Effects of Cold Jet Atmospheric Pressure Plasma on the Structural Characteristics and Immunoreactivity of Celiac-Toxic Peptides and Wheat Storage Proteins

 ,
, 


Abstract
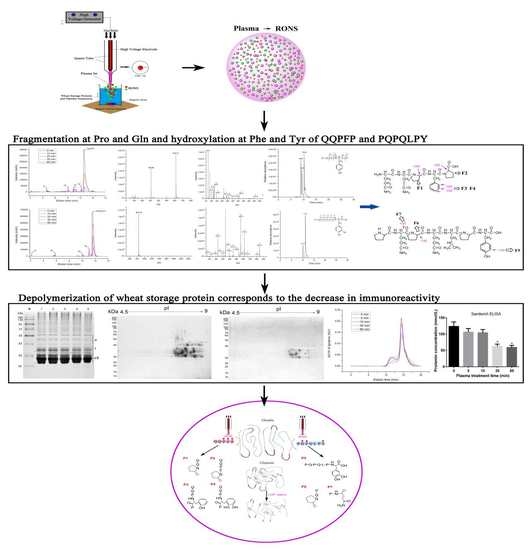
1. Introduction
2. Results
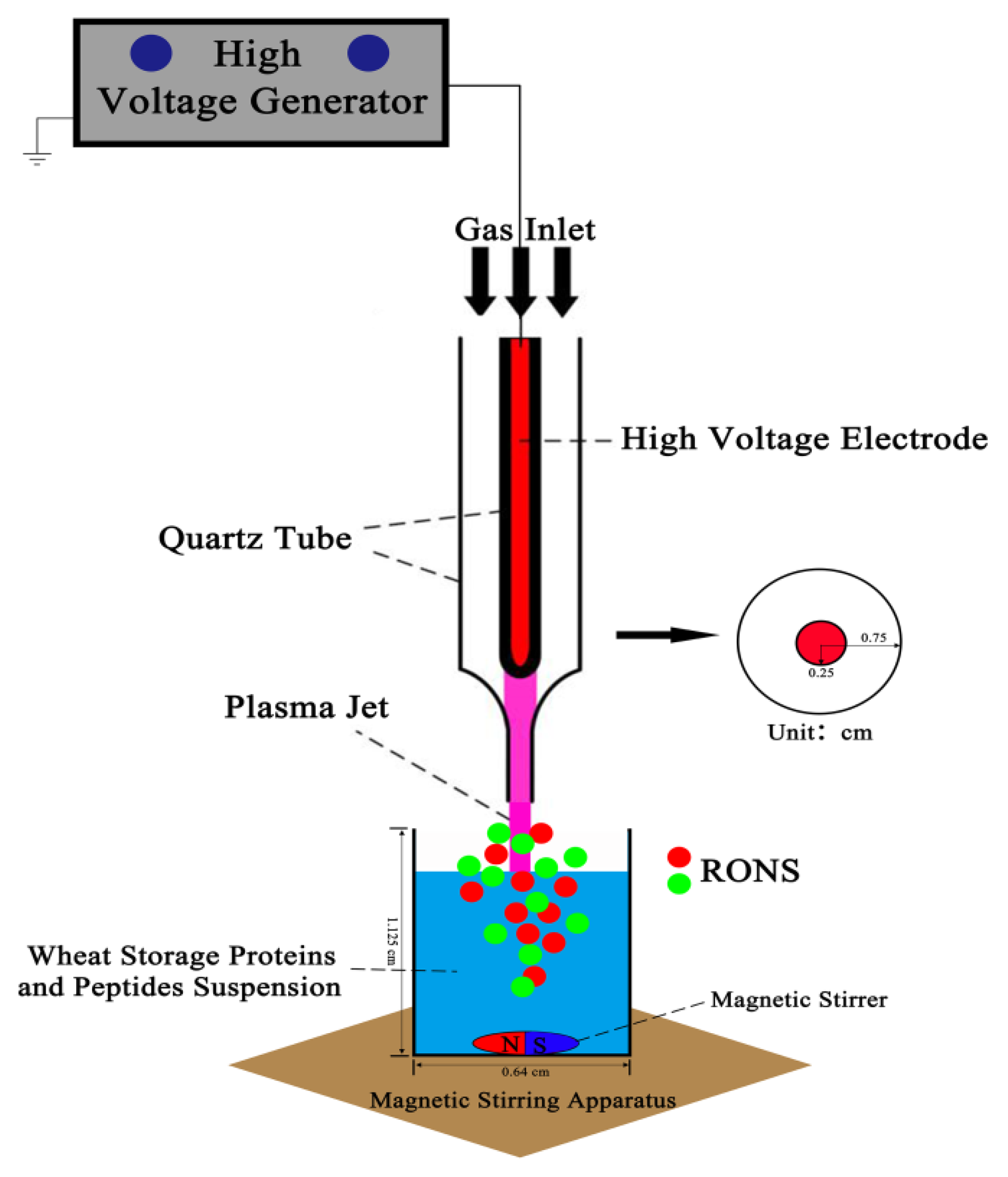
2.1. Experiment Setup, Discharge, Current and Optical Emission Spectrum (OES) Measurements
2.2. Generation of Several RONS Induced by CJAP Plasma
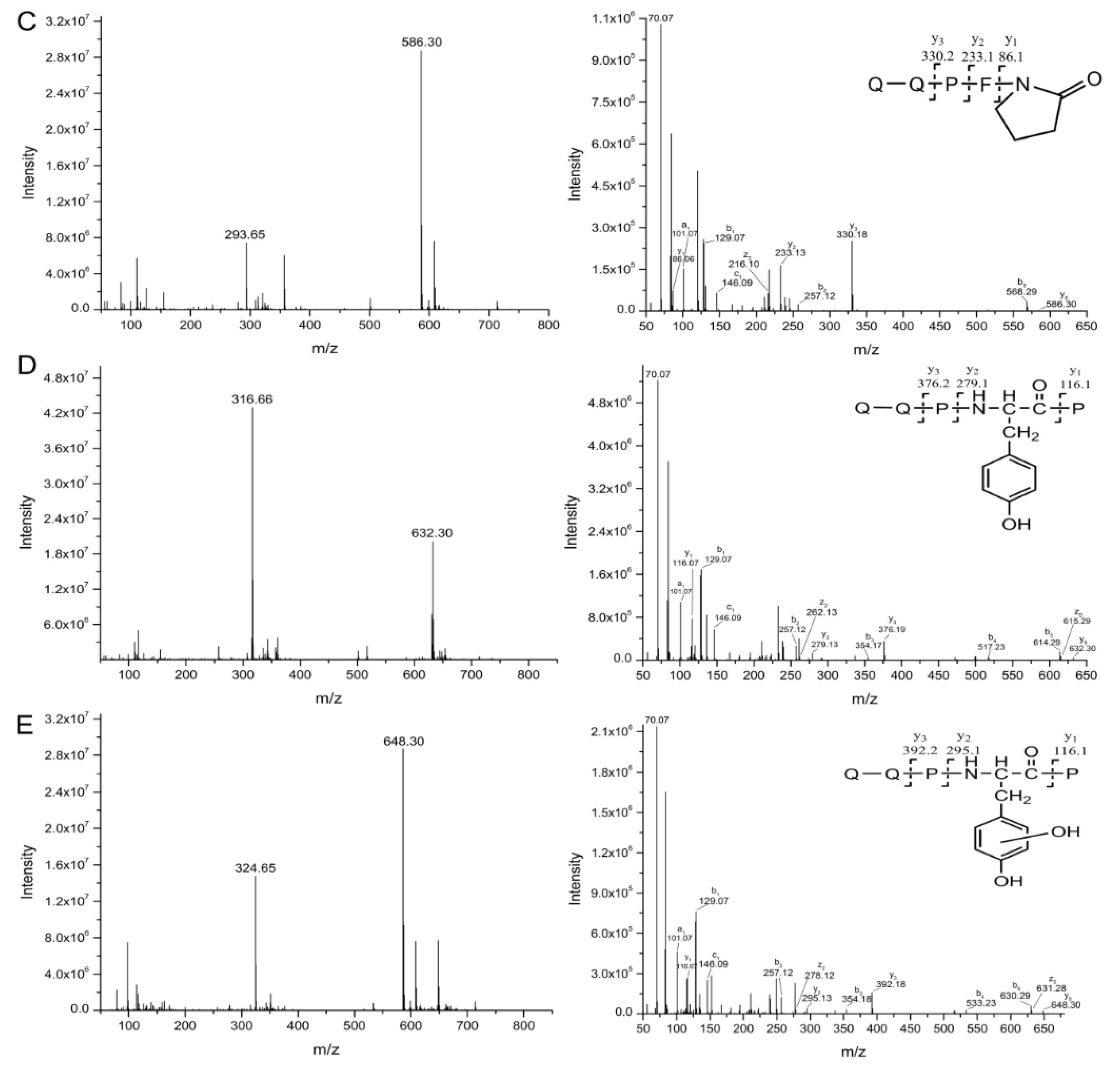
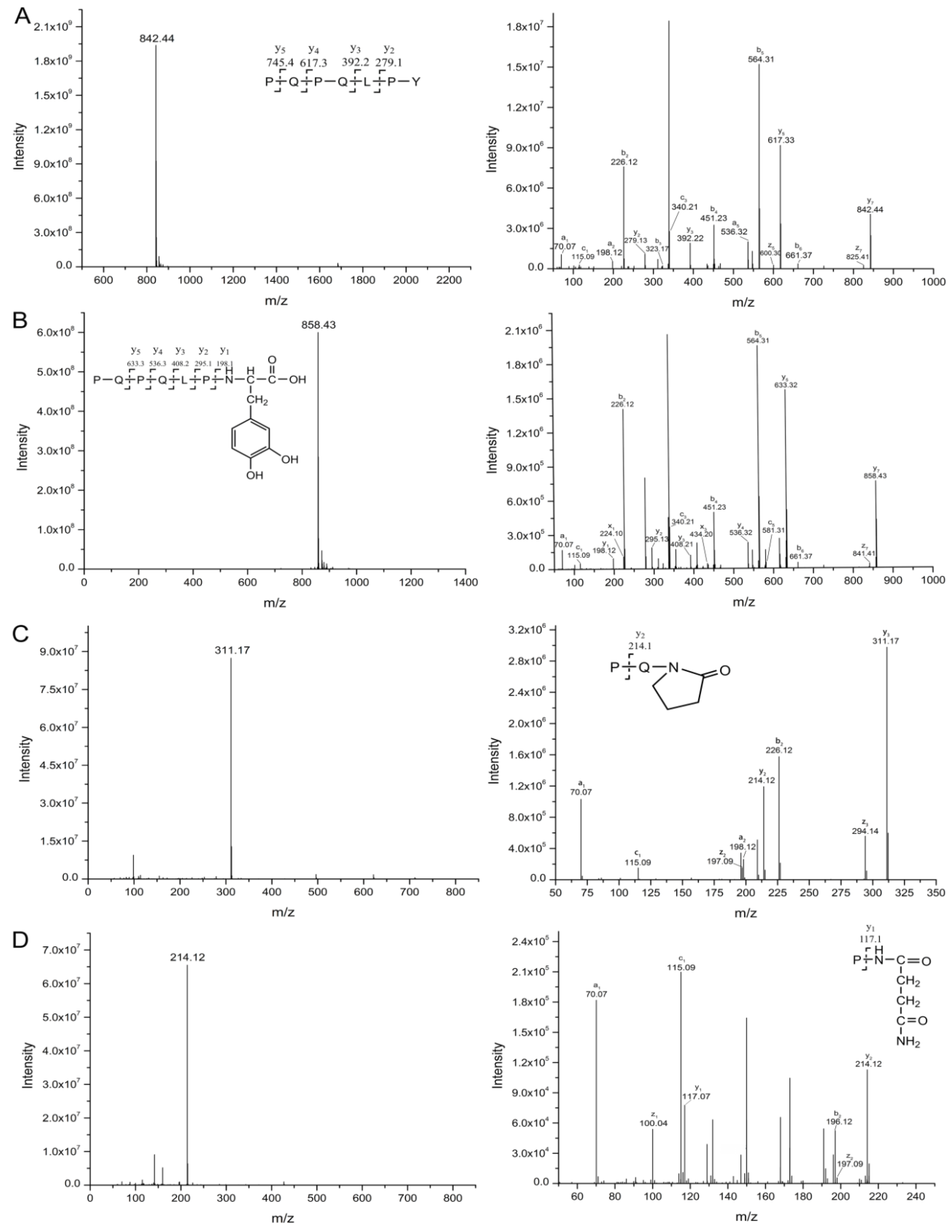
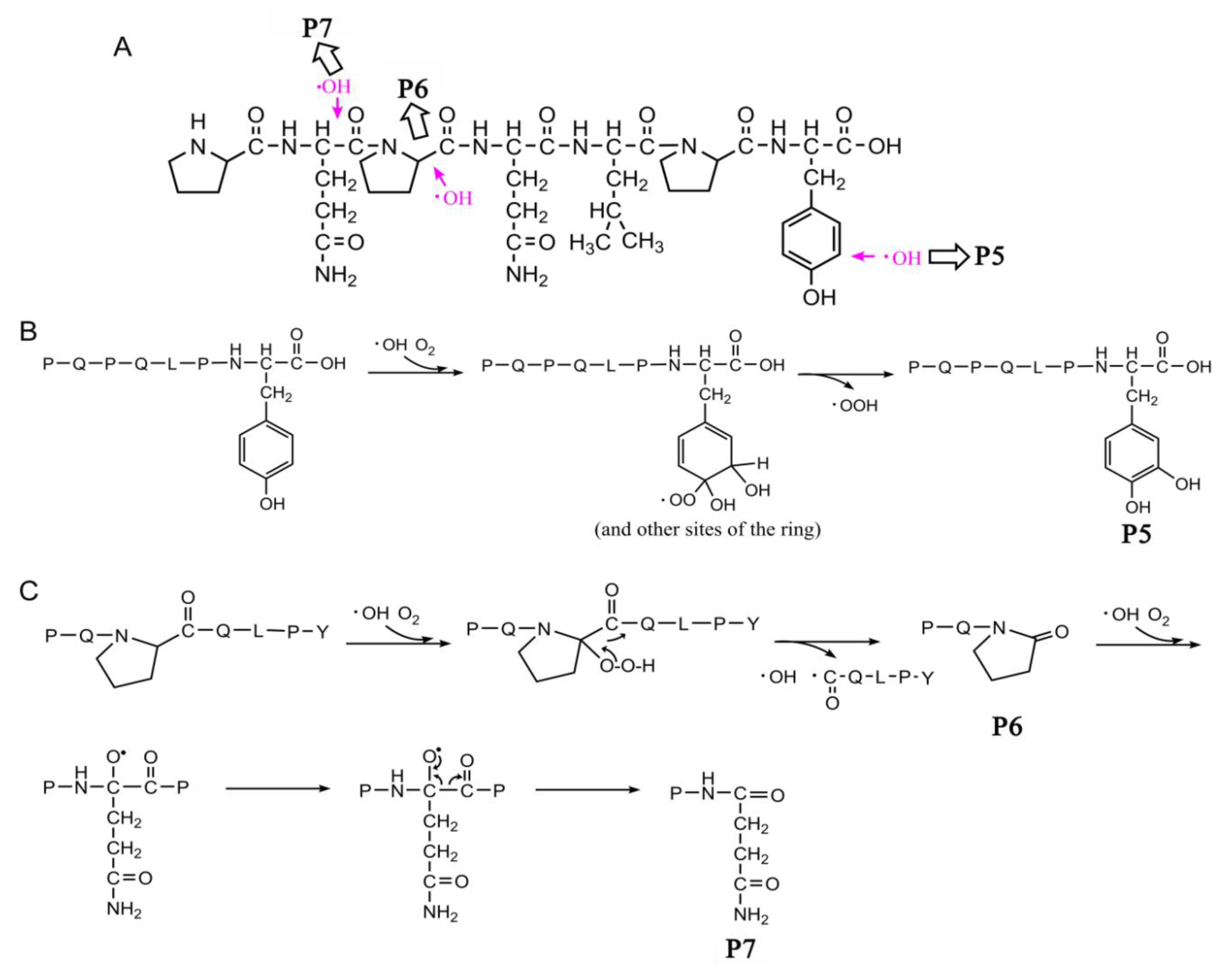
2.3. Oxidation and Hydroxylation Gives Rise to P1~P7 after CJAP Plasma Treatment
2.4. Modification of Specific Pro, Gln, Phe, and Tyr Residues Leads to Degradation of the Model Peptides
2.5. Agglomeration State of Wheat Gliadins Analyzed by SDS-PAGE
2.6. Decrease in Quantity of Wheat Gliadins Treated with CJAP Plasma
2.7. LC-MS Identification of Proteins in 2-DE Spots
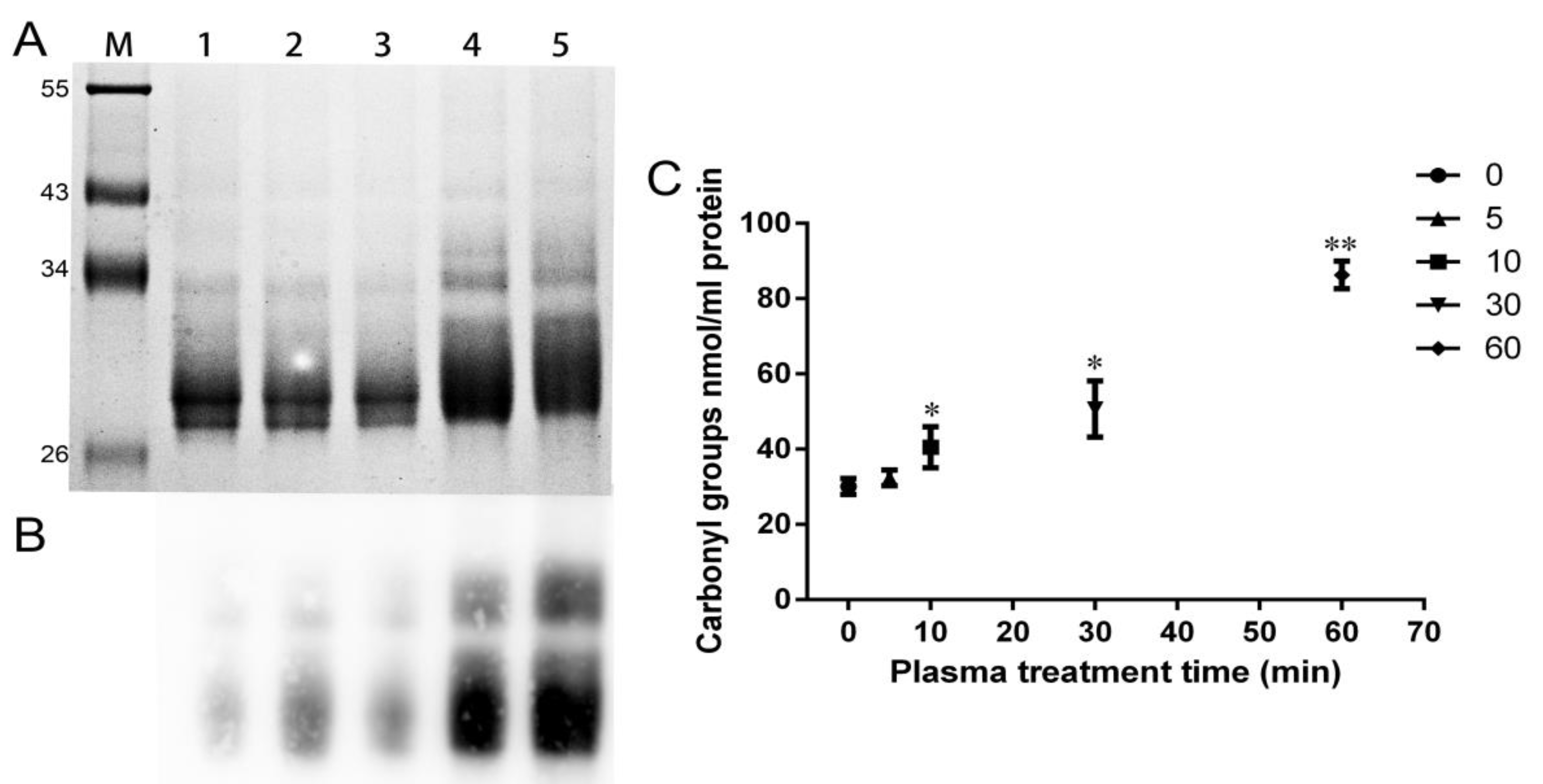
2.8. Increment of Carbonyl Groups in Plasma-Treated Wheat Gliadins
2.9. Immunological Activity Evaluation of Wheat Gliadins in ELISA
2.10. Size Distribution Change Analysis of Gluten Proteins before and after CJAP Plasma Treatment
3. Discussion
3.1. CJAP Plasma can Generate a Variety of RONS
3.2. CJAP Plasma-Induced Oxidation and Hydroxylation at Specific Pro, Gln, Phe, and Tyr Residues Lead to Reduction and Modification of Celiac-Toxic Peptides
3.3. Decreased Immunoreactivity of Wheat Gliadins Modified by the CJAP Plasma
3.4. Decrease in Large-Sized Polymers through Longer Exposure to CJAP Plasma
3.5. Proposed Mechanism of the Model Peptide and Wheat Storage Protein Modified by CJAP Plasma
4. Materials and Methods
4.1. Materials
4.2. Discharge, Current and Optical Emission Spectra (OES) Detection
4.3. Extraction of Gliadins from Wheat
4.4. O3 Measurement
4.5. H2O2 and pH Measurement
4.6. •OH Measurement
4.7. Total Content of NO2− and NO3− Determination
4.8. Identification of CJAP Plasma Treatment Fragments by Orbitrap HR-LC-MS/MS
