2016; pp. 69–101. [Google Scholar]
- Banca Dati di Composizione degli Alimenti per Studi Epidemiologici in Italia (BDA). Available online: http://www.bda-ieo.it/wordpress/en/?page_id=23 (accessed on 16 January 2020).
- U.S. Department of Agriculture, Agricultural Research Service. FoodData Central. 2019. Available online: https://fdc.nal.usda.gov (accessed on 16 January 2020).
- Ferreri, C.; Masi, A.; Sansone, A.; Giacometti, G.; Larocca, A.; Menounou, G.; Scanferlato, R.; Tortorella, S.; Rota, D.; Conti, M. Fatty acids in membranes as homeostatic, metabolic and nutritional biomarkers: Recent advancements in analytics and diagnostics. Diagnostics 2017, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Abedi, E.; Sahari, M.A. Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food Sci. Nutr. 2014, 2, 443–463. [Google Scholar] [CrossRef] [PubMed]
- Brenna, J.T.; Salem Jr, N.; Sinclair, A.J.; Cunnane, S.C. α-Linolenic acid supplementation and conversion to n-3 long-chain polyunsaturated fatty acids in humans. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 85–91. [Google Scholar] [CrossRef]
- Harnack, K.; Andersen, G.; Somoza, V. Quantitation of alpha-linolenic acid elongation to eicosapentaenoic and docosahexaenoic acid as affected by the ratio of n6/n3 fatty acids. Nutr. Metab. (Lond.) 2009, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.-S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef] [PubMed]
- James, M.J.; Gibson, R.A.; Cleland, L.G. Dietary polyunsaturated fatty acids and inflammatory mediator production 1,2. Am. J. Clin. Nutr. 2000, 71, 343. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Sergeant, S.; Rahbar, E.; Chilton, F.H. Gamma-linolenic acid, dihommo-gamma linolenic, eicosanoids and inflammatory processes. Eur. J. Pharmacol. 2016, 785, 77–86. [Google Scholar] [CrossRef]
- Quinn, P.; Wang, X. Lipids in Health and Disease; Quinn, P.J., Wang, X., Eds.; Springer Science & Business Media, B.V.: Haarlem, The Netherlands, 2008; Volume 49, ISBN 1402088310. [Google Scholar]
- Duplus, E.; Glorian, M.; Forest, C. Fatty acid regulation of gene transcription. J. Biol. Chem. 2000, 275, 30749–30752. [Google Scholar] [CrossRef]
- Deckelbaum, R.J.; Worgall, T.S.; Seo, T. n-3 fatty acids and gene expression. Am. J. Clin. Nutr. 2006, 83, 1520S–1525S. [Google Scholar] [CrossRef]
- Adams, C.M.; Reitz, J.; De Brabander, J.K.; Feramisco, J.D.; Li, L.; Brown, M.S.; Goldstein, J.L. Cholesterol and 25-hydroxycholesterol inhibit activation of SREBPs by different mechanisms, both involving SCAP and Insigs. J. Biol. Chem. 2004, 279, 52772–52780. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.A.; Hamilton, J.A.; Worgall, T.S.; Deckelbaum, R.J. Free fatty acids modulate intermembrane trafficking of cholesterol by increasing lipid mobilities: Novel 13C NMR analyses of free cholesterol partitioning. Biochemistry 2003, 42, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Reddy, J.K. Transcription coactivators for peroxisome proliferator-activated receptors. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2007, 1771, 936–951. [Google Scholar] [CrossRef]
- Matsuura, H.; Adachi, H.; Smart, R.C.; Xu, X.; Arata, J.; Jetten, A.M. Correlation between expression of peroxisome proliferator-activated receptor β and squamous differentiation in epidermal and tracheobronchial epithelial cells. Mol. Cell. Endocrinol. 1999, 147, 85–92. [Google Scholar] [CrossRef]
- Rivier, M.; Safonova, I.; Lebrun, P.; Michel, S.; Griffiths, C.E.M.; Ailhaud, G. Differential expression of peroxisome proliferator-activated receptor subtypes during the differentiation of human keratinocytes. J. Investig. Dermatol. 1998, 111, 1116–1121. [Google Scholar] [CrossRef]
- Westergaard, M.; Henningsen, J.; Kratchmarova, I.; Kristiansen, K.; Svendsen, M.L.; Johansen, C.; Jensen, U.B.; Schrøder, H.D.; Berge, R.K.; Iversen, L. Modulation of keratinocyte gene expression and differentiation by PPAR-selective ligands and tetradecylthioacetic acid. J. Investig. Dermatol. 2001, 116, 702–712. [Google Scholar] [CrossRef]
- Hofmann, C.; Lorenz, K.; Braithwaite, S.S.; Colca, J.R.; Palazuk, B.J.; Hotamisligil, G.S.; Spiegelman, B.M. Altered gene expression for tumor necrosis factor-alpha and its receptors during drug and dietary modulation of insulin resistance. Endocrinology 1994, 134, 264–270. [Google Scholar] [CrossRef]
- Ricote, M.; Li, A.C.; Willson, T.M.; Kelly, C.J.; Glass, C.K. The peroxisome proliferator-activated receptor-γ is a negative regulator of macrophage activation. Nature 1998, 391, 79. [Google Scholar] [CrossRef]
- Jiang, C.; Ting, A.T.; Seed, B. PPAR-γ agonists inhibit production of monocyte inflammatory cytokines. Nature 1998, 391, 82. [Google Scholar] [CrossRef]
- Gosset, P.; Charbonnier, A.; Delerive, P.; Fontaine, J.; Staels, B.; Pestel, J.; Tonnel, A.; Trottein, F. Peroxisome proliferator-activated receptor γ activators affect the maturation of human monocyte-derived dendritic cells. Eur. J. Immunol. 2001, 31, 2857–2865. [Google Scholar] [CrossRef]
