Geomagnetic Shielding Enhances Radiation Resistance by Promoting DNA Repair Process in Human Bronchial Epithelial Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
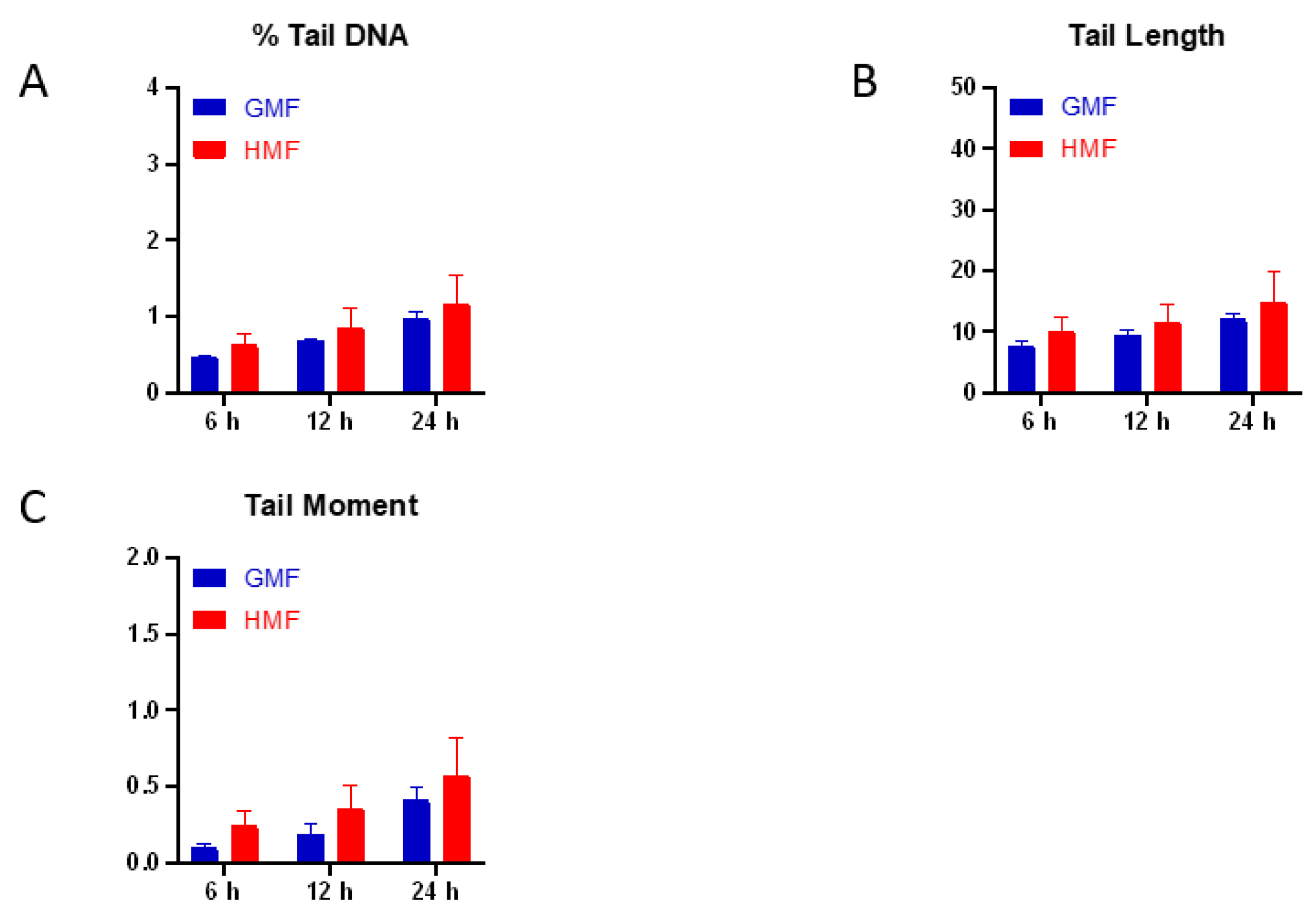
2.1. Geomagnetic Shielding Alone Does Not Affect DNA Integrity
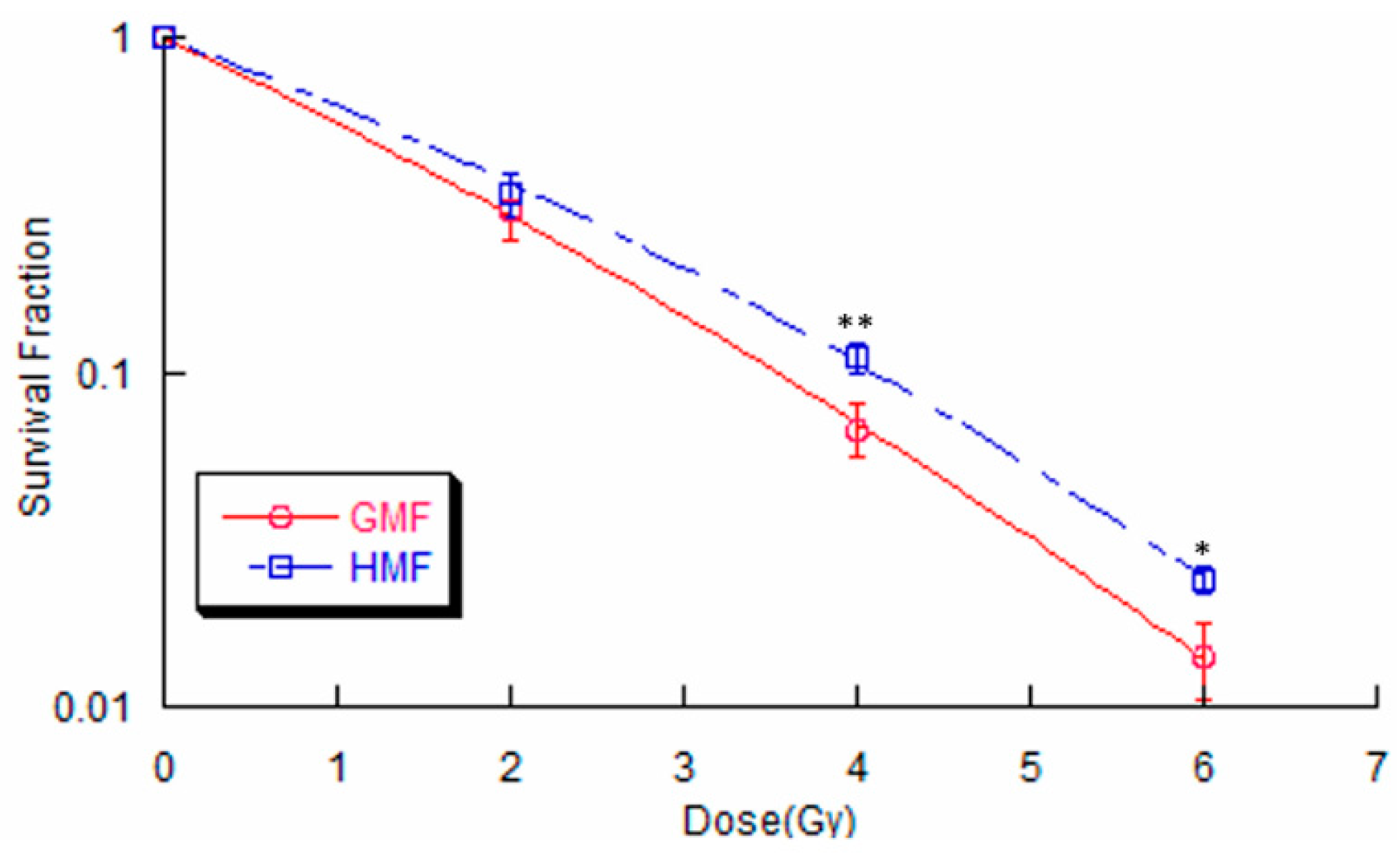
2.2. Geomagnetic Shielding Enhances Radiation Resistance in BEAS-2B Cells
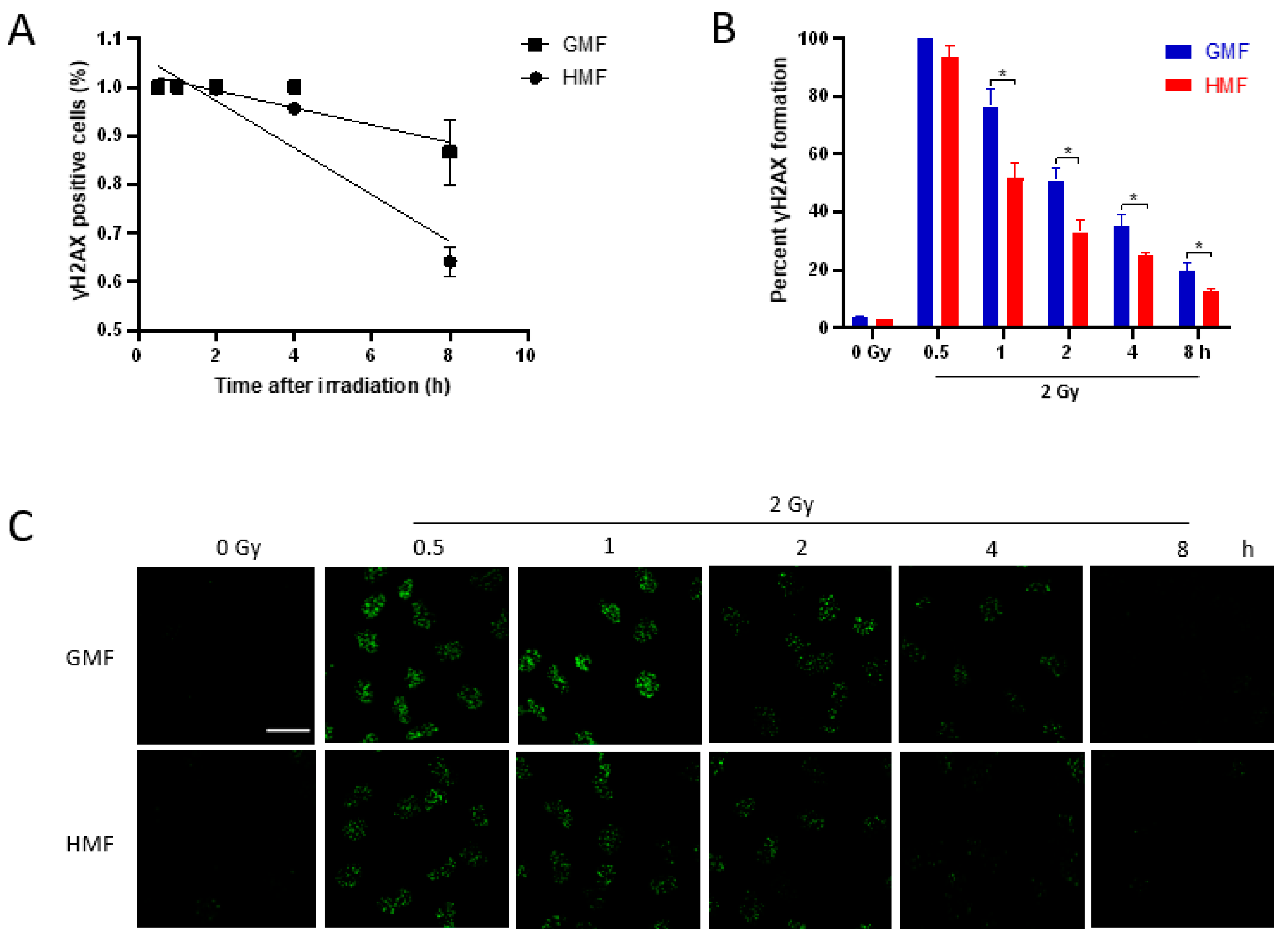
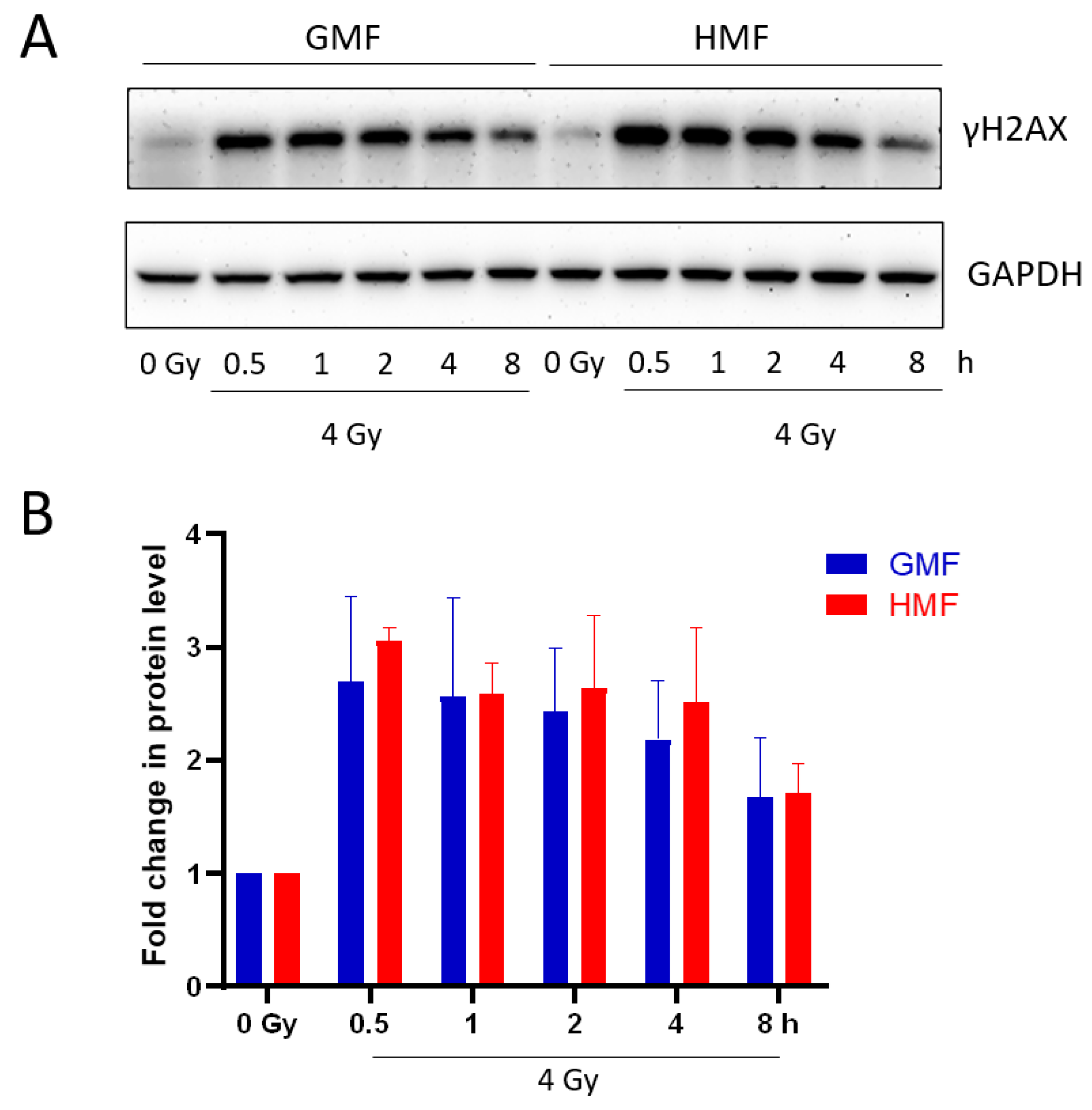
2.3. Geomagnetic Shielding Accelerates the Decline of IR Induced γH2AX Foci
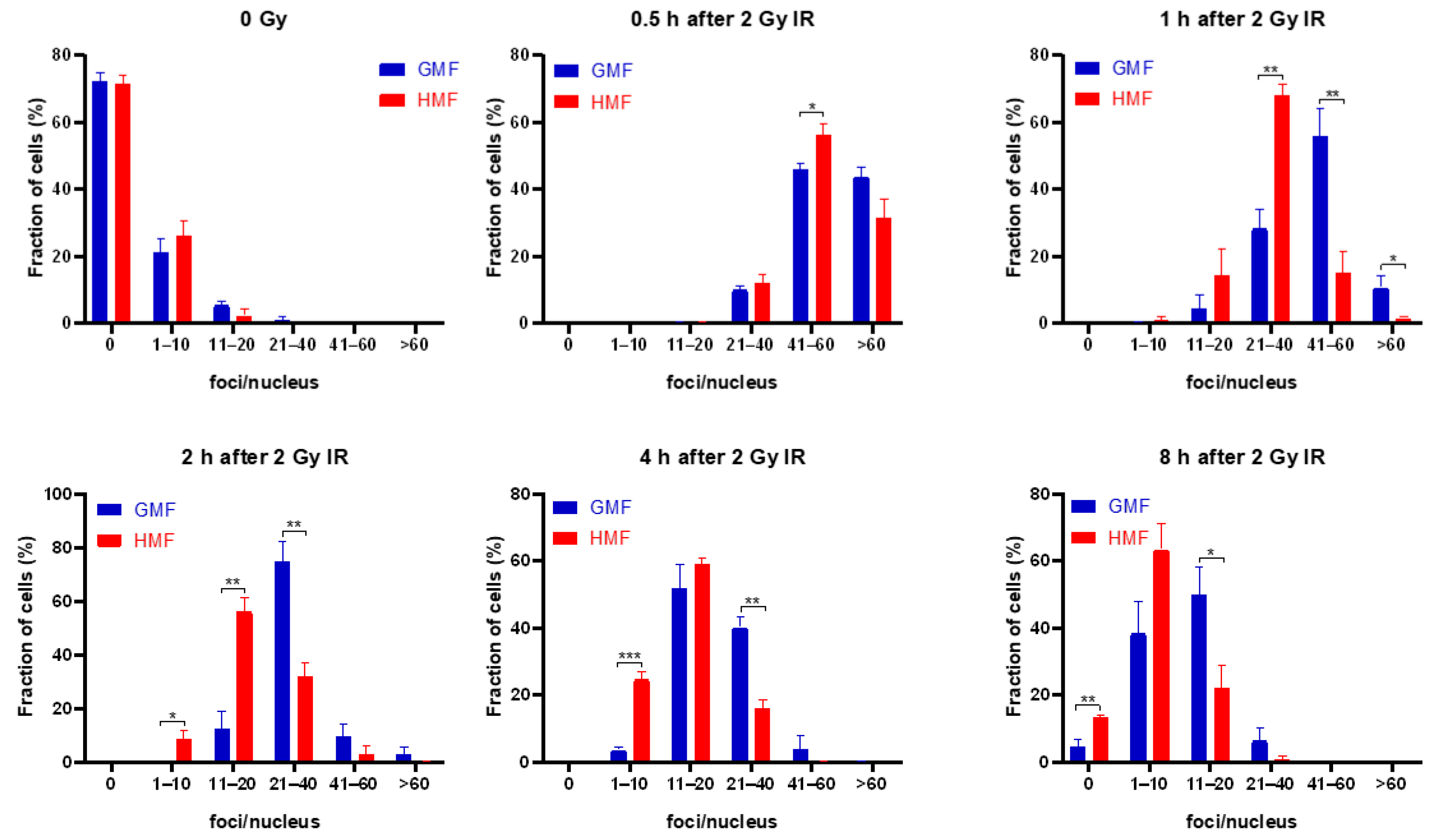
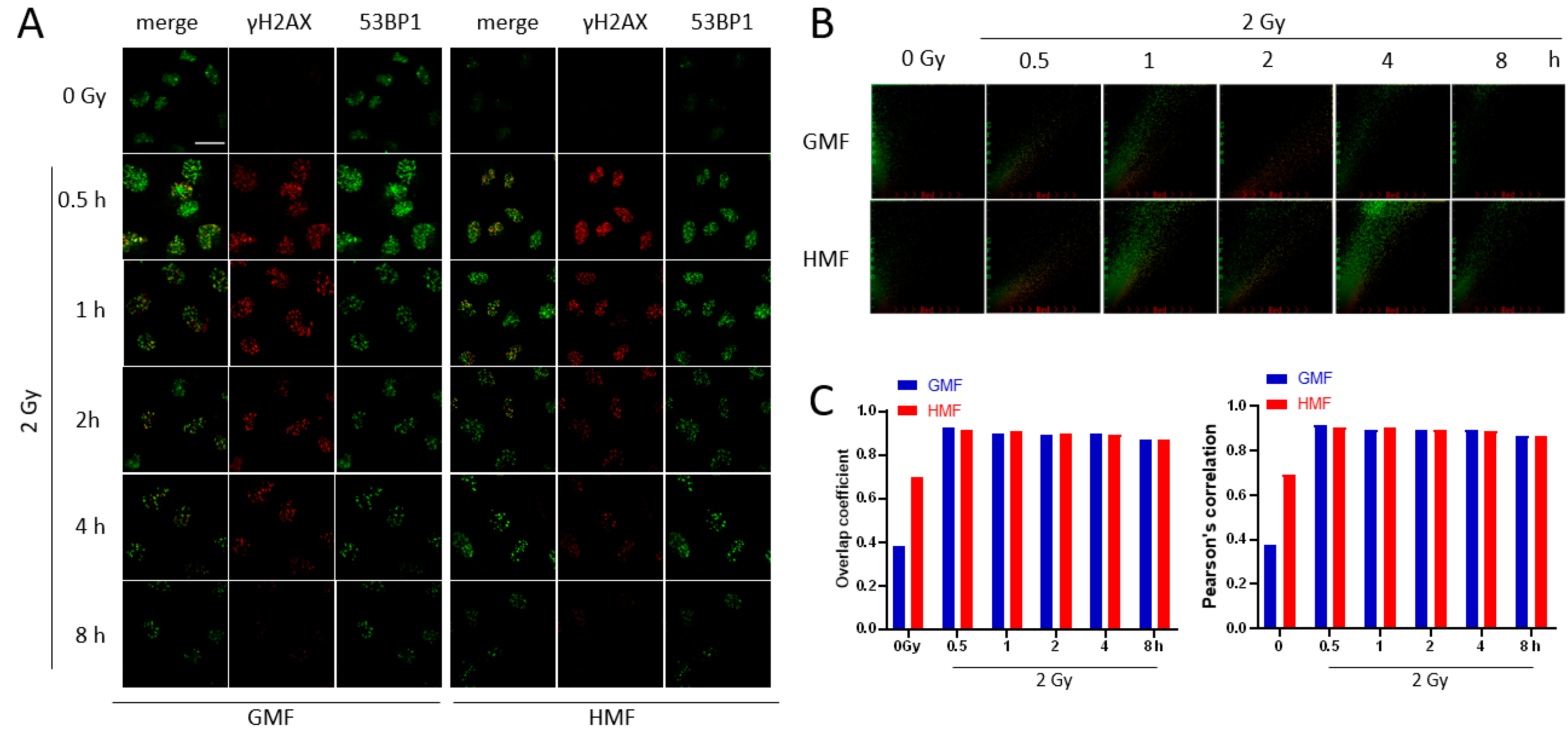
2.4. Geomagnetic Shielding Promotes DNA Repair Process at the IR Induced DSB Sites
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. The HMF Conditions
4.3. Irradiation
4.4. Alkaline Comet Assay
4.5. Cell Survival Assay
4.6. Immunofluorescent Staining
4.7. Western Blotting
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DSB | DNA double-strand break |
| GMF | Geomagnetic field |
| HMF | Hypomagnetic field |
| HR | Homologous recombination |
| IR | Ionizing radiation |
| NHEJ | Non-homologous end-joining |
| 53BP1 | p53 binding protein 1 |
| ROS | Reactive oxygen species |
| SF | Surviving fraction |
References
- Steinhilber, F.; Abreu, J.A.; Beer, J.; McCracken, K.G. Interplanetary magnetic field during the past 9300 years inferred from cosmogenic radionuclides. J. Geophys. Res. Space 2010, 115, A01104. [Google Scholar] [CrossRef]
