1. Introduction
Mammalian skin is composed of three layers: epidermis, dermis and hypodermis. Epidermis is built up by stratified keratinocytes separated from the underlying dermis by a basement membrane. Dermis is comprised of both a fibroblast-rich dermis and an underlying dermal white adipose tissue (dWAT). Hypodermis, formed by connective tissues and adipose tissues, in rodents is separated from the dermis by a layer of striated muscle (
panniculus carnosus). dWAT forms the adipose compartment within the dermis which is primarily composed of adipocytes defined as intradermal adipocytes [
1]. This unique population of fat cells actively participates in the various physiological and pathological processes in the skin. A decrease in the ambient temperature increases the thickness of the dWAT layer, which indicates the possible role of intradermal adipocytes in generating heat and maintaining a constant temperature, thus participating in thermoregulation [
2]. Studies by Festa et al. showed that the resident intradermal adipocytes contribute to follicular stem cell activation and control hair follicle regeneration by producing platelet-derived growth factor (PDGF) signals [
3]. dWAT was additionally identified as contributor in the immune defense response induced by bacterial infections of the skin [
4]. It was also delineated that resident intradermal adipocytes participate in the skin wound healing process by promoting fibroblast production and increasing their migration into the wound bed [
5]. Moreover, loss of dWAT is observed in murine models of dermal fibrosis, whereas a lack of dWAT is characteristic for patients with systematic sclerosis (SSc; scleroderma) [
6]. Treatment with bleomycin (an inducer of dermal fibrosis in mice), resulted in the reduction of the dWAT layer and a decrease in the levels of adipogenic markers (