- Faveeuw, C.; Fougeray, S.; Angeli, V.; Fontaine, J.; Chinetti, G.; Gosset, P.; Delerive, P.; Maliszewski, C.; Capron, M.; Staels, B. Peroxisome proliferator-activated receptor γ activators inhibit interleukin-12 production in murine dendritic cells. FEBS Lett. 2000, 486, 261–266. [Google Scholar] [CrossRef]
- Martin, H. Role of PPAR-gamma in inflammation. Prospects for therapeutic intervention by food components. Mutat. Res. Mol. Mech. Mutagen. 2010, 690, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Ide, T.; Shimano, H.; Yahagi, N.; Amemiya-Kudo, M.; Matsuzaka, T.; Yatoh, S.; Kitamine, T.; Okazaki, H.; Tamura, Y. Cross-talk between peroxisome proliferator-activated receptor (PPAR) α and liver X receptor (LXR) in nutritional regulation of fatty acid metabolism. I. PPARs suppress sterol regulatory element binding protein-1c promoter through inhibition of LXR signali. Mol. Endocrinol. 2003, 17, 1240–1254. [Google Scholar]
- Cagen, L.M.; Xiong, D.; Wilcox, H.G.; Raghow, R. Insulin activates the rat sterol-regulatory-element-binding protein 1c (SREBP-1c) promoter through the combinatorial actions of SREBP, LXR, Sp-1 and NF-Y cis-acting elements. Biochem. J. 2005, 385, 207–216. [Google Scholar] [CrossRef]
- Ou, J.; Tu, H.; Shan, B.; Luk, A.; DeBose-Boyd, R.A.; Bashmakov, Y.; Goldstein, J.L.; Brown, M.S. Unsaturated fatty acids inhibit transcription of the sterol regulatory element-binding protein-1c (SREBP-1c) gene by antagonizing ligand-dependent activation of the LXR. Proc. Natl. Acad. Sci. USA 2001, 98, 6027–6032. [Google Scholar] [CrossRef]
- Erkkilä, A.T.; Lichtenstein, A.H.; Mozaffarian, D.; Herrington, D.M. Fish intake is associated with a reduced progression of coronary artery atherosclerosis in postmenopausal women with coronary artery disease. Am. J. Clin. Nutr. 2004, 80, 626–632. [Google Scholar] [CrossRef]
- Li, A.C.; Binder, C.J.; Gutierrez, A.; Brown, K.K.; Plotkin, C.R.; Pattison, J.W.; Valledor, A.F.; Davis, R.A.; Willson, T.M.; Witztum, J.L. Differential inhibition of macrophage foam-cell formation and atherosclerosis in mice by PPARα, β/δ, and γ. J. Clin. Investig. 2004, 114, 1564–1576. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K.; Kaisho, T. Toll-like receptors: Critical proteins linking innate and acquired immunity. Nat. Immunol. 2001, 2, 675. [Google Scholar] [CrossRef]
- Kumar, H.; Kawai, T.; Akira, S. Toll-like receptors and innate immunity. Biochem. Biophys. Res. Commun. 2009, 388, 621–625. [Google Scholar] [CrossRef]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-like receptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [CrossRef]
- Barrat, F.J.; Coffman, R.L. Development of TLR inhibitors for the treatment of autoimmune diseases. Immunol. Rev. 2008, 223, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.S. Toll-like receptors in skin. Adv. Dermatol. 2008, 24, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Van der Graaf, C.; Van der Meer, J.W.M.; Kullberg, B.J. Toll-like receptors and the host defense against microbial pathogens: Bringing specificity to the innate-immune system. J. Leukoc. Biol. 2004, 75, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Fatty acids and in fl ammation: The cutting edge between food and pharma. Eur. J. Pharmacol. 2011, 668, S50–S58. [Google Scholar] [CrossRef]
- Mobraten, K.; Haug, T.M.; Kleiveland, C.R.; Lea, T. Omega-3 and omega-6 PUFAs induce the same GPR120-mediated signalling events, but with different kinetics and intensity in Caco-2 cells. Lipids Health Dis. 2013, 12, 101. [Google Scholar] [CrossRef]
- Valins, W.; Amini, S.; Berman, B. The expression of toll-like receptors in dermatological diseases and the therapeutic effect of current and newer topical toll-like receptor modulators. J. Clin. Aesthet. Dermatol. 2010, 3, 20. [Google Scholar]
- McInturff, J.E.; Modlin, R.L.; Kim, J. The role of toll-like receptors in the pathogenesis and treatment of dermatological disease. J. Investig. Dermatol. 2005, 125, 1–8. [Google Scholar] [CrossRef]
- McCusker, M.M.; Grant-Kels, J.M. Healing fats of the skin: The structural and immunologic roles of the ω-6 and ω-3 fatty acids. Clin. Dermatol. 2010, 28, 440–451. [Google Scholar] [CrossRef]
- Wertz, P.W. Biochemistry of Human Stratum Corneum Lipids. In Skin Barrier; Peter, M., Elias, P.M., Feingold, K.R., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 33–42. [Google Scholar]
- Wright, S. Essential fatty acids and the skin. Br. J. Dermatol. 1991, 125, 503–515. [Google Scholar] [CrossRef]
- Akiyama, M. The roles of ABCA12 in epidermal lipid barrier formation and keratinocyte differentiation. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2014, 1841, 435–440. [Google Scholar] [CrossRef]