4.9. Quantitative 2-DE and LC-MS Analysis
4.10. Qualitative and Quantitative Detection of Carbonyl Groups
4.11. Sandwich Enzyme-Linked Immunosorbent Assay (ELISA)
4.12. SE-HPLC Analysis
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CJAP | cold jet atmospheric pressure plasma |
| RONS | reactive oxygen and nitrogen species |
| CD | celiac disease |
| QQPFP | Gln-Gln-Pro-Phe-Pro |
| PQPQLPY | Pro-Gln-Pro-Gln-Leu-Pro-Tyr |
| MW | molecular weight |
| kDa | kiloDalton |
| SDS | Sodium dodecyl sulfate |
| DNPH 2 | 4-dinitrophenyl-hydrazine |
| EDTA | ethylenediamine-tetraacetic acid |
| DTT | dithiothreitol |
| AC | alternating current |
| OES | optical emission spectra |
| H.V. | high-voltage |
| DPD | N,N-diethyl-p-phenyl-enediamine |
| TA | terephthalic acid |
| HR-LC-MS/MS | high resolution-liquid chromatography-mass spectrometry/mass spectrometry |
| EIC | extract ion chromatograms |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| 2-DE | two dimensional electrophoresis |
| IEF | isoelectric focusing |
| ELISA | sandwich enzyme-linked immunosorbent assay |
| SE-HPLC | size exclusion high performance liquid phase chromatography |
| HMW | high molecular weight; LMW, low-molecular-weight |
References
- Shewry, P. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef] [PubMed]
- Valerii, M.; Ricci, C.; Spisni, E.; Silvestro, R.; Fazio, L.; Cavazza, E.; Lanzini, A.; Campieri, M.; Dalpiaz, A.; Pavan, B. Responses of peripheral blood mononucleated cells from non-celiac gluten sensitive patients to various cereal sources. Food Chem. 2015, 176, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-León, S.; Gil-Humanes, J.; Ozuna, C.; Giménez, M.; Sousa, C.; Voytas, D.; Barro, F. Low-gluten, non-transgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 2017, 16, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Gianfrani, C.; Siciliano, R.; Facchiano, A.; Camarca, A.; Mazzeo, M.; Costantini, S.; Salvati, V.; Maurano, F.; Mazzarella, G.; Iaquinto, G.; et al. Transamidation of wheat flour inhibits the response to gliadin of intestinal T cells in celiac disease. Gastroenterology 2007, 133, 780–789. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Sontag-Strohm, T.; Stoddard, F.; Kato, Y. Oxidation of proline decreases immunoreactivity and alters structure of barley prolamin. Food Chem. 2017, 214, 597–605. [Google Scholar] [CrossRef]
- Wongsagonsup, R.; Deeyai, P.; Chaiwat, W.; Horrungsiwat, S.; Leejariensuk, K.; Suphantharika, M.; Fuongfuchat, A.; Dangtip, S. Modification of tapioca starch by non-chemical route using jet atmospheric argon plasma. Carbohydr. Polym. 2014, 102, 790–798. [Google Scholar] [CrossRef]
- Cuadrado, C.; Cabanillas, B.; Pedrosa, M.M.; Varela, A.; Guillamon, E.; Muzquiz, M.; Crespo, J.; Rodriguez, J.; Burbano, C. Influence of thermal processing on IgE reactivity to lentil and chickpea proteins. Mol. Nutr. Food Res. 2010, 53, 1462–1468. [Google Scholar] [CrossRef]
- Sandhu, H.; Manthey, F.; Simsek, S. Ozone gas affects physical and chemical properties of wheat (Triticum aestivum L.) starch. Carbohydr. Polym. 2012, 87, 1261–1268. [Google Scholar] [CrossRef]
- Misra, N.; Kaur, S.; Tiwari, B.; Kaur, A.; Singh, N.; Cullen, P. Atmospheric pressure cold plasma (ACP) treatment of wheat flour. Food Hydrocoll. 2015, 44, 115–121. [Google Scholar] [CrossRef]
- Bogaerts, A.; Neyts, E.; Gijbels, R.; Van der, J. Gas discharge plasmas and their applications. Spectrochim. Acta Part B At. Spectrosc. 2002, 57, 609–658. [Google Scholar] [CrossRef]
- Ikawa, S.; Kitano, K.; Hamaguchi, S. Effects of pH on bacterial inactivation in aqueous solutions due to low-temperature atmospheric pressure plasma application. Plasma Process. Polym. 2010, 7, 33–42. [Google Scholar] [CrossRef]
- Keidar, M.; Shashurin, A.; Volotskova, O.; Ann, M.; Srinivasan, P.; Sandler, A.; Trink, B. Cold atmospheric plasma in cancer therapy. Phys. Plasmas 2013, 20, 057101. [Google Scholar] [CrossRef]
