Hypoxic-Inflammatory Responses under Acute Hypoxia: In Vitro Experiments and Prospective Observational Expedition Trial

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Results
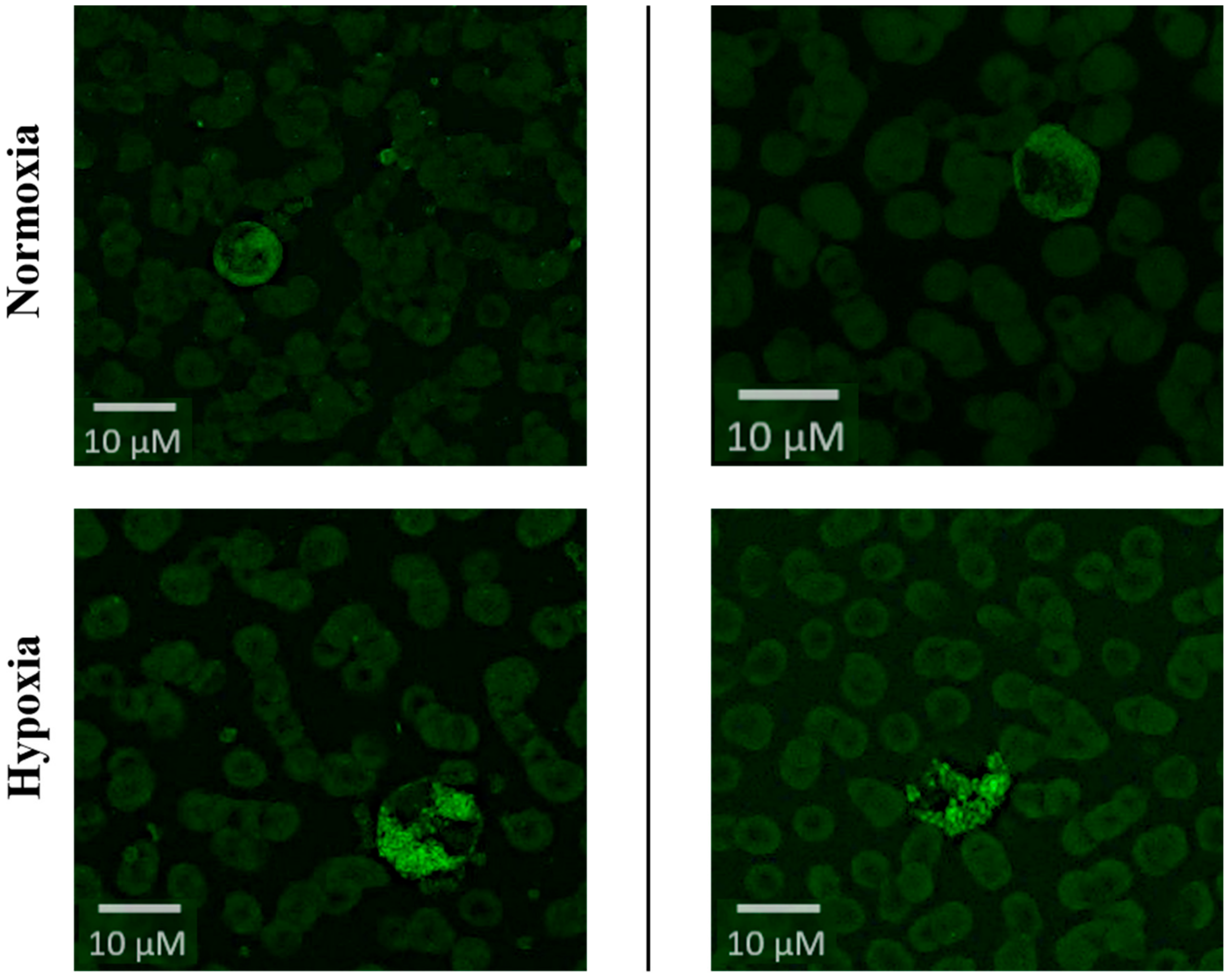
2.1. Induction of Hypoxic-Inflammatory Pathways
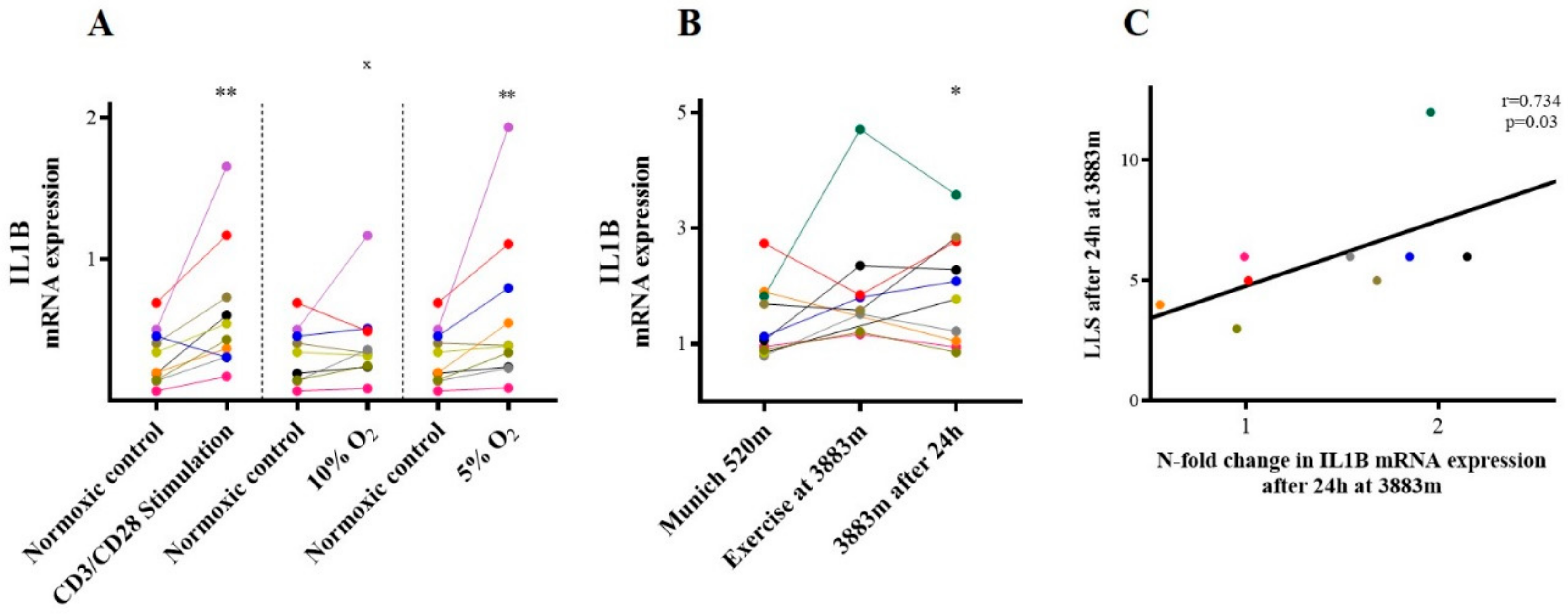
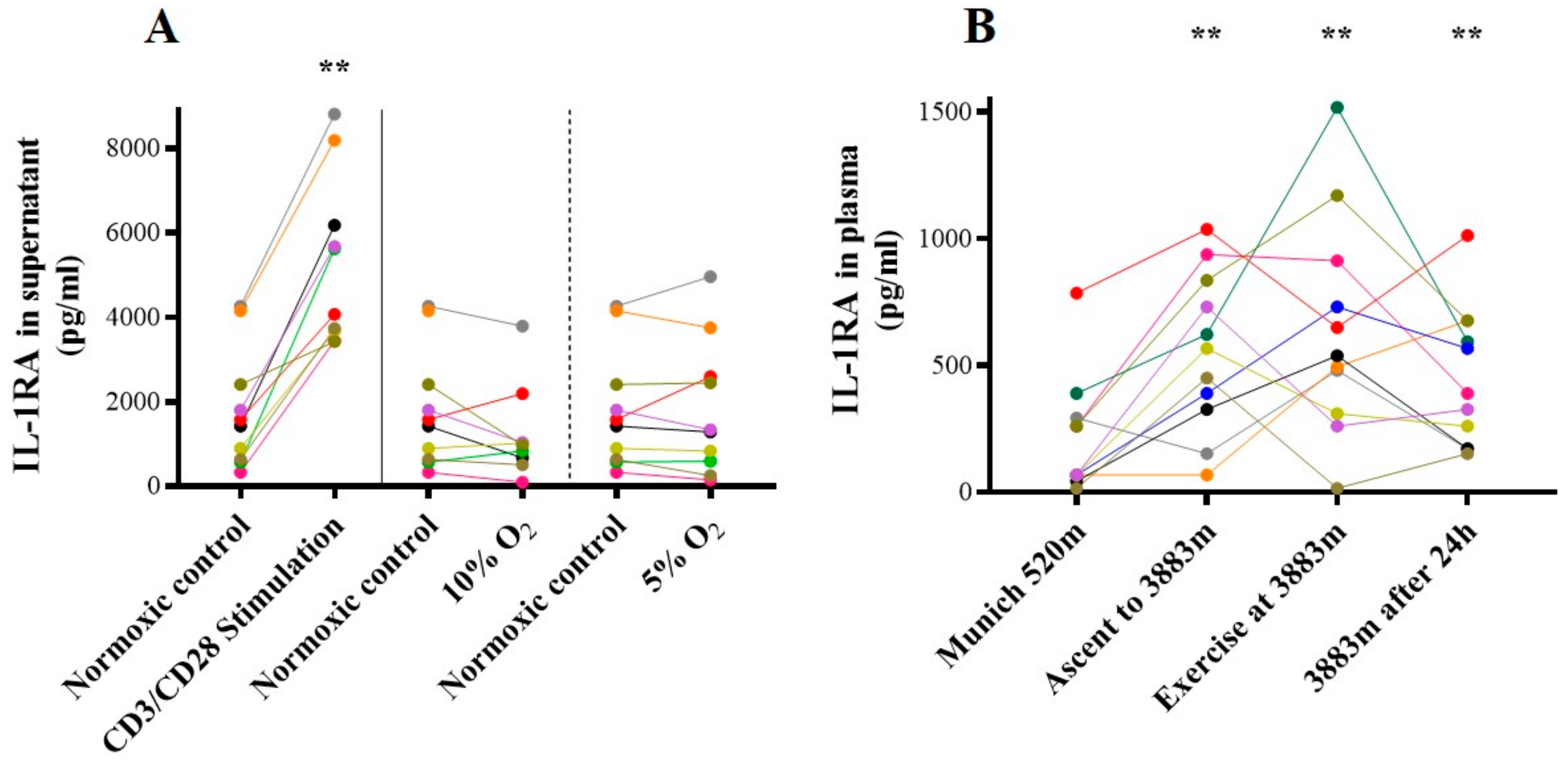
2.2. IL-1β
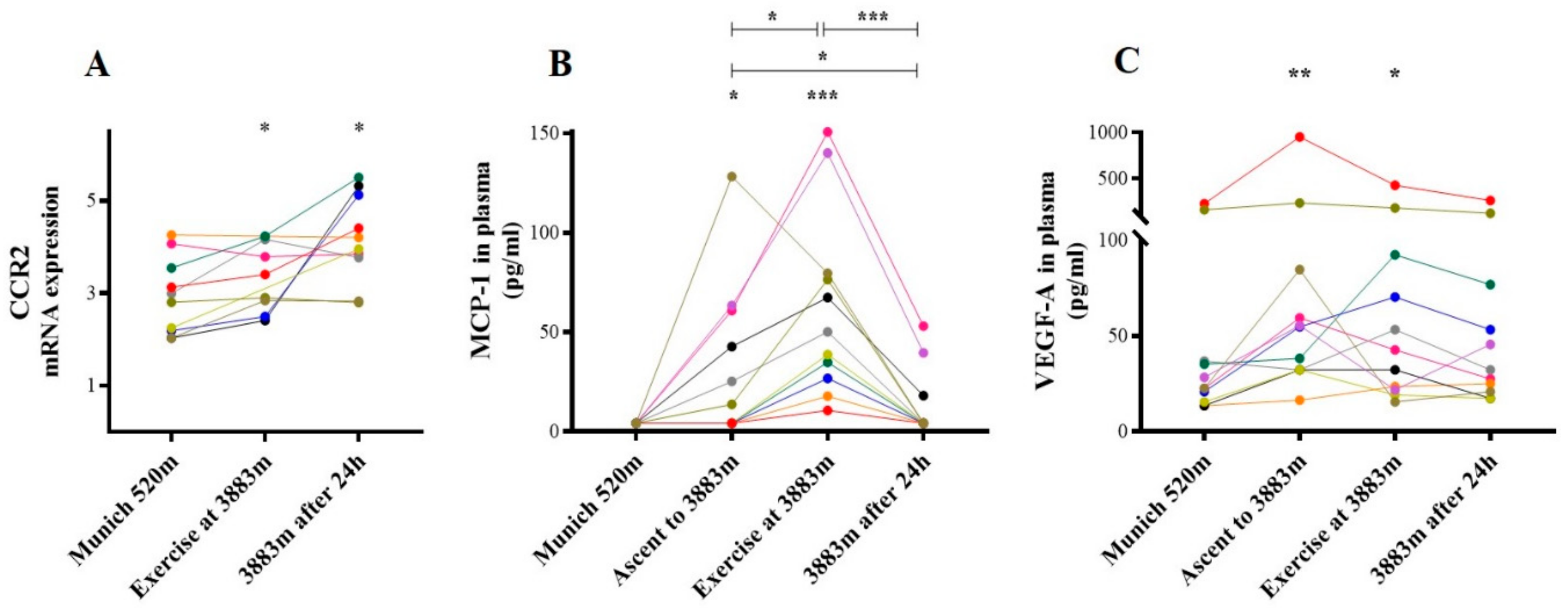
2.3. Chemokine Receptors
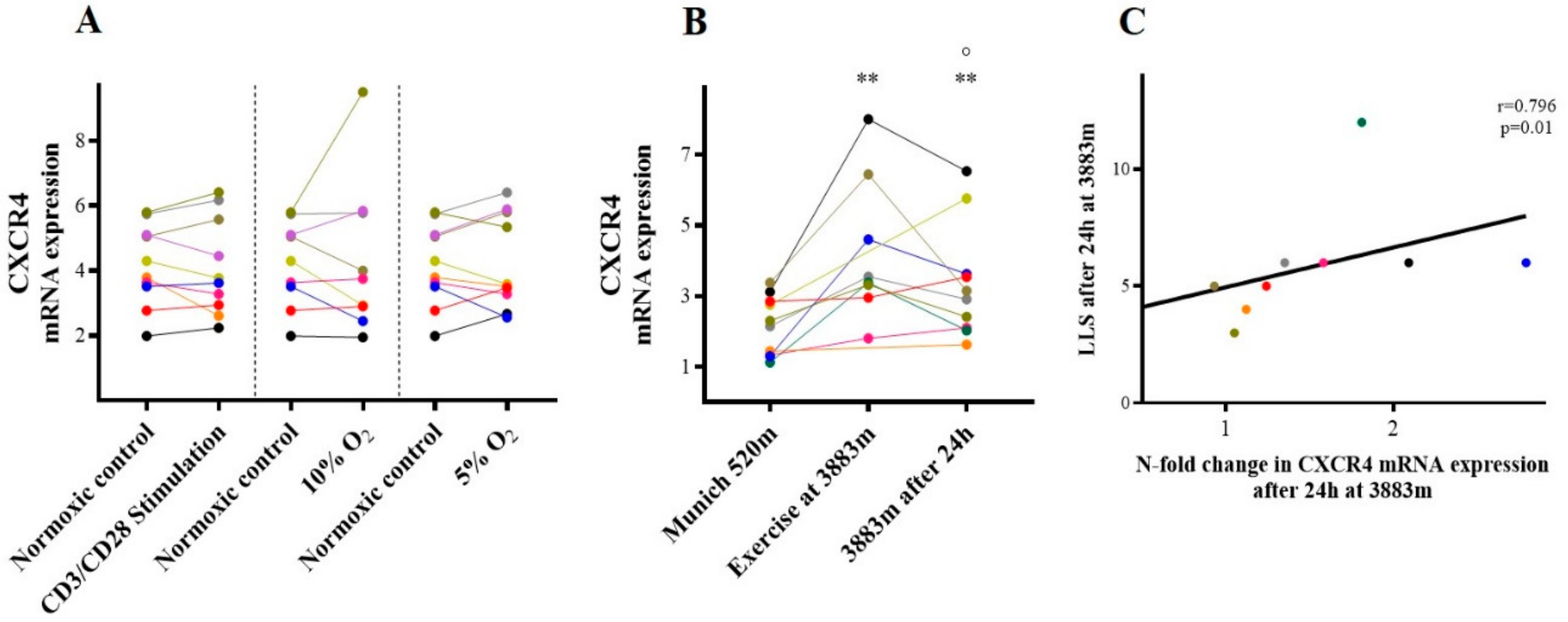
2.3.1. CXCR-4/SDF-1
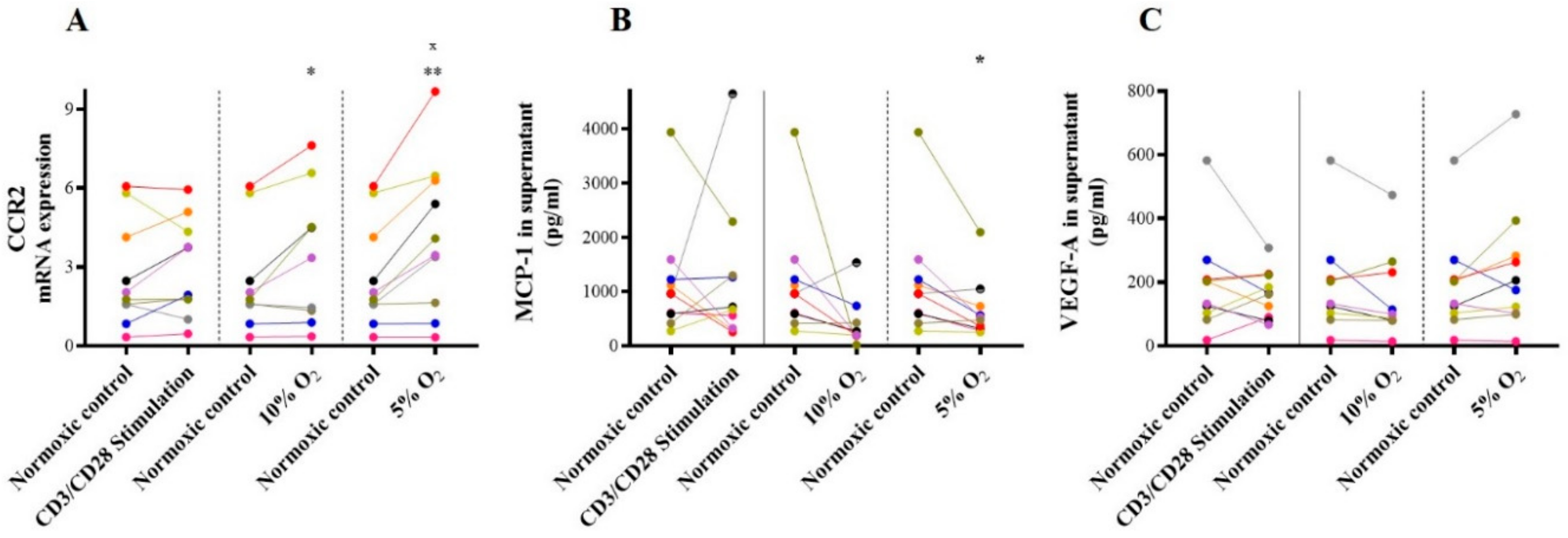
2.3.2. CCR-2/MCP-1 and VEGF-A
2.4. Proinflammatory Cytokines
2.4.1. Interleukin-3
2.4.2. Interleukin-6
2.4.3. TNF-α
2.5. Anti-Inflammatory Cytokines
2.5.1. IL-10
2.5.2. IL-1RA
3. Discussion
4. Materials and Methods
4.1. Volunteers Characteristics
4.2. In Vitro Experiments
4.2.1. PBMC Isolation
4.2.2. Cell CULTURE Experiments
- patm = 625.9 hPa = 469 mmHg
- Sea level: 1013 hPa, FiO2 = 0.21
- Little Matterhorn summit (3883 m a.s.l.): FiO2 = 625.9 hPa/1013 hPa × 0.21= 0.13.
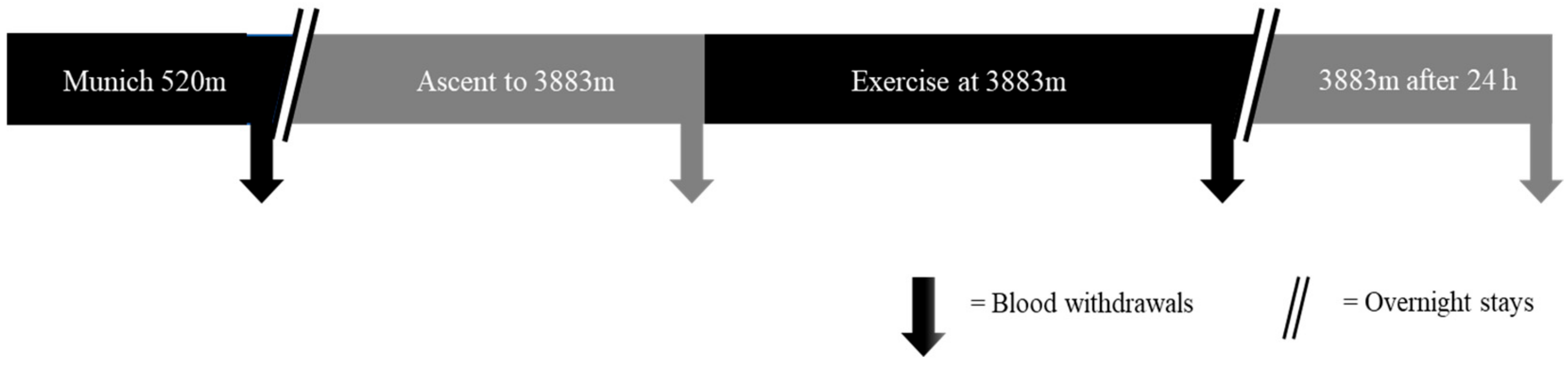
4.3. Expedition Trial
4.4. Molecular Analyses
4.4.1. HIF-Visualization
4.4.2. Real-Time PCR (qPCR)
4.4.3. Cytokine Measurements from Plasma and Supernatant
4.5. Clinical Analyses
Acute Mountain Sickness
4.6. Statistical Analysis
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abe, T.; Kitaoka, Y.; Kikuchi, D.M.; Takeda, K.; Numata, O.; Takemasa, T. High-intensity interval training-induced metabolic adaptation coupled with an increase in Hif-1α and glycolytic protein expression. J. Appl. Physiol. 2015, 119, 1297–1302. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, M.E.; Rundqvist, H. Skeletal muscle hypoxia-inducible factor-1 and exercise. Exp. Physiol. 2016, 101, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Shen, S.-M.; Zhao, X.-Y.; Chen, G.-Q. Targeted genes and interacting proteins of hypoxia inducible factor-1. Int. J. Biochem. Mol. Biol. 2012, 3, 165–178. [Google Scholar] [PubMed]
- Eltzschig, H.K.; Carmeliet, P. Hypoxia and inflammation. N. Engl. J. Med. 2011, 364, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Frede, S.; Berchner-Pfannschmidt, U.; Fandrey, J. Regulation of Hypoxia-Inducible Factors During Inflammation. In Methods in Enzymology; Academic Press: Camebridge, MA, USA, 2007; Vol. 435, pp. 403–419. [Google Scholar]
