Advances in Transgenic Mouse Models to Study Infections by Human Pathogenic Viruses


Abstract
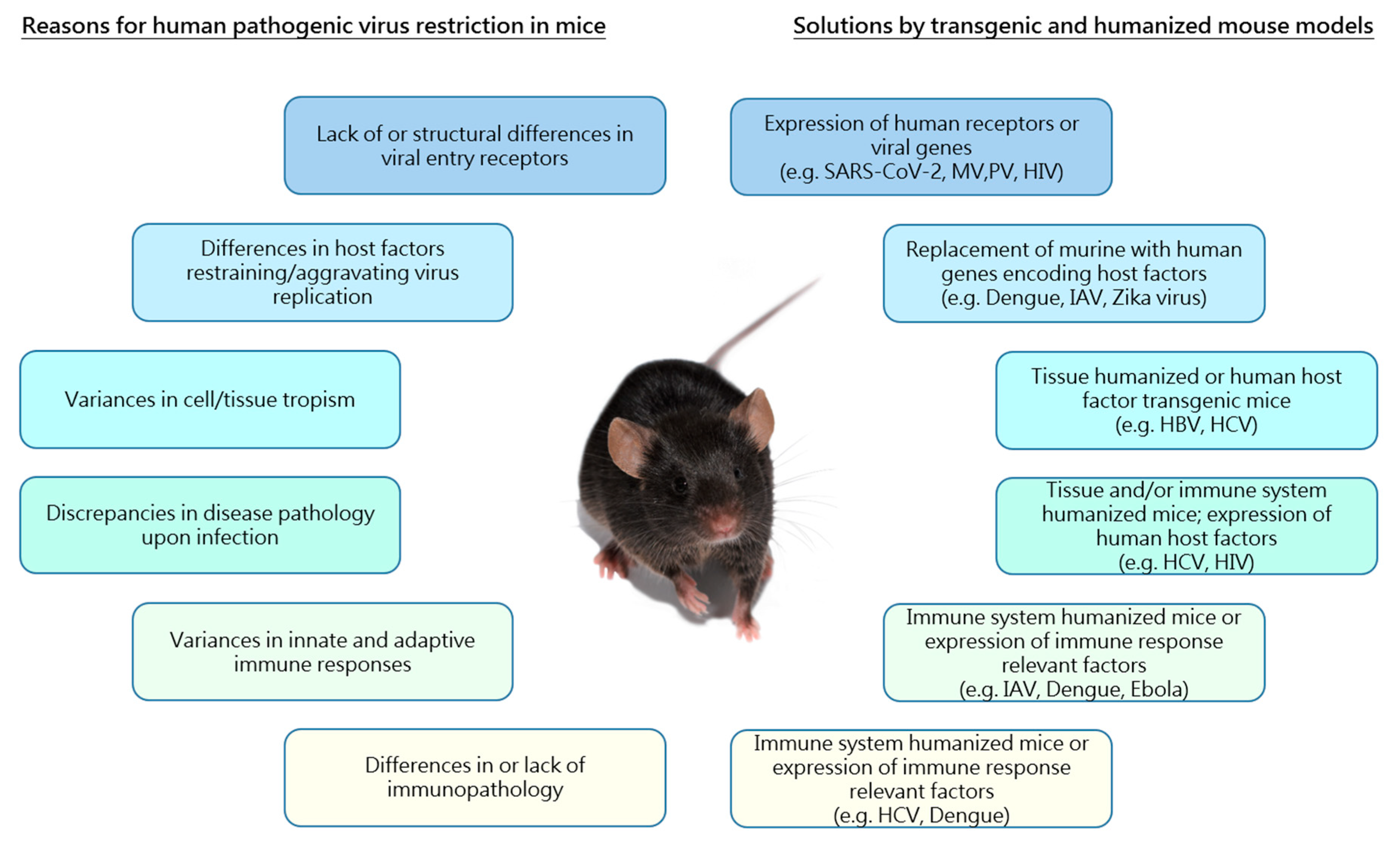
1. Introduction
2. Transgenic Mice Expressing Viral Genes

{kind=link}
| Virus | Entry Receptors | Virus Genome Transgenic Mice | Entry Receptor Transgenic Mice | Transgenic Mice with Changes in Host Factor Expression | Transgenic Tissue/Immune System Humanized Mice |
|---|---|---|---|---|---|
| HIV | CD4, CCR5, CXCR4 | vHIVwttg [32,33,34] vHIVtattg [35,36,37,38,39,40,41] HIVneftg [42,43,44,45,46,47,48,49] HIVdelgag/poltg [50] HIVdelgag/pol/envtg [51] | hCD4/hCCR5 [52,53] | hCD4/hCCR5/ hcyclin T1 [54] CCR5-/-/HIVgp120tg [55] | MISTRG [56] DRAG [57] NOG-EXL [58] |
| Polio | PVR/CD155 | hPVR [59,60,61,62,63,64] | hPVR/IFNAR−/− [65] | ||
| Measles | CD46, CD150, Nectin-4 | hCD46 [66,67] hCD150 [68,69] hCD46/hCD150 [70] | hCD46/IFNAR−/− [71,72] | ||
| HCV | CD81, Occludin, CLDN1; CARB1 | vHCVNS3/4Atg [23] vHCVcore,E1,E2tg [24] vHCVE1,E2tg [25] vHCVFL-Ntg, vHCVS-Ntg [27] vHCVcoretg [29] | hCD81/hOccludin [73,74,75] | AFC8-hu-HSC/Hep [76,77] MUP-uPA [78] | |
| HBV | NTCP | vHBVtg [6,12,79,80] vHBVenvelopetg [4] vHBVdelcoretg [5] vHBVcoretg [8] vHBVHBeAgtg [9] vHBVHBsAg,pre-S,Xtg [7] vHBVHBxtg [10,11] vHBVHBstg [81] vHBVpreS/Stg [82] | vHBVtg/HNF1−/− [83] vHBVHBstg/Abcb4−/− [84] | A2/NSG—hu HSC/Hep [85] MUP-uPA [78,86,87] | |
| SARS-CoV-2 | ACE-2, TMPRSS2 | hACE2 [88,89,90,91,92] | |||
| Influenza | S,3-/2,6-sialic acid residues | hMxA [93] hHLA-A2 [94,95] hHLA-DR3 [96] | DRAGA [97] | ||
| Dengue | Heparan sulfate, DC-SIGN/L-SIGN, mannose receptor, laminin receptor | hTNFα [98] hHLA-A, B [99] IFNAR−/−/HLA-A, B, DRB1 [100,101] | NSG-A2 [102] NSG-SGM3 [103] | ||
| Ebola | Different attaching factors, i.e., C-type lectins, T-cell immunoglobulin and mucin domain 1, tyrosine kinase receptor Axl, Niemann-Pick C1 | HLA-DR3 [104] HLA-A11/DR1 [105] HLA-A2 [106] | NSG-A2 [107,108] NSG-SGM3 [109,110] | ||
| Zika | Glycosaminoglycans NCAM1 | hSTAT2 [111] IFNAR−/−/HLA-B, A, DRB1 [101,112,113] |
3. Human Cell Surface Receptor Supplemented Mice
4. Transgenic Integration of Human Host/Immune Response Factors
5. Transgenic Immune System Humanized Mouse Models
6. Transgenic Humanized Mouse Models for Human-Specific Bloodborne and Hepatotropic Viruses
7. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Feitelson, M.A.; Lee, J. Hepatitis B virus integration, fragile sites, and hepatocarcinogenesis. Cancer Lett. 2007, 252, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhuang, Q.; Wang, Y.; Zhang, T.; Zhao, J.; Zhang, Y.; Zhang, J.; Lin, Y.; Yuan, Q.; Xia, N.; et al. HBV life cycle is restricted in mouse hepatocytes expressing human NTCP. Cell. Mol. Immunol. 2014, 11, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, T.; Guilhot, S.; Klopchin, K.; Moss, B.; Pinkert, C.A.; Palmiter, R.D.; Brinster, R.L.; Kanagawa, O.; Chisari, F.V. Immunobiology and pathogenesis of hepatocellular injury in hepatitis B virus transgenic mice. Science 1990, 248, 361–364. [Google Scholar] [CrossRef]
- Babinet, C.; Farza, H.; Morello, D.; Hadchouel, M.; Pourcel, C. Specific expression of hepatitis B surface antigen (HBsAg) in transgenic mice. Science 1985, 230, 1160–1163. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.D.; DeLoia, J.A.; elAwady, M.K.; Gearhart, J.D. Tissue preferential expression of the hepatitis B virus (HBV) surface antigen gene in two lines of HBV transgenic mice. J. Virol. 1988, 62, 649–654. [Google Scholar] [CrossRef]
- Chisari, F.V.; Pinkert, C.A.; Milich, D.R.; Filippi, P.; Mclachlan, A.; Palmiter, R.D.; Brinster, R.L. A Transgenic Mouse Model of the Chronic Hepatitis B Surface Antigen Carrier State. Science 1985, 230, 1157–1160. [Google Scholar] [CrossRef]
- Guidotti, L.G.; Martinez, V.; Loh, Y.T.; Rogler, C.E.; Chisari, F.V. Hepatitis B virus nucleocapsid particles do not cross the hepatocyte nuclear membrane in transgenic mice. J. Virol. 1994, 68, 5469–5475. [Google Scholar] [CrossRef]
- Milich, D.R.; Jones, J.E.; Hughes, J.L.; Prices, J.; Raney, A.K.; Mclachlan, A. Is a function of the secreted hepatitis B e antigen to induce immunologic tolerance in utero? Proc. Natl. Acad. Sci. USA 1990, 87, 6599–6603. [Google Scholar] [CrossRef]
- Kim, C.M.; Koike, K.; Saito, I.; Miyamura, T.; Jay, G. HBx gene of hepatitis B virus induces liver cancer in transgenic mice. Nature 1991, 351, 317–320. [Google Scholar] [CrossRef]
- Lee, T.H.; Finegold, M.J.; Shen, R.F.; DeMayo, J.L.; Woo, S.L.; Butel, J.S. Hepatitis B virus transactivator X protein is not tumorigenic in transgenic mice. J. Virol. 1990, 64, 5939–5947. [Google Scholar] [CrossRef] [PubMed]
- Guidotti, L.G.; Matzke, B.; Schaller, H.; Chisari, F.V. High-level hepatitis B virus replication in transgenic mice. J. Virol. 1995, 69, 6158–6169. [Google Scholar] [CrossRef] [PubMed]
- Weber, O.; Schlemmer, K.H.; Hartmann, E.; Hagelschuer, I.; Paessens, A.; Graef, E.; Deres, K.; Goldmann, S.; Niewoehner, U.; Stoltefuss, J.; et al. Inhibition of human hepatitis B virus (HBV) by a novel non-nucleosidic compound in a transgenic mouse model. Antivir. Res. 2002, 54, 69–78. [Google Scholar] [CrossRef]
- Wieland, S.F.; Guidotti, L.G.; Chisari, F.V. Intrahepatic Induction of Alpha/Beta Interferon Eliminates Viral RNA-Containing Capsids in Hepatitis B Virus Transgenic Mice. J. Virol. 2000, 74, 4165–4173. [Google Scholar] [CrossRef]
- Julander, J.G.; Colonno, R.J.; Sidwell, R.W.; Morrey, J.D. Characterization of antiviral activity of entecavir in transgenic mice expressing hepatitis B virus. Antivir. Res. 2003, 59, 155–161. [Google Scholar] [CrossRef]
- Hwang, J.-R.; Park, S.-G. Mouse models for hepatitis B virus research. Lab. Anim. Res. 2018, 34, 85. [Google Scholar] [CrossRef]
