Abstract
Endometriosis is a chronic, inflammatory, hormone-dependent disease characterized by histological lesions produced by the presence of endometrial tissue outside the uterine cavity. Despite the fact that an estimated 176 million women are affected worldwide by this gynecological disorder, risk factors that cause endometriosis have not been properly defined and current treatments are not efficient. Although the interaction between diet and human health has been the focus of many studies, little information about the correlation of foods and their bioactive derivates with endometriosis is available. In this framework, Brassica crops have emerged as potential candidates for ameliorating the chronic inflammatory condition of endometriosis, due to their abundant content of health-promoting compounds such as glucosinolates and their hydrolysis products, isothiocyanates. Several inflammation-related signaling pathways have been included among the known targets of isothiocyanates, but those involving aquaporin water channels have an important role in endometriosis. Therefore, the aim of this review is to highlight the promising effects of the phytochemicals present in Brassica spp. as major candidates for inclusion in a dietary approach aiming to improve the inflammatory condition of women affected with endometriosis. This review points out the potential roles of glucosinolates and isothiocyanates from Brassicas as anti-inflammatory compounds, which might contribute to a reduction in endometriosis symptoms. In view of these promising results, further investigation of the effect of glucosinolates on chronic inflammatory diseases, either as diet coadjuvants or as therapeutic molecules, should be performed. In addition, we highlight the involvement of aquaporins in the maintenance of immune homeostasis. In brief, glucosinolates and the modulation of cellular water by aquaporins could shed light on new approaches to improve the quality of life for women with endometriosis.
1. Introduction
Endometriosis is a frequently diagnosed, incurable, hormone-dependent gynecological disorder characterized by a chronic inflammatory profile and histological lesions generated by the abnormal growth of endometrial-like tissue outside the uterine cavity. These lesions mainly occur within the peritoneal cavity and are engrafted in different locations such as the peritoneum wall, ovaries, colon, and bladder, although it can also develop in distant organs, such as the liver, lung, and brain, among others [1,2]. Endometriosis may appear in diverse forms and locations, and thus it can be classified as peritoneal endometriosis; ovarian endometriosis; deep infiltrating endometriosis (DIE); or extragenital endometriosis [1,2].
It is estimated that approximately 176 million women worldwide are affected by endometriosis, which represents about 10% of women of reproductive age [2]. However, it is suspected that these data could underestimate the real number of affected women, since many of them are accidentally diagnosed after going into surgery for the treatment of other pathologies [1,2]. Women affected by endometriosis may present severe symptoms, which include, among others, chronic pelvic pain, dysmenorrhea, and dyspareunia [1,2,3,4]. In this context, 2–4% of women of reproductive age may suffer from sexual dysfunction due to the painful symptoms produced during intercourse [5]. Furthermore, pelvic pain can be incapacitating, especially in cases of DIE [6,7]. Endometriosis causes infertility in approximately 30% of affected women, likely because of the endometriotic lesion scars present in the reproductive organs [1,2,8]. Furthermore, it has been demonstrated that endometriosis is a disease with a high impact on health care costs, being comparable with other chronic diseases such as diabetes [9].
On the other hand, knowledge of the direct influence of diet on human health has grown quickly in recent years [10,11]. However, to date, no conclusive results relating a diet rich in vegetables with endometriosis development or an improvement in its symptoms have been found [12]. In this regard, Parazzini et al. [13] performed a case-control study with 504 women suffering from endometriosis. Food frequency questionnaires revealed that women with a higher intake of vegetables showed a lower endometriosis risk. Nevertheless, another case-control study based on dietary questionnaires found no association between a higher intake of vegetables and a decrease in endometriosis risk [14]. A more recent study showed that women who consumed more than one serving of cruciferous vegetables per day had a 13% higher risk of developing endometriosis compared with those who ingested them once a week or less [15]. However, these data came from retrospective studies based on self-reported questionnaires, while there is little or no information on the effect of a diet rich in Brassica spp. for women affected with endometriosis and even less information about the effect of the bioactive compounds present in cruciferous foods.
Therefore, in this review, we aim to point out the promising effect of the phytochemicals present in Brassica vegetables as key components of a diet intended to ameliorate the inflammatory condition of endometriosis patients. We bring together the connections between endometriosis and the scarce available information on the mechanistic effects of the biocomponents present in Brassica (glucosinolates and their conjugates: Isothiocyanates and thiols) in this pathology, also taking into account the role of aquaporins.
2. Etiology of Endometriosis
Endometriosis is considered a benign disease as it is not caused by transformed malignant cells. Although the mechanisms responsible for the origin and further progression of endometriosis have not been established, it has been proposed that endometriosis may be caused by an increased rate of cell proliferation and/or a decreased apoptotic rate. In this regard, the balance between cell proliferation and apoptosis is altered in the eutopic endometrium of endometriosis patients [16]. Additionally, it has been related to an elevated risk of developing ovarian cancer and, in fact, it shares common characteristics with neoplasia, including hyper-proliferative activity and cell growth, invasion of adjacent tissues, defective apoptotic ability, neoangiogenesis, and sustained local inflammatory responses [17,18,19,20,21].
The etiology of endometriosis is still unknown. The classical explanation for the development of endometriosis involves the escape of endometrial tissue fragments from the uterus into the peritoneal cavity by a retrograde menstruation mechanism, which allows the adhesion and growth of this tissue inside the abdominal cavity [22]. However, while retrograde menstruation is a common process, only 10% of women have endometriosis; therefore, multiple factors must be concomitantly acting to cause the disease. Recently, it has become widely accepted that the actual origin of endometriosis must have multiple causes. In this regard, different individual genetic and environmental predisposition factors, together with a deregulated and inefficient immune system, would allow the attachment and growth of endometrial ectopic tissue [23], which plays a key role in the onset and further progression of the disease. Other postulated hypotheses include (a) the occurrence of coelomic metaplasia, i.e., the peritoneal coelomic epithelium differentiates into endometrial-like cells; (b) neonatal spread of somatic stem cells and peritoneal implantation that remains latent until menarche; and (c) abnormal embryonic Müllerian ducts remains (from which the uterus, fallopian tubes, and upper vagina, develop) [23,24]. Furthermore, epidemiological and experimental evidence suggests that intrauterine exposure to high levels of endocrine-disrupting chemicals during the prenatal stage may play a role in the physiopathology of endometriosis in adulthood [25,26,27,28]. However, these theories have not conclusively demonstrated a unique causal association for all cases of endometriosis.
Furthermore, many epidemiological studies have explored the potential association of endometriosis with multiple risk factors. As recently reviewed by Parazzini et al., 2017 [13], Parasar et al., 2017 [29] and, Shafrir et al., 2018 [30], early age at menarche, taller height, lower adiposity, and low body mass index (BMI) [31,32], as well as infertility [33] have been associated with a higher risk of developing endometriosis, while a higher parity and higher BMI are inversely associated with endometriosis. Additionally, high exposure to environmental endocrine disrupting compounds (EDCs), such as phthalates, bisphenols, dioxins, polychlorinated biphenyls (PCBs), pesticides, and many others contaminants, has been associated with a higher endometriosis risk [34,35,36,37]. On the other hand, inconsistent results have been reported from studies associating endometriosis with several modifiable lifestyle risks factors, including smoking and frequent exercise [38,39,40], as well as caffeine or alcohol intake and level of education [29,41].
Current treatments are based on the estrogen dependence of this pathology [42,43]. However, those hormone-based treatments have negative effects on the associated infertility problem and lead to the induction of disorders similar to menopause, including the loss of bone density. All effects associated with this therapy, together with its low efficiency in many patients, have resulted in the administration of the drugs being stopped, allowing the progression of endometriotic lesions and adherences outside the uterus and increasing the ratio of repeated surgical invasive procedures [1,2,44]. Therefore, there is a great need to identify new molecular targets and potential therapies to facilitate the development of non-invasive techniques and/or to allow better treatment of this pathology.
3. Inflammatory Background of Endometriosis
The most characteristic histological features of endometriosis lesions are local inflammation and peritoneal fibrosis. The invasive capacity and resistance to apoptosis differ between the endometrium of women with and without endometriosis, as well as between ectopic and eutopic endometrium of patients with endometriosis. This suggests that the ectopic inflammatory environment plays an important role in the development of endometriotic lesions [1,2,24,45,46,47,48]. Furthermore, whilst retrograde migrated endometrial cells in healthy women are cleared by peritoneal immune cells, ectopic endometrial tissue in endometriosis patients escapes immune surveillance, survives, and is implanted in the peritoneum and other organs.
Macrophages are crucial cells that are involved not only in the immune response induced by infected, injured, or tumor stimuli but also in the promotion of the appropriate regulatory response necessary to restore tissue homeostasis [49]. Macrophages are the prevalent immune cell type in the peritoneal fluid of women with endometriosis and, consequently, they must play an important role in its pathophysiology [50,51]. There is accumulated evidence of an altered and dysfunctional activity of macrophages isolated from the ectopic endometrial tissue located in the peritoneal cavity [50,52]. In this regard, an M2 polarization profile, which corresponds to alternative activation of macrophages towards a tissue-reparative phenotype, would permit the survival, growth, and neovascularization of the ectopic endometriotic tissue [50,51,52,53,54,55,56]. Additionally, other studies have shown a classical M1 polarization profile of macrophage activation in the eutopic endometrium of women with endometriosis in comparison with corresponding healthy controls [57]. The knowledge of molecular mechanisms involved in the activation and polarization of macrophages is crucial to understand the pathophysiology of endometriosis and to allow the design of new therapies. In this regard, it is of great importance to use a proper method to obtain human peritoneal macrophages [58]. On the other hand, dysfunctional natural killer (NK) cells [59] that express a higher number of KIR (killing inhibitory receptors) [60] and a lower level of KAR (killing activating receptors), could play a role in the unsuitable activation and polarization of macrophages by inhibiting their phagocytic capacity and the expression of scavenger receptors [61]. Furthermore, these NK cells induce the activation of T regulatory (Treg) lymphocytes, whose inhibitory activity would allow endometrial cells to escape from local immune surveillance [62,63,64,65,66]. Additionally, it has been described that the number of peritoneal Myeloid-Derived Suppressor Cells (MDSCs) rapidly increases in the presence of endometrial tissue, thus contributing to the inhibited immune response [48,67]. Recently Gou et al. [68] showed that ERβ modulates the production of CCL2 (C-C motif chemokine ligand 2, also known as monocyte chemoattractant protein (MCP)-1) via NF-κB (nuclear factor kappa-light-chain enhancer of activated B cells) signaling in endometriotic stromal cells, recruiting macrophages to ectopic lesions and, thus, promoting the pathogenesis.