- Schafer, S.T.; Frede, S.; Winning, S.; Bick, A.; Roshangar, P.; Fandrey, J.; Peters, J.; Adamzik, M. Hypoxia-inducible factor and target gene expression are decreased in patients with sepsis: Prospective observational clinical and cellular studies. Anesthesiology 2013, 118, 1426–1436. [Google Scholar] [CrossRef]
- Hotchkiss, R.S.; Coopersmith, C.M.; McDunn, J.E.; Ferguson, T.A. The sepsis seesaw: Tilting toward immunosuppression. Nat. Med. 2009, 15, 496–497. [Google Scholar] [CrossRef]
- Frede, S.; Stockmann, C.; Freitag, P.; Fandrey, J. Bacterial lipopolysaccharide induces HIF-1 activation in human monocytes via p44/42 MAPK and NF-κB. Biochem. J. 2006, 396, 517–527. [Google Scholar] [CrossRef]
- Frede, S.; Stockmann, C.; Winning, S.; Freitag, P.; Fandrey, J. Hypoxia-inducible factor (HIF) 1alpha accumulation and HIF target gene expression are impaired after induction of endotoxin tolerance. J. Immunol. 2009, 182, 6470–6476. [Google Scholar] [CrossRef]
- Winning, S.; Splettstoesser, F.; Fandrey, J.; Frede, S. Acute hypoxia induces HIF-independent monocyte adhesion to endothelial cells through increased intercellular adhesion molecule-1 expression: The role of hypoxic inhibition of prolyl hydroxylase activity for the induction of NF-kappa B. J. Immunol. 2010, 185, 1786–1793. [Google Scholar] [CrossRef]
- Zepeda, A.B.; Pessoa, A.; Castillo, R.L.; Figueroa, C.A.; Pulgar, V.M.; Farías, J.G. Cellular and molecular mechanisms in the hypoxic tissue: Role of HIF-1 and ROS. Cell Biochem. Funct. 2013, 31, 451–459. [Google Scholar] [CrossRef]
- Drewry, A.M.; Hotchkiss, R.S. Revising definitions of sepsis. Nat. Rev. Nephrol. 2015, 11, 326–328. [Google Scholar] [CrossRef] [PubMed]
- Seeley, J.J.; Ghosh, S. Molecular mechanisms of innate memory and tolerance to LPS. J. Leukoc. Biol. 2017, 101, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.A. The sepsis seesaw: Seeking a heart salve. Nat. Med. 2009, 15, 497–498. [Google Scholar] [CrossRef]
- Buroker, N.E.; Ning, X.-H.; Zhou, Z.-N.; Li, K.; Cen, W.-J.; Wu, X.-F.; Zhu, W.-Z.; Scott, C.R.; Chen, S.-H. EPAS1 and EGLN1 associations with high altitude sickness in Han and Tibetan Chinese at the Qinghai-Tibetan Plateau. Blood Cells Mol. Dis. 2012, 49, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Buroker, N.E.; Ning, X.-H.; Zhou, Z.-N.; Li, K.; Cen, W.