- Hovnanian, A. Harlequin ichthyosis unmasked: A defect of lipid transport. J. Clin. Investig. 2005, 115, 1708–1710. [Google Scholar] [CrossRef] [PubMed]
- Menon, G.K.; Feingold, K.R.; Moser, A.H.; Brown, B.E.; Elias, P.M. De novo sterologenesis in the skin. II. Regulation by cutaneous barrier requirements. J. Lipid Res. 1985, 26, 418–427. [Google Scholar]
- Grubauer, G.; Feingold, K.R.; Elias, P.M. Relationship of epidermal lipogenesis to cutaneous barrier function. J. Lipid Res. 1987, 28, 746–752. [Google Scholar]
- Bhawracharyya, A.K.; Connor, W.E.; Specror, A.A. Excretion of Sterols from the Skin of Normal and Hypercholesterolemic Humans. J. Clin. Investig. 1972, 51, 2060–2070. [Google Scholar] [CrossRef]
- Nikkari, T.; Schreibman, P.H.; Ahrens, E.H. Isotope kinetics of human skin cholesterol secretion. J. Exp. Med. 1975, 141, 620–634. [Google Scholar] [CrossRef]
- Peng, W.; Novak, N. Pathogenesis of atopic dermatitis. Clin. Exp. Allergy 2015, 45, 566–574. [Google Scholar] [CrossRef]
- David Boothe, W.; Tarbox, J.A.; Tarbox, M.B. Atopic Dermatitis: Pathophysiology. Adv. Exp. Med. Biol. 2017, 1027, 21–37. [Google Scholar]
- Palmer, C.N.A.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.D. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441. [Google Scholar] [CrossRef]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; McLean, W.H.I. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122, 1285–1294. [Google Scholar] [CrossRef]
- Guttman-yassky, E.; Krueger, J.G. ScienceDirect Atopic dermatitis and psoriasis: Two different immune diseases or one spectrum? Curr. Opin. Immunol. 2019, 48, 68–73. [Google Scholar] [CrossRef]
- Ferreri, C.; Angelini, F.; Chatgilialoglu, C.; Dellonte, S.; Moschese, V.; Rossi, P.; Chini, L. Trans fatty acids and atopic eczema/dermatitis syndrome: The relationship with a free radical cis-trans isomerization of membrane lipids. Lipids 2005, 40, 661–667. [Google Scholar] [CrossRef]
- Kew, S.; Mesa, M.D.; Tricon, S.; Buckley, R.; Minihane, A.M.; Yaqoob, P. Effects of oils rich in eicosapentaenoic and docosahexaenoic acids on immune cell composition and function in healthy humans. Am. J. Clin. Nutr. 2004, 79, 674–681. [Google Scholar] [CrossRef]
- Li, X.; Bi, X.; Wang, S.; Zhang, Z.; Li, F.; Zhao, A.Z. Therapeutic potential of ω-3 polyunsaturated fatty acids in human autoimmune diseases. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Denburg, J.A.; Hatfield, H.M.; Cyr, M.M.; Hayes, L.; Holt, P.G.; Sehmi, R.; Dunstan, J.A.; Prescott, S.L. Fish oil supplementation in pregnancy modifies neonatal progenitors at birth in infants at risk of atopy. Pediatr. Res. 2005, 57, 276–281. [Google Scholar] [CrossRef]
- Newburg, D.S.; Walker, W.A. Protection of the neonate by the innate immune system of developing gut and of human milk. Pediatr. Res. 2007, 61, 2–8. [Google Scholar] [CrossRef]
- Gdalevich, M.; Mimouni, D.; David, M.; Mimouni, M. Breast-feeding and the onset of atopic dermatitis in childhood: A systematic review and meta-analysis of prospective studies. J. Am. Acad. Dermatol. 2001, 45, 520–527. [Google Scholar] [CrossRef]
- Hoppu, U.; Rinne, M.; Lampi, A.M.; Isolauri, E. Breast milk fatty acid composition is associated with development of atopic dermatitis in the infant. J. Pediatr. Gastroenterol. Nutr. 2005, 41, 335–338. [Google Scholar] [CrossRef]
- Barcelos, R.C.S.; de Mello-Sampayo, C.; Antoniazzi, C.T.D.; Segat, H.J.; Silva, H.; Veit, J.C.; Piccolo, J.; Emanuelli, T.; Bürger, M.E.; Silva-Lima, B.; et al. Oral supplementation with fish oil reduces dryness and pruritus in the acetone-induced dry skin rat model. J. Dermatol. Sci. 2015, 79, 298–304. [Google Scholar] [CrossRef]
- Fang, Y.P.; Yang, S.H.; Lee, C.H.; Aljuffali, I.A.; Kao, H.C.; Fang, J.Y. What is the discrepancy between drug permeation into/across intact and diseased skins? Atopic dermatitis as a model. Int. J. Pharm. 2016, 497, 277–286. [Google Scholar] [CrossRef]
- Elias, P.M.; Feingold, K.R.; Fartasch, M. The epidermal lamellar body as a multifunctional secretory organelle. In Skin Barrier; Elias, P.M., Feingold, K.R., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 281–292. [Google Scholar]
- Mueller, M.; Lukas, B.; Novak, J.; Simoncini, T.; Genazzani, A.R.; Jungbauer, A. Oregano: A source for peroxisome proliferator-activated receptor γ antagonists. J. Agric. Food Chem. 2008, 56, 11621–11630. [Google Scholar] [CrossRef]
- Zhou, X.R.; Sun, C.H.; Liu, J.R.; Zhao, D. Dietary conjugated linoleic acid increases PPARγ gene expression in adipose tissue of obese rat, and improves insulin resistance. Growth Horm. IGF Res. 2008, 18, 361–368. [Google Scholar] [CrossRef]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2-nonsteroidal antiinflammatory drugs and transcellular processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef]
- Gronert, K. Lipid autacoids in inflammation and injury responses: A matter of privilege. Mol. Interv. 2008, 8, 28–35. [Google Scholar] [CrossRef]
