Nemo-Like Kinase in Development and Diseases: Insights from Mouse Studies


{kind=link}
Abstract
1. Introduction
1.1. Wnt Signalling Pathway
1.2. Nemo-Like Kinase
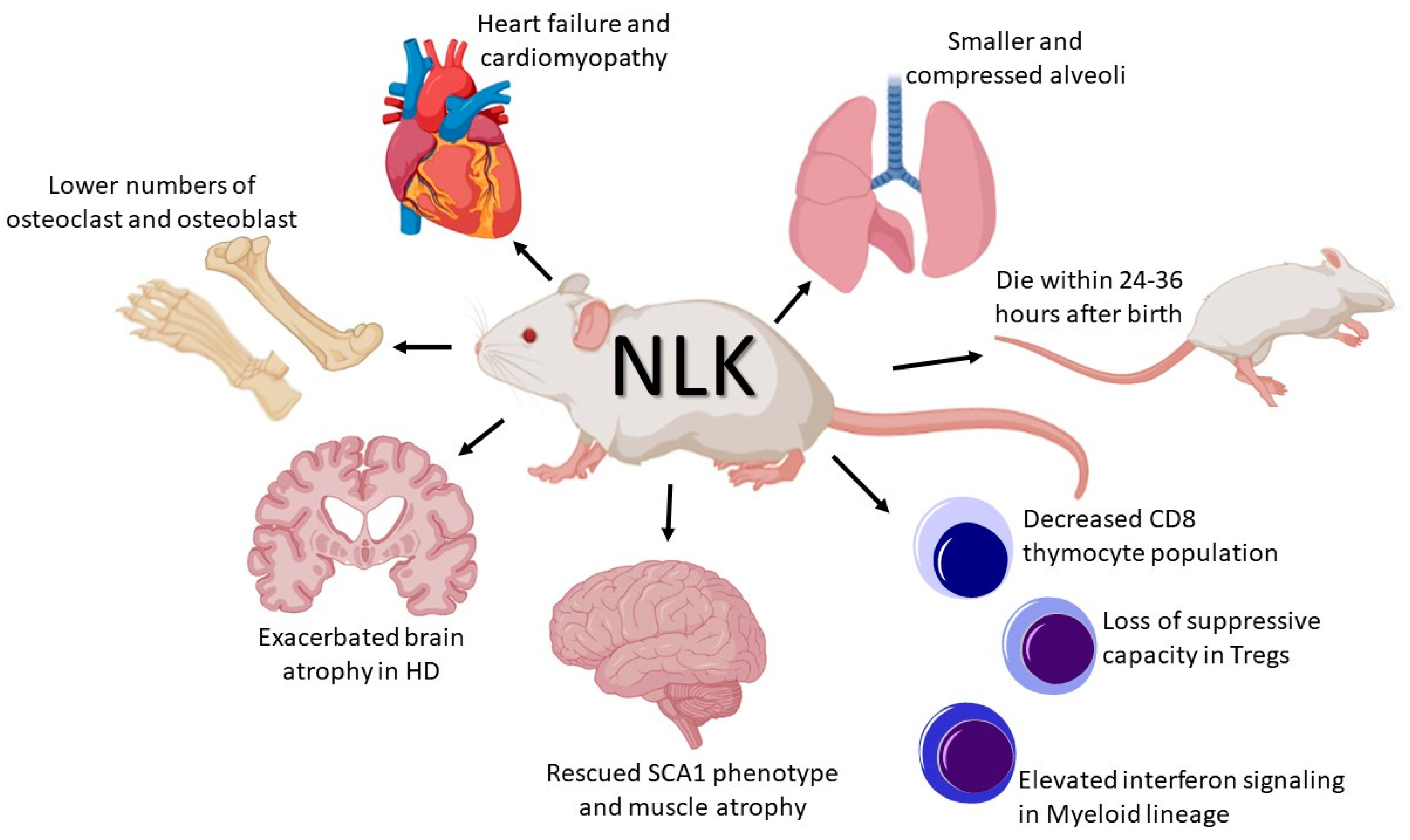
2. NLK and Development
3. NLK and the Immune System
4. NLK and Neuronal Disorders
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Steinhart, Z.; Angers, S. Wnt signaling in development and tissue homeostasis. Development 2018, 145, dev146589. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.P.; Chopra, V.L. Effect of the Wingless (wg1) mutation on wing and haltere development in Drosophila melanogaster. Dev. Biol. 1976, 48, 461–465. [Google Scholar] [CrossRef]
- Nüsslein-Volhard, C.; Wieschaus, E. Mutations affecting segment number and polarity in Drosophila. Nature 1980, 287, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R.; Varmus, H.E. Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome. Cell 1982, 31, 99–109. [Google Scholar] [CrossRef]
- Van Ooyen, A.; Nusse, R. Structure and nucleotide sequence of the putative mammary oncogene int-1; proviral insertions leave the protein-encoding domain intact. Cell 1984, 39, 233–240. [Google Scholar] [CrossRef]
- Rijsewijk, F.; Schuermann, M.; Wagenaar, E.; Parren, P.; Weigel, D.; Nusse, R. The Drosophila homolog of the mouse mammary oncogene int-1 is identical to the segment polarity gene wingless. Cell 1987, 50, 649–657. [Google Scholar] [CrossRef]
- Nusse, R.; Brown, A.; Papkoff, J.; Scambler, P.; Shackleford, G.; McMahon, A.; Moon, R.; Varmus, H. A new nomenclature for int-1 and related genes: The Wnt gene family. Cell 1991, 64, 231. [Google Scholar] [CrossRef]
- Komiya, Y.; Habas, R. Wnt signal transduction pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef]
- Latres, E.; Chiaur, D.S.; Pagano, M. The human F box protein β-Trcp associates with the Cul1/Skp1 complex and regulates the stability of β-catenin. Oncogene 1999, 18, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Wnt/β-Catenin Signaling in Development and Disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Coulombe, P.; Meloche, S. Atypical mitogen-activated protein kinases: Structure, regulation and functions. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2007, 1773, 1376–1387. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.-W.; Benzer, S. Rotation of photoreceptor clusters in the developing drosophila eye requires the nemo gene. Cell 1994, 78, 125–136. [Google Scholar] [CrossRef]