- Berguig, M.S.; Hamoudi, M.; Lemouel, J.L.; Cohen, Y. Validate global mapping of internal lunar magnetic field. Arab. J. Geosci. 2013, 6, 1063–1072. [Google Scholar] [CrossRef]
- Watters, T.R.; McGovern, P.J.; Irwin, R.P. Hemispheres apart: The crustal dichotomy on Mars. Annu. Rev. Earth Planet. Sci. 2007, 35, 621–652. [Google Scholar] [CrossRef]
- Cucinotta, F.A. Space radiation risks for astronauts on multiple International Space Station missions. PLoS ONE 2014, 9, e96099. [Google Scholar] [CrossRef] [PubMed]
- Pearl, L.H.; Schierz, A.C.; Ward, S.E.; Al-Lazikani, B.; Pearl, F.M. Therapeutic opportunities within the DNA damage response. Nat. Rev. Cancer 2015, 15, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Kopanev, V.I.; Efimenko, G.D.; Shakula, A.V. Biological effect of a hypogeomagnetic environment on an organism. Biol. Bull. Acad. Sci. USSR 1979, 6, 289–298. [Google Scholar] [PubMed]
- Fu, J.P.; Mo, W.C.; Liu, Y.; Bartlett, P.F.; He, R.Q. Elimination of the geomagnetic field stimulates the proliferation of mouse neural progenitor and stem cells. Protein Cell 2016, 7, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Binhi, V.N.; Prato, F.S. Biological effects of the hypomagnetic field: An analytical review of experiments and theories. PLoS ONE 2017, 12, e0179340. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.; Choi, H.; Park, H.; Cho, B.; Kim, S.; Kim, J. Effects of a hypomagnetic field on DNA methylation during the differentiation of embryonic stem cells. Sci. Rep. 2019, 9, 1333. [Google Scholar] [CrossRef]
- Asashima, M.; Shimada, K.; Pfeiffer, C.J. Magnetic shielding induces early developmental abnormalities in the newt, Cynops pyrrhogaster. Bioelectromagnetics 1991, 12, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Mo, W.C.; Liu, Y.; Cooper, H.M.; He, R.Q. Altered development of Xenopus embryos in a hypogeomagnetic field. Bioelectromagnetics 2012, 33, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Bliss, V.L.; Heppner, F.H. Circadian activity rhythm influenced by near zero magnetic field. Nature 1976, 261, 411–412. [Google Scholar] [CrossRef] [PubMed]
- Zamoshchina, T.A.; Krivova, N.A.; Khodanovich, M.; Trukhanov, K.A.; Tukhvatulin, R.T.; Zaeva, O.B.; Zelenskaia, A.E.; Gul, E.V. Influence of simulated hypomagnetic environment in a far space flight on the rhythmic structure of rat’s behavior. Aviakosm. Ekolog. Med. 2012, 46, 17–23. [Google Scholar] [PubMed]
- Zhang, B.; Lu, H.; Xi, W.; Zhou, X.; Xu, S.; Zhang, K.; Jiang, J.; Li, Y.; Guo, A. Exposure to hypomagnetic field space for multiple generations causes amnesia in Drosophila melanogaster. Neurosci. Lett. 2004, 371, 190–195. [Google Scholar] [CrossRef]