- Serhan, C.N.; Hong, S.; Gronert, K.; Colgan, S.P.; Devchand, P.R.; Mirick, G.; Moussignac, R.L. Resolvins: A family of bioactive products of omega-3 fatty acid transformation circuits initiated by aspirin treatment that counter proinflammation signals. J. Exp. Med. 2002, 196, 1025–1037. [Google Scholar] [CrossRef]
- Ohira, T.; Arita, M.; Omori, K.; Recchiuti, A.; Van Dykeand, T.E.; Serhan, C.N. Resolvin E1 receptor activation signals phosphorylation and phagocytosis. J. Biol. Chem. 2010, 285, 3451–3461. [Google Scholar] [CrossRef]
- Arita, M.; Bianchini, F.; Aliberti, J.; Sher, A.; Chiang, N.; Hong, S.; Yang, R.; Petasis, N.A.; Serhan, C.N. Stereochemical assignment, antiinflammatory properties, and receptor for the omega-3 lipid mediator resolvin E1. J. Exp. Med. 2005, 201, 713–722. [Google Scholar] [CrossRef]
- Hasturk, H.; Kantarci, A.; Ohira, T.; Arita, M.; Ebrahimi, N.; Chiang, N.; Petasis, N.A.; Levy, B.D.; Serhan, C.N.; Van Dyke, T.E. RvE1 protects from local inflammation and osteoclast-mediated bone destruction in periodontitis. FASEB J. 2006, 20, 401–403. [Google Scholar] [CrossRef]
- Ariel, A.; Serhan, C.N. Resolvins and protectins in the termination program of acute inflammation. Trends Immunol. 2007, 28, 176–183. [Google Scholar] [CrossRef]
- Kim, T.-H.; Kim, G.-D.; Jin, Y.-H.; Park, Y.S.; Park, C.-S. Omega-3 fatty acid-derived mediator, Resolvin E1, ameliorates 2, 4-dinitrofluorobenzene-induced atopic dermatitis in NC/Nga mice. Int. Immunopharmacol. 2012, 14, 384–391. [Google Scholar] [CrossRef]
- Nishi, K.; Kanayama, Y.; Kim, I.-H.; Nakata, A.; Nishiwaki, H.; Sugahara, T. Docosahexaenoyl ethanolamide mitigates IgE-mediated allergic reactions by inhibiting mast cell degranulation and regulating allergy-related immune cells. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Huang, T.-H.; Wang, P.-W.; Yang, S.-C.; Chou, W.-L.; Fang, J.-Y. Cosmetic and therapeutic applications of fish oil’s fatty acids on the skin. Mar. Drugs 2018, 16, 256. [Google Scholar] [CrossRef] [PubMed]
- Horrobin, D.F. Essential fatty acid metabolism and its modification in atopic eczema. Am. J. Clin. Nutr. 2000, 71, 367s–372s. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.H.; Hardy, G.; Alany, R.G. Borage oil in the treatment of atopic dermatitis. Nutrition 2010, 26, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Brosche, T.; Platt, D. Effect of borage oil consumption on fatty acid metabolism, transepidermal water loss and skin parameters in elderly people. Arch. Gerontol Geriatr 2000, 30, 139–150. [Google Scholar] [CrossRef]
- Van Gool, C.; Zeegers, M.P.A.; Thijs, C. Oral essential fatty acid supplementation in atopic dermatitis—A meta-analysis of placebo-controlled trials. Br. J. Dermatol. 2004, 150, 728–740. [Google Scholar] [CrossRef]
- Muggli, R. Systemic evening primrose oil improves the biophysical skin parameters of healthy adults. Int. J. Cosmet. Sci. 2005, 27, 243–249. [Google Scholar] [CrossRef]
- Neukam, K.; De Spirt, S.; Stahl, W.; Bejot, M.; Maurette, J.-M.; Tronnier, H.; Heinrich, U. Supplementation of flaxseed oil diminishes skin sensitivity and improves skin barrier function and condition. Skin Pharmacol. Physiol. 2011, 24, 67–74. [Google Scholar] [CrossRef]
- Kawamura, A.; Ooyama, K.; Kojima, K.; Kachi, H.; Abe, T.; Amano, K.; Aoyama, T. Dietary supplementation of gamma-linolenic acid improves skin parameters in subjects with dry skin and mild atopic dermatitis. J. Oleo Sci. 2011, 60, 597–607. [Google Scholar] [CrossRef]
- Mathias, R.A.; Pani, V.; Chilton, F.H. Genetic Variants in the FADS Gene: Implications for Dietary Recommendations for Fatty Acid Intake. Curr. Nutr. Rep. 2014, 3, 139–148. [Google Scholar] [CrossRef]
- Ziboh, V.A.; Miller, C.C.; Cho, Y. Metabolism of polyunsaturated fatty acids by skin epidermal enzymes: Generation of antiinflammatory and antiproliferative metabolites. Am. J. Clin. Nutr. 2000, 71, 361s–366s. [Google Scholar] [CrossRef]
- Barham, J.B.; Edens, M.B.; Fonteh, A.N.; Johnson, M.M.; Easter, L.; Chilton, F.H. Addition of eicosapentaenoic acid to γ-linolenic acid–supplemented diets prevents serum arachidonic acid accumulation in humans. J. Nutr. 2000, 130, 1925–1931. [Google Scholar] [CrossRef] [PubMed]
- Mir, M. Echium oil: A valuable source of n-3 and n-6 fatty acids. OCL 2008, 15, 252–256. [Google Scholar] [CrossRef]
- Strannegård, I.L.; Svennerholm, L.; Strannegård, O. Essential fatty acids in serum lecithin of children with atopic dermatitis and in umbilical cord serum of infants with high or low IgE levels. Int. Arch. Allergy Appl. Immunol. 1987, 82, 422–423. [Google Scholar] [CrossRef] [PubMed]