- Lee, H.; Kim, G.; Kim, J.; Park, J.; Lee, J.; Kim, G. Tooth bleaching with nonthermal atmospheric pressure plasma. J. Endod. 2009, 35, 587–591. [Google Scholar] [CrossRef]
- Heinlin, J.; Isbary, G.; Stolz, W.; Morfill, G.; Landthaler, M.; Shimizu, T.; Steffes, B.; Nosenko, T.; Zimmermann, J.; Karrer, S. Plasma applications in medicine with a special focus on dermatology. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 1–11. [Google Scholar] [CrossRef]
- Osman, A.; Uhlig, H.; Valdes, I.; Amin, M.; Méndez, E.; Mothes, T. A monoclonal antibody that recognizes a potential coeliac-toxic repetitive pentapeptide epitope in gliadins. Eur. J. Gastroenterol. Hepatol. 2001, 13, 1189–1193. [Google Scholar] [CrossRef]
- Arentz–Hansen, H.; Mcadam, S.; Molberg, Ø.; Fleckenstein, B.; Lundin, K.; Jørgensen, T.; Jung, G.; Roepstorff, P.; Sollid, L. Celiac lesion T cells recognize epitopes that cluster in regions of gliadins rich in proline residues. Gastroenterology 2002, 123, 803–809. [Google Scholar] [CrossRef]
- Morón, B.; Cebolla, A.; Manyani, H.; Alvarez-Maqueda, M.; Megías, M.; Thomas, M.; López, M.; Sousa, C. Sensitive detection of cereal fractions that are toxic to celiac disease patients by using monoclonal antibodies to a main immunogenic wheat peptide. Am. J. Clin. Nutr. 2008, 87, 405–414. [Google Scholar] [CrossRef]
- Vader, L.; Stepniak, D.; Bunnik, E.; Kooy, Y.; Haan, W.; Drijfhout, J.; Veelen, P.; Koning, F. Characterization of cereal toxicity for celiac disease patients based on protein homology in grains. Gastroenterology 2003, 125, 1105–1113. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, D.; Liu, D.; Cui, Q.; Cai, H.; Li, Q.; Chen, H.; Kong, M. Production of simplex RNS and ROS by nanosecond pulse N2/O2 plasma jets with homogeneous shielding gas for inducing myeloma cell apoptosis. J. Phys. D Appl. Phys. 2017, 50, 195204. [Google Scholar] [CrossRef]