- Xiao, Y.; Wang, Q.; Xu, M.L.; Jiang, J.C.; Li, B. Chicks incubated in hypomagnetic field need more exogenous noradrenaline for memory consolidation. Adv. Space Res. 2009, 44, 226–232. [Google Scholar] [CrossRef]
- Del Seppia, C.; Luschi, P.; Ghione, S.; Crosio, E.; Choleris, E.; Papi, F. Exposure to a hypogeomagnetic field or to oscillating magnetic fields similarly reduce stress-induced analgesia in C57 male mice. Life Sci. 2000, 66, 1299–1306. [Google Scholar] [CrossRef]
- Prato, F.S.; Robertson, J.A.; Desjardins, D.; Hensel, J.; Thomas, A.W. Daily repeated magnetic field shielding induces analgesia in CD-1 mice. Bioelectromagnetics 2005, 26, 109–117. [Google Scholar] [CrossRef]
- Gurfinkel, I.I.; Vasin, A.L.; Matveeva, T.A.; Sasonko, M.L. Evaluation of the hypomagnetic environment effects on capillary blood circulation, blood pressure and heart rate. Aviakosm. Ekolog. Med. 2014, 48, 24–30. [Google Scholar] [CrossRef]
- Mo, W.C.; Zhang, Z.J.; Liu, Y.; Bartlett, P.F.; He, R.Q. Magnetic shielding accelerates the proliferation of human neuroblastoma cell by promoting G1-phase progression. PLoS ONE 2013, 8, e54775. [Google Scholar] [CrossRef]
- Mo, W.C.; Liu, Y.; He, R.Q. A Biological Perspective of The Hypomagnetic Field: From Definition Towards Mechanism. Prog. Biochem. Biophys. 2012, 39, 835–842. [Google Scholar]