- Galli, E.; Picardo, M.; Chini, L.; Passi, S.; Moschese, V.; Terminali, O.; Paone, F.; Fraioli, G.; Rossi, P. Analysis of polyunsaturated fatty acids in newborn sera: A screening tool for atopic disease? Br. J. Dermatol. 1994, 130, 752–756. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.; Zelczak, G.; Lentze, M.J. Abnormal fatty acid composition in infants at high risk of atopic disease. J. Pediatr. Gastroenterol. Nutr. 1997, 24, 492. [Google Scholar] [CrossRef]
- Laitinen, K.; Hoppu, U.; Hämäläinen, M.; Linderborg, K.; Moilanen, E.; Isolauri, E. Breast milk fatty acids may link innate and adaptive immune regulation: Analysis of soluble CD14, prostaglandin E2, and fatty acids. Pediatr. Res. 2006, 59, 723–727. [Google Scholar] [CrossRef][Green Version]
- Elias, S.L.; Innis, S.M. Infant plasma trans, n-6, and n-3 fatty acids and conjugated linoleic acids are related to maternal plasma fatty acids, length of gestation, and birth weight and length. Am. J. Clin. Nutr. 2001, 73, 807–814. [Google Scholar] [CrossRef]
- Blaess, M. Derailed Ceramide Metabolism in Atopic Dermatitis (AD): A Causal Starting Point for a Personalized (Basic) Therapy. Int. J. Mol. Sci. 2019, 20, 3967. [Google Scholar] [CrossRef]
- Greb, J.E.; Goldminz, A.M.; Elder, J.T.; Lebwohl, M.G.; Gladman, D.D.; Wu, J.J.; Mehta, N.N.; Finlay, A.Y.; Gottlieb, A.B. Psoriasis. Nat. Rev. Dis. Prim. 2016, 2, 16082. [Google Scholar] [CrossRef]
- Kagami, S.; Rizzo, H.L.; Lee, J.J.; Koguchi, Y.; Blauvelt, A. Circulating Th17, Th22, and Th1 cells are increased in psoriasis. J. Investig. Dermatol. 2010, 130, 1373–1383. [Google Scholar] [CrossRef]
- Qin, S.; Wen, J.; Bai, X.-C.; Chen, T.-Y.; Zheng, R.-C.; Zhou, G.; Ma, J.; Feng, J.-Y.; Zhong, B.-L.; Li, Y.-M. Endogenous n-3 polyunsaturated fatty acids protect against imiquimod-induced psoriasis-like inflammation via the IL-17/IL-23 axis. Mol. Med. Rep. 2014, 9, 2097–2104. [Google Scholar] [CrossRef] [PubMed]
- Gürel, G.; Sabah-Özcan, S. Evaluation of Toll-like receptor expression profile in patients with psoriasis vulgaris. Gene 2019, 702, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Duarte, G.V.; Follador, I.; Cavalheiro, C.M.A.; Silva, T.S.; de Oliveira, M.D. Psoriasis and obesity: Literature review and recommendations for management. An. Bras. Dermatol. 2010, 85, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Puzenat, E.; Bronsard, V.; Prey, S.; Gourraud, P.; Aractingi, S.; Bagot, M.; Cribier, B.; Joly, P.; Jullien, D.; Le Maitre, M. What are the best outcome measures for assessing plaque psoriasis severity? A systematic review of the literature. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 10–16. [Google Scholar] [CrossRef]
- Barrea, L.; Balato, N.; Di Somma, C.; Macchia, P.E.; Napolitano, M.; Savanelli, M.C.; Esposito, K.; Colao, A.; Savastano, S. Nutrition and psoriasis: Is there any association between the severity of the disease and adherence to the Mediterranean diet? J. Transl. Med. 2015, 13, 18. [Google Scholar] [CrossRef]
- Machado-Pinto, J.; Diniz, M.S.; Bavoso, N.C. Psoriasis: New comorbidities. An. Bras. Dermatol. 2016, 91, 8–14. [Google Scholar] [CrossRef]
- Miossec, P.; Kolls, J.K. Targeting IL-17 and TH17 cells in chronic inflammation. Nat. Rev. Drug Discov. 2012, 11, 763. [Google Scholar] [CrossRef]
- Bianchi, E.; Rogge, L. The IL-23/IL-17 pathway in human chronic in fl ammatory diseases—New insight from genetics and targeted therapies. Genes Immun. 2019, 415–425. [Google Scholar] [CrossRef]
- Li, S.S.; Liu, Y.; Li, H.; Wang, L.P.; Xue, L.F.; Yin, G.S.; Wu, X.S. Identification of psoriasis vulgaris biomarkers in human plasma by non-targeted metabolomics based on UPLC-Q-TOF/MS. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3940–3950. [Google Scholar]
- Clark, C.C.T.; Taghizadeh, M.; Nahavandi, M.; Jafarnejad, S. Efficacy of ω-3 supplementation in patients with psoriasis: A meta-analysis of randomized controlled trials. Clin. Rheumatol. 2019, 38, 977–988. [Google Scholar] [CrossRef]
- Voorhees, J.J. Leukotrienes and other lipoxygenase products in the pathogenesis and therapy of psoriasis and other dermatoses. Arch. Dermatol. 1983, 119, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Madden, S.K.; Flanagan, K.L.; Jones, G. How lifestyle factors and their associated pathogenetic mechanisms impact psoriasis. Clin. Nutr. 2019. [Google Scholar] [CrossRef] [PubMed]
- Namazi, M.R. Why is psoriasis uncommon in Africans? The influence of dietary factors on the expression of psoriasis. Int. J. Dermatol. 2004, 43, 391–392. [Google Scholar] [CrossRef] [PubMed]
- Horrobin, D.F. Low prevalences of coronary heart disease (CHD), psoriasis, asthma and rheumatoid arthritis in Eskimos: Are they caused by high dietary intake of eicosapentaenoic acid (EPA), a genetic variation of essential fatty acid (EFA) metabolism or a combination of. Med. Hypotheses 1987, 22, 421–428. [Google Scholar] [CrossRef]
- Boelsma, E.; Hendriks, H.F.J.; Roza, L. Nutritional skin care: Health effects of micronutrients and fatty acids. Am. J. Clin. Nutr. 2001, 73, 853–864. [Google Scholar] [CrossRef]