- Brott, B.K.; Pinsky, B.A.; Erikson, R.L. Nlk is a murine protein kinase related to Erk/MAP kinases and localized in the nucleus. Proc. Natl. Acad. Sci. USA 1998, 95, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Ishitani, T.; Ninomiya-Tsuji, J.; Nagai, S.I.; Nishita, M.; Meneghini, M.; Barker, N.; Waterman, M.; Bowerman, B.; Clevers, H.; Shibuya, H.; et al. The TAK1-NLK-MAPK-related pathway antagonizes signalling between beta-catenin and transcription factor TCF. Nature 1999, 399, 798–802. [Google Scholar] [CrossRef]
- Ishitani, T.; Ninomiya-Tsuji, J.; Matsumoto, K. Regulation of Lymphoid Enhancer Factor 1/T-Cell Factor by Mitogen-Activated Protein Kinase-Related Nemo-Like Kinase-Dependent Phosphorylation in Wnt/β-Catenin Signaling. Mol. Cell. Biol. 2003, 23, 1379–1389. [Google Scholar] [CrossRef]
- Li, M.; Wang, H.; Huang, T.; Wang, J.; Ding, Y.; Li, Z.; Zhang, J.; Li, L. TAB2 scaffolds TAK1 and NLK in repressing canonical Wnt signaling. J. Biol. Chem. 2010, 285, 13397–13404. [Google Scholar] [CrossRef]
- Yamada, M.; Ohnishi, J.; Ohkawara, B.; Iemura, S.; Satoh, K.; Hyodo-Miura, J.; Kawachi, K.; Natsume, T.; Shibuya, H. NARF, an nemo-like kinase (NLK)-associated ring finger protein regulates the ubiquitylation and degradation of T cell factor/lymphoid enhancer factor (TCF/LEF). J. Biol. Chem. 2006, 281, 20749–20760. [Google Scholar] [CrossRef]
- Ishitani, T.; Ishitani, S. Nemo-like kinase, a multifaceted cell signaling regulator. Cell. Signal. 2013, 25, 190–197. [Google Scholar] [CrossRef]
- Ota, S.; Ishitani, S.; Shimizu, N.; Matsumoto, K.; Itoh, M.; Ishitani, T. NLK positively regulates Wnt/β-catenin signalling by phosphorylating LEF1 in neural progenitor cells. EMBO J. 2012, 31, 1904–1915. [Google Scholar] [CrossRef] [PubMed]
- Ohkawara, B.; Shirakabe, K.; Hyodo-Miura, J.; Matsuo, R.; Ueno, N.; Matsumoto, K.; Shibuya, H. Role of the TAK1-NLK-STAT3 pathway in TGF-beta-mediated mesoderm induction. Genes Dev. 2004, 18, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Kojima, H.; Sasaki, T.; Ishitani, T.; Iemura, S.I.; Zhao, H.; Kaneko, S.; Kunimoto, H.; Natsume, T.; Matsumoto, K.; Nakajima, K. STAT3 regulates Nemo-like kinase by mediating its interaction with IL-6-stimulated TGFβ-activated kinase 1 for STAT3 Ser-727 phosphorylation. Proc. Natl. Acad. Sci. USA 2005, 102, 4524–4529. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lin, Q.; Ni, T.; Zhao, J.; Lin, F.; Lu, X.; Lv, Y.; Ren, S.; Liu, Z.; Zhang, T.; et al. NLK interacts with 14-3-3ζ to restore the expression of E-cadherin. Oncol. Rep. 2020, 43, 1845–1852. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Z.; Zhang, H.H.; Liang, J.B.; Song, Y.; Jin, B.X.; Xing, N.N.; Fan, G.C.; Du, R.L.; Zhang, X.D. Nemo-like kinase (NLK) negatively regulates NF-kappa B activity through disrupting the interaction of TAK1 with IKKβ. Biochim. Biophys. Acta 2014, 1843, 1365–1372. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ishitani, T.; Hirao, T.; Suzuki, M.; Isoda, M.; Ishitani, S.; Harigaya, K.; Kitagawa, M.; Matsumoto, K.; Itoh, M. Nemo-like kinase suppresses Notch signalling by interfering with formation of the Notch active transcriptional complex. Nat. Cell Biol. 2010, 12, 278–285. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, Y.; He, Y.; Li, J. The emerging role of Nemo-like kinase (NLK) in the regulation of cancers. Tumor Biol. 2015, 36, 9147–9152. [Google Scholar] [CrossRef]
- Yasuda, J.; Tsuchiya, A.; Yamada, T.; Sakamoto, M.; Sekiya, T.; Hirohashi, S. Nemo-like kinase induces apoptosis in DLD-1 human colon cancer cells. Biochem. Biophys. Res. Commun. 2003, 308, 227–233. [Google Scholar] [CrossRef]
- Shaw-Hallgren, G.; Masoumi, K.C.; Zarrizi, R.; Hellman, U.; Karlsson, P.; Helou, K.; Massoumi, R. Association of Nuclear-Localized Nemo-Like Kinase with Heat-Shock Protein 27 Inhibits Apoptosis in Human Breast Cancer Cells. PLoS ONE 2014, 9, e96506. [Google Scholar] [CrossRef]
- Yang, Y.; Zhe, H.; Massoumi, R.; Ke, H. Decreased expression of nemo-like kinase in melanoma is correlated with increased vascularity and metastasis. Melanoma Res. 2019, 29, 376–381. [Google Scholar] [CrossRef]
- Shen, N.; Duan, X.H.; Wang, X.L.; Yang, Q.Y.; Feng, Y.; Zhang, J.X. Effect of NLK on the proliferation and invasion of laryngeal carcinoma cells by regulating CDCP1. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6226–6233. [Google Scholar] [PubMed]
- Röttinger, E.; Croce, J.; Lhomond, G.; Besnardeau, L.; Gache, C.; Lepage, T. Nemo-like kinase (NLK) acts downstream of Notch/Delta signalling to downregulate TCF during mesoderm induction in the sea urchin embryo. Development 2006, 133, 4341. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, C.J.; Moon, R.T. Nemo-like kinase is an essential co-activator of Wnt signaling during early zebrafish development. Development 2004, 131, 2899. [Google Scholar] [CrossRef] [PubMed]