-J.; Wu, X.-F.; Zhu, W.-Z.; Scott, C.R.; Chen, S.-H. SNPs, linkage disequilibrium, and chronic mountain sickness in Tibetan Chinese. Hypoxia 2017, 5, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.; Zhang, J.; Jin, J.; Qin, J.; Li, H.; Huang, L. Variants of the low oxygen sensors EGLN1 and HIF-1AN associated with acute mountain sickness. Int. J. Mol. Sci. 2014, 15, 21777–21787. [Google Scholar] [CrossRef]
- Song, D.; Li, L.-S.; Arsenault, P.R.; Tan, Q.; Bigham, A.W.; Heaton-Johnson, K.J.; Master, S.R.; Lee, F.S. Defective Tibetan PHD2 binding to p23 links high altitude adaption to altered oxygen sensing. J. Biol. Chem. 2014, 289, 14656–14665. [Google Scholar] [CrossRef]
- Mazzeo, R.S. Altitude, exercise and immune function. Exerc. Immunol. Rev. 2005, 11, 6–16. [Google Scholar]
- Swenson, E.R. Hypoxic pulmonary vasoconstriction. High. Alt. Med. Biol. 2013, 14, 101–110. [Google Scholar] [CrossRef]
- Labrousse-Arias, D.; Castillo-González, R.; Rogers, N.M.; Torres-Capelli, M.; Barreira, B.; Aragonés, J.; Cogolludo, Á.; Isenberg, J.S.; Calzada, M.J. HIF-2α-mediated induction of pulmonary thrombospondin-1 contributes to hypoxia-driven vascular remodelling and vasoconstriction. Cardiovasc. Res. 2016, 109, 115–130. [Google Scholar] [CrossRef]
- Dötsch, A.; Eisele, L.; Rabeling, M.; Rump, K.; Walstein, K.; Bick, A.; Cox, L.; Engler, A.; Bachmann, H.S.; Jöckel, K.-H.; et al. Hypoxia Inducible Factor-2 Alpha and Prolinhydroxylase 2 Polymorphisms in Patients with Acute Respiratory Distress Syndrome (ARDS). Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Höcker, A.; Rabeling, M.; Bick, A.; Cox, L.; Kreuzer, M.; Engler, A.; Walstein, K.; Bachmann, H.S.; Jöckel, K.-H.; Eisele, L.; et al. Hypoxia inducible factor-1 alpha and prolinhydroxlase 2 polymorphisms in patients with severe sepsis: A prospective observational trial. BMC Anesthesiol. 2016, 16, 656–659. [Google Scholar] [CrossRef] [PubMed]
- Kammerer, T.; Faihs, V.; Hulde, N.; Bayer, A.; Hübner, M.; Brettner, F.; Karlen, W.; Kröpfl, J.M.; Rehm, M.; Spengler, C.; et al. Changes of hemodynamic and cerebral oxygenation after exercise in normobaric and hypobaric hypoxia: Associations with acute mountain sickness. Ann. Occup. Environ. Med. 2018, 30, 66. [Google Scholar] [CrossRef]