Additionally, multiple inflammatory mediators are secreted into the endometriotic milieu by several immune cells [69,70]. These mediators include interleukin (IL)-1β (induces the expression of cyclooxygenase 2 (COX-2) and vascular endothelial growth factor (VEGF), increasing angiogenesis) [71,72]; IL-6 (inhibits NK cell-mediated cytotoxicity) [73]; IL-8, which induces migration and increases the survival, proliferation, and angiogenesis of migrated endometrium [74,75]; transforming growth factor (TGF)-β, which is involved in post-surgical tissue adherence [62]; IL-27, which promotes the development of endometriotic lesions by inducing the secretion of IL-10 by T helper (TH) 17 cells [76]; IL-32, which increases cell viability and invasiveness [77]; IL-33, which exacerbates endometriotic lesions by polarizing peritoneal macrophages to M2 subtype secreting IL-1β [55]; prostaglandin E2 (PGE2), which, together with IL-4, induces the synthesis of estrogens, increases the production of fibroblast growth factor (FGF)-9, and inhibits the rate of apoptosis [78]; macrophage inhibitory factor (MIF), which induces angiogenesis and endometriotic cell expression of VEGF, IL-8, and MCP-1 (CCL2) [79]; MCP-1 (CCL2), which recruits monocytes [75,80]; and soluble intercellular adhesion molecule (sICAM)-1 [81], among others [69,70]. Additionally, the Akt (also known as protein kinase B) and mitogen-activated protein kinase (MEK)/extracellular-signal regulated kinase (ERK) intracellular signaling routes, together with signal transducer and activator of transcription (STAT) proteins and SMAD transcription factors have been described as key molecular pathways that play a role in the mechanisms leading to the development of endometriosis [82,83,84], as reviewed by Aznaurova et al. [85]; McKinnon et al. [86]; Riccio et al. [87]; Patel et al. [47]; and Zhang et al. [48].
4. Endometriosis and Diet
There is also a great interest in uncovering the impact of diet components on both endometriosis development and treatment, mostly based on growing evidence of their relationships with chronic inflammation, redox state, estrogen activity, prostaglandin metabolism, and the menstrual cycle, among others. Although epidemiological findings obtained from these studies are difficult to consistently replicate, recent reports and reviews suggest that fruits and vegetables, calcium and vitamin D from dairy products, and fish oils and Omega-3 polyunsaturated fatty acids are related to a lower risk of endometriosis [88,89]. On the contrary, a high intake of trans fats and red meat, processed or not, is highly associated with endometriosis development [88], while consumption of poultry, fish, shellfish, and eggs is not associated with this disease [90,91]. Recently, Nodler et al. [92], in a prospective cohort study, showed that greater dairy consumption, specifically yogurt and ice cream, during adolescence is associated with a lower risk of laparoscopically-confirmed endometriosis [92]. Regarding fruit and vegetable consumption and endometriosis risk, the recent work of Harris et al. [15] in a wide prospective cohort study reported an inverse association between greater fruit intake and risk of laparoscopically confirmed endometriosis, especially for citrus fruits (women with ≥ 1 servings of citrus fruits/day had a 22% lower endometriosis risk (95% CI = 0.69–0.89; p = 0.004) compared to those consuming < 1 serving/week. Furthermore, consumption of ≥ 1 servings/day of cruciferous vegetables (cauliflower, cabbage and Brussels sprouts, but not broccoli) was significantly associated with a higher endometriosis risk [15]. As Huijs and Nap [93] stated in their review, the effect of specific food products on endometriosis symptoms and whether they provide or not a synergic effect remains unclear.
The link between diet and endometriosis highlights the potential of the anti-inflammatory compounds present in foods to alleviate endometriosis symptoms and their progression. However, there are current gaps that preclude obtaining reproducible and clear interpretable results. First of all, these studies are based on retrospective or case-control populations and diet intake relays in self-reported food frequency questionnaires, which could lead to a bias whereby females report greater intake of those foods considered healthier. Additionally, contribution of the level of EDC food contamination, for example, through pesticides in fruits and vegetables, or estrogenic EDCs present in farmed animal foods, cannot be taken into account. All of these considerations underscore the necessity to perform new case-control studies in endometriosis patients eating strictly controlled diets (type of food, cooking and food preparation, and duration of the intervention) using foods rich in anti-inflammatory compounds by studying the level of endometriosis inflammatory markers and the clinical evolution.
5. Brassicaceae and Their Bioactive Compounds in Inflammation
As mentioned before, Brassica crops are well known for their high content in health-promoting compounds [94,95]. Specifically, glucosinolates (GLSs) and their bioactive breakdown product, ITCs, have shown different types of activity, such as induction of detoxification Phase II enzymes [96] and anti-tumorigenic [97] or anti-inflammatory effects [98]. Glucosinolates are secondary plant metabolites that are mainly present in vegetables from the Brassicaceae family. Their basic structure is a thiohydrozimate-O-sulfonate group linked with a glucose, whose side chain can vary depending on the amino acid from which they are derived [99]. Inside the plant cells, GLSs are stable, but when tissue disruption takes places, for example, chewing or wounding, these biomolecules are hydrolyzed by the enzyme myrosinase (EC 3.2.1.147), producing ITCs [100]. Cooking methods can also affect the content in glucosinolates and its degree of conversion to ITCs. For example, steaming, microwave cooking, and stir frying are processes that reduce the glucosinolate content the least and can even improve its extraction from the food matrix [101]. However, boiling or blanching usually inactivate myrosinase while leaching glucosinolates and breakdown products into the water [102]. A link between chronic inflammatory-related diseases, such as cancer or obesity, and these bioactive ITCs has been reported [10,103]. Furthermore, different ITCs are involved in diverse mechanisms and pathways of inflammatory processes; this analyzed here under the perspective of endometriosis.
5.1. Aliphatic ITCs and Related Metabolites
One of the most frequently studied aliphatic ITCs is sulforaphane (SFN), the resulting hydrolysis derivative of glucoraphanin, due to its effects on human health. SFN is widely considered to be a multi-faceted agent due to its role in several cellular pathways—attenuating, reversing, or even blocking different activities in the cellular metabolism [97]. For example, it has been reported that SFN specially induces the activation of Phase II detoxification enzymes, alters cellular signaling pathways, and takes part in the suppression of pro-inflammatory responses [104]. Specifically, SFN has been described as an inductor of the Nrf2 transcription factor (Figure 1A), which is responsible for the transcription of various genes involved in antioxidant activities or anti-inflammatory pathways [105]. The influence of SFN is based on its ability to bind the cysteine residues present in the Nrf2 repressor Keap1. under normal conditions, Keap1 promotes the degradation of Nrf2 in the proteasome, but when SFN interacts with it, Nrf2 is released and translocated to the nucleus [106].
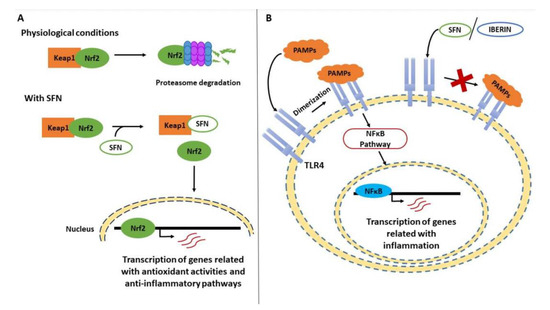
Figure 1.
Effect of Brassicaceae bioactives compounds upon the transcription of genes related with inflammation. (A) In physiological conditions, Keap1 promotes the degradation of Nrf2 by the proteasome. However, when sulforaphane (SFN) interacts with Keap1 by binding its cysteine residues, Nef2 is released and translocated to the nucleus. In this way, Nrf2 up regulates the transcription of genes related with antioxidant activities and anti-inflammatory pathways. (B) In an inflammatory process, Toll-like receptor 4 (TLR4) undergoes dimerization after recognizing pathogen-associated molecular patterns (PAMPs), activating the NFκB pathway and up-regulating genes related with inflammation. When SFN and iberin interact with TLR4, it cannot oligomerizate, and the inflammatory response is inhibited.
Furthermore, SFN is involved in the NFκB pathway, decreasing its ability to bind to target genes related to the inflammatory response, such as pro-inflammatory interleukins, including tumor necrosis factor (TNF)-α [97]. In this signaling cascade, Toll-like receptor (TLR)-4 is the first element, and it is responsible for recognizing pathogen-associated molecular patterns (PAMPs) or intrinsic molecules (Figure 1B), such as fibronectin and heparan sulfate [107,108]. It has been reported that SFN can bind to the cysteine residues in this receptor, suppressing its oligomerization and inhibiting the inflammatory response [109].
Additionally, several pre-clinical studies have demonstrated the efficiency of this ITC in models of chronic inflammatory diseases [110]. For example, Zhao et al. [111] demonstrated that the administration of SFN to a nephropathy murine model reduced the level of reactive oxygen species (ROS) and activated Nrf2 transcription factor. However, little to no information about the effects in models of endometriosis has been found. Recently, Zhou et al. [112] revealed that SFN attenuated endometriosis symptoms in rats by diminishing the levels of TNF-α and IL-6 in peritoneal fluid and plasma. Moreover, SFN seems to decrease the expression of VEGF, affecting the neoangiogenesis of the endometriotic foci. In addition, SFN has been reported to inhibit the growth of ectopic endometrial tissue in sciatic endometriosis rat models, showing a decrease in both the size of lesions and the VEGF level [113]. However, more studies are needed in order to determine the effect of SFN in long-term dietary interventions.
SFN has a structural analog, sulforaphene (SFE), which differs only by having a double bond in the alkyl chain. It is derived from the aliphatic GLS glucoraphenin and can be found mainly in radishes [114]. Although research on SFE has been focused on its anti-carcinogenic and pro-apoptotic effects [115], little is known about its direct effect on inflammatory processes. However, it is known that SFE can lose its double bond, turning into SFN, whose properties are better known [116,117].
Another aliphatic ITC present in Brassica foods, including broccoli sprouts, is erucin [118]. Although an inter-conversion of SFN to erucin and vice versa has been reported in humans after its consumption, the effects of erucin in inflammation are also significant [119]. For example, it has been reported that it decreases the DNA-binding capacity of NFκB, thus reducing the transcription of target genes related to the inflammatory process, including TNF-α, IL-6, COX-2, and inducible nitric oxide synthase (iNOS) [120]. Additionally, glucoiberin is a glucosinolate that is present in products such as cabbage and kale, and its degradation product is iberin [121,122]. As mentioned previously, TLRs play an important role in the induction of the innate immune response, and iberin can prevent the dimerization of TLRs, down-regulating NFκB signaling [107,123].
In oilseed rape and other herbaceous Brassicas, the aliphatic ITC allyl-ITC (AITC) derived from sinigrin does not have sulfur atoms in its side chain, unlike SFN and SFE. The action of AITC in the NFκB pathway when administered to the LPS-activated macrophage RAW 264.7 has been reported to decrease the production of TNF-α, IL-6, and nitric oxide [124]. In addition, in vivo studies performed in rat models of mammary carcinogenesis showed that AITC administration did not provoke a decrease in p65-NFκB expression and, thus, in pro-inflammatory cytokines such as IL-6 [125]. The same group also reported that AITC has a preventive effect on DMBA-induced mammary carcinogenesis by modulating the aryl hydrocarbon receptor (AhR)/Nrf2 signaling pathway [126]. This cytoplasmic receptor and transcription factor has a major role in environmental pollutant detoxification [127]. Nevertheless, recent studies have highlighted the role of AhR as a negative regulator of the immune response [128]. In this regard, it has been reported that AhR-null mice produce higher levels of pro-inflammatory cytokines, such as TNF-α and IL-12 [129].