- Pizzini, A.; Lunger, L.; Sonnweber, T.; Weiss, G.; Tancevski, I. The Role of Omega-3 Fatty Acids in the Setting of Coronary Artery Disease and COPD: A Review. Nutrients 2018, 10, 1864. [Google Scholar] [CrossRef]
- Gupta, A.K.; Ellis, C.N.; Goldfarb, M.T.; Hamilton, T.A.; Voorhees, J.J. The Role of Fish Oil in Psoriasis: A Randomized, Double-Blind, Placebo-Controlled Study to Evaluate the Effect of Fish Oil and Topical Corticosteroid Therapy in Psoriasis. Int. J. Dermatol. 1990, 29, 591–595. [Google Scholar] [CrossRef]
- Upala, S.; Yong, W.C.; Theparee, T.; Sanguankeo, A. Effect of omega-3 fatty acids on disease severity in patients with psoriasis: A systematic review. Int. J. Rheum. Dis. 2017, 20, 442–450. [Google Scholar] [CrossRef]
- Feingold, K.R. Thematic review series: Skin lipids. The role of epidermal lipids in cutaneous permeability barrier homeostasis. J. Lipid Res. 2007, 48, 2531–2546. [Google Scholar] [CrossRef]
- Ricketts, J.R.; Rothe, M.J.; Grant-Kels, J.M. Nutrition and psoriasis. Clin. Dermatol. 2010, 28, 615–626. [Google Scholar] [CrossRef]
- Das, U.N. Essential fatty acids-a review. Curr. Pharm. Biotechnol. 2006, 7, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Arican, O.; Aral, M.; Sasmaz, S.; Ciragil, P. Serum levels of TNF-α, IFN-γ, IL-6, IL-8, IL-12, IL-17, and IL-18 in patients with active psoriasis and correlation with disease severity. Mediators Inflamm. 2005, 2005, 273–279. [Google Scholar] [CrossRef]
- Sawada, Y.; Honda, T.; Nakamizo, S.; Otsuka, A.; Ogawa, N.; Kobayashi, Y.; Nakamura, M.; Kabashima, K. Resolvin E1 attenuates murine psoriatic dermatitis. Sci. Rep. 2018, 8, 11873. [Google Scholar] [CrossRef] [PubMed]
- Maurice, P.D.L.; Allen, B.R.; Barkley, A.S.J.; Cockbill, S.R.; Stammers, J.; Bather, P.C. The effects of dietary supplementation with fish oil in patients with psoriasis. Br. J. Dermatol. 1987, 117, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Soyland, E.; Lea, T.; Sandstad, B.; Section, A.D. Dietary supplementation with very long-chain n-3 fatty acids in man decreases expression of the interleukin-2 receptor (CD25) on mitogen-stimulated lymphocytes from patients with inflammatory skin diseases. Eur. J. Clin. Investig. 1993, 2, 236–242. [Google Scholar] [CrossRef]
- Allen, B.R. Fish oil in combination with other therapies in the treatment of psoriasis. World Rev. Nutr. Diet. 1991, 66, 436–445. [Google Scholar]
- Rossmeisl, M.; Jelenik, T.; Jilkova, Z.; Slamova, K.; Kus, V.; Hensler, M.; Medrikova, D.; Povysil, C.; Flachs, P.; Mohamed-Ali, V. Prevention and reversal of obesity and glucose intolerance in mice by DHA derivatives. Obesity 2009, 17, 1023–1031. [Google Scholar] [CrossRef]
- Armstrong, A.W.; Harskamp, C.T.; Armstrong, E.J. The association between psoriasis and obesity: A systematic review and meta-analysis of observational studies. Nutr. Diabetes 2012, 2, e54. [Google Scholar] [CrossRef]
- Melnik, B.C. Western Diet-Mediated mTORC1-Signaling in Acne, Psoriasis, Atopic Dermatitis, and Related Diseases of Civilization: Therapeutic Role of Plant-Derived Natural mTORC1 Inhibitors. In Bioactive Dietary Factors and Plant Extracts in Dermatology; Watson, R.R., Zibadi, S., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 397–419. [Google Scholar]
- Timoszuk, M.; Bielawska, K.; Skrzydlewska, E. Evening primrose (Oenothera biennis) biological activity dependent on chemical composition. Antioxidants 2018, 7, 108. [Google Scholar] [CrossRef]
- Dowlatshahi, E.A.; Van Der Voort, E.A.M.; Arends, L.R.; Nijsten, T. Markers of systemic inflammation in psoriasis: A systematic review and meta-analysis. Br. J. Dermatol. 2013, 169, 266–282. [Google Scholar] [CrossRef]
- Lima, E.A.; Lima, M.M.D.A.; Marques, C.D.L.; Duarte, A.L.B.P.; Pita, I.R.; Pita, M.G.R. Peroxisome proliferator-activated receptor agonists (PPARs): A promising prospect in the treatment of psoriasis and psoriatic arthritis. An. Bras. Dermatol. 2013, 88, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.N.; Varani, J.; Fisher, G.J.; Zeigler, M.E.; Pershadsingh, H.A.; Benson, S.C.; Chi, Y.; Kurtz, T.W. Troglitazone improves psoriasis and normalizes models of proliferative skin disease: Ligands for peroxisome proliferator-activated receptor-γ inhibit keratinocyte proliferation. Arch. Dermatol. 2000, 136, 609–616. [Google Scholar] [CrossRef]