- Roach, R.C.; Hackett, P.H.; Oelz, O.; Bärtsch, P.; Luks, A.M.; MacInnis, M.J.; Baillie, J.K. Lake Louise AMS Score Consensus Committee The 2018 Lake Louise Acute Mountain Sickness Score. High. Alt. Med. Biol. 2018, 19, 4–6. [Google Scholar] [CrossRef]
- Schaber, M.; Leichtfried, V.; Fries, D.; Wille, M.; Gatterer, H.; Faulhaber, M.; Würtinger, P.; Schobersberger, W. Influence of Acute Normobaric Hypoxia on Hemostasis in Volunteers with and without Acute Mountain Sickness. Biomed. Res. Int. 2015, 2015, 593938. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Deng, J.; Wei, X.; Jayasuriya, C.T.; Zhou, J.; Chen, Q.; Zhang, J.; Wei, L.; Wei, F. Blockade of hypoxia-induced CXCR4 with AMD3100 inhibits production of OA-associated catabolic mediators IL-1β and MMP-13. Mol. Med. Rep. 2016, 14, 1475–1482. [Google Scholar] [CrossRef]
- Dai, T.; Hu, Y.; Zheng, H. Hypoxia increases expression of CXC chemokine receptor 4 via activation of PI3K/Akt leading to enhanced migration of endothelial progenitor cells. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1820–1827. [Google Scholar]
- Choi, S.; Chung, H.; Hong, H.; Kim, S.Y.; Kim, S.-E.; Seoh, J.-Y.; Moon, C.M.; Yang, E.G.; Oh, E.-S. Inflammatory hypoxia induces syndecan-2 expression through IL-1β-mediated FOXO3a activation in colonic epithelia. FASEB J. 2017, 31, 1516–1530. [Google Scholar] [CrossRef]
- De Vries, H.E.; Blom-Roosemalen, M.C.M.; van Oosten, M.; de Boer, A.G.; van Berkel, T.J.C.; Breimer, D.D.; Kuiper, J. The influence of cytokines on the integrity of the blood-brain barrier in vitro. J. Neuroimmunol. 1996, 64, 37–43. [Google Scholar] [CrossRef]
- Wang, C.; Jiang, H.; Duan, J.; Chen, J.; Wang, Q.; Liu, X.; Wang, C. Exploration of Acute Phase Proteins and Inflammatory Cytokines in Early Stage Diagnosis of Acute Mountain Sickness. High. Alt. Med. Biol. 2018, 19, 170–177. [Google Scholar] [CrossRef]
- Momcilović, M.; Mostarica-Stojković, M.; Miljković, D. CXCL12 in control of neuroinflammation. Immunol. Res. 2012, 52, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ransohoff, R.M. Multiple roles of chemokine CXCL12 in the central nervous system: A migration from immunology to neurobiology. Prog. Neurobiol. 2008, 84, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Huang, X.; Zhao, T.; Qiao, M.; Zhao, X.; Zhao, M.; Xu, L.; Zhao, Y.; Wu, L.; Wu, K.; et al. Hypoxia augments LPS-induced inflammation and triggers high altitude cerebral edema in mice. Brain Behav. Immun. 