- Yang, P.L.; Althage, A.; Chung, J.; Chisari, F.V. Hydrodynamic injection of viral DNA: A mouse model of acute hepatitis B virus infection. Proc. Natl. Acad. Sci. USA 2002, 99, 13825–13830. [Google Scholar] [CrossRef]
- Huang, L.-R.; Wu, H.-L.; Chen, P.-J.; Chen, D.-S. An immunocompetent mouse model for the tolerance of human chronic hepatitis B virus infection. Proc. Natl. Acad. Sci. USA 2006, 103, 17862–17867. [Google Scholar] [CrossRef]
- Dion, S.; Bourgine, M.; Godon, O.; Levillayer, F.; Michel, M.-L. Adeno-Associated Virus-Mediated Gene Transfer Leads to Persistent Hepatitis B Virus Replication in Mice Expressing HLA-A2 and HLA-DR1 Molecules. J. Virol. 2013, 87, 5554–5563. [Google Scholar] [CrossRef]
- Yang, D.; Liu, L.; Zhu, D.; Peng, H.; Su, L.; Fu, Y.X.; Zhang, L. A mouse model for HBV immunotolerance and immunotherapy. Cell. Mol. Immunol. 2014, 11, 71–78. [Google Scholar] [CrossRef]
- Lin, S.R.; Yang, H.C.; Kuo, Y.T.; Liu, C.J.; Yang, T.Y.; Sung, K.C.; Lin, Y.Y.; Wang, H.Y.; Wang, C.C.; Shen, Y.C.; et al. The CRISPR/Cas9 system facilitates clearance of the intrahepatic HBV templates in vivo. Mol. Ther. Nucleic Acids 2014, 3, e186. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sheng, C.; Liu, H.; Wang, S.; Zhao, J.; Yang, L.; Jia, L.; Li, P.; Wang, L.; Xie, J.; et al. Inhibition of HBV expression in HBV transgenic mice using AAV-delivered CRISPR-SaCas9. Front. Immunol. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Frelin, L.; Brenndörfer, E.D.; Ahlén, G.; Weiland, M.; Hultgren, C.; Alheim, M.; Glaumann, H.; Rozell, B.; Milich, D.R.; Bode, J.G.; et al. The hepatitis C virus and immune evasion: Non-structural 3/4A transgenic mice are resistant to lethal tumour necrosis factor α mediated liver disease. Gut 2006, 55, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Furusaka, A.; Koziel, M.J.; Chung, R.T.; Wang, T.C.; Schmidt, E.V.; Liang, T.J. Transgenic expression of hepatitis C virus structural proteins in the mouse. Hepatology 1997, 25, 1014–1021. [Google Scholar] [CrossRef]
- Koike, K.; Moriya, K.; Ishibashi, K.; Matsuura, Y.; Suzuki, T.; Saito, I.; Iino, S.; Kurokawa, K.; Miyamura, T. Expression of hepatitis C virus envelope proteins in transgenic mice. J. Gen. Virol. 1995, 76, 3031–3038. [Google Scholar] [CrossRef]
- Alonzi, T.; Agrati, C.; Costabile, B.; Cicchini, C.; Amicone, L.; Cavallari, C.; della Rocca, C.; Folgori, A.; Fipaldini, C.; Poccia, F.; et al. Steatosis and intrahepatic lymphocyte recruitment in hepatitis C virus transgenic mice. J. Gen. Virol. 2004, 85, 1509–1520. [Google Scholar] [CrossRef]
- Lerat, H.; Honda, M.; Beard, M.R.; Loesch, K.; Sun, J.; Yang, Y.; Okuda, M.; Gosert, R.; Xiao, S.Y.; Weinman, S.A.; et al. Steatosis and liver cancer in transgenic mice expressing the structural and nonstructural proteins of hepatitis C virus. Gastroenterology 2002, 122, 352–365. [Google Scholar] [CrossRef]
- Moriya, K.; Fujie, H.; Shintani, Y.; Yotsuyanagi, H.; Tsutsumi, T.; Ishibashi, K.; Matsuura, Y.; Kimura, S.; Miyamura, T.; Koike, K. The core protein of hepatitis C virus induces hepatocellular carcinoma in transgenic mice. Nat. Med. 1998, 4, 1065–1067. [Google Scholar] [CrossRef]
- Jia, F.; Diao, P.; Wang, X.; Hu, X.; Kimura, T.; Nakamuta, M.; Nakamura, I.; Shirotori, S.; Sato, Y.; Moriya, K.; et al. Dietary Restriction Suppresses Steatosis-Associated Hepatic Tumorigenesis in Hepatitis C Virus Core Gene Transgenic Mice. Liver Cancer 2020, 9, 529–548. [Google Scholar] [CrossRef]
- Burm, R.; Collignon, L.; Mesalam, A.A.; Meuleman, P. Animal models to study hepatitis C virus infection. Front. Immunol. 2018, 9, 1032. [Google Scholar] [CrossRef]
- Klopstock, N.; Katzenellenbogen, M.; Pappo, O.; Sklair-Levy, M.; Olam, D.; Mizrahi, L.; Potikha, T.; Galun, E.; Goldenberg, D. HCV tumor promoting effect is dependent on host genetic background. PLoS ONE 2009, 4, e5025. [Google Scholar] [CrossRef] [PubMed]
- Hanna, Z.; Kay, D.G.; Cool, M.; Jothy, S.; Rebai, N.; Jolicoeur, P. Transgenic Mice Expressing Human Immunodeficiency Virus Type 1 in Immune Cells Develop a Severe AIDS-Like Disease. J. Virol. 1998, 72, 121. [Google Scholar] [CrossRef] [PubMed]
- Leonard, J.M.; Abramczuk, J.W.; Pezen, D.S.; Rutledge, R.; Belcher, J.H.; Hakim, F.; Shearer, G.; Lamperth, L.; Travis, W.; Fredrickson, T.; et al. Development of disease and virus recovery in transgenic mice containing HIV proviral DNA. Science 1988, 242, 1665. [Google Scholar] [CrossRef]
- Goudreau, G.; Carpenter, S.; Beaulieu, N.; Jolicoeur, P. Vacuolar myelopathy in transgenic mice expressing human immunodeficiency virus type 1 proteins under the regulation of the myelin basic protein gene promoter. Nat. Med. 1996, 2, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.; Hinrichs, S.H.; Reynolds, R.K.; Luciw, P.A.; Jay, G. The HIV tat gene induces dermal lesions resembling Kaposi’s sarcoma in transgenic mice. Nature 1988, 335, 606–611. [Google Scholar] [CrossRef]
- Brady, H.J.; Abraham, D.J.; Pennington, D.J.; Miles, C.G.; Jenkins, S.; Dzierzak, E.A. Altered cytokine expression in T lymphocytes from human immunodeficiency virus Tat transgenic mice. J. Virol. 1995, 69, 7622–7629. [Google Scholar] [CrossRef]
- Hatziioannou, T.; Evans, D.T. Animal models for HIV/AIDS research. Nat. Rev. Microbiol. 2012, 10, 852–867. [Google Scholar] [CrossRef]
- Bieniasz, P.D.; Cullen, B.R. Multiple Blocks to Human Immunodeficiency Virus Type 1 Replication in Rodent Cells. J.Virol. 2000, 74, 9868–9877. [Google Scholar] [CrossRef]
- Vellutini, C.; Horschowski, N.; Philippon, V.; Gambarelli, D.; Nave, K.-A.; Filippi, P. Development of Lymphoid Hyperplasia in Transgenic Mice Expressing the HIV tat Gene. Aids Res. Hum. Retrovir. 1995, 11, 21–29. [Google Scholar] [CrossRef]