5.2. Indoles and Related Compounds
The hydrolysis of glucobrassicin, a major indole glucosinolate present in Brassicas, including broccoli or cabbage [130], produces indole-3-carbinol (I3C). I3C can be converted into its dimeric condensation product 3,3-diindolylmethane (3,3-DIM) [131]. Both biomolecules are AhR agonists that decrease the level of IL-1β and increase the rate of the detoxification cascade [132]. In addition, when I3C was administrated to murine models of doxorubicin-induced damage, positive stimulation of the Nrf2 response and down-regulation of p50-NFκB expression was observed [133]. However, when macrophages derived from monocytes of systemic lupus erythematosus patients were treated with I3C, a switch toward a M2 phenotype was described [134]. As mentioned before, M2 macrophages could help with the survival of endometrial cysts [54]. Thus, in the absence of in vivo studies of I3C applied to endometriosis, the specific effect on this disease remains unknown.
On the other hand, 3,3-DIM alone has been reported to inhibit the inflammatory response by also down-regulating the NFκB pathway and decreasing the levels of PGE2, TNF-α, IL-6, and IL-1β in murine macrophage cell cultures [135], and the oral administration of 3,3-DIM to a model of acute colitis in mice provoked the down-regulation of various types of VEGF and the expression of the VEGF receptor-2 [136]. This function would be beneficial for endometriosis patients to reduce the neoangiogenesis of lesions. In addition, the effects of 3,3-DIM have been also tested in combination with dienogest, a common progestrin that is commonly prescribed to treat pain associated with endometriosis. Women who were administered with this combination for 3 months reported less endometriosis-associated pelvic pain [137]. However, only eight patients finished the study; therefore, studies with a larger number of volunteers are needed to better understand its effects.
Ascorbigen is a biomolecule obtained from glucobrassicin degradation products generated after the enzymatic hydrolysis of I3C in the presence of ascorbic acid that has shown high antioxidant potential [138]. Fermented cabbage extracts enriched in ascorbigen were found to have improved antioxidant capacity and nitric oxide inhibition in RAW 264.7 murine cell cultures [139]. Nonetheless, information about the exact effects of this metabolite in inflammation is very limited since no recent in vivo studies are available.
6. Aquaporins in Endometriosis
Aquaporins (AQPs) belong to the membrane intrinsic protein (MIP) family [140] and are found in practically all living organisms [141]. AQPs are tetrameric, small (25–34 kDa), hydrophobic proteins that constitute integral membrane channels [142]. The main function of AQPs is water transport and flux water regulation in cellular membranes [143]. Since their discovery, AQPs have been the focus of many studies, given the importance of water transport in all biological processes, and have been shown to be involved in several diseases of different tissues [144]. Specifically, nine of the thirteen total AQPs (AQP1, 2, 3, 4, 5, 6, 8, 9, and 11) discovered in mammals have been identified as existing in the human female reproductive tract [145]. Proper water transport and homeostasis is crucial for normal reproductive performance and, thus, these AQPs play key roles in the female reproductive system, as they are involved in vaginal lubrication, cervical ripening, implantation, uterine water inhibition, blastocyst formation, ovum transport, follicle maturation, and oocyte cryopreservation [145]. However, apart from the main described functions, some aquaporins are also involved in biological alterations of the reproductive system such as cell migration, proliferation, carcinogenesis, and inflammatory processes [146,147]. As previously described, these processes are crucial for the development of endometriosis, and thus, the link between AQPs and the pathogenesis of endometriosis must be further explored [148]. AQP1 might have a role in endometriosis via the Wnt/β-catenin signaling pathway. Hence, AQP1 gene silencing entails a decrease in the adhesion and invasion of ectopic endometrial cells, as well as a decrease in angiogenesis and an increase in the apoptosis rate of these cells [149]. In the same way, AQP5, whose expression is menstrual cycle-dependent [150], is more highly expressed in eutopic as compared with ectopic endometrial cells in women with endometriosis [151]. Another investigation reported that AQP5 expression was induced by estrogen through activation of the estrogen-response element (ERE) in the promoter region of the AQP5 gene [152]. Therefore, it could be related to endometrial cell invasion and proliferation. This aspect was observed in a study where overexpression of AQP5 promoted endometrial tumor cell migration, while knockdown of AQP5 decreased migration of these cells [153]. Moreover, Choi et al. [154] described that AQP2 and AQP8 expression was significantly increased in the eutopic endometrial cells of patients with endometriosis as compared with a control group. Also, this work stated that the endometrial expression of AQP9 was significantly decreased in the eutopic endometrium of patients with endometriosis as compared with that of healthy women. Besides, it was demonstrated that the down-regulation of AQP9 increased cell migration and invasion through the activation of the ERK/p38 MAPK signaling pathways. These results are consistent with the demonstration of changes in the expression of AQP9 in several types of cancer and inflammatory diseases [155], which suggests that AQP9 could also play an important role in the pathogenesis of endometriosis.
Additionally, it has been reported that the expression of AQP1, present in macrophages, regulated their migration in a murine model of peritonitis [156], showing that functional AQP1 suppresses the migration of resting macrophages. Further, the expression of AQP1 was found to attenuate macrophage-mediated inflammatory responses in LPS-induced acute kidney injury by inhibiting ERK and p38 MAPK activation [157]. In addition, AQP3 has shown to be a key component in the immune function of macrophages through its participation in water and glycerol membrane transportation, as well as in their phagocytic and migration activity [147]. Furthermore, AQP4 blockade alleviated the development and severity of irradiated lung damage in a mouse model. This was associated with attenuated infiltration of inflammatory cells, decreased production of pro-inflammatory cytokines, and inhibited activation of M2 macrophages [158].
Therefore, the available results suggest that a relation between AQPs and endometriosis would not only be mediated by their role as water membrane channels (Figure 2), which is their major physiological function [143]. Their involvement in several intracellular signaling pathways as signal regulating factors (via transporting molecules or coupling with other proteins) [159] should be taken into account. Particularly, AQPs 1, 2, 8, and 9 could be related to endometriosis due to their participation in different signaling pathways such as Wnt/β-catenin, ERK/p38 MAPKm, and PI3K/Akt [149,152,154]. Hence, more studies are needed to reveal the actual signaling regulatory function of AQPs. In contrast, the roles of AQP1 and 5 in different inflammatory processes, whose regulation is altered in endometriosis (displaying maintained or increased expression), could be mediated by their roles in water fluid homeostasis as water transporters, and thus their potential relevance in endometriosis should be also further investigated [147,160].
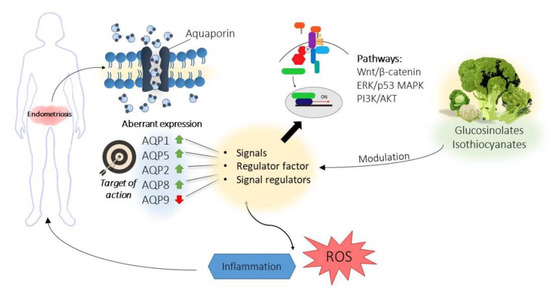
Figure 2.
Role of aquaporins in endometriosis. Aquaporins are involved in different signaling pathways related with inflammation and endometriosis. Besides, aquaporins present an aberrant expression in endometrial tissues of women with endometriosis and therefore, they could be the target of therapies based on bioactive compounds derived from brassicas.
Modulation of Aquaporins by Bioactive Compounds: Further Targeted Therapies
The modulation of the presence or functionality of aquaporins by bioactive compounds or drugs may provide new opportunities for therapeutic applications in a variety of diseases that involve alterations of cellular homeostasis and inflammatory processes [161]. In this sense, gating of the AQP channel by different non-covalent small molecule inhibitors has been reported [162]. Blockers of AQP4 have demonstrated a positive effect in ischemia-mediated cerebral edema models [163]. Also, some of the quaternary ammonium compounds [164,165,166], arylsulfonamide-based carbonic anhydrase inhibitors [167,168], and certain anti-epileptics [169] have been suggested to be selective blockers of AQP1 and AQP4. Furthermore, on the other hand, there are pharmacological synergists to AQP functionality, i.e., the aryl sulfonamide-related derivatives [170] and the chemical derivative of the arylsulfonamide compound furosemide [166], that directly and specifically potentiate AQP-mediated water transport.
Aberrant expression of AQPs in eutopic and ectopic endometrial cells could be a target of action against endometriosis disease. Some studies of endometriosis cell models have reported a deficit in antioxidant enzymes and, thus, a low antioxidant capacity. Therefore, excess accumulation of reactive oxygen species could play a role in the infertility associated with endometriosis [171]. AQP8 has been reported as a fine regulator of redox signal transduction since it regulates permeability to water and H2O2 under stress conditions [172]. As described above, AQP8 is overexpressed in eutopic endometrial cells of women with endometriosis [154]. Hence, the regulation of AQP8 expression could reduce the level of ROS in cells, therefore reducing oxidative stress. This could favor better evolution of endometriosis. In this sense, a recent study carried out in the erythromegakaryocytic cell line B1647 showed a downregulation of AQP8 expression with low doses of SFN (10–30 µM) [173]. Therefore, SFN could be a good therapy to regulate AQP8 expression in endometrial cells and decrease oxidative stress in these cells.
Furthermore, if we consider that endometriosis is an estrogen-dependent disease [174], new therapies that counter the effect of estradiol need to be explored. Estrogens are steroid hormones that regulate AQP expression in the female reproductive system [175]. The expression of AQP2 and AQP5, which is up-regulated in endometrial cells and seem to play a role in the estrogen-related migration, invasion, adhesion, and proliferation of these cells, could be regulated to act on the ERE located in the promoters of the genes encoding AQP2 and AQP5 [152,176]. In some studies, SFN [177] and I3C [178,179,180] have been shown to be modulators of the estrogen effects in several types of cancer. Given the common features of cancer and endometriosis, these compounds could induce changes in the ERE of the AQP5 gene, thus regulating its expression. In the same way, it has been reported that sulforaphane protects the central nervous system in traumatic brain injury patients by reducing the loss of AQP4 in the injury core and increasing the level of AQP4 in the penumbra region [181,182].
All of the above data indicate that AQP activity can be modified through distinct mechanisms. However, it is also important to consider that pharmacological and bioactive natural compounds might interact regarding AQP expression and/or functionality, reflecting diverse affinity and efficacy. Therefore, it has become clear that the investigation of plant biomolecules as glucosinolates/isotiocyanates that target the modulation of AQPs is an interesting research objective, especially as these are natural compounds with fewer side effects than other therapeutic agents.
7. Conclusions and Perspectives
To summarize the results presented in this review, macrophages are key immune cells in endometriotic lesions, which are characterized by an anomalous inflammatory environment and cell growth in women with endometriosis. We have presented and discussed a new approach based on the potential role of glucosinolates/isothiocyanates from brassicas as putative natural anti-inflammatory compounds, which could reduce the symptoms and progression of endometriosis, improving the quality of life for affected women. The potential role of glucosinolates in the development chronic inflammatory diseases should be investigated since there is still little information in this respect. In connection to that, water transport and solute movement, in which AQPs are involved, are crucial events not only in the maintenance of immune homeostasis but also in human pathophysiology. The reported importance of AQPs in different inflammatory processes could be integrated in the role played by macrophages in endometriosis. Therefore, the modulation of cellular water and solute transport through AQPs by glucosinolates may provide new opportunities for the design of preventive and therapeutic approaches to improve the quality of life of women with endometriosis.