- Belyaev, I.; Alipov, Y.D.; Harms-Ringdahl, M. Effects of zero magnetic field on the conformation of chromatin in human cells. Biochim. Biophys. Acta 1997, 1336, 465–473. [Google Scholar] [CrossRef]
- Martino, C.F.; Castello, P.R. Modulation of hydrogen peroxide production in cellular systems by low level magnetic fields. PLoS ONE 2011, 6, e22753. [Google Scholar] [CrossRef] [PubMed]
- Muller, B.; Ellinwood, N.M.; Lorenz, B.; Stieger, K. Detection of DNA Double Strand Breaks by gammaH2AX Does Not Result in 53bp1 Recruitment in Mouse Retinal Tissues. Front. Neurosci. 2018, 12, 286. [Google Scholar] [CrossRef] [PubMed]
- Jezkova, L.; Zadneprianetc, M.; Kulikova, E.; Smirnova, E.; Bulanova, T.; Depes, D.; Falkova, I.; Boreyko, A.; Krasavin, E.; Davidkova, M.; et al. Particles with similar LET values generate DNA breaks of different complexity and reparability: A high-resolution microscopy analysis of gammaH2AX/53BP1 foci. Nanoscale 2018, 10, 1162–1179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.T.; Zhang, Z.J.; Mo, W.C.; Hu, P.D.; Ding, H.M.; Liu, Y.; Hua, Q.; He, R.Q. Shielding of the geomagnetic field reduces hydrogen peroxide production in human neuroblastoma cell and inhibits the activity of CuZn superoxide dismutase. Protein Cell 2017, 8, 527–537. [Google Scholar] [CrossRef]
- Rogakou, E.P.; Boon, C.; Redon, C.; Bonner, W.M. Megabase chromatin domains involved in DNA double-strand breaks in vivo. J. Cell Biol. 1999, 146, 905–916. [Google Scholar] [CrossRef]