- Ohene-Nyako, M.; Nass, S.R.; Hahn, Y.K.; Knapp, P.E.; Hauser, K.F. Morphine and HIV-1 Tat interact to cause region-specific hyperphosphorylation of tau in transgenic mice. Neurosci. Lett. 2020, 135502. [Google Scholar] [CrossRef]
- Hermes, D.J.; Jacobs, I.R.; Key, M.C.; League, A.F.; Yadav-Samudrala, B.J.; Xu, C.; McLane, V.D.; Nass, S.R.; Jiang, W.; Meeker, R.B.; et al. Escalating morphine dosing in HIV-1 Tat transgenic mice with sustained Tat exposure reveals an allostatic shift in neuroinflammatory regulation accompanied by increased neuroprotective non-endocannabinoid lipid signaling molecules and amino acids. J. Neuroinflammation 2020, 17, 345. [Google Scholar] [CrossRef] [PubMed]
- Brady, H.J.; Pennington, D.J.; Miles, C.G.; Dzierzak, E.A. CD4 cell surface downregulation in HIV-1 Nef transgenic mice is a consequence of intracellular sequestration. Embo J. 1993, 12, 4923–4932. [Google Scholar] [CrossRef] [PubMed]
- Dickie, P.; Ramsdell, F.; Notkins, A.L.; Venkatesan, S. Spontaneous and Inducible Epidermal Hyperplasia in Transgenic Mice Expressing HIV-1 Nef. Virology 1993, 197, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Rahim, M.M.A.; Chrobak, P.; Hu, C.; Hanna, Z.; Jolicoeur, P. Adult AIDS-like disease in a novel inducible human immunodeficiency virus type 1 Nef transgenic mouse model: CD4+ T-cell activation is Nef dependent and can occur in the absence of lymphophenia. J. Virol. 2009, 83, 11830–11846. [Google Scholar] [CrossRef]
- Hanna, Z.; Kay, D.G.; Rebai, N.; Guimond, A.; Jothy, S.; Jolicoeur, P. Nef Harbors a Major Determinant of Pathogenicity for an AIDS-like Disease Induced by HIV-1 in Transgenic Mice. Cell 1998, 95, 163–175. [Google Scholar] [CrossRef]
- Weng, X.; Priceputu, E.; Chrobak, P.; Poudrier, J.; Kay, D.G.; Hanna, Z.; Mak, T.W.; Jolicoeur, P. CD4+ T Cells from CD4C/HIVNef Transgenic Mice Show Enhanced Activation In Vivo with Impaired Proliferation In Vitro but Are Dispensable for the Development of a Severe AIDS-Like Organ Disease. J. Virol. 2004, 78, 5244. [Google Scholar] [CrossRef]
- Lindemann, D.; Wilhelm, R.; Renard, P.; Althage, A.; Zinkernagel, R.; Mous, J. Severe immunodeficiency associated with a human immunodeficiency virus 1 NEF/3’-long terminal repeat transgene. J. Exp. Med. 1994, 179, 797–807. [Google Scholar] [CrossRef]
- Radja, F.; Kay, D.G.; Albrecht, S.; Jolicoeur, P. Oligodendrocyte-Specific Expression of Human Immunodeficiency Virus Type 1 Nef in Transgenic Mice Leads to Vacuolar Myelopathy and Alters Oligodendrocyte Phenotype In Vitro. J. Virol. 2003, 77, 11745. [Google Scholar] [CrossRef]
- Skowronski, J.; Parks, D.; Mariani, R. Altered T cell activation and development in transgenic mice expressing the HIV-1 nef gene. EMBO J. 1993, 12, 703–713. [Google Scholar] [CrossRef]
- Kopp, J.B.; Klotman, M.E.; Adler, S.H.; Bruggeman, L.A.; Dickie, P.; Marinos, N.J.; Eckhaus, M.; Bryant, J.L.; Notkins, A.L.; Klotman, P.E. Progressive glomerulosclerosis and enhanced renal accumulation of basement membrane components in mice transgenic for human immunodeficiency virus type 1 genes. Proc. Natl. Acad. Sci. USA 1992, 89, 1577. [Google Scholar] [CrossRef]
- Tinkle, B.T.; Ngo, L.; Luciw, P.A.; Maciag, T.; Jay, G. Human immunodeficiency virus-associated vasculopathy in transgenic mice. J. Virol. 1997, 71, 4809–4814. [Google Scholar] [CrossRef] [PubMed]
- Kaul, M.; Ma, Q.; Medders, K.E.; Desai, M.K.; Lipton, S.A. HIV-1 coreceptors CCR5 and CXCR4 both mediate neuronal cell death but CCR5 paradoxically can also contribute to protection. Cell Death Differ. 2007, 14, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.; Horner, J.W.; Pettoello-Mantovani, M.; Raker, C.; Yurasov, S.; Depinho, R.A.; Goldstein, H. Mice transgenic for human CD4 and CCR5 are susceptible to HIV infection. Proc. Natl. Acad. Sci. USA 1997, 94, 14637–14641. [Google Scholar] [CrossRef] [PubMed]
- Seay, K.; Qi, X.; Zheng, J.H.; Zhang, C.; Chen, K.; Dutta, M.; Deneroff, K.; Ochsenbauer, C.; Kappes, J.C.; Littman, D.R.; et al. Mice Transgenic for CD4-Specific Human CD4, CCR5 and Cyclin T1 Expression: A New Model for Investigating HIV-1 Transmission and Treatment Efficacy. PLoS ONE 2013, 8, e63537. [Google Scholar] [CrossRef] [PubMed]
- Maung, R.; Hoefer, M.M.; Sanchez, A.B.; Sejbuk, N.E.; Medders, K.E.; Desai, M.K.; Catalan, I.C.; Dowling, C.C.; de Rozieres, C.M.; Garden, G.A.; et al. CCR5 knockout prevents neuronal injury and behavioral impairment induced in a transgenic mouse model by a CXCR4-using HIV-1 glycoprotein 120. J. Immunol. 2014, 193, 1895–1910. [Google Scholar] [CrossRef] [PubMed]
- Ivic, S.; Rochat, M.-A.; Li, D.; Audigé, A.; Schlaepfer, E.; Münz, C.; Manz, M.G.; Speck, R.F. Differential Dynamics of HIV Infection in Humanized MISTRG versus MITRG Mice. ImmunoHorizons 2017, 1, 162–175. [Google Scholar] [CrossRef]
- Kim, J.; Peachman, K.K.; Jobe, O.; Morrison, E.B.; Allam, A.; Jagodzinski, L.; Casares, S.A.; Rao, M. Tracking Human Immunodeficiency Virus-1 Infection in the Humanized DRAG Mouse Model. Front. Immunol. 2017, 8, 1405. [Google Scholar] [CrossRef]
- Perdomo-Celis, F.; Medina-Moreno, S.; Davis, H.; Bryant, J.; Zapata, J.C. HIV Replication in Humanized IL-3/GM-CSF-Transgenic NOG Mice. Pathogens 2019, 8, 33. [Google Scholar] [CrossRef]
- Ren, R.; Costantini, F.; Gorgacz, E.J.; Lee, J.J.; Racaniello, V.R. Transgenic mice expressing a human poliovirus receptor: A new model for poliomyelitis. Cell 1990, 63, 353–362. [Google Scholar] [CrossRef]
- Koike, S.; Taya, C.; Kurata, T.; Abe, S.; Ise, I.; Yonekawa, H.; Nomoto, A. Transgenic mice susceptible to poliovirus. Proc. Natl. Acad. Sci. USA 1991, 88, 951–955. [Google Scholar] [CrossRef]