8. Materials and Methods
A non-systematic approach was chosen and a narrative synthesis of the results of the searched articles was carried out in accordance with Gasparian et al. [183] and Saracci et al. [184].
Author Contributions
Conceptualization: all coauthors; methodology: P.G.-I., L.Y.-M., A.J.R.-A., D.A.M.; data description and interpretation: all coauthors; conclusions and discussion of perspectives: all coauthors; writing—original draft preparation: P.G.-I., L.Y.-M., P.G.-P., A.J.R.-A., D.A.M.; final editing: all coauthors; project administration and supervision, A.J.R.-A., D.A.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by Fundación Seneca—Murcia Regional Agency for Science and Technology through the Project Ref. 20855/PI/18. P.G.-I. was funded by a grant from the Fundación Séneca-CARM, Spain (21273/FPI/19), and L.Y.-M. was funded by a grant from the Spanish Ministerio de Ciencia, Educación y Universidades (FPU17/02261). This work was also partially supported by the Spanish Ministerio de Economía, Industria y Competitividad under Grant AGL2016-80247-C2-1-R.
Acknowledgments
The authors A.J.R.-A., M.M.-E and P.G.-P. also thank the donors who supported Precipita crowdfunding (PR200; Endometriosis), especially Macarena Guillamón, Mercedes Jimenez, Ramón Diaz Petrel, and Victoria Millor. We also thank the association “Endomequé”, founded by Lucia Arnaiz García, for their generous financial support of scientific endometriosis research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vercellini, P.; Viganò, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261–275. [Google Scholar] [CrossRef]
- Zondervan, K.T.; Becker, C.M.; Koga, K.; Missmer, S.A.; Taylor, R.N.; Viganò, P. Endometriosis. Nat. Rev. Dis. Primers 2018, 4, 9. [Google Scholar] [CrossRef]
- As-Sanie, S.; Black, R.; Giudice, L.C.; Gray Valbrun, T.; Gupta, J.; Jones, B.; Laufer, M.R.; Milspaw, A.T.; Missmer, S.A.; Norman, A.; et al. Assessing research gaps and unmet needs in endometriosis. Am. J. Obstet. Gynecol. 2019, 221, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Denny, E.; Mann, C.H. A clinical overview of endometriosis: A misunderstood disease. Br. J. Nurs. 2007, 16, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, V.L.; de Franciscis, P.; Barra, F.; Schiattarella, A.; Tropea, A.; Tesarik, J.; Shah, M.; Kahramanoglu, I.; Marques Cerentini, T.; Ponta, M.; et al. Sexuality in women with endometriosis: A critical narrative review. Minerva Med. 2020, 111, 79–89. [Google Scholar] [PubMed]
- Jones, G.; Jenkinson, C.; Kennedy, S. The impact of endometriosis upon quality of life: A qualitative analysis. J. Psychosom. Obstet. Gynaecol. 2004, 25, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Aerts, L.; Grangier, L.; Streuli, I.; Dällenbach, P.; Marci, R.; Wenger, J.-M.; Pluchino, N. Psychosocial impact of endometriosis: From co-morbidity to intervention. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 50, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.A.; Templeton, A.A.; Thomson, L.; Fraser, C. Menstrual symptoms in women with pelvic endometriosis. Br. J. Obstet. Gynaecol. 1991, 98, 558–563. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, V.L.; De Franciscis, P.; Barra, F.; Schiattarella, A.; Török, P.; Shah, M.; Karaman, E.; Marques Cerentini, T.; Di Guardo, F.; Gullo, G.; et al. Quality of life in women with endometriosis: A narrative overview. Minerva Med. 2020, 111, 68–78. [Google Scholar] [CrossRef] [PubMed]
- López-Chillón, M.T.; Carazo-Díaz, C.; Prieto-Merino, D.; Zafrilla, P.; Moreno, D.A.; Villaño, D. Effects of long-term consumption of broccoli sprouts on inflammatory markers in overweight subjects. Clin. Nutr. 2019, 38, 745–752. [Google Scholar] [CrossRef]
- Finicelli, M.; Squillaro, T.; Di Cristo, F.; Di Salle, A.; Melone, M.A.B.; Galderisi, U.; Peluso, G. Metabolic syndrome, Mediterranean diet, and polyphenols: Evidence and perspectives. J. Cell. Physiol. 2019, 234, 5807–5826. [Google Scholar] [CrossRef] [PubMed]
- Soave, I.; Occhiali, T.; Wenger, J.M.; Pluchino, N.; Caserta, D.; Marci, R. Endometriosis and food habits: Can diet make the difference? J. Endometr. Pelvic Pain Disord. 2018, 10, 59–71. [Google Scholar] [CrossRef]
- Parazzini, F.; Esposito, G.; Tozzi, L.; Noli, S.; Bianchi, S. Epidemiology of endometriosis and its comorbidities. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 209, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Trabert, B.; Peters, U.; De Roos, A.J.; Scholes, D.; Holt, V.L. Diet and risk of endometriosis in a population-based case-control study. Br. J. Nutr. 2011, 105, 459–467. [Google Scholar] [CrossRef]
- Harris, H.R.; Eke, A.C.; Chavarro, J.E.; Missmer, S.A. Fruit and vegetable consumption and risk of endometriosis. Hum. Reprod. 2018, 33, 715–727. [Google Scholar] [CrossRef]
- Agic, A.; Djalali, S.; Diedrich, K.; Hornung, D. Apoptosis in Endometriosis. Gynecol. Obstet. Investig. 2009, 68, 217–223. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, M.A.; Lee, M.; Suh, D.H.; Kim, K.; No, J.H.; Chung, H.H.; Kim, Y.B.; Song, Y.S. Effect of endometriosis on the prognosis of ovarian clear cell carcinoma: A two-center cohortsStudy and meta-analysis. Ann. Surg. Oncol. 2015, 22, 2738–2745. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, T.H.; Chung, H.H.; Song, Y.S. Risk and prognosis of ovarian cancer in women with endometriosis: A meta-analysis. Br. J. Cancer 2014, 110, 1878–1890. [Google Scholar] [CrossRef]
- Machado-Linde, F.; Sánchez-Ferrer, M.L.; Cascales, P.; Torroba, A.; Orozco, R.; Silva Sánchez, Y.; Nieto, A.; Fiol, G. Prevalence of endometriosis in epithelial ovarian cancer. Analysis of the associated clinical features and study on molecular mechanisms involved in the possible causality. Eur. J. Gynaecol. Oncol. 2015, 36, 21–24. [Google Scholar]
- Pearce, C.L.; Templeman, C.; Rossing, M.A.; Lee, A.; Near, A.M.; Webb, P.M.; Nagle, C.M.; Doherty, J.A.; Cushing-Haugen, K.L.; Wicklund, K.G.; et al. Association between endometriosis and risk of histological subtypes of ovarian cancer: A pooled analysis of case-control studies. Lancet. Oncol. 2012, 13, 385–394. [Google Scholar] [CrossRef]
- Wei, J.-J.; William, J.; Bulun, S. Endometriosis and ovarian cancer: A review of clinical, pathologic, and molecular aspects. Int. J. Gynecol. Pathol. 2011, 30, 553–568. [Google Scholar] [CrossRef] [PubMed]
- Sampson, J.A. Metastatic or embolic endometriosis, due to the menstrual dissemination of endometrial tissue into the venous circulation. Am. J. Pathol. 1927, 3, 93–110.43. [Google Scholar]
- Sourial, S.; Tempest, N.; Hapangama, D.K. Theories on the pathogenesis of endometriosis. Int. J. Reprod. Med. 2014, 2014, 179515. [Google Scholar] [CrossRef] [PubMed]
- Asghari, S.; Valizadeh, A.; Aghebati-Maleki, L.; Nouri, M.; Yousefi, M. Endometriosis: Perspective, lights, and shadows of etiology. Biomed. Pharmacother. 2018, 106, 163–174. [Google Scholar] [CrossRef]
- García-Peñarrubia, P.; Ruiz-Alcaraz, A.J.; Martínez-Esparza, M.; Marín, P.; Machado-Linde, F. Hypothetical roadmap towards endometriosis: Prenatal endocrine-disrupting chemical pollutant exposure, anogenital distance, gut-genital microbiota and subclinical infections. Hum. Reprod. Update 2020, 26, 214–246. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Chen, X.; Zhao, Y.; Cao, B.; Zhao, W. Effects of prenatal environmental exposures on the development of endometriosis in female offspring. Reprod. Sci. 2016, 23, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Missmer, S.A.; Hankinson, S.E.; Spiegelman, D.; Barbieri, R.L.; Michels, K.B.; Hunter, D.J. In utero exposures and the incidence of endometriosis. Fertil. Steril. 2004, 82, 1501–1508. [Google Scholar] [CrossRef]
- Upson, K.; Sathyanarayana, S.; Scholes, D.; Holt, V.L. Early-life factors and endometriosis risk. Fertil. Steril. 2015, 104, 964–971.e5. [Google Scholar] [CrossRef]
- Parasar, P.; Ozcan, P.; Terry, K.L. Endometriosis: Epidemiology, diagnosis and clinical management. Curr. Obstet. Gynecol. Rep. 2017, 6, 34–41. [Google Scholar] [CrossRef]
- Shafrir, A.L.; Farland, L.V.; Shah, D.K.; Harris, H.R.; Kvaskoff, M.; Zondervan, K.; Missmer, S.A. Risk for and consequences of endometriosis: A critical epidemiologic review. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 51, 1–15. [Google Scholar] [CrossRef]
- Backonja, U.; Hediger, M.L.; Chen, Z.; Lauver, D.R.; Sun, L.; Peterson, C.M.; Buck Louis, G.M. Beyond body mass index: Using anthropometric measures and body composition indicators to assess odds of an endometriosis diagnosis. J. Womens. Health (Larchmt) 2017, 26, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Hediger, M.L.; Hartnett, H.J.; Buck Louis, G.M. Association of endometriosis with body size and figure. Fertil. Steril. 2005, 84, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Allebeck, P.; Mishra, G.D.; Koupil, I. Developmental origins of endometriosis: A Swedish cohort study. J. Epidemiol. Community Health 2019, 73, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Cano-Sancho, G.; Ploteau, S.; Matta, K.; Adoamnei, E.; Louis, G.B.; Mendiola, J.; Darai, E.; Squifflet, J.; Le Bizec, B.; Antignac, J.-P. Human epidemiological evidence about the associations between exposure to organochlorine chemicals and endometriosis: Systematic review and meta-analysis. Environ. Int. 2019, 123, 209–223. [Google Scholar] [CrossRef]