2017, 64, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Manthiram, K.; Zhou, Q.; Aksentijevich, I.; Kastner, D.L. The monogenic autoinflammatory diseases define new pathways in human innate immunity and inflammation. Nat. Immunol. 2017, 18, 832–842. [Google Scholar] [CrossRef]
- Giesbrecht, K.; Eberle, M.-E.; Wölfle, S.J.; Sahin, D.; Sähr, A.; Oberhardt, V.; Menne, Z.; Bode, K.A.; Heeg, K.; Hildebrand, D. IL-1β As Mediator of Resolution That Reprograms Human Peripheral Monocytes toward a Suppressive Phenotype. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Liu, B.; Chen, J.; Zhang, L.; Gao, Y.; Cui, J.; Zhang, E.; Xu, G.; Liang, Y.; Liang, Y.; Wang, J.; et al. IL-10 Dysregulation in Acute Mountain Sickness Revealed by Transcriptome Analysis. Front. Immunol. 2017, 8, 628. [Google Scholar] [CrossRef]
- Lundeberg, J.; Feiner, J.R.; Schober, A.; Sall, J.W.; Eilers, H.; Bickler, P.E. Increased Cytokines at High Altitude: Lack of Effect of Ibuprofen on Acute Mountain Sickness, Physiological Variables, or Cytokine Levels. High. Alt. Med. Biol. 2018, 19, 249–258. [Google Scholar] [CrossRef]
- Swenson, E.R.; MacDonald, A.; Vatheuer, M.; Maks, C.; Treadwell, A.; Allen, R.; Schoene, R.B. Acute mountain sickness is not altered by a high carbohydrate diet nor associated with elevated circulating cytokines. Aviat. Space Environ. Med. 1997, 68, 499–503. [Google Scholar]
- Schild, M.; Eichner, G.; Beiter, T.; Zügel, M.; Krumholz-Wagner, I.; Hudemann, J.; Pilat, C.; Krüger, K.; Niess, A.M.; Steinacker, J.M.; et al. Effects of Acute Endurance Exercise on Plasma Protein Profiles of Endurance-Trained and Untrained Individuals over Time. Mediators Inflamm. 2016, 2016, 4851935. [Google Scholar] [CrossRef]
- Barron, A.M.; Tokunaga, M.; Zhang, M.-R.; Ji, B.; Suhara, T.; Higuchi, M. Assessment of neuroinflammation in a mouse model of obesity and β-amyloidosis using PET. J. Neuroinflamm. 2016, 13, 221. [Google Scholar] [CrossRef]
- Prendergast, C.T.; Anderton, S.M. Immune cell entry to central nervous system—Current understanding and prospective therapeutic targets. Endocr. Metab. Immune Disord. Drug Targets 2009, 9, 315–327. [Google Scholar] [CrossRef]