- Shad, A.; Li, C.; Zuo, J.; Liu, J.; Dar, A.; Wang, Z. Understanding the ozonated degradation of sulfadimethoxine, exploration of reaction site, and classification of degradation products. Chemosphere 2018, 212, 228–236. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Yang, H.; Wang, X.; Xie, Y. Oxidation of isoprothiolane by ozone and chlorine: Reaction kinetics and mechanism. Chemosphere 2019, 232, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, B.; Sornsakdanuphap, J.; Hong, Y.; Uhm, H.; Weltmann, K.; Choi, E. The effect of the gap distance between an atmospheric-pressure plasma jet nozzle and liquid surface on OH and N2 species concentrations. Phys. Plasmas 2017, 24, 073502. [Google Scholar] [CrossRef]
- Liu, F.; Lai, S.; Tong, H.; Lakey, P.; Shiraiwa, M.; Weller, M.; Pöschl, U.; Kampf, C. Release of free amino acids upon oxidation of peptides and proteins by hydroxyl radicals. Anal. Bioanal. Chem. 2017, 409, 2411–2420. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Kanerva, P.; Salovaara, H.; Sontag-Strohm, T. Degradation of C-hordein by metal-catalysed oxidation. Food Chem. 2016, 196, 1256–1263. [Google Scholar] [CrossRef] [PubMed]
- Bruggeman, P.; Schram, D. On OH production in water containing atmospheric pressure plasmas. Plasma Sources Sci. Technol. 2010, 19, 045025. [Google Scholar] [CrossRef]
- Pei, X.; Lu, Y.; Wu, S.; Xiong, Q.; Lu, X. A study on the temporally and spatially resolved OH radical distribution of a room-temperature atmospheric-pressure plasma jet by laser-induced fluorescence imaging. Plasma Sources Sci. Technol. 2013, 22, 025023. [Google Scholar] [CrossRef]
- Capitelli, M.; Ferreira, C.; Gordiets, B.; Osipov, A. Plasma kinetics in atmospheric gases. Vacuum 2001, 62, 388–389. [Google Scholar]
- Liu, D.; Liu, Z.; Chen, C.; Yang, A.; Li, D.; Rong, M.; Chen, H.; Kong, M. Aqueous reactive species induced by a surface air discharge: Heterogeneous mass transfer and liquid chemistry pathways. Sci. Rep. 2016, 6, 23737. [Google Scholar] [CrossRef]