- Hunt, P.A.; Sathyanarayana, S.; Fowler, P.A.; Trasande, L. Female reproductive disorders, diseases, and costs of exposure to endocrine disrupting chemicals in the European Union. J. Clin. Endocrinol. Metab. 2016, 101, 1562–1570. [Google Scholar] [CrossRef]
- Smarr, M.M.; Kannan, K.; Buck Louis, G.M. Endocrine disrupting chemicals and endometriosis. Fertil. Steril. 2016, 106, 959–966. [Google Scholar] [CrossRef]
- Wen, X.; Xiong, Y.; Qu, X.; Jin, L.; Zhou, C.; Zhang, M.; Zhang, Y. The risk of endometriosis after exposure to endocrine-disrupting chemicals: A meta-analysis of 30 epidemiology studies. Gynecol. Endocrinol. 2019, 35, 645–650. [Google Scholar] [CrossRef]
- Matalliotakis, I.M.; Cakmak, H.; Fragouli, Y.G.; Goumenou, A.G.; Mahutte, N.G.; Arici, A. Epidemiological characteristics in women with and without endometriosis in the Yale series. Arch. Gynecol. Obstet. 2008, 277, 389–393. [Google Scholar] [CrossRef]
- Heilier, J.-F.; Donnez, J.; Nackers, F.; Rousseau, R.; Verougstraete, V.; Rosenkranz, K.; Donnez, O.; Grandjean, F.; Lison, D.; Tonglet, R. Environmental and host-associated risk factors in endometriosis and deep endometriotic nodules: A matched case–control study. Environ. Res. 2007, 103, 121–129. [Google Scholar] [CrossRef]
- Parazzini, F.; Cipriani, S.; Bravi, F.; Pelucchi, C.; Chiaffarino, F.; Ricci, E.; Viganò, P. A metaanalysis on alcohol consumption and risk of endometriosis. Am. J. Obstet. Gynecol. 2013, 209, 106.e1–106.e10. [Google Scholar] [CrossRef]
- Saha, R.; Kuja-Halkola, R.; Tornvall, P.; Marions, L. Reproductive and lifestyle factors associated with endometriosis in a large cross-sectional population sample. J. Womens. Health (Larchmt) 2017, 26, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Rižner, T.L. Estrogen metabolism and action in endometriosis. Mol. Cell. Endocrinol. 2009, 307, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, S.; Remorgida, V.; Maganza, C.; Venturini, P.L.; Salvatore, S.; Papaleo, E.; Candiani, M.; Leone Roberti Maggiore, U. Aromatase and endometriosis: Estrogens play a role. Ann. N. Y. Acad. Sci. 2014, 1317, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Simoens, S.; Hummelshoj, L.; D’Hooghe, T. Endometriosis: Cost estimates and methodological perspective. Hum. Reprod. Update 2007, 13, 395–404. [Google Scholar] [CrossRef]
- Klemmt, P.A.B.; Starzinski-Powitz, A. Molecular and cellular pathogenesis of endometriosis. Curr. Women’s Health Rev. 2018, 14, 106–116. [Google Scholar] [CrossRef]
- Miller, J.E.; Ahn, S.H.; Monsanto, S.P.; Khalaj, K.; Koti, M.; Tayade, C. Implications of immune dysfunction on endometriosis associated infertility. Oncotarget 2017, 8, 7138–7147. [Google Scholar] [CrossRef]
- Patel, B.G.; Lenk, E.E.; Lebovic, D.I.; Shu, Y.; Yu, J.; Taylor, R.N. Pathogenesis of endometriosis: Interaction between Endocrine and inflammatory pathways. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 50, 50–60. [Google Scholar] [CrossRef]
- Zhang, T.; De Carolis, C.; Man, G.C.W.; Wang, C.C. The link between immunity, autoimmunity and endometriosis: A literature update. Autoimmun. Rev. 2018, 17, 945–955. [Google Scholar] [CrossRef]
- Pepe, G.; Locati, M.; Della Torre, S.; Mornata, F.; Cignarella, A.; Maggi, A.; Vegeto, E. The estrogen-macrophage interplay in the homeostasis of the female reproductive tract. Hum. Reprod. Update 2018, 24, 652–672. [Google Scholar] [CrossRef]
- Capobianco, A.; Rovere-Querini, P. Endometriosis, a disease of the macrophage. Front. Immunol. 2013, 4, 9. [Google Scholar] [CrossRef]
- Bacci, M.; Capobianco, A.; Monno, A.; Cottone, L.; Di Puppo, F.; Camisa, B.; Mariani, M.; Brignole, C.; Ponzoni, M.; Ferrari, S.; et al. Macrophages are alternatively activated in patients with endometriosis and required for growth and vascularization of lesions in a mouse model of disease. Am. J. Pathol. 2009, 175, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Xie, H.; Yao, S.; Liang, Y. Macrophage and nerve interaction in endometriosis. J. Neuroinflamm. 2017, 14, 53. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Li, D.; Yuan, M.; Li, Q.; Zhen, Q.; Li, N.; Wang, G. Macrophages alternatively activated by endometriosis-exosomes contribute to the development of lesions in mice. Mol. Hum. Reprod. 2019, 25, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Liu, X.; Wang, H.; Guo, S.-W. The M2a macrophage subset may be critically involved in the fibrogenesis of endometriosis in mice. Reprod. Biomed. Online 2018, 37, 254–268. [Google Scholar] [CrossRef]
- Ono, Y.; Yoshino, O.; Hiraoka, T.; Akiyama, I.; Sato, E.; Ito, M.; Kobayashi, M.; Nakashima, A.; Wada, S.; Onda, T.; et al. IL-33 Exacerbates endometriotic lesions via polarizing peritoneal macrophages to M2 subtype. Reprod. Sci. 2020, 27, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Hudson, Q.J.; Ashjaei, K.; Perricos, A.; Kuessel, L.; Husslein, H.; Wenzl, R.; Yotova, I. Endometriosis Patients Show an Increased M2 Response in the Peritoneal CD14+low/CD68+low Macrophage Subpopulation Coupled with an Increase in the T-helper 2 and T-regulatory Cells. Reprod. Sci. 2020, 27, 1920–1931. [Google Scholar] [CrossRef]
- Takebayashi, A.; Kimura, F.; Kishi, Y.; Ishida, M.; Takahashi, A.; Yamanaka, A.; Wu, D.; Zheng, L.; Takahashi, K.; Suginami, H.; et al. Subpopulations of macrophages within eutopic endometrium of endometriosis patients. Am. J. Reprod. Immunol. 2015, 73, 221–231. [Google Scholar] [CrossRef]
- Ruiz-Alcaraz, A.J.; Martínez-Banaclocha, H.; Marín-Sánchez, P.; Carmona-Martínez, V.; Iniesta-Albadalejo, M.A.; Tristán-Manzano, M.; Tapia-Abellán, A.; García-Peñarrubia, P.; Machado-Linde, F.; Pelegrín, P.; et al. Isolation of functional mature peritoneal macrophages from healthy humans. Immunol. Cell Biol. 2020, 98, 114–126. [Google Scholar] [CrossRef]
- Yu, J.-J.; Sun, H.-T.; Zhang, Z.-F.; Shi, R.-X.; Liu, L.-B.; Shang, W.-Q.; Wei, C.-Y.; Chang, K.-K.; Shao, J.; Wang, M.-Y.; et al. IL15 promotes growth and invasion of endometrial stromal cells and inhibits killing activity of NK cells in endometriosis. Reproduction 2016, 152, 151–160. [Google Scholar] [CrossRef]
- Wu, M.Y.; Yang, J.H.; Chao, K.H.; Hwang, J.L.; Yang, Y.S.; Ho, H.N. Increase in the expression of killer cell inhibitory receptors on peritoneal natural killer cells in women with endometriosis. Fertil. Steril. 2000, 74, 1187–1191. [Google Scholar] [CrossRef]
- Chuang, P.-C.; Wu, M.-H.; Shoji, Y.; Tsai, S.-J. Downregulation of CD36 results in reduced phagocytic ability of peritoneal macrophages of women with endometriosis. J. Pathol. 2009, 219, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Hanada, T.; Tsuji, S.; Nakayama, M.; Wakinoue, S.; Kasahara, K.; Kimura, F.; Mori, T.; Ogasawara, K.; Murakami, T. Suppressive regulatory T cells and latent transforming growth factor-β-expressing macrophages are altered in the peritoneal fluid of patients with endometriosis. Reprod. Biol. Endocrinol. 2018, 16, 9. [Google Scholar] [CrossRef] [PubMed]
- Christodoulakos, G.; Augoulea, A.; Lambrinoudaki, I.; Sioulas, V.; Creatsas, G. Pathogenesis of endometriosis: The role of defective “immunosurveillance”. Eur. J. Contracept. Reprod. Health Care 2007, 12, 194–202. [Google Scholar] [CrossRef]
- Gogacz, M.; Winkler, I.; Bojarska-Junak, A.; Tabarkiewicz, J.; Semczuk, A.; Rechberger, T.; Adamiak, A. T regulatory lymphocytes in patients with endometriosis. Mol. Med. Rep. 2014, 10, 1072–1076. [Google Scholar] [CrossRef]
- de Barros, I.B.L.; Malvezzi, H.; Gueuvoghlanian-Silva, B.Y.; Piccinato, C.A.; Rizzo, L.V.; Podgaec, S. What do we know about regulatory T cells and endometriosis? A systematic review. J. Reprod. Immunol. 2017, 120, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-Q.; Zhou, W.-J.; Luo, X.-Z.; Tao, Y.; Li, D.-J. Synergistic effect of regulatory T cells and proinflammatory cytokines in angiogenesis in the endometriotic milieu. Hum. Reprod. 2017, 32, 1304–1317. [Google Scholar] [CrossRef] [PubMed]
- Elkabets, M.; Ribeiro, V.S.G.; Dinarello, C.A.; Ostrand-Rosenberg, S.; Di Santo, J.P.; Apte, R.N.; Vosshenrich, C.A.J. IL-1β regulates a novel myeloid-derived suppressor cell subset that impairs NK cell development and function. Eur. J. Immunol. 2010, 40, 3347–3357. [Google Scholar] [CrossRef]
- Gou, Y.; Li, X.; Li, P.; Zhang, H.; Xu, T.; Wang, H.; Wang, B.; Ma, X.; Jiang, X.; Zhang, Z. Estrogen receptor β upregulates CCL2 via NF-κB signaling in endometriotic stromal cells and recruits macrophages to promote the pathogenesis of endometriosis. Hum. Reprod. 2019, 34, 646–658. [Google Scholar] [CrossRef]
- Mu, F.; Harris, H.R.; Rich-Edwards, J.W.; Hankinson, S.E.; Rimm, E.B.; Spiegelman, D.; Missmer, S.A. A Prospective Study of Inflammatory Markers and Risk of Endometriosis. Am. J. Epidemiol. 2018, 187, 515–522. [Google Scholar] [CrossRef]
- Nothnick, W.; Alali, Z. Recent advances in the understanding of endometriosis: The role of inflammatory mediators in disease pathogenesis and treatment. F1000Research 2016, 5, 186. [Google Scholar] [CrossRef]