- Hanley, K.; Kömüves, L.G.; Bass, N.M.; He, S.; Jiang, Y.; Crumrine, D.; Appel, R.; Friedman, M.; Bettencourt, J.; Min, K. Fetal Epidermal Differentiation and Barrier Development In Vivo is Accelerated by Nuclear Hormone Receptor Activators1. J. Investig. Dermatol. 1999, 113, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Bongartz, T.; Coras, B.; Vogt, T.; Scholmerich, J.; Muller-Ladner, U. Treatment of active psoriatic arthritis with the PPARγ ligand pioglitazone: An open-label pilot study. Rheumatology 2004, 44, 126–129. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, X.; Chen, X.; Xue, F.; Zheng, J. Topical application of a linoleic acid-ceramide containing moisturizer exhibit therapeutic and preventive benefits for psoriasis vulgaris: A randomized controlled trial. Dermatol. Ther. 2015, 28, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Bhoir, S.S.; Vishwapathi, V.; Singh, K.K. Antipsoriatic potential of Annona squamosa seed oil: An in vitro and in vivo evaluation. Phytomedicine 2019, 54, 265–277. [Google Scholar] [CrossRef]
- Tuchayi, S.M.; Makrantonaki, E.; Ganceviciene, R.; Dessinioti, C.; Feldman, S.R.; Zouboulis, C.C. Acne vulgaris. Nat. Rev. Dis. Prim. 2015, 1, 15029. [Google Scholar] [CrossRef]
- Layton, A.M. Optimal management of acne to prevent scarring and psychological sequelae. Am. J. Clin. Dermatol. 2001, 2, 135–141. [Google Scholar] [CrossRef]
- Cong, T.-X.; Hao, D.; Wen, X.; Li, X.-H.; He, G.; Jiang, X. From pathogenesis of acne vulgaris to anti-acne agents. Arch. Dermatol. Res. 2019, 311, 337–349. [Google Scholar] [CrossRef]
- Gollnick, H.P.M. From new findings in acne pathogenesis to new approaches in treatment. J. Eur. Acad. Dermatology Venereol. 2015, 29, 1–7. [Google Scholar] [CrossRef]
- Kircik, L.H. What’s new in the management of acne vulgaris. Cutis 2019, 104, 48–52. [Google Scholar] [PubMed]
- Claudel, J.-P.; Auffret, N.; Leccia, M.T.; Poli, F.; Dréno, B. Acne and nutrition: Hypotheses, myths and facts. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 1631–1637. [Google Scholar] [CrossRef] [PubMed]
- Rezaković, S.; Bukvić, Z.M.; Basta-Juzbašić, A. Acne and diet: Facts and controversies. Acta Dermatovenerol. Croat. ADC 2012, 20, 170–174. [Google Scholar]
- Jung, J.Y.; Yoon, M.Y.; Min, S.U.; Hong, J.S.; Choi, Y.S.; Suh, D.H. The influence of dietary patterns on acne vulgaris in Koreans. Eur. J. Dermatol. 2010, 20, 768–772. [Google Scholar]
- Di Landro, A.; Cazzaniga, S.; Parazzini, F.; Ingordo, V.; Cusano, F.; Atzori, L.; Cutrì, F.T.; Musumeci, M.L.; Zinetti, C.; Pezzarossa, E. Family history, body mass index, selected dietary factors, menstrual history, and risk of moderate to severe acne in adolescents and young adults. J. Am. Acad. Dermatol. 2012, 67, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Bowe, W.P.; Joshi, S.S.; Shalita, A.R. Diet and acne. J. Am. Acad. Dermatol. 2010, 63, 124–141. [Google Scholar] [CrossRef]
- Tanghetti, E.A. The role of inflammation in the pathology of acne. J. Clin. Aesthet. Dermatol. 2013, 6, 27. [Google Scholar]
- Snodgrass, R.G.; Huang, S.; Choi, I.-W.; Rutledge, J.C.; Hwang, D.H. Inflammasome-mediated secretion of IL-1β in human monocytes through TLR2 activation; modulation by dietary fatty acids. J. Immunol. 2013, 191, 4337–4347. [Google Scholar] [CrossRef]
- Dréno, B. What is new in the pathophysiology of acne, an overview. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 8–12. [Google Scholar] [CrossRef]
- Dreno, B.; Gollnick, H.P.M.; Kang, S.; Thiboutot, D.; Bettoli, V.; Torres, V.; Leyden, J.; Acne, G.A. to I.O. in Understanding innate immunity and inflammation in acne: Implications for management. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 3–11. [Google Scholar] [CrossRef]
- Graham, G.M.; Farrar, M.D.; Cruse-Sawyer, J.E.; Holland, K.T.; Ingham, E. Proinflammatory cytokine production by human keratinocytes stimulated with Propionibacterium acnes and P. acnes GroEL. Br. J. Dermatol. 2004, 150, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, I.; Danby, F.W.; Ju, Q.; Wang, X.; Xiang, L.F.; Xia, L.; Chen, W.; Nagy, I.; Picardo, M.; Suh, D.H. New developments in our understanding of acne pathogenesis and treatment. Exp. Dermatol. 2009, 18, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Contassot, E.; French, L.E. New insights into acne pathogenesis: Propionibacterium acnes activates the inflammasome. J. Investig. Dermatol. 2014, 134, 310–313. [Google Scholar] [CrossRef]
- Kim, H.H.; Lee, Y.; Eun, H.C.; Chung, J.H. Eicosapentaenoic acid inhibits TNF-α-induced matrix metalloproteinase-9 expression in human keratinocytes, HaCaT cells. Biochem. Biophys. Res. Commun. 2008, 368, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Moon, S.Y.; Sohn, M.Y.; Lee, W.J. Insulin-like growth factor-1 increases the expression of inflammatory biomarkers and sebum production in cultured sebocytes. Ann. Dermatol. 2017, 29, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Seifert, M.F.; Ney, D.M.; Grahn, M.; Grant, A.L.; Allen, K.G.D.; Watkins, B.A. Dietary conjugated linoleic acids alter serum IGF-I and IGF binding protein concentrations and reduce bone formation in rats fed (n-6) or (n-3) fatty acids. J. Bone Miner. Res. 1999, 14, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C. Linking diet to acne metabolomics, inflammation, and comedogenesis: An update. Clin. Cosmet. Investig. Dermatol. 2015, 8, 371. [Google Scholar] [CrossRef]