- Zangouei, M.; Haynes, B. The Role of Atomic Oxygen and Ozone in the Plasma and Post-plasma Catalytic Removal of N2O. Plasma Chem. Plasma Process. 2019, 39, 89–108. [Google Scholar] [CrossRef]
- Xu, G.; Chance, M. Hydroxyl radical-mediated modification of proteins as probes for structural proteomics. Chem. Rev. 2007, 107, 3514–3543. [Google Scholar] [CrossRef]
- Jones, B.; Vergne, M.; Bunk, D.; Locascio, L.; Hayes, M. Cleavage of peptides and proteins using light-generated radicals from titanium dioxide. Anal. Chem. 2007, 79, 1327–1332. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Uchida, K.; Kawakishi, S. Oxidative fragmentation of collagen and prolyl peptide by Cu (II)/H2O2. Conversion of proline residue to 2-pyrrolidone. J. Biol. Chem. 1992, 267, 23646–23651. [Google Scholar] [PubMed]
- Uchida, K.; Kato, Y.; Kawakishi, S. A novel mechanism for oxidative cleavage of prolyl peptides induced by the hydroxyl radical. Biochem. Biophys. Res. Commun. 1990, 169, 265–271. [Google Scholar] [CrossRef]
- Stadtman, E.; Levine, R. Free radical-mediated oxidation of free amino acids and amino acid residues in proteins. Amino Acids 2003, 25, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Morgan, P.; Pattison, D.; Davies, M. Quantification of hydroxyl radical-derived oxidation products in peptides containing glycine, alanine, valine, and proline. Free Radic. Biol. Med. 2012, 52, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Broeck, H.; Cordewener, J.; Nessen, M.; America, A.; Meer, I. Label free targeted detection and quantification of celiac disease immunogenic epitopes by mass spectrometry. J. Chromatogr. A 2015, 1391, 60–71. [Google Scholar] [CrossRef]
- Zhou, L.; Kooy-Winkelaar, Y.M.C.; Cordfunke, R.A.; Dragan, I.; Thompson, A.; Drijfhout, J.W.; Van Veelen, P.A.; Chen, H.; Koning, F. Abrogation of immunogenic properties of gliadin peptides through transamidation by microbial transglutaminase is acyl-acceptor dependent. J. Agric. Food Chem. 2017, 65, 7542–7552. [Google Scholar] [CrossRef]