- Huang, F.; Cao, J.; Liu, Q.; Zou, Y.; Li, H.; Yin, T. MAPK/ERK signal pathway involved expression of COX-2 and VEGF by IL-1β induced in human endometriosis stromal cells in vitro. Int. J. Clin. Exp. Pathol. 2013, 6, 2129–2136. [Google Scholar] [PubMed]
- McLaren, J. Vascular endothelial growth factor and endometriotic angiogenesis. Hum. Reprod. Update 2000, 6, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-J.; Jeung, I.C.; Park, A.; Park, Y.-J.; Jung, H.; Kim, T.-D.; Lee, H.G.; Choi, I.; Yoon, S.R. An increased level of IL-6 suppresses NK cell activity in peritoneal fluid of patients with endometriosis via regulation of SHP-2 expression. Hum. Reprod. 2014, 29, 2176–2189. [Google Scholar] [CrossRef] [PubMed]
- Sikora, J.; Smycz-Kubańska, M.; Mielczarek-Palacz, A.; Kondera-Anasz, Z. Abnormal peritoneal regulation of chemokine activation-The role of IL-8 in pathogenesis of endometriosis. Am. J. Reprod. Immunol. 2017, 77, e12622. [Google Scholar] [CrossRef]
- Ulukus, M.; Ulukus, E.C.; Tavmergen Goker, E.N.; Tavmergen, E.; Zheng, W.; Arici, A. Expression of interleukin-8 and monocyte chemotactic protein 1 in women with endometriosis. Fertil. Steril. 2009, 91, 687–693. [Google Scholar] [CrossRef]
- Chang, K.-K.; Liu, L.-B.; Jin, L.-P.; Zhang, B.; Mei, J.; Li, H.; Wei, C.-Y.; Zhou, W.-J.; Zhu, X.-Y.; Shao, J.; et al. IL-27 triggers IL-10 production in Th17 cells via a c-Maf/RORγt/Blimp-1 signal to promote the progression of endometriosis. Cell Death Dis. 2017, 8, e2666. [Google Scholar] [CrossRef]
- Lee, M.-Y.; Kim, S.H.; Oh, Y.S.; Heo, S.-H.; Kim, K.-H.; Chae, H.D.; Kim, C.-H.; Kang, B.M. Role of interleukin-32 in the pathogenesis of endometriosis: In vitro, human and transgenic mouse data. Hum. Reprod. 2018, 33, 807–816. [Google Scholar] [CrossRef]
- Urata, Y.; Osuga, Y.; Akiyama, I.; Nagai, M.; Izumi, G.; Takamura, M.; Hasegawa, A.; Harada, M.; Hirata, T.; Hirota, Y.; et al. Interleukin-4 and prostaglandin E2 synergistically up-regulate 3β-hydroxysteroid dehydrogenase type 2 in endometrioma stromal cells. J. Clin. Endocrinol. Metab. 2013, 98, 1583–1590. [Google Scholar] [CrossRef]
- Veillat, V.; Carli, C.; Metz, C.N.; Al-Abed, Y.; Naccache, P.H.; Akoum, A. Macrophage migration inhibitory factor elicits an angiogenic phenotype in human ectopic endometrial cells and triggers the production of major angiogenic factors via CD44, CD74, and MAPK signaling pathways. J. Clin. Endocrinol. Metab. 2010, 95, E403–E412. [Google Scholar] [CrossRef]
- Tan, A.; Luo, R.; Liang, H.; Li, M.; Ruan, P. Bioinformatics approach reveals the key role of C-X-C motif chemokine receptor 2 in endometriosis development. Mol. Med. Rep. 2018, 18, 2841–2849. [Google Scholar] [CrossRef]
- Kuessel, L.; Wenzl, R.; Proestling, K.; Balendran, S.; Pateisky, P.; Yotova, I.; Yerlikaya, G.; Streubel, B.; Husslein, H. Soluble VCAM-1/soluble ICAM-1 ratio is a promising biomarker for diagnosing endometriosis. Hum. Reprod. 2017, 32, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-Q.; Li, H.-P.; Meng, Y.-H.; Wang, X.-Q.; Zhu, X.-Y.; Mei, J.; Li, D.-J. Chemokine CCL2 enhances survival and invasiveness of endometrial stromal cells in an autocrine manner by activating Akt and MAPK/Erk1/2 signal pathway. Fertil. Steril. 2012, 97, 919–929.e1. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Y.-D.; Chen, S.-L.; Chen, X.; Ye, D.-S.; Zhou, X.-Y.; Zhe, J.; Zhang, J. Down-regulation of long non-coding RNA MALAT1 inhibits granulosa cell proliferation in endometriosis by up-regulating P21 via activation of the ERK/MAPK pathway. Mol. Hum. Reprod. 2019, 25, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Darcha, C. Co-operation between the AKT and ERK signaling pathways may support growth of deep endometriosis in a fibrotic microenvironment in vitro. Hum. Reprod. 2015, 30, 1606–1616. [Google Scholar] [CrossRef]
- Aznaurova, Y.B.; Zhumataev, M.B.; Roberts, T.K.; Aliper, A.M.; Zhavoronkov, A.A. Molecular aspects of development and regulation of endometriosis. Reprod. Biol. Endocrinol. 2014, 12, 50. [Google Scholar] [CrossRef]
- McKinnon, B.D.; Kocbek, V.; Nirgianakis, K.; Bersinger, N.A.; Mueller, M.D. Kinase signalling pathways in endometriosis: Potential targets for non-hormonal therapeutics. Hum. Reprod. Update 2016, 22, 382–403. [Google Scholar] [CrossRef]
- Riccio, L.d.G.C.; Santulli, P.; Marcellin, L.; Abrão, M.S.; Batteux, F.; Chapron, C. Immunology of endometriosis. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 50, 39–49. [Google Scholar] [CrossRef]
- Parazzini, F.; Viganò, P.; Candiani, M.; Fedele, L. Diet and endometriosis risk: A literature review. Reprod. Biomed. Online 2013, 26, 323–336. [Google Scholar] [CrossRef]
- Jurkiewicz-Przondziono, J.; Lemm, M.; Kwiatkowska-Pamuła, A.; Ziółko, E.; Wójtowicz, M.K. Influence of diet on the risk of developing endometriosis. Ginekol. Pol. 2017, 88, 96–102. [Google Scholar] [CrossRef]
- Yamamoto, A.; Harris, H.R.; Vitonis, A.F.; Chavarro, J.E.; Missmer, S.A. A prospective cohort study of meat and fish consumption and endometriosis risk. Am. J. Obstet. Gynecol. 2018, 219, 178.e1–178.e10. [Google Scholar] [CrossRef]
- Simmen, R.C.M.; Kelley, A.S. Seeing red: Diet and endometriosis risk. Ann. Transl. Med. 2018, 6, S119. [Google Scholar] [CrossRef] [PubMed]
- Nodler, J.L.; Harris, H.R.; Chavarro, J.E.; Frazier, A.L.; Missmer, S.A. Dairy consumption during adolescence and endometriosis risk. Am. J. Obstet. Gynecol. 2019, 222, 257. [Google Scholar] [CrossRef] [PubMed]
- Huijs, E.; Nap, A. The effects of nutrients on symptoms in women with endometriosis: A systematic review. Reprod. Biomed. Online 2020, 41, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, E.; Kazimierczak, R.; Marszałek, K.; Drela, N.; Kiernozek, E.; Toomik, P.; Matt, D.; Luik, A.; Rembiałkowska, E. The nutritive value of organic and conventional white cabbage (Brassica oleracea L. var. capitata) and anti-apoptotic activity in gastric adenocarcinoma cells of sauerkraut juice produced therof. J. Agric. Food Chem. 2017, 65, 8171–8183. [Google Scholar] [CrossRef]
- Wieczorek, M.N.; Walczak, M.; Skrzypczak-Zielińska, M.; Jeleń, H.H. Bitter taste of Brassica vegetables: The role of genetic factors, receptors, isothiocyanates, glucosinolates, and flavor context. Crit. Rev. Food Sci. Nutr. 2018, 58, 3130–3140. [Google Scholar] [CrossRef]
- Russo, M.; Spagnuolo, C.; Russo, G.L.; Skalicka-Woźniak, K.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S.F.; Nabavi, S.M. Nrf2 targeting by sulforaphane: A potential therapy for cancer treatment. Crit. Rev. Food Sci. Nutr. 2018, 58, 1391–1405. [Google Scholar] [CrossRef]
- Bayat Mokhtari, R.; Baluch, N.; Homayouni, T.S.; Morgatskaya, E.; Kumar, S.; Kazemi, P.; Yeger, H. The role of Sulforaphane in cancer chemoprevention and health benefits: A mini-review. J. Cell Commun. Signal. 2018, 12, 91–101. [Google Scholar] [CrossRef]
- Burčul, F.; Generalić Mekinić, I.; Radan, M.; Rollin, P.; Blažević, I. Isothiocyanates: Cholinesterase inhibiting, antioxidant, and anti-inflammatory activity. J. Enzyme Inhib. Med. Chem. 2018, 33, 577–582. [Google Scholar] [CrossRef]
- Agerbirk, N.; Olsen, C.E. Glucosinolate structures in evolution. Phytochemistry 2012, 77, 16–45. [Google Scholar] [CrossRef]
- Bhat, R.; Vyas, D. Myrosinase: Insights on structural, catalytic, regulatory, and environmental interactions. Crit. Rev. Biotechnol. 2019, 39, 508–523. [Google Scholar] [CrossRef]
- Nugrahedi, P.Y.; Verkerk, R.; Widianarko, B.; Dekker, M. A Mechanistic Perspective on Process-Induced Changes in Glucosinolate Content in Brassica Vegetables: A Review. Crit. Rev. Food Sci. Nutr. 2015, 55, 823–838. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.; Carrascosa, C.; Raposo, A. Influence of Different Cooking Methods on the Concentration of Glucosinolates and Vitamin C in Broccoli. Food Bioprocess Technol. 2017, 10, 1387–1411. [Google Scholar] [CrossRef]
- Bessler, H.; Djaldetti, M. Broccoli and human health: Immunomodulatory effect of sulforaphane in a model of colon cancer. Int. J. Food Sci. Nutr. 2018, 69, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Houghton, C.A.; Fassett, R.G.; Coombes, J.S. Sulforaphane and other nutrigenomic Nrf2 activators: Can the clinician’s expectation be matched by the reality? Oxid. Med. Cell. Longev. 2016, 2016, 785186. [Google Scholar] [CrossRef] [PubMed]
- Eren, E.; Tufekci, K.U.; Isci, K.B.; Tastan, B.; Genc, K.; Genc, S. Sulforaphane inhibits lipopolysaccharide-induced inflammation, cytotoxicity, oxidative stress, and miR-155 expression and switches to mox phenotype through activating extracellular signal-regulated kinase 1/2-nuclear factor erythroid 2-related factor 2/An. Front. Immunol. 2018, 23, 9–36. [Google Scholar]
- Greaney, A.J.; Maier, N.K.; Leppla, S.H.; Moayeri, M. Sulforaphane inhibits multiple inflammasomes through an Nrf2-independent mechanism. J. Leukoc. Biol. 2016, 99, 189–199. [Google Scholar] [CrossRef]
- Okamura, Y.; Watari, M.; Jerud, E.S.; Young, D.W.; Ishizaka, S.T.; Rose, J.; Chow, J.C.; Strauss, J.F. The extra domain A of fibronectin activates Toll-like Receptor 4. J. Biol. Chem. 2001, 276, 10229–10233. [Google Scholar] [CrossRef]
- O’Callaghan, P.; Zhang, X.; Li, J.P. Heparan sulfate proteoglycans as relays of neuroinflammation. J. Histochem. Cytochem. 2018, 66, 305–319. [Google Scholar] [CrossRef]