- Mojsilovic-Petrovic, J.; Callaghan, D.; Cui, H.; Dean, C.; Stanimirovic, D.B.; Zhang, W. Hypoxia-inducible factor-1 (HIF-1) is involved in the regulation of hypoxia-stimulated expression of monocyte chemoattractant protein-1 (MCP-1/CCL2) and MCP-5 (Ccl12) in astrocytes. J. Neuroinflamm. 2007, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.-S.; Peng, J.; Murugan, M.; Feng, L.-J.; Liu, J.-L.; Eyo, U.B.; Zhou, L.-J.; Mogilevsky, R.; Wang, W.; Wu, L.-J. Chemokine CCL2–CCR2 Signaling Induces Neuronal Cell Death via STAT3 Activation and IL-1β Production after Status Epilepticus. J. Neurosci. 2017, 37, 7878–7892. [Google Scholar] [CrossRef] [PubMed]
- Palazon, A.; Tyrakis, P.A.; Macias, D.; Veliça, P.; Rundqvist, H.; Fitzpatrick, S.; Vojnovic, N.; Phan, A.T.; Loman, N.; Hedenfalk, I.; et al. An HIF-1α/VEGF-A Axis in Cytotoxic T Cells Regulates Tumor Progression. Cancer Cell 2017, 32, 669–683.e5. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Babapoor-Farrokhran, S.; Rodrigues, M.; Deshpande, M.; Puchner, B.; Kashiwabuchi, F.; Hassan, S.J.; Asnaghi, L.; Handa, J.T.; Merbs, S.; et al. Hypoxia-inducible factor 1 upregulation of both VEGF and ANGPTL4 is required to promote the angiogenic phenotype in uveal melanoma. Oncotarget 2016, 7, 7816–7828. [Google Scholar] [CrossRef]
- Hassan, A.; Arnold, B.M.; Caine, S.; Toosi, B.M.; Verge, V.M.K.; Muir, G.D. Acute intermittent hypoxia and rehabilitative training following cervical spinal injury alters neuronal hypoxia- and plasticity-associated protein expression. PLoS ONE 2018, 13, e0197486. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, M.; Tokai, H.; Takehara, Y.; Haraguchi, Y.; Asada, A.; Utsumi, K.; Inoue, M. Interrelation between oxygen tension and nitric oxide in the respiratory system. Am. J. Respir. Crit. Care Med. 2000, 162, 1257–1261. [Google Scholar] [CrossRef]
- Westendorf, A.M.; Skibbe, K.; Adamczyk, A.; Buer, J.; Geffers, R.; Hansen, W.; Pastille, E.; Jendrossek, V. Hypoxia Enhances Immunosuppression by Inhibiting CD4+ Effector T Cell Function and Promoting Treg Activity. Cell. Physiol. Biochem. 2017, 41, 1271–1284. [Google Scholar] [CrossRef]
- Hatfield, S.; Veszeleiova, K.; Steingold, J.; Sethuraman, J.; Sitkovsky, M. Mechanistic Justifications of Systemic Therapeutic Oxygenation of Tumors to Weaken the Hypoxia Inducible Factor 1α-Mediated Immunosuppression. Adv. Exp. Med. Biol. 2019, 1136, 113–121. [Google Scholar]