- Aten, J.A.; Stap, J.; Krawczyk, P.M.; van Oven, C.H.; Hoebe, R.A.; Essers, J.; Kanaar, R. Dynamics of DNA double-strand breaks revealed by clustering of damaged chromosome domains. Science 2004, 303, 92–95. [Google Scholar] [CrossRef]
- Chowdhury, D.; Keogh, M.C.; Ishii, H.; Peterson, C.L.; Buratowski, S.; Lieberman, J. gamma-H2AX dephosphorylation by protein phosphatase 2A facilitates DNA double-strand break repair. Mol. Cell 2005, 20, 801–809. [Google Scholar] [CrossRef]
- Krogan, N.J.; Lam, M.H.; Fillingham, J.; Keogh, M.C.; Gebbia, M.; Li, J.; Datta, N.; Cagney, G.; Buratowski, S.; Emili, A.; et al. Proteasome involvement in the repair of DNA double-strand breaks. Mol. Cell 2004, 16, 1027–1034. [Google Scholar] [CrossRef]
- Bouquet, F.; Muller, C.; Salles, B. The loss of gammaH2AX signal is a marker of DNA double strand breaks repair only at low levels of DNA damage. Cell Cycle 2006, 5, 1116–1122. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, D. Repair pathway choice for double-strand breaks. Essays Biochem. 2020, 64, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Shibata, A.; Jeggo, P.A. Roles for the DNA-PK complex and 53BP1 in protecting ends from resection during DNA double-strand break repair. J. Radiat. Res. 2020, 61, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Ward, I.M.; Minn, K.; van Deursen, J.; Chen, J. p53 Binding protein 53BP1 is required for DNA damage responses and tumor suppression in mice. Mol. Cell Biol. 2003, 23, 2556–2563. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, P.; Song, L.; Bai, L.; Huen, M.S.Y.; Liu, Y.; Lu, L.Y. 53BP1 loss rescues embryonic lethality but not genomic instability of BRCA1 total knockout mice. Cell Death Differ. 2020, 27, 2552–2567. [Google Scholar] [CrossRef] [PubMed]
- Frohns, A.; Frohns, F.; Naumann, S.C.; Layer, P.G.; Lobrich, M. Inefficient double-strand break repair in murine rod photoreceptors with inverted heterochromatin organization. Curr. Biol. 2014, 24, 1080–1090. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kohutova, A.; Raska, J.; Kruta, M.; Seneklova, M.; Barta, T.; Fojtik, P.; Jurakova, T.; Walter, C.A.; Hampl, A.; Dvorak, P.; et al. Ligase 3-mediated end-joining maintains genome stability of human embryonic stem cells. FASEB J. 2019, 33, 6778–6788. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.A.; Sun, J.; Kono, K.; Horikoshi, Y.; Ikura, T.; Tong, X.; Haraguchi, T.; Tashiro, S. Regulation of homologous recombinational repair by lamin B1 in radiation-induced DNA damage. FASEB J. 2015, 29, 2514–2525. [Google Scholar] [CrossRef]
- Adler, J.; Parmryd, I. Quantifying colocalization by correlation: The Pearson correlation coefficient is superior to the Mander’s overlap coefficient. Cytom. A 2010, 77, 733–742. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, X.; Ali, Y.F.; Liu, C.; Hong, Z.; Luo, W.; Nie, J.; Li, B.; Jiao, Y.; Liu, N.-A. Geomagnetic Shielding Enhances Radiation Resistance by Promoting DNA Repair Process in Human Bronchial Epithelial Cells. Int. J. Mol. Sci. 2020, 21, 9304. https://doi.org/10.3390/ijms21239304
Xue X, Ali YF, Liu C, Hong Z, Luo W, Nie J, Li B, Jiao Y, Liu N-A. Geomagnetic Shielding Enhances Radiation Resistance by Promoting DNA Repair Process in Human Bronchial Epithelial Cells. International Journal of Molecular Sciences. 2020; 21(23):9304. https://doi.org/10.3390/ijms21239304
Chicago/Turabian StyleXue, Xunwen, Yasser F. Ali, Caorui Liu, Zhiqiang Hong, Wanrong Luo, Jing Nie, Bingyan Li, Yang Jiao, and Ning-Ang Liu. 2020. "Geomagnetic Shielding Enhances Radiation Resistance by Promoting DNA Repair Process in Human Bronchial Epithelial Cells" International Journal of Molecular Sciences 21, no. 23: 9304. https://doi.org/10.3390/ijms21239304
APA StyleXue, X., Ali, Y. F., Liu, C., Hong, Z., Luo, W., Nie, J., Li, B., Jiao, Y., & Liu, N.-A. (2020). Geomagnetic Shielding Enhances Radiation Resistance by Promoting DNA Repair Process in Human Bronchial Epithelial Cells. International Journal of Molecular Sciences, 21(23), 9304. https://doi.org/10.3390/ijms21239304