- Zhang, S.; Racaniello, V.R. Expression of the poliovirus receptor in intestinal epithelial cells is not sufficient to permit poliovirus replication in the mouse gut. J. Virol. 1997, 71, 4915–4920. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Toyoda, H.; Linehan, M.; Iwasaki, A.; Nomoto, A.; Bernhardt, G.; Cello, J.; Wimmer, E. Poliomyelitis in transgenic mice expressing CD155 under the control of the Tage4 promoter after oral and parenteral poliovirus inoculation. J. Gen. Virol. 2014, 95, 1668–1676. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S.; Hix, L.; Sigal, L.J.; Andino, R. Poliovirus pathogenesis in a new poliovirus receptor transgenic mouse model: Age-dependent paralysis and a mucosal route of infection. J. Gen. Virol. 2002, 83, 1707–1720. [Google Scholar] [CrossRef] [PubMed]
- Ida-Hosonuma, M.; Iwasaki, T.; Taya, C.; Sato, Y.; Li, J.; Nagata, N.; Yonekawa, H.; Koike, S. Comparison of neuropathogenicity of poliovirus in two transgenic mouse strains expressing human poliovirus receptor with different distribution patterns. J. Gen. Virol. 2002, 83, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Ida-Hosonuma, M.; Iwasaki, T.; Yoshikawa, T.; Nagata, N.; Sato, Y.; Sata, T.; Yoneyama, M.; Fujita, T.; Taya, C.; Yonekawa, H.; et al. The Alpha/Beta Interferon Response Controls Tissue Tropism and Pathogenicity of Poliovirus. J. Virol. 2005, 79, 4460–4469. [Google Scholar] [CrossRef] [PubMed]
- Rall, G.F.; Manchester, M.; Daniels, L.R.; Callahan, E.M.; Belman, A.R.; Oldstone, M.B.A. A transgenic mouse model for measles virus infection of the brain. Proc. Natl. Acad. Sci. USA 1997, 94, 4659–4663. [Google Scholar] [CrossRef] [PubMed]
- Kemper, C.; Leung, M.; Stephensen, C.B.; Pinkert, C.A.; Liszewski, M.K.; Cattaneo, R.; Atkinson, J.P. Membrane cofactor protein (MCP; CD46) expression in transgenic mice. Clin. Exp. Immunol. 2001, 124, 180–189. [Google Scholar] [CrossRef]
- Ohno, S.; Ono, N.; Seki, F.; Takeda, M.; Kura, S.; Tsuzuki, T.; Yanagi, Y. Measles Virus Infection of SLAM (CD150) Knockin Mice Reproduces Tropism and Immunosuppression in Human Infection. J. Virol. 2007, 81, 1650–1659. [Google Scholar] [CrossRef]
- Welstead, G.G.; Iorio, C.; Draker, R.; Bayani, J.; Squire, J.; Vongpunsawad, S.; Cattaneo, R.; Richardson, C.D. Measles virus replication in lymphatic cells and organs of CD150 (SLAM) transgenic mice. Proc. Natl. Acad. Sci. USA 2005, 102, 16415–16420. [Google Scholar] [CrossRef]
- Shingai, M.; Inoue, N.; Okuno, T.; Okabe, M.; Akazawa, T.; Miyamoto, Y.; Ayata, M.; Honda, K.; Kurita-Taniguchi, M.; Matsumoto, M.; et al. Wild-Type Measles Virus Infection in Human CD46/CD150-Transgenic Mice: CD11c-Positive Dendritic Cells Establish Systemic Viral Infection. J. Immunol. 2005, 175, 3252–3261. [Google Scholar] [CrossRef]
- Mrkic, B.; Pavlovic, J.; Rülicke, T.; Volpe, P.; Buchholz, C.J.; Hourcade, D.; Atkinson, J.P.; Aguzzi, A.; Cattaneo, R. Measles Virus Spread and Pathogenesis in Genetically Modified Mice. J. Virol. 1998, 72, 7420–7427. [Google Scholar] [CrossRef] [PubMed]
- Marty, R.R.; Knuchel, M.C.; Morin, T.N.A.; Naim, H.Y. An immune competent mouse model for the characterization of recombinant measles vaccines. Hum. Vaccines Immunother. 2015, 11, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Dorner, M.; Horwitz, J.A.; Robbins, J.B.; Barry, W.T.; Feng, Q.; Mu, K.; Jones, C.T.; Schoggins, J.W.; Catanese, M.T.; Burton, R.; et al. A genetically humanized mouse model for hepatitis C virus infection. Nature 2011, 474, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Dorner, M.; Horwitz, J.A.; Donovan, B.M.; Labitt, R.N.; Budell, W.C.; Friling, T.; Vogt, A.; Catanese, M.T.; Satoh, T.; Kawai, T.; et al. Completion of the entire hepatitis C virus life cycle in genetically humanized mice. Nature 2013, 501, 237–241. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, Y.; Zhang, C.; Chen, H.; Feng, J.; Chi, X.; Pan, Y.; Du, J.; Guo, M.; Cao, H.; et al. Persistent hepatitis C virus infections and hepatopathological manifestations in immune-competent humanized mice. Cell Res. 2014, 24, 1050–1066. [Google Scholar] [CrossRef]
- Bility, M.T.; Zhang, L.; Washburn, M.L.; Curtis, T.A.; Kovalev, G.I.; Su, L. Generation of a humanized mouse model with both human immune system and liver cells to model hepatitis C virus infection and liver immunopathogenesis. Nat. Protoc. 2012, 7, 1608–1617. [Google Scholar] [CrossRef]
- Washburn, M.L.; Bility, M.T.; Zhang, L.; Kovalev, G.I.; Buntzman, A.; Frelinger, J.A.; Barry, W.; Ploss, A.; Rice, C.M.; Su, L. A humanized mouse model to study hepatitis C virus infection, immune response, and liver disease. Gastroenterology 2011, 140, 1334–1344. [Google Scholar] [CrossRef]
- Tesfaye, A.; Stift, J.; Maric, D.; Cui, Q.; Dienes, H.-P.; Feinstone, S.M. Chimeric Mouse Model for the Infection of Hepatitis B and C Viruses. PLoS ONE 2013, 8, e77298. [Google Scholar] [CrossRef]
- Fumagalli, V.; di Lucia, P.; Venzin, V.; Bono, E.B.; Jordan, R.; Frey, C.R.; Delaney, W.; Chisari, F.V.; Guidotti, L.G.; Iannacone, M. Serum HBsAg clearance has minimal impact on CD8+ T cell responses in mouse models of HBV infection. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Liu, P.J.; Harris, J.M.; Marchi, E.; D’Arienzo, V.; Michler, T.; Wing, P.A.C.; Magri, A.; Ortega-Prieto, A.M.; van de Klundert, M.; Wettengel, J.; et al. Hypoxic gene expression in chronic hepatitis B virus infected patients is not observed in state-of-the-art in vitro and mouse infection models. Sci. Rep. 2020, 10, 14101. [Google Scholar] [CrossRef]