- Scanferlato, R.; Bortolotti, M.; Sansone, A.; Chatgilialoglu, C.; Polito, L.; De Spirito, M.; Maulucci, G.; Bolognesi, A.; Ferreri, C. Hexadecenoic fatty acid positional isomers and de novo PUFA synthesis in colon cancer cells. Int. J. Mol. Sci. 2019, 20, 832. [Google Scholar] [CrossRef]
- Sansone, A.; Tolika, E.; Louka, M.; Sunda, V.; Deplano, S.; Melchiorre, M.; Anagnostopoulos, D.; Chatgilialoglu, C.; Formisano, C.; Di Micco, R. Hexadecenoic fatty acid isomers in human blood lipids and their relevance for the interpretation of lipidomic profiles. PLoS ONE 2016, 11, e0152378. [Google Scholar] [CrossRef]
- Jung, J.Y.; Kwon, H.H.; Hong, J.S.; Yoon, J.Y.; Park, M.S.; Jang, M.Y.; Suh, D.H. Effect of dietary supplementation with omega-3 fatty acid and gamma-linolenic acid on acne vulgaris: A randomised, double-blind, controlled trial. Acta Derm. Venereol. 2014, 94, 521–526. [Google Scholar] [CrossRef]
- Ziboh, V.A.; Cho, Y.; Mani, I.; Xi, S. Biological significance of essential fatty acids/prostanoids/lipoxygenase-derived monohydroxy fatty acids in the skin. Arch. Pharm. Res. 2002, 25, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Rubin, M.G.; Kim, K.; Logan, A.C. Acne vulgaris, mental health and omega-3 fatty acids: A report of cases. Lipids Health Dis. 2008, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Khayef, G.; Young, J.; Burns-Whitmore, B.; Spalding, T. Effects of fish oil supplementation on inflammatory acne. Lipids Health Dis. 2012, 11, 165. [Google Scholar] [CrossRef] [PubMed]
- Park, K.Y.; Ko, E.J.; Kim, I.S.; Li, K.; Kim, B.J.; Seo, S.J. The Effect of Evening Primrose Oil for the Prevention of Xerotic Cheilitis in Acne Patients Being Treated with Isotretinoin: A Pilot Study. Ann. Dermatol. 2014, 26, 706–712. [Google Scholar] [CrossRef]
- Krishna, S.; Okhovat, J.-P.; Kim, J.; Kim, C.N. Influence of omega-3 fatty acids on triglyceride levels in patients using isotretinoin. JAMA Dermatol. 2015, 151, 101–102. [Google Scholar] [CrossRef][Green Version]



{kind=link}
{kind=link}
| Food Source | ω-3 | ω-6 | Reference | |||||
|---|---|---|---|---|---|---|---|---|
| ALA * | EPA * | DHA * | LA * | ARA * | DPA * | |||
| Oil | corn | 0.6 | - | - | 49.83 | - | - | [13] |
| sunflower | 0.33 | - | - | 49.89 | - | - | ||
| soybean | 7.6 | - | - | 51.36 | - | - | ||
| wheat germ | 5.3 | - | - | 55.1 | - | - | ||
| Canola | 9.15 | - | - | 18.65 | - | - | [14] | |
| Safflower | 0.1 | - | - | 12.72 | - | - | ||
| Flaxseed | 53.37 | - | - | 14.33 | - | - | ||
| Fish oil | Salmon | - | 13.3 | 18.23 | - | - | 2.99 | [14] |
| Sardine | - | 10.15 | 10.66 | - | - | 1.97 | ||
| Herring | - | 6.28 | 4.21 | - | - | 0.62 | ||
| menhaden | - | 13.18 | 8.56 | - | - | 4.92 | ||
| Vegetables | lettuce, raw | 0.15 | - | - | 0.06 | - | - | [13] |
| green broccoli, raw | 0.11 | - | - | 0.03 | - | - | ||
| brussels sprouts, raw | 0.17 | - | - | 0.08 | - | - | ||
| Fish | salmon, raw | 0.09 | 0.89 | 1.19 | 0.15 | 0.05 | - | [13] |
| herring, raw | 0.19 | 1.09 | 1.01 | 0.22 | 0.1 | - | ||
| sardine, raw | - | 0.51 | 1.16 | 0.06 | 0.04 | - | ||
| trout, raw | 0.1 | 0.15 | 0.5 | 0.37 | 0.05 | - | ||
| cod, dried | - | 0.02 | 0.62 | 0.03 | 0.12 | - | ||
| Meat | lamb, lean meat | 0.11 | - | - | 0.11 | - | - | [13] |
| pork, fat and lean meat, without visible fat | - | - | - | 1.63 | 0.03 | - | ||
| beef, veal, 4 months, lean meat, without visible fat | 0.08 | - | - | 0.13 | - | - | ||
| Seeds | chia, dried | 17.83 | - | - | 5.84 | - | - | [14] |
| walnuts, dried | 6.64 | - | - | 34.02 | - | - | [13] | |
| hazelnuts, dried | 0.11 | - | - | 5.09 | - | - | ||
| almond, dried | 0.3 | - | - | 10.54 | - | - | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balić, A.; Vlašić, D.; Žužul, K.; Marinović, B.; Bukvić Mokos, Z. Omega-3 Versus Omega-6 Polyunsaturated Fatty Acids in the Prevention and Treatment of Inflammatory Skin Diseases. Int. J. Mol. Sci. 2020, 21, 741. https://doi.org/10.3390/ijms21030741
Balić A, Vlašić D, Žužul K, Marinović B, Bukvić Mokos Z. Omega-3 Versus Omega-6 Polyunsaturated Fatty Acids in the Prevention and Treatment of Inflammatory Skin Diseases. International Journal of Molecular Sciences. 2020; 21(3):741. https://doi.org/10.3390/ijms21030741
Chicago/Turabian StyleBalić, Anamaria, Domagoj Vlašić, Kristina Žužul, Branka Marinović, and Zrinka Bukvić Mokos. 2020. "Omega-3 Versus Omega-6 Polyunsaturated Fatty Acids in the Prevention and Treatment of Inflammatory Skin Diseases" International Journal of Molecular Sciences 21, no. 3: 741. https://doi.org/10.3390/ijms21030741
APA StyleBalić, A., Vlašić, D., Žužul, K., Marinović, B., & Bukvić Mokos, Z. (2020). Omega-3 Versus Omega-6 Polyunsaturated Fatty Acids in the Prevention and Treatment of Inflammatory Skin Diseases. International Journal of Molecular Sciences, 21(3), 741. https://doi.org/10.3390/ijms21030741