- Ota, R.; Kotani, T.; Yamashita, M. Possible Involvement of Nemo-like Kinase 1 in Xenopus Oocyte Maturation as a Kinase Responsible for Pumilio1, Pumilio2, and CPEB Phosphorylation. Biochemistry 2011, 50, 5648–5659. [Google Scholar] [CrossRef]
- Cheng, X.; Liang, J.; Teng, Y.; Fu, J.; Miao, S.; Zong, S.; Wang, L. Nemo-like kinase promotes etoposide-induced apoptosis of male germ cell-derived GC-1 cells in vitro. FEBS Lett. 2012, 586, 1497–1503. [Google Scholar] [CrossRef]
- Ke, H.; Masoumi, K.C.; Ahlqvist, K.; Seckl, M.J.; Rydell-Törmänen, K.; Massoumi, R. Nemo-like kinase regulates the expression of vascular endothelial growth factor (VEGF) in alveolar epithelial cells. Sci. Rep. 2016, 6, 23987. [Google Scholar] [CrossRef]
- Masoumi, K.C.; Daams, R.; Sime, W.; Siino, V.; Ke, H.; Levander, F.; Massoumi, R. NLK-mediated phosphorylation of HDAC1 negatively regulates Wnt signaling. Mol. Biol. Cell 2017, 28, 346–355. [Google Scholar] [CrossRef]
- Liu, R.; Khalil, H.; Lin, S.-C.J.; Sargent, M.A.; York, A.J.; Molkentin, J.D. Nemo-Like Kinase (NLK) Is a Pathological Signaling Effector in the Mouse Heart. PLoS ONE 2016, 11, e0164897. [Google Scholar] [CrossRef]
- Canalis, E.; Kranz, L.; Zanotti, S. Nemo-Like Kinase Regulates Postnatal Skeletal Homeostasis. J. Cell. Physiol. 2014, 229, 1736–1743. [Google Scholar] [CrossRef]
- Staal, F.J.T.; Luis, T.C.; Tiemessen, M.M. WNT signalling in the immune system: WNT is spreading its wings. Nat. Rev. Immunol. 2008, 8, 581–593. [Google Scholar] [CrossRef]
- Chae, W.-J.; Bothwell, A.L.M. Canonical and Non-Canonical Wnt Signaling in Immune Cells. Trends Immunol. 2018, 39, 830–847. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, R.; Fang, X.; Sun, Z. β-catenin/TCF-1 pathway in T cell development and differentiation. J. Neuroimmune Pharmacol. Off. J. Soc. Neuroimmune Pharmacol. 2012, 7, 750–762. [Google Scholar] [CrossRef] [PubMed]
- Van de Wetering, M.; de Lau, W.; Clevers, H. WNT signaling and lymphocyte development. Cell 2002, 109, S13–S19. [Google Scholar] [CrossRef]
- Van Loosdregt, J.; Coffer, P.J. The Role of WNT Signaling in Mature T Cells: T Cell Factor Is Coming Home. J. Immunol. 2018, 201, 2193. [Google Scholar] [CrossRef] [PubMed]
- Daams, R.; Sime, W.; Leandersson, K.; Sitnicka, E.; Massoumi, R. Deletion of Nemo-like Kinase in T Cells Reduces Single-Positive CD8(+) Thymocyte Population. J. Immunol. 2020, 205, 1830–1841. [Google Scholar] [CrossRef] [PubMed]