- Churin, Y.; Roderfeld, M.; Stiefel, J.; Würger, T.; Schröder, D.; Matono, T.; Mollenkopf, H.-J.; Montalbano, R.; Pompaiah, M.; Reifenberg, K.; et al. Pathological Impact of Hepatitis B Virus Surface Proteins on the Liver Is Associated with the Host Genetic Background. PLoS ONE 2014, 9, e90608. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.-W.; Liang, K.-H.; Lin, W.-R.; Huang, Y.-H.; Huang, S.-F.; Chen, T.-C.; Yeh, C.-T. Hepatocarcinogenesis in transgenic mice carrying hepatitis B virus pre-S/S gene with the sW172* mutation. Oncogenesis 2016, 5, e273. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Raney, A.K.; Eggers, C.M.; Kline, E.F.; Guidotti, L.G.; Pontoglio, M.; Yaniv, M.; McLachlan, A. Nuclear Covalently Closed Circular Viral Genomic DNA in the Liver of Hepatocyte Nuclear Factor 1α-Null Hepatitis B Virus Transgenic Mice. J. Virol. 2001, 75, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Irungbam, K.; Roderfeld, M.; Glimm, H.; Hempel, F.; Schneider, F.; Hehr, L.; Glebe, D.; Churin, Y.; Morlock, G.; Yüce, I.; et al. Cholestasis impairs hepatic lipid storage via AMPK and CREB signaling in hepatitis B virus surface protein transgenic mice. Lab. Investig. 2020, 100, 1411–1424. [Google Scholar] [CrossRef] [PubMed]
- Bility, M.T.; Cheng, L.; Zhang, Z.; Luan, Y.; Li, F.; Chi, L.; Zhang, L.; Tu, Z.; Gao, Y.; Fu, Y.; et al. Hepatitis B Virus Infection and Immunopathogenesis in a Humanized Mouse Model: Induction of Human-Specific Liver Fibrosis and M2-Like Macrophages. PLoS Pathog. 2014, 10. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, N.; Tesfaye, A.; Feinstone, S.; Kumar, A. HCV infection-associated hepatocellular carcinoma in humanized mice. Infect. Agents Cancer 2015, 10, 24. [Google Scholar] [CrossRef]
- Bobardt, M.; Hansson, M.J.; Mayo, P.; Ure, D.; Foster, R.; Gallay, P. Structurally distinct cyclosporin and sanglifehrin analogs CRV431 and NV556 suppress established HCV infection in humanized-liver mice. PLoS ONE 2020, 15, e0237236. [Google Scholar] [CrossRef]
- McCray, P.B.; Pewe, L.; Wohlford-Lenane, C.; Hickey, M.; Manzel, L.; Shi, L.; Netland, J.; Jia, H.P.; Halabi, C.; Sigmund, C.D.; et al. Lethal Infection of K18-hACE2 Mice Infected with Severe Acute Respiratory Syndrome Coronavirus. J. Virol. 2007, 81, 813–821. [Google Scholar] [CrossRef]
- Winkler, E.S.; Bailey, A.L.; Kafai, N.M.; Nair, S.; McCune, B.T.; Yu, J.; Fox, J.M.; Chen, R.E.; Earnest, J.T.; Keeler, S.P.; et al. SARS-CoV-2 infection of human ACE2-transgenic mice causes severe lung inflammation and impaired function. Nat. Immunol. 2020. [Google Scholar] [CrossRef]
- Oladunni, F.S.; Park, J.-G.; Pino Tamayo, P.; Gonzalez, O.; Akhter, A.; Allué-Guardia, A.; Olmo-Fontánez, A.; Gautam, S.; Ye, C.; Chiem, K.; et al. Lethality of SARS-CoV-2 infection in K18 human angiotensin converting enzyme 2 transgenic mice. bioRxiv 2020. [Google Scholar] [CrossRef]
- Bao, L.; Deng, W.; Huang, B.; Gao, H.; Liu, J.; Ren, L.; Wei, Q.; Yu, P.; Xu, Y.; Qi, F.; et al. The pathogenicity of SARS-CoV-2 in hACE2 transgenic mice. Nature 2020, 583, 830–833. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.H.; Chen, Q.; Gu, H.J.; Yang, G.; Wang, Y.X.; Huang, X.Y.; Liu, S.S.; Zhang, N.N.; Li, X.F.; Xiong, R.; et al. A Mouse Model of SARS-CoV-2 Infection and Pathogenesis. Cell Host Microbe 2020, 28, 124–133.e4. [Google Scholar] [CrossRef]
- Deeg, C.M.; Hassan, E.; Mutz, P.; Rheinemann, L.; Götz, V.; Magar, L.; Schilling, M.; Kallfass, C.; Nürnberger, C.; Soubies, S.; et al. In vivo evasion of MxA by avian influenza viruses requires human signature in the viral nucleoprotein. J. Exp. Med. 2017, 214, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- di Mario, G.; Sciaraffia, E.; Facchini, M.; Gubinelli, F.; Soprana, E.; Panigada, M.; Bernasconi, V.; Garulli, B.; Siccardi, A.; Donatelli, I.; et al. Protective immunity against influenza in HLA-A2 transgenic mice by modified vaccinia virus Ankara vectored vaccines containing internal influenza proteins. Pathog. Glob. Health 2017, 111, 76–82. [Google Scholar] [CrossRef] [PubMed]
- di Mario, G.; Garulli, B.; Sciaraffia, E.; Facchini, M.; Donatelli, I.; Castrucci, M.R. A heat-inactivated H7N3 vaccine induces cross-reactive cellular immunity in HLA-A2.1 transgenic mice. Virol. J. 2016, 13, 4–13. [Google Scholar] [CrossRef]
- Moise, L.; Tassone, R.; Latimer, H.; Terry, F.; Levitz, L.; Haran, J.P.; Ross, T.M.; Boyle, C.; Martin, W.D.; de Groot, A.S. Immunization with cross-conserved H1N1 influenza CD4+T-cell epitopes lowers viral burden in HLA DR3 transgenic mice. Hum. Vaccines Immunother. 2013, 9, 2060–2068. [Google Scholar] [CrossRef]
- Mendoza, M.; Ballesteros, A.; Qiu, Q.; Pow Sang, L.; Shashikumar, S.; Casares, S.; Brumeanu, T.D. Generation and testing anti-influenza human monoclonal antibodies in a new humanized mouse model (DRAGA: HLA-A2. HLA-DR4. Rag1 KO. IL-2Rγc KO. NOD). Hum. Vaccines Immunother. 2018, 14, 345–360. [Google Scholar] [CrossRef]
- Jhan, M.K.; HuangFu, W.C.; Chen, Y.F.; Kao, J.C.; Tsai, T.T.; Ho, M.R.; Shen, T.J.; Tseng, P.C.; Wang, Y.T.; Lin, C.F. Anti-TNF-α restricts dengue virus-induced neuropathy. J. Leukoc. Biol. 2018, 104, 961–968. [Google Scholar] [CrossRef]
- Roth, C.; Cantaert, T.; Colas, C.; Prot, M.; Casadémont, I.; Levillayer, L.; Thalmensi, J.; Langlade-Demoyen, P.; Gerke, C.; Bahl, K.; et al. A Modified mRNA Vaccine Targeting Immunodominant NS Epitopes Protects Against Dengue Virus Infection in HLA Class I Transgenic Mice. Front. Immunol. 2019, 10, 1424. [Google Scholar] [CrossRef]
- Weiskopf, D.; Yauch, L.E.; Angelo, M.A.; John, D.V.; Greenbaum, J.A.; Sidney, J.; Kolla, R.V.; de Silva, A.D.; de Silva, A.M.; Grey, H.; et al. Insights into HLA-Restricted T Cell Responses in a Novel Mouse Model of Dengue Virus Infection Point toward New Implications for Vaccine Design. J. Immunol. 2011, 187, 4268. [Google Scholar] [CrossRef]