- Schäfer, S.T.; Gessner, S.; Scherag, A.; Rump, K.; Frey, U.H.; Siffert, W.; Westendorf, A.M.; Steinmann, J.; Peters, J.; Adamzik, M. Hydrocortisone Fails to Abolish NF-κB1 Protein Nuclear Translocation in Deletion Allele Carriers of the NFKB1 Promoter Polymorphism (-94ins/delATTG) and Is Associated with Increased 30-Day Mortality in Septic Shock. PLoS ONE 2014, 9, e104953. [Google Scholar] [CrossRef]
- Fenn, W.O.; Rahn, H.; Otis, A.B. A theoretical study of the composition of the alveolar air at altitude. Am. J. Physiol. 1946, 146, 637–653. [Google Scholar] [CrossRef] [PubMed]
- Richard, N.A.; Koehle, M.S. Differences in cardio-ventilatory responses to hypobaric and normobaric hypoxia: A review. Aviat. Space Environ. Med. 2012, 83, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Van der Heide, V.; Möhnle, P.; Rink, J.; Briegel, J.; Kreth, S. Down-regulation of MicroRNA-31 in CD4+ T Cells Contributes to Immunosuppression in Human Sepsis by Promoting TH2 Skewing. Anesthesiology 2016, 124, 908–922. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, S.T.; Franken, L.; Adamzik, M.; Schumak, B.; Scherag, A.; Engler, A.; Schönborn, N.; Walden, J.; Koch, S.; Baba, H.A.; et al. Mitochondrial DNA: An Endogenous Trigger for Immune Paralysis. Anesthesiology 2016, 124, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef]
- Chavakis, E.; Choi, E.Y.; Chavakis, T. Novel aspects in the regulation of the leukocyte adhesion cascade. Thromb. Haemost. 2009, 102, 191–197. [Google Scholar] [CrossRef]
- Martin, C.; Burdon, P.C.E.; Bridger, G.; Gutierrez-Ramos, J.C.; Williams, T.J.; Rankin, S.M. Chemokines acting via CXCR2 and CXCR4 control the release of neutrophils from the bone marrow and their return following senescence. Immunity 2003, 19, 583–593. [Google Scholar] [CrossRef]
- Massena, S.; Christoffersson, G.; Vågesjö, E.; Seignez, C.; Gustafsson, K.; Binet, F.; Herrera Hidalgo, C.; Giraud, A.; Lomei, J.; Weström, S.; et al. Identification and characterization of VEGF-A-responsive neutrophils expressing CD49d, VEGFR1, and CXCR4 in mice and humans. Blood 2015, 126, 2016–2026. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volunteer Characteristics | N = 11 |
|---|---|
| Gender, female/male (numbers; %) | 5/6 (45/55) |
| Age (Years; MV ± SD) | 36.4 ± 7 |