- Fleskens, V.; Minutti, C.M.; Wu, X.; Wei, P.; Pals, C.E.; McCrae, J.; Hemmers, S.; Groenewold, V.; Vos, H.J.; Rudensky, A.; et al. Nemo-like Kinase Drives Foxp3 Stability and Is Critical for Maintenance of Immune Tolerance by Regulatory T Cells. Cell Rep. 2019, 26, 3600–3612.e6. [Google Scholar] [CrossRef] [PubMed]
- Kortenjann, M.; Nehls, M.; Smith, A.J.; Carsetti, R.; Schueler, J.; Koehler, G.; Boehm, T. Abnormal bone marrow stroma in mice deficient for nemo-like kinase, Nlk. Eur. J. Immunol. 2001, 31, 3580–3587. [Google Scholar] [CrossRef]
- Swann, J.B.; Happe, C.; Boehm, T. Elevated levels of Wnt signaling disrupt thymus morphogenesis and function. Sci. Rep. 2017, 7, 785. [Google Scholar] [CrossRef]
- Li, S.Z.; Shu, Q.P.; Song, Y.; Zhang, H.H.; Liu, Y.; Jin, B.X.; Liuyu, T.Z.; Li, C.; Huang, X.C.; Du, R.L.; et al. Phosphorylation of MAVS/VISA by Nemo-like kinase (NLK) for degradation regulates the antiviral innate immune response. Nat. Commun. 2019, 10, 3233. [Google Scholar] [CrossRef]
- Ju, H.; Kokubu, H.; Todd, T.W.; Kahle, J.J.; Kim, S.; Richman, R.; Chirala, K.; Orr, H.T.; Zoghbi, H.Y.; Lim, J. Polyglutamine disease toxicity is regulated by Nemo-like kinase in spinocerebellar ataxia type 1. J. Neurosci. 2013, 33, 9328–9336. [Google Scholar] [CrossRef]
- Todd, T.W.; Kokubu, H.; Miranda, H.C.; Cortes, C.J.; La Spada, A.R.; Lim, J. Nemo-like kinase is a novel regulator of spinal and bulbar muscular atrophy. Elife 2015, 4, e08493. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, X.; Liu, H.; LeBron, J.; Alexandris, A.; Peng, Q.; Gu, H.; Yang, F.; Li, Y.; Wang, R.; et al. Nemo-like kinase reduces mutant huntingtin levels and mitigates Huntington’s disease. Hum. Mol. Genet. 2020, 29, 1340–1352. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daams, R.; Massoumi, R. Nemo-Like Kinase in Development and Diseases: Insights from Mouse Studies. Int. J. Mol. Sci. 2020, 21, 9203. https://doi.org/10.3390/ijms21239203
Daams R, Massoumi R. Nemo-Like Kinase in Development and Diseases: Insights from Mouse Studies. International Journal of Molecular Sciences. 2020; 21(23):9203. https://doi.org/10.3390/ijms21239203
Chicago/Turabian StyleDaams, Renée, and Ramin Massoumi. 2020. "Nemo-Like Kinase in Development and Diseases: Insights from Mouse Studies" International Journal of Molecular Sciences 21, no. 23: 9203. https://doi.org/10.3390/ijms21239203
APA StyleDaams, R., & Massoumi, R. (2020). Nemo-Like Kinase in Development and Diseases: Insights from Mouse Studies. International Journal of Molecular Sciences, 21(23), 9203. https://doi.org/10.3390/ijms21239203