- Youn, H.S.; Kim, Y.S.; Park, Z.Y.; Kim, S.Y.; Choi, N.Y.; Joung, S.M.; Seo, J.A.; Lim, K.-M.; Kwak, M.-K.; Hwang, D.H.; et al. Sulforaphane Suppresses Oligomerization of TLR4 in a Thiol-Dependent Manner. J. Immunol. 2010, 184, 305–319. [Google Scholar] [CrossRef]
- Mazarakis, N.; Snibson, K.; Licciardi, P.V.; Karagiannis, T.C. The potential use of L-sulforaphane for the treatment of chronic inflammatory diseases: A review of the clinical evidence. Clin. Nutr. 2019, 39, 664–675. [Google Scholar] [CrossRef]
- Zhao, Z.; Liao, G.; Zhou, Q.; Lv, D.; Holthfer, H.; Zou, H. Sulforaphane attenuates contrast-induced nephropathy in rats via Nrf2/HO-1 pathway. Oxid. Med. Cell. Longev. 2016, 2016, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Hong, Y.; Lv, Y. Sulforaphane attenuates endometriosis in rat models through inhibiting PI3K/Akt signaling pathway. Dose-Response 2019, 17, 1559325819855538. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Z.; Lu, X.; Meng, J.; Qin, X.; Jiang, J. Anti-nociceptive and anti-inflammatory effects of sulforaphane on sciatic endometriosis in a rat model. Neurosci. Lett. 2020, 723, 134858. [Google Scholar] [CrossRef] [PubMed]
- Baenas, N.; Piegholdt, S.; Schloesser, A.; Moreno, D.A.; García-Viguera, C.; Rimbach, G.; Wagner, A.E. Metabolic activity of radish sprouts derived isothiocyanates in drosophila melanogaster. Int. J. Mol. Sci. 2016, 17, 251. [Google Scholar] [CrossRef] [PubMed]
- Pawlik, A.; Wała, M.; Hać, A.; Felczykowska, A.; Herman-Antosiewicz, A. Sulforaphene, an isothiocyanate present in radish plants, inhibits proliferation of human breast cancer cells. Phytomedicine 2017, 29, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pocasap, P.; Weerapreeyakul, N.; Barusrux, S. Cancer preventive effect of Thai rat-tailed radish (Raphanus sativus L. var. caudatus Alef). J. Funct. Foods 2013, 5, 1372–1381. [Google Scholar] [CrossRef]
- Baenas, N.; Suárez-Martínez, C.; García-Viguera, C.; Moreno, D.A. Bioavailability and new biomarkers of cruciferous sprouts consumption. Food Res. Int. 2017. [Google Scholar] [CrossRef]
- Westphal, A.; Riedl, K.M.; Cooperstone, J.L.; Kamat, S.; Balasubramaniam, V.M.; Schwartz, S.J.; Böhm, V. High-pressure processing of broccoli sprouts: Influence on bioactivation of glucosinolates to isothiocyanates. J. Agric. Food Chem. 2017, 65, 8578–8585. [Google Scholar] [CrossRef]
- Clarke, J.D.; Hsu, A.; Riedl, K.; Bella, D.; Schwartz, S.J.; Stevens, J.F.; Ho, E. Bioavailability and inter-conversion of sulforaphane and erucin in human subjects consuming broccoli sprouts or broccoli supplement in a cross-over study design. Pharmacol. Res. 2011, 64, 456–463. [Google Scholar] [CrossRef]
- Cho, H.J.; Lee, K.W.; Yoon Park, J.H. Erucin exerts anti-inflammatory properties in murine macrophages and mouse skin: Possible mediation through the inhibition of NFκB signaling. Int. J. Mol. Sci. 2013, 14, 20564–20577. [Google Scholar] [CrossRef]
- Luang-In, V.; Deeseenthum, S.; Udomwong, P.; Saengha, W.; Gregori, M. Formation of sulforaphane and iberin products from Thai cabbage fermented by myrosinase-positive bacteria. Molecules 2018, 23, 955. [Google Scholar] [CrossRef] [PubMed]
- Baenas, N.; Marhuenda, J.; García-Viguera, C.; Zafrilla, P.; Moreno, D.A. Influence of cooking methods on glucosinolates and isothiocyanates content in novel cruciferous foods. Foods 2019, 8, 257. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Nakashima, F.; Honda, K.; Lu, Y.J.; Kondo, T.; Ushida, Y.; Aizawa, K.; Suganuma, H.; Oe, S.; Tanaka, H.; et al. Toll-like receptors as a target of food-derived anti-inflammatory compounds. J. Biol. Chem. 2014, 289, 32757–32772. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.J.; Chen, B.H.; Inbaraj, B.S.; Chien, J.T. Preparation of allyl isothiocyanate nanoparticles, their anti-inflammatory activity towards RAW 264.7 macrophage cells and anti-proliferative effect on HT1376 bladder cancer cells. J. Sci. Food Agric. 2019, 99, 3106–3116. [Google Scholar] [CrossRef] [PubMed]
- Rajakumar, T.; Pugalendhi, P.; Jayaganesh, R.; Ananthakrishnan, D.; Gunasekaran, K. Effect of allyl isothiocyanate on NF-κB signaling in 7,12-dimethylbenz(a)anthracene and N-methyl-N-nitrosourea-induced mammary carcinogenesis. Breast Cancer 2018, 25, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Rajakumar, T.; Pugalendhi, P.; Thilagavathi, S.; Ananthakrishnan, D.; Gunasekaran, K. Allyl isothiocyanate, a potent chemopreventive agent targets AhR/Nrf2 signaling pathway in chemically induced mammary carcinogenesis. Mol. Cell. Biochem. 2018, 437, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bessede, A.; Gargaro, M.; Pallotta, M.T.; Matino, D.; Servillo, G.; Brunacci, C.; Bicciato, S.; Mazza, E.M.C.; Macchiarulo, A.; Vacca, C.; et al. Aryl hydrocarbon receptor control of a disease tolerance defence pathway. Nature 2014, 511, 184–190. [Google Scholar] [CrossRef]
- Murray, I.A.; Patterson, A.D.; Perdew, G.H. Aryl hydrocarbon receptor ligands in cancer: Friend and foe. Nat. Rev. Cancer 2014, 14, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, Y.; de Dios Rosado, J.; Vega, L.; Elizondo, G.; Estrada-Muñiz, E.; Saavedra, R.; Juárez, I.; Rodríguez-Sosa, M. The unexpected role for the aryl hydrocarbon receptor on susceptibility to experimental toxoplasmosis. J. Biomed. Biotechnol. 2010, 2010, 505694. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.M.; Park, B.; Dang, Y.M.; Kim, S.Y.; Seo, H.Y. Simultaneous direct determination of 15 glucosinolates in eight Brassica species by UHPLC-Q-Orbitrap-MS. Food Chem. 2019, 282, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Grose, K.R.; Bjeldanes, L.F. Oligomerization of Indole-3-carbinol in aqueous acid. Chem. Res. Toxicol. 1992, 5, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.Y.; Pham, Q.; Kim, Y.S. Elucidating the role of CD84 and AHR in modulation of LPS-induced cytokines production by cruciferous vegetable-derived compounds indole-3-carbinol and 3,3′-diindolylmethane. Int. J. Mol. Sci. 2018, 19, 339. [Google Scholar] [CrossRef] [PubMed]
- Hajra, S.; Patra, A.R.; Basu, A.; Bhattacharya, S. Prevention of doxorubicin (DOX)-induced genotoxicity and cardiotoxicity: Effect of plant derived small molecule indole-3-carbinol (I3C) on oxidative stress and inflammation. Biomed. Pharmacother. 2018, 101, 228–243. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, S.; Memarian, A.; Sedighi, S.; Behnampour, N.; Yazdani, Y. Immunoregulatory effects of indole-3-carbinol on monocyte-derived macrophages in systemic lupus erythematosus: A crucial role for aryl hydrocarbon receptor. Autoimmunity 2018, 51, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Seon, M.R.; Lee, Y.M.; Kim, J.; Kim, J.-K.; Kim, S.G.; Park, J.H.Y. 3,3′-Diindolylmethane suppresses the inflammatory response to lipopolysaccharide in murine macrophages. J. Nutr. 2008, 138, 17–23. [Google Scholar] [CrossRef]
- Jeon, E.J.; Davaatseren, M.; Hwang, J.T.; Park, J.H.; Hur, H.J.; Lee, A.S.; Sung, M.J. Effect of oral administration of 3,3′-Diindolylmethane on dextran sodium sulfate-induced acute colitis in mice. J. Agric. Food Chem. 2016, 64, 7702–7709. [Google Scholar] [CrossRef]
- Morales-Prieto, D.M.; Herrmann, J.; Osterwald, H.; Kochhar, P.S.; Schleussner, E.; Markert, U.R.; Oettel, M. Comparison of dienogest effects upon 3,3′–diindolylmethane supplementation in models of endometriosis and clinical cases. Reprod. Biol. 2018, 18, 252–258. [Google Scholar] [CrossRef]
- Tai, A.; Fukunaga, K.; Ohno, A.; Ito, H. Antioxidative properties of ascorbigen in using multiple antioxidant assays. Biosci. Biotechnol. Biochem. 2014, 78, 1723–1730. [Google Scholar] [CrossRef]
- Martinez-Villaluenga, C.; Peñas, E.; Sidro, B.; Ullate, M.; Frias, J.; Vidal-Valverde, C. White cabbage fermentation improves ascorbigen content, antioxidant and nitric oxide production inhibitory activity in LPS-induced macrophages. LWT Food Sci. Technol. 2012, 46, 77–83. [Google Scholar] [CrossRef]
- Magni, F.; Sarto, C.; Ticozzi, D.; Soldi, M.; Bosso, N.; Mocarelli, P.; Kienle, M.G. Proteomic knowledge of human aquaporins. Proteomics 2006, 6, 5637–5649. [Google Scholar] [CrossRef]
- Dajani, S.; Saripalli, A.; Sharma-Walia, N. Water transport proteins–aquaporins (AQPs) in cancer biology. Oncotarget 2018, 9, 36392–36405. [Google Scholar] [CrossRef] [PubMed]
- Agre, P.; Kozono, D. Aquaporin water channels: Molecular mechanisms for human diseases. FEBS Letters 2003, 555, 72–78. [Google Scholar] [CrossRef]
- Madeira, A.; Moura, T.F.; Soveral, G. Detecting aquaporin function and regulation. Front. Chem. 2016, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Verkman, A.S. Aquaporins in clinical medicine. Annu. Rev. Med. 2012, 63, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Jiang, Z.; Bazer, F.W.; Johnson, G.A.; Burghardt, R.C.; Wu, G. Aquaporins in the female reproductive system of mammals. Front. Biosci. 2015, 20, 838–871. [Google Scholar]
- Zhang, D.; Tan, Y.J.; Qu, F.; Sheng, J.Z.; Huang, H.F. Functions of water channels in male and female reproductive systems. Mol. Aspects Med. 2012, 33, 676–690. [Google Scholar] [CrossRef]
- Meli, R.; Pirozzi, C.; Pelagalli, A. New perspectives on the potential role of aquaporins (AQPs) in the physiology of inflammation. Front. Physiol. 2018, 16, 101. [Google Scholar] [CrossRef]