- Wen, J.; Wang, Y.-T.; Valentine, K.M.; dos Santos Alves, R.P.; Xu, Z.; Regla-Nava, J.A.; Ngono, A.E.; Young, M.P.; Ferreira, L.C.S.; Shresta, S. CD4(+) T Cells Cross-Reactive with Dengue and Zika Viruses Protect against Zika Virus Infection. Cell Rep. 2020, 31, 107566. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.; Pearson, T.; Friberg, H.; Shultz, L.D.; Greiner, D.L.; Rothman, A.L.; Mathew, A. Dengue virus infection and virus-specific HLA-A2 restricted immune responses in humanized NOD-scid IL2rγnull mice. PLoS ONE 2009, 4, e7251. [Google Scholar] [CrossRef] [PubMed]
- Jangalwe, S.; Shultz, L.D.; Mathew, A.; Brehm, M.A. Improved B cell development in humanized NOD-scid IL2Rγ null mice transgenically expressing human stem cell factor, granulocyte-macrophage colony-stimulating factor and interleukin-3. Immun. Inflamm. Dis. 2016, 4, 427–440. [Google Scholar] [CrossRef] [PubMed]
- Bounds, C.E.; Terry, F.E.; Moise, L.; Hannaman, D.; Martin, W.D.; de Groot, A.S.; Suschak, J.J.; Dupuy, L.C.; Schmaljohn, C.S. An immunoinformatics-derived DNA vaccine encoding human class II T cell epitopes of Ebola virus, Sudan virus, and Venezuelan equine encephalitis virus is immunogenic in HLA transgenic mice. Hum. Vaccines Immunother. 2017, 13, 2824–2836. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, P.; Song, N.; Jiang, Y.; Zeng, Y.; Zhao, G.; Fa, Y.; Ye, H.; Lone, Y.; Zhou, Y.; et al. Identification of novel HLA-A11-restricted T-cell epitopes in the Ebola virus nucleoprotein. Microbes Infect. 2019, 21, 56–62. [Google Scholar] [CrossRef]
- Sundar, K.; Boesen, A.; Coico, R. Computational prediction and identification of HLA-A2.1-specific Ebola virus CTL epitopes. Virology 2007, 360, 257–263. [Google Scholar] [CrossRef]
- Escudero-Pérez, B.; Ruibal, P.; Rottstegge, M.; Lüdtke, A.; Port, J.R.; Hartmann, K.; Gómez-Medina, S.; Müller-Guhl, J.; Nelson, E.V.; Krasemann, S.; et al. Comparative pathogenesis of Ebola virus and Reston virus infection in humanized mice. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Lüdtke, A.; Oestereich, L.; Ruibal, P.; Wurr, S.; Pallasch, E.; Bockholt, S.; Ip, W.H.; Rieger, T.; Gómez-Medina, S.; Stocking, C.; et al. Ebola virus disease in mice with transplanted human hematopoietic stem cells. J. Virol. 2015, 89, 4700–4704. [Google Scholar] [CrossRef]
- Spengler, J.R.; Saturday, G.; Lavender, K.J.; Martellaro, C.; Keck, J.G.; Nichol, S.T.; Spiropoulou, C.F.; Feldmann, H.; Prescott, J. Severity of disease in humanized mice infected with ebola virus or reston virus is associated with magnitude of early viral replication in liver. J. Infect. Dis. 2018, 217, 58–63. [Google Scholar] [CrossRef]
- Spengler, J.R.; Lavender, K.J.; Martellaro, C.; Carmody, A.; Kurth, A.; Keck, J.G.; Saturday, G.; Scott, D.P.; Nichol, S.T.; Hasenkrug, K.J.; et al. Ebola Virus Replication and Disease Without Immunopathology in Mice Expressing Transgenes to Support Human Myeloid and Lymphoid Cell Engraftment. J. Infect. Dis. 2016, 214, S308–S318. [Google Scholar] [CrossRef]
- Gorman, M.J.; Caine, E.A.; Zaitsev, K.; Begley, M.C.; Weger-lucarelli, J.; Uccellini, M.B.; Tripathi, S.; Morrison, J.; Yount, B.L.; Dinnon, K.H., III; et al. An immunocompetent mouse model of Zika virus infection. Stem Cells Dev. 2018, 23, 672–685. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Tang, W.W.; Sheets, N.; Ellison, J.; Sette, A.; Kim, K.; Shresta, S. Identification of Zika virus epitopes reveals immunodominant and protective roles for dengue virus cross-reactive CD8+ T cells. Nat. Microbiol. 2017, 2, 17036. [Google Scholar] [CrossRef] [PubMed]
- Elong Ngono, A.; Syed, T.; Nguyen, A.-V.; Regla-Nava, J.A.; Susantono, M.; Spasova, D.; Aguilar, A.; West, M.; Sparks, J.; Gonzalez, A.; et al. CD8(+) T cells mediate protection against Zika virus induced by an NS3-based vaccine. Sci. Adv. 2020, 6, eabb2154. [Google Scholar] [CrossRef] [PubMed]
- Kuba, K.; Imai, Y.; Rao, S.; Gao, H.; Guo, F.; Guan, B.; Huan, Y.; Yang, P.; Zhang, Y.; Deng, W.; et al. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nat. Med. 2005, 11, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Soldatov, V.O.; Kubekina, M.V.; Silaeva, Y.Y.; Bruter, A.V.; Deykin, A.V. On the way from SARS-CoV-sensitive mice to murine COVID-19 model. Res. Results Pharmacol. 2020, 6, 1–7. [Google Scholar] [CrossRef]
- Bodian, D. Emerging Concept of Poliomyelitis Infection. Science 1955, 122, 105–108. [Google Scholar] [CrossRef]
- Sabin, A.B. Pathogenesis of Poliomyelitis Reappraisal in the Light of New Data. Science 1956, 123, 1151–1157. [Google Scholar] [CrossRef]
- Gromeier, M.; Alexander, L.; Wimmer, E. Internal ribosomal entry site substitution eliminates neurovirulence in intergeneric poliovirus recombinants. Proc. Natl. Acad. Sci. USA 1996, 93, 2370–2375. [Google Scholar] [CrossRef]
- Ohka, S.; Igarashi, H.; Nagata, N.; Sakai, M.; Koike, S.; Nochi, T.; Kiyono, H.; Nomoto, A. Establishment of a Poliovirus Oral Infection System in Human Poliovirus Receptor-Expressing Transgenic Mice That Are Deficient in Alpha/Beta Interferon Receptor. J. Virol. 2007, 81, 7902–7912. [Google Scholar] [CrossRef]
- Lindenmann, J.; Klein, P.A. Further Studies on the Resistance of Mice to Myxoviruses. Archiv für die gesamte Virusforschung 1966, 19, 1–12. [Google Scholar] [CrossRef]
- Ibricevic, A.; Pekosz, A.; Walter, M.J.; Newby, C.; Battaile, J.T.; Brown, E.G.; Holtzman, M.J.; Brody, S.L. Influenza Virus Receptor Specificity and Cell Tropism in Mouse and Human Airway Epithelial Cells. J. Virol. 2006, 80, 7469–7480. [Google Scholar] [CrossRef] [PubMed]