| Height (cm; MV ± SD) | 178 ± 6 |
| Bodyweight (kg; MV ± SD) | 72.7 ± 10 |
| Body Mass Index (kg/m2; MV ± SD) | 22.7 ± 2 |
| Heart rate (min−1; MV ± SD) | 61 ± 9 |
| Peripheral oxygen saturation | 97 ± 1 |
| (SpO2; %; MV ± SD) |
| Genes | Primer Sequences, UPL Probes and Assays |
|---|---|
| TBP forward | 5′ GAACATCATGGATCAGAACAACA 3′ |
| TBP reverse | 5′ ATAGGGATTCCGGGAGTCAT 3′ |
| UPL Probe Nr. 87 | |
| SDHA forward | 5′ GAGGCAGGGTTTAATACAGCA 3′ |
| SDHA reverse | 5′ CCAGTTGTCCTCCTCCATGT 3′ |
| UPL Probe Nr. 132 | |
| CCR-2 forward | 5′ TGAGACAAGCCACAAGCTGA 3′ |
| CCR-2 reverse | 5′ TTCTGATAAACCGAGAACGAGAT 3′ |
| UPL Probe Nr. 56 | |
| IL-1β forward | 5′ GAGGCACAAGGCACAACAG 3′ |
| IL-1β reverse | 5′ CCATGGCTGCTTCAGACAC 3′ |
| UPL Probe Nr. 41 | |
| CXCR-4 | Assay ID 110817 |
| VEGF-A | Assay ID 140396 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kammerer, T.; Faihs, V.; Hulde, N.; Stangl, M.; Brettner, F.; Rehm, M.; Horstmann, M.; Kröpfl, J.; Spengler, C.; Kreth, S.; et al. Hypoxic-Inflammatory Responses under Acute Hypoxia: In Vitro Experiments and Prospective Observational Expedition Trial. Int. J. Mol. Sci. 2020, 21, 1034. https://doi.org/10.3390/ijms21031034
Kammerer T, Faihs V, Hulde N, Stangl M, Brettner F, Rehm M, Horstmann M, Kröpfl J, Spengler C, Kreth S, et al. Hypoxic-Inflammatory Responses under Acute Hypoxia: In Vitro Experiments and Prospective Observational Expedition Trial. International Journal of Molecular Sciences. 2020; 21(3):1034. https://doi.org/10.3390/ijms21031034
Chicago/Turabian StyleKammerer, Tobias, Valentina Faihs, Nikolai Hulde, Manfred Stangl, Florian Brettner, Markus Rehm, Mareike Horstmann, Julia Kröpfl, Christina Spengler, Simone Kreth, and et al. 2020. "Hypoxic-Inflammatory Responses under Acute Hypoxia: In Vitro Experiments and Prospective Observational Expedition Trial" International Journal of Molecular Sciences 21, no. 3: 1034. https://doi.org/10.3390/ijms21031034
APA StyleKammerer, T., Faihs, V., Hulde, N., Stangl, M., Brettner, F., Rehm, M., Horstmann, M., Kröpfl, J., Spengler, C., Kreth, S., & Schäfer, S. (2020). Hypoxic-Inflammatory Responses under Acute Hypoxia: In Vitro Experiments and Prospective Observational Expedition Trial. International Journal of Molecular Sciences, 21(3), 1034. https://doi.org/10.3390/ijms21031034