- Peter, H.; Benjamin, L.; Ruby, G. Celiac disease. J. Allergy Clin. Immunol. 2015, 135, 1099–1106. [Google Scholar]
- Fatma, B.; Barbara, P.; Andrea, F.; Stefano, S. A complete mass spectrometry (MS)-based peptidomic description of gluten peptides generated during in vitro gastrointestinal digestion of durum wheat: Implication for celiac disease. J. Am. Soc. Mass Spectrom. 2019, 30, 1481–1490. [Google Scholar]
- Shi, H.; Cooper, B.; Stroshine, R.; Ileleji, K.; Keener, K. Structures of degradation products and degradation pathways of aflatoxin B1 by high-voltage atmospheric cold plasma (HVACP) treatment. J. Agric. Food Chem. 2017, 65, 6222–6230. [Google Scholar] [CrossRef]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–1193. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.; Tatham, A. The prolamin storage proteins of cereal seeds: Structure and evolution. Biochem. J. 1990, 267, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nagib, A.; Tifenn, D.; Mohammad-Zaman, N.; Setsuko, K. Tissue-specific defense and thermo-adaptive mechanisms of soybean seedlings under heat stress revealed by proteomic approach. J. Proteome Res. 2010, 9, 4189–4204. [Google Scholar]
- Martinez-Esteso, M.J.; NøRgaard, J.; Brohée, M.; Haraszi, R.; Maquet, A.; O’Connor, G. Defining the wheat gluten peptide fingerprint via a discovery and targeted proteomics approach. J. Proteom. 2016, 147, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, Y.; Ma, F.; Zeng, J.; Chang, J.; Chen, M.; Li, K.; Yang, G.; Wang, Y.; He, G. Effect of extra cysteine residue of new mutant 1Ax1 subunit on the functional properties of common wheat. Sci. Rep. 2017, 7, 7510. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot | G1 | G2 | G3 |
|---|---|---|---|
| Predominant protein | Gamma gliadin | Gamma gliadin | Gamma gliadin |
| Accession number | F2XAR5 | Q30DX7 | K7WVC4 |
| MW | 37,564 | 34,502 | 33,067 |
| pI | 8.51 | 8.57 | 8.88 |