- van Riel, D.; Munster, V.J.; de Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Kuiken, T. Human and avian influenza viruses target different cells in the lower respiratory tract of humans and other mammals. Am. J. Pathol. 2007, 171, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Haller, O.; Acklin, M.; Staeheli, P. Influenza Virus Resistance of Wild Mice: Wild-Type and Mutant Mx Alleles Occur at Comparable Frequencies. J. Interferon Res. 1987, 7, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Staeheli, P.; Grob, R.; Meier, E.; Sutcliffe, J.G.; Haller, O. Influenza virus-susceptible mice carry Mx genes with a large deletion or a nonsense mutation. Mol. Cell. Biol. 1988, 8, 4518–4523. [Google Scholar] [CrossRef]
- Tumpey, T.M.; Szretter, K.J.; van Hoeven, N.; Katz, J.M.; Kochs, G.; Haller, O.; García-Sastre, A.; Staeheli, P. The Mx1 Gene Protects Mice against the Pandemic 1918 and Highly Lethal Human H5N1 Influenza Viruses. J. Virol. 2007, 81, 10818–10821. [Google Scholar] [CrossRef]
- Haller, O.; Staeheli, P.; Schwemmle, M.; Kochs, G. Mx GTPases: Dynamin-like antiviral machines of innate immunity. Trends Microbiol. 2015, 23, 154–163. [Google Scholar] [CrossRef]
- Haller, O.; Kochs, G. Mx genes: Host determinants controlling influenza virus infection and trans-species transmission. Hum. Genet. 2020, 139, 695–705. [Google Scholar] [CrossRef]
- Reiser, M.; Wieland, A.; Plachter, B.; Mertens, T.; Greiner, J.; Schirmbeck, R. The Immunodominant CD8 T Cell Response to the Human Cytomegalovirus Tegument Phosphoprotein pp65 495–503 Epitope Critically Depends on CD4 T Cell Help in Vaccinated HLA-A*0201 Transgenic Mice. J. Immunol. 2011, 187, 2172–2180. [Google Scholar] [CrossRef]
- Himoudi, N.; Abraham, J.-D.; Fournillier, A.; Lone, Y.C.; Joubert, A.; de Beeck, A.O.; Freida, D.; Lemonnier, F.; Kieny, M.P.; Inchauspé, G. Comparative Vaccine Studies in HLA-A2.1-Transgenic Mice Reveal a Clustered Organization of Epitopes Presented in Hepatitis C Virus Natural Infection. J. Virol. 2002, 76, 12735–12746. [Google Scholar] [CrossRef]
- Moise, L.; Buller, R.M.; Schriewer, J.; Lee, J.; Frey, S.E.; Weiner, D.B.; Martin, W.; de Groot, A.S. VennVax, a DNA-prime, peptide-boost multi-T-cell epitope poxvirus vaccine, induces protective immunity against vaccinia infection by T cell response alone. Vaccine 2011, 29, 501–511. [Google Scholar] [CrossRef]
- Yong, K.S.M.; Her, Z.; Chen, Q. Humanized Mice as Unique Tools for Human-Specific Studies. Arch. Immunol. Ther. Exp. 2018, 66, 245–266. [Google Scholar] [CrossRef] [PubMed]
- Morillon, Y.M.; Sabzevari, A.; Schlom, J.; Greiner, J.W. The Development of Next-generation PBMC Humanized Mice for Preclinical Investigation of Cancer Immunotherapeutic Agents. Anticancer Res. 2020, 40, 5329–5341. [Google Scholar] [CrossRef] [PubMed]
- Bosma, G.C.; Custer, R.P.; Bosma, M.J. A severe combined immunodeficiency mutation in the mouse. Nature 1983, 301, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Bosma, M.J.; Carroll, A.M. The SCID Mouse Mutant: Definition, Characterization, and Potential Uses. Annu. Rev. Immunol. 1991, 9, 323–350. [Google Scholar] [CrossRef] [PubMed]
- Shinkai, Y.; Rathbun, G.; Lam, K.-P.; Oltz, E.M.; Stewart, V.; Mendelsohn, M.; Charron, J.; Datta, M.; Young, F.; Stall, A.M.; et al. RAG-2-deficient mice lack mature lymphocytes owing to inability to initiate V(D)J rearrangement. Cell 1992, 68, 855–867. [Google Scholar] [CrossRef]
- Rochman, Y.; Spolski, R.; Leonard, W.J. New insights into the regulation of T cells by gamma(c) family cytokines. Nat. Rev. Immunol. 2009, 9, 480–490. [Google Scholar] [CrossRef]
- Pearson, T.; Markees, T.G.; Serreze, D.V.; Pierce, M.A.; Marron, M.P.; Wicker, L.S.; Peterson, L.B.; Shultz, L.D.; Mordes, J.P.; Rossini, A.A.; et al. Genetic Disassociation of Autoimmunity and Resistance to Costimulation Blockade-Induced Transplantation Tolerance in Nonobese Diabetic Mice. J. Immunol. 2003, 171, 185. [Google Scholar] [CrossRef]
- Ishikawa, F.; Yasukawa, M.; Lyons, B.; Yoshida, S.; Miyamoto, T.; Yoshimoto, G.; Watanabe, T.; Akashi, K.; Shultz, L.D.; Harada, M. Development of functional human blood and immune systems in NOD/SCID/IL2 receptor {gamma} chain(null) mice. Blood 2005, 106, 1565–1573. [Google Scholar] [CrossRef]
- Ito, M.; Hiramatsu, H.; Kobayashi, K.; Suzue, K.; Kawahata, M.; Hioki, K.; Ueyama, Y.; Koyanagi, Y.; Sugamura, K.; Tsuji, K.; et al. NOD/SCID/γcnull mouse: An excellent recipient mouse model for engraftment of human cells. Blood 2002, 100, 3175–3182. [Google Scholar] [CrossRef]
- Shultz, L.D.; Lyons, B.L.; Burzenski, L.M.; Gott, B.; Chen, X.; Chaleff, S.; Kotb, M.; Gillies, S.D.; King, M.; Mangada, J.; et al. Human Lymphoid and Myeloid Cell Development in NOD/LtSz-scid IL2R gamma null Mice Engrafted with Mobilized Human Hemopoietic Stem Cells. J. Immunol. 2005, 174, 6477. [Google Scholar] [CrossRef]
- Pearson, T.; Greiner, D.L.; Shultz, L.D. Creation of “humanized” mice to study human immunity. Curr. Protoc. Immunol. 2008. [Google Scholar] [CrossRef] [PubMed]
- Brehm, M.A.; Cuthbert, A.; Yang, C.; Miller, D.M.; DiIorio, P.; Laning, J.; Burzenski, L.; Gott, B.; Foreman, O.; Kavirayani, A.; et al. Parameters for establishing humanized mouse models to study human immunity: Analysis of human hematopoietic stem cell engraftment in three immunodeficient strains of mice bearing the IL2rgamma(null) mutation. Clin. Immunol. 2010, 135, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Duchosal, M.A.; Eming, S.A.; McConahey, P.J.; Dixon, F.J. Characterization of hu-PBL-SCID mice with high human immunoglobulin serum levels and graft-versus-host disease. Am. J. Pathol. 1992, 141, 1097–1113. [Google Scholar] [PubMed]
- Tary-Lehmann, M.; Saxon, A.; Lehmann, P.V. The human immune system in hu-PBL-SCID mice. Immunol. Today 1995, 16, 529–533. [Google Scholar] [CrossRef]
- Lan, P.; Tonomura, N.; Shimizu, A.; Wang, S.; Yang, Y.-G. Reconstitution of a functional human immune system in immunodeficient mice through combined human fetal thymus/liver and CD34+ cell transplantation. Blood 2006, 108, 487–492. [Google Scholar] [CrossRef]