- Riemma, G.; Laganà, A.S.; Schiattarella, A.; Garzon, S.; Cobellis, L.; Autiero, R.; Licciardi, F.; Corte, L.D.; La Verde, M.; De Franciscis, P. Ion channels in the pathogenesis of endometriosis: A cutting-edge point of view. Int. J. Mol. Sci. 2020, 21, 1114. [Google Scholar] [CrossRef]
- Shu, C.; Shu, Y.; Gao, Y.; Chi, H.; Han, J. Inhibitory effect of AQP1 silencing on adhesion and angiogenesis in ectopic endometrial cells of mice with endometriosis through activating the Wnt signaling pathway. Cell Cycle 2019, 18, 2026–2039. [Google Scholar] [CrossRef]
- Skowronski, M.T. Distribution and quantitative changes in amounts of aquaporin 1, 5 and 9 in the pig uterus during the estrous cycle and early pregnancy. Reprod. Biol. Endocrinol. 2010, 8, 1–11. [Google Scholar] [CrossRef]
- Jiang, X.X.; Wu, R.J.; Xu, K.H.; Zhou, C.Y.; Guo, X.Y.; Sun, Y.L.; Lin, J. Immunohistochemical detection of aquaporin expression in eutopic and ectopic endometria from women with endometriomas. Fertil. Steril. 2010, 94, 1229–1234. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.X.; Fei, X.W.; Zhao, L.; Ye, X.L.; Xin, L.B.; Qu, Y.; Xu, K.H.; Wu, R.J.; Lin, J. Aquaporin 5 plays a role in estrogen-induced ectopic implantation of endometrial stromal cells in endometriosis. PLoS ONE 2015, 10, e0145290. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.X.; Xu, K.H.; Ma, J.Y.; Tian, Y.H.; Guo, X.Y.; Lin, J.; Wu, R.J. Reduced migration of Ishikawa cells associated with downregulation of aquaporin-5. Oncol. Lett. 2012, 4, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Park, J.H.; Yoon, J.-K.; Yoon, J.S.; Kim, J.S.; Lee, J.H.; Yun, B.H.; Park, J.H.; Seo, S.K.; Cho, S.; et al. Potential roles of aquaporin 9 in the pathogenesis of endometriosis. MHR Basic Sci. Reprod. Med. 2019, 125, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.G.; Li, C.F.; Liu, M.; Chen, X.F.; Shuai, K.; Kong, X.; Lv, L.; Mei, Z. chuan Aquaporin 9 is down-regulated in hepatocellular carcinoma and its over-expression suppresses hepatoma cell invasion through inhibiting epithelial-to-mesenchymal transition. Cancer Lett. 2016, 378, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Tyteca, D.; Nishino, T.; Debaix, H.; Van Smissen, P.D.; N’Kuli, F.; Hoffmann, D.; Cnops, Y.; Rabolli, V.; Van Loo, G.; Beyaert, R.; et al. Regulation of macrophage motility by the water channel aquaporin-1: Crucial role of M0/M2 phenotype switch. PLoS ONE 2015, 10, e0117398. [Google Scholar] [CrossRef]
- Li, B.; Liu, C.; Tang, K.; Dong, X.; Xue, L.; Su, G.; Zhang, W.; Jin, Y. Aquaporin-1 attenuates macrophage-mediated inflammatory responses by inhibiting p38 mitogen-activated protein kinase activation in lipopolysaccharide-induced acute kidney injury. Inflamm. Res. 2019, 68, 1035–1047. [Google Scholar] [CrossRef]
- Li, Y.; Lu, H.; Lv, X.; Tang, Q.; Li, W.; Zhu, H.; Long, Y. Blockade of Aquaporin 4 Inhibits Irradiation-Induced Pulmonary Inflammation and Modulates Macrophage Polarization in Mice. Inflammation 2018, 41, 2196–2203. [Google Scholar] [CrossRef]
- Kitchen, P.; Day, R.E.; Salman, M.M.; Conner, M.T.; Bill, R.M.; Conner, A.C. Beyond water homeostasis: Diverse functional roles of mammalian aquaporins. Biochim. Biophys. Acta Gen. Subj. 2015, 1850, 2410–2421. [Google Scholar] [CrossRef]
- Lin, Y.H.; Chen, Y.H.; Chang, H.Y.; Au, H.K.; Tzeng, C.R.; Huang, Y.H. Chronic niche inflammation in endometriosis-associated infertility: Current understanding and future therapeutic strategies. Int. J. Mol. Sci. 2018, 19, 2385. [Google Scholar] [CrossRef]
- Martínez-Ballesta, M.d.C.; Bou, G.; Carvajal, M. Aquaporins as targets of pharmacological plant-derived compounds. Phytochem. Rev. 2014, 13, 573–586. [Google Scholar] [CrossRef]
- Tanimura, Y.; Hiroaki, Y.; Fujiyoshi, Y. Acetazolamide reversibly inhibits water conduction by aquaporin-4. J. Struct. Biol. 2009, 166, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, H.; Huber, V.J.; Tsujita, M.; Nakada, T. Pretreatment with a novel aquaporin 4 inhibitor, TGN-020, significantly reduces ischemic cerebral edema. Neurol. Sci. 2011, 32, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Brooks, H.L.; Regan, J.W.; Yool, A.J. Inhibition of aquaporin-1 water permeability by tetraethylammonium: Involvement of the loop E pore region. Mol. Pharmacol. 2000, 57, 1021–1026. [Google Scholar] [PubMed]
- Detmers, F.J.M.; De Groot, B.L.; Müller, E.M.; Hinton, A.; Konings, I.B.M.; Sze, M.; Flitsch, S.L.; Grubmüller, H.; Deen, P.M.T. Quaternary ammonium compounds as water channel blockers: Specificity, potency, and site of action. J. Biol. Chem. 2006, 281, 14207–14214. [Google Scholar] [CrossRef] [PubMed]
- Yool, A.J.; Morelle, J.; Cnops, Y.; Verbavatz, J.M.; Campbell, E.M.; Beckett, E.A.H.; Booker, G.W.; Flynn, G.; Devuyst, O. AqF026 is a pharmacologic agonist of the water channel aquaporin-1. J. Am. Soc. Nephrol. 2013, 24, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wang, X.; Chang, Y.; Zhang, J.; Song, Q.; Yu, H.; Li, X. Acetazolamide inhibits osmotic water permeability by interaction with aquaporin-1. Anal. Biochem. 2006, 350, 165–170. [Google Scholar] [CrossRef]
- Huber, V.J.; Tsujita, M.; Yamazaki, M.; Sakimura, K.; Nakada, T. Identification of arylsulfonamides as Aquaporin 4 inhibitors. Bioorg. Med. Chem. Lett. 2007, 17, 1270–1273. [Google Scholar] [CrossRef]
- Huber, V.J.; Tsujita, M.; Kwee, I.L.; Nakada, T. Inhibition of Aquaporin 4 by antiepileptic drugs. Bioorg. Med. Chem. 2009, 17, 418–424. [Google Scholar] [CrossRef]
- Devuyst, O.; Yool, A.J. Aquaporin-1: New developments and perspectives for peritoneal dialysis. Perit. Dial. Int. 2010, 30, 135–141. [Google Scholar] [CrossRef]
- Szczepańska, M.; Koźlik, J.; Skrzypczak, J.; Mikołajczyk, M. Oxidative stress may be a piece in the endometriosis puzzle. Fertil. Steril. 2003, 79, 1288–1293. [Google Scholar] [CrossRef]
- Bestetti, S.; Medraño-Fernandez, I.; Galli, M.; Ghitti, M.; Bienert, G.P.; Musco, G.; Orsi, A.; Rubartelli, A.; Sitia, R. A persulfidation-based mechanism controls aquaporin-8 conductance. Sci. Adv. 2018, 4, eaar5770. [Google Scholar] [CrossRef] [PubMed]
- Prata, C.; Facchini, C.; Leoncini, E.; Lenzi, M.; Maraldi, T.; Angeloni, C.; Zambonin, L.; Hrelia, S.; Fiorentini, D. Sulforaphane modulates AQP8-linked redox signalling in leukemia cells. Oxid. Med. Cell. Longev. 2018, 2018, 4125297. [Google Scholar] [CrossRef]
- Kitawaki, J.; Kado, N.; Ishihara, H.; Koshiba, H.; Kitaoka, Y.; Honjo, H. Endometriosis: The pathophysiology as an estrogen-dependent disease. J. Steroid Biochem. Mol. Biol. 2002, 83, 149–155. [Google Scholar] [CrossRef]
- Oliveira, C.A.; Carnes, K.; França, L.R.; Hermo, L.; Hess, R.A. Aquaporin-1 and -9 are differentially regulated by oestrogen in the efferent ductule epithelium and initial segment of the epididymis. Biol. Cell 2005, 97, 385–395. [Google Scholar] [CrossRef]
- Zou, L.B.; Zhang, R.J.; Tan, Y.J.; Ding, G.L.; Shi, S.; Zhang, D.; He, R.H.; Liu, A.X.; Wang, T.T.; Leung, P.C.K.; et al. Identification of estrogen response element in the aquaporin-2 gene that mediates estrogen-induced cell migration and invasion in human endometrial carcinoma. J. Clin. Endocrinol. Metab. 2011, 96, E1399–E1408. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Wang, L.; Zhang, Z.; Chen, F.; Wu, Q.; Li, L. Sulforaphane-induced metabolomic responses with epigenetic changes in estrogen receptor positive breast cancer cells. FEBS Open Bio 2018, 8, 2022–2034. [Google Scholar] [CrossRef] [PubMed]
- Marconett, C.N.; Singhal, A.K.; Sundar, S.N.; Firestone, G.L. Indole-3-carbinol disrupts estrogen receptor-alpha dependent expression of insulin-like growth factor-1 receptor and insulin receptor substrate-1 and proliferation of human breast cancer cells. Mol. Cell. Endocrinol. 2012, 363, 74–84. [Google Scholar] [CrossRef]
- Meng, Q.; Yuan, F.; Goldberg, I.D.; Rosen, E.M.; Auborn, K.; Fan, S. Indole-3-Carbinol Is a Negative Regulator of Estrogen Receptor-α Signaling in Human Tumor Cells. J. Nutr. 2000, 130, 2927–2931. [Google Scholar] [CrossRef]
- Amare, D.E.; Bovee, T.F.H.; Mulder, P.P.J.; Hamers, A.; Hoogenboom, R.L.A.P. Acid condensation products of indole-3-carbinol and their in-vitro (anti)estrogenic, (anti)androgenic and aryl hydrocarbon receptor activities. Arab. J. Chem. 2020, 13, 7199–7211. [Google Scholar] [CrossRef]
- Zhao, J.; Moore, A.N.; Clifton, G.L.; Dash, P.K. Sulforaphane enhances aquaporin-4 expression and decreases cerebral edema following traumatic brain injury. J. Neurosci. Res. 2005, 82, 499–506. [Google Scholar] [CrossRef]
- Dash, P.K.; Zhao, J.; Orsi, S.A.; Zhang, M.; Moore, A.N. Sulforaphane improves cognitive function administered following traumatic brain injury. Neurosci. Lett. 2009, 460, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Gasparyan, A.Y.; Ayvazyan, L.; Blackmore, H.; Kitas, G.D. Writing a narrative biomedical review: Considerations for authors, peer reviewers, and editors. Rheumatol. Int. 2011, 31, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Saracci, C.; Mahamat, M.; Jacquérioz, F. How to write a narrative literature review article? Rev. Med. Suisse 2019, 15, 1694–1698. [Google Scholar] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).