| Protein score | 94 | 1762 | 2831 |
| Protein match | 2 | 91 (54) | 62 (52) |
| Protein coverage | - | 25.5 | 15.8 |
| Sequence | R.APFASIVAGIGGQ. (ppm: 5.18) R.SLVLQTLPSMCSVYVPPECSIMR.A (ppm: −44.56) | R.TTTSVPFGVGTGVGAY. (ppm: 8.86) R.ILPTMCSVNVPLYR.T (ppm: 6.64) R.ILPTMCSVNVPLYR.T (ppm: 6.85) K.VFLQQQCSPVAMPQR.L (ppm: 9.19) R.SQMLQQSSCHVMQQQCCQQLPQIPQQSR.Y (ppm: 8.80) | R.SDCQVMQQQCCQQLAQIPR.Q (ppm: 6.96) R.SDCQVMQQQCCQQLAQIPR.Q (ppm: 0.35) R.QPQQPFYQQPQQTFPQPQQAFPHQPK.Q (ppm: 5.81) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, F.; Xie, X.; Zhang, Y.; Duan, J.; Ma, M.; Wang, Y.; Qiu, D.; Lu, X.; Yang, G.; He, G. Effects of Cold Jet Atmospheric Pressure Plasma on the Structural Characteristics and Immunoreactivity of Celiac-Toxic Peptides and Wheat Storage Proteins. Int. J. Mol. Sci. 2020, 21, 1012. https://doi.org/10.3390/ijms21031012
Sun F, Xie X, Zhang Y, Duan J, Ma M, Wang Y, Qiu D, Lu X, Yang G, He G. Effects of Cold Jet Atmospheric Pressure Plasma on the Structural Characteristics and Immunoreactivity of Celiac-Toxic Peptides and Wheat Storage Proteins. International Journal of Molecular Sciences. 2020; 21(3):1012. https://doi.org/10.3390/ijms21031012
Chicago/Turabian StyleSun, Fusheng, Xiaoxue Xie, Yufan Zhang, Jiangwei Duan, Mingyu Ma, Yaqiong Wang, Ding Qiu, Xinpei Lu, Guangxiao Yang, and Guangyuan He. 2020. "Effects of Cold Jet Atmospheric Pressure Plasma on the Structural Characteristics and Immunoreactivity of Celiac-Toxic Peptides and Wheat Storage Proteins" International Journal of Molecular Sciences 21, no. 3: 1012. https://doi.org/10.3390/ijms21031012
APA StyleSun, F., Xie, X., Zhang, Y., Duan, J., Ma, M., Wang, Y., Qiu, D., Lu, X., Yang, G., & He, G. (2020). Effects of Cold Jet Atmospheric Pressure Plasma on the Structural Characteristics and Immunoreactivity of Celiac-Toxic Peptides and Wheat Storage Proteins. International Journal of Molecular Sciences, 21(3), 1012. https://doi.org/10.3390/ijms21031012