- Covassin, L.; Jangalwe, S.; Jouvet, N.; Laning, J.; Burzenski, L.; Shultz, L.D.; Brehm, M.A. Human immune system development and survival of non-obese diabetic (NOD)-scid IL2rγ(null) (NSG) mice engrafted with human thymus and autologous haematopoietic stem cells. Clin. Exp. Immunol. 2013, 174, 372–388. [Google Scholar] [CrossRef]
- Hu, Z.; van Rooijen, N.; Yang, Y.-G. Macrophages prevent human red blood cell reconstitution in immunodeficient mice. Blood 2011, 118, 5938–5946. [Google Scholar] [CrossRef]
- Chen, Q.; He, F.; Kwang, J.; Chan, J.K.Y.; Chen, J. GM-CSF and IL-4 stimulate antibody responses in humanized mice by promoting T, B, and dendritic cell maturation. J. Immunol. 2012, 189, 5223–5229. [Google Scholar] [CrossRef]
- Lang, J.; Kelly, M.; Freed, B.M.; McCarter, M.D.; Kedl, R.M.; Torres, R.M.; Pelanda, R. Studies of lymphocyte reconstitution in a humanized mouse model reveal a requirement of T cells for human B cell maturation. J. Immunol. 2013, 190, 2090–2101. [Google Scholar] [CrossRef]
- Tsuji, M.; Akkina, R. Editorial: Development of Humanized Mouse Models for Infectious Diseases and Cancer. Front. Immunol. 2020, 10, 3051. [Google Scholar] [CrossRef]
- Ito, R.; Takahashi, T.; Katano, I.; Kawai, K.; Kamisako, T.; Ogura, T.; Ida-Tanaka, M.; Suemizu, H.; Nunomura, S.; Ra, C.; et al. Establishment of a Human Allergy Model Using Human IL-3/GM-CSF–Transgenic NOG Mice. J. Immunol. 2013, 191, 2890. [Google Scholar] [CrossRef] [PubMed]
- Nicolini, F.E.; Cashman, J.D.; Hogge, D.E.; Humphries, R.K.; Eaves, C.J. NOD/SCID mice engineered to express human IL-3, GM-CSF and Steel factor constitutively mobilize engrafted human progenitors and compromise human stem cell regeneration. Leukemia 2004, 18, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Rongvaux, A.; Willinger, T.; Martinek, J.; Strowig, T.; Gearty, S.V.; Teichmann, L.L.; Saito, Y.; Marches, F.; Halene, S.; Palucka, A.K.; et al. Development and function of human innate immune cells in a humanized mouse model. Nat. Biotechnol. 2014, 32, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Shultz, L.D.; Saito, Y.; Najima, Y.; Tanaka, S.; Ochi, T.; Tomizawa, M.; Doi, T.; Sone, A.; Suzuki, N.; Fujiwara, H.; et al. Generation of functional human T-cell subsets with HLA-restricted immune responses in HLA class I expressing NOD/SCID/IL2r gamma(null) humanized mice. Proc. Natl. Acad. Sci. USA 2010, 107, 13022–13027. [Google Scholar] [CrossRef]
- Danner, R.; Chaudhari, S.N.; Rosenberger, J.; Surls, J.; Richie, T.L.; Brumeanu, T.-D.; Casares, S. Expression of HLA class II molecules in humanized NOD.Rag1KO.IL2RgcKO mice is critical for development and function of human T and B cells. PLoS ONE 2011, 6, e19826. [Google Scholar] [CrossRef]
- Majji, S.; Wijayalath, W.; Shashikumar, S.; Brumeanu, T.D.; Casares, S. Humanized DRAGA mice immunized with Plasmodium falciparum sporozoites and chloroquine elicit protective pre-erythrocytic immunity. Malar. J. 2018, 17, 114. [Google Scholar] [CrossRef]
- Zeng, Y.; Liu, B.; Rubio, M.T.; Wang, X.; Ojcius, D.M.; Tang, R.; Durrbach, A.; Ru, Z.; Zhou, Y.; Lone, Y.C. Creation of an immunodeficient HLA-transgenic mouse (HUMAMICE) and functional validation of human immunity after transfer of HLA-matched human cells. PLoS ONE 2017, 12, e173754. [Google Scholar] [CrossRef]
- Marsden, M.D. Benefits and limitations of humanized mice in HIV persistence studies. Retrovirology 2020, 17, 7. [Google Scholar] [CrossRef]
- Weichseldorfer, M.; Heredia, A.; Reitz, M.; Bryant, J.L.; Latinovic, O.S. Use of Humanized Mouse Models for Studying HIV-1 Infection, Pathogenesis and Persistence. J. AIDS HIV Treat. 2020, 2, 23–29. [Google Scholar]
- Begum, F.; Das, S.; Mukherjee, D.; Mal, S.; Ray, U. Insight into the Tropism of Dengue Virus in Humans. Viruses 2019, 11, 1136. [Google Scholar] [CrossRef]
- Martina, B.E.E.; Koraka, P.; Osterhaus, A.D.M.E. Dengue virus pathogenesis: An integrated view. Clin. Microbiol. Rev. 2009, 22, 564–581. [Google Scholar] [CrossRef] [PubMed]
- Mathew, A. Humanized mouse models to study human cell-mediated and humoral responses to dengue virus. Curr. Opin. Virol. 2017, 25, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Coronel-Ruiz, C.; Gutiérrez-Barbosa, H.; Medina-Moreno, S.; Velandia-Romero, M.L.; Chua, J.V.; Castellanos, J.E.; Zapata, J.C. Humanized mice in dengue research: A comparison with other mouse models. Vaccines 2020, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Yong, K.S.M.; Her, Z.; Chen, Q. Humanized Mouse Models for the Study of Hepatitis C and Host Interactions. Cells 2019, 8, 604. [Google Scholar] [CrossRef] [PubMed]
- Lai, F.; Chen, Q. Humanized mouse models for the study of infection and pathogenesis of human viruses. Viruses 2018, 10, 643. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masemann, D.; Ludwig, S.; Boergeling, Y. Advances in Transgenic Mouse Models to Study Infections by Human Pathogenic Viruses. Int. J. Mol. Sci. 2020, 21, 9289. https://doi.org/10.3390/ijms21239289
Masemann D, Ludwig S, Boergeling Y. Advances in Transgenic Mouse Models to Study Infections by Human Pathogenic Viruses. International Journal of Molecular Sciences. 2020; 21(23):9289. https://doi.org/10.3390/ijms21239289
Chicago/Turabian StyleMasemann, Dörthe, Stephan Ludwig, and Yvonne Boergeling. 2020. "Advances in Transgenic Mouse Models to Study Infections by Human Pathogenic Viruses" International Journal of Molecular Sciences 21, no. 23: 9289. https://doi.org/10.3390/ijms21239289
APA StyleMasemann, D., Ludwig, S., & Boergeling, Y. (2020). Advances in Transgenic Mouse Models to Study Infections by Human Pathogenic Viruses. International Journal of Molecular Sciences, 21(23), 9289. https://doi.org/10.3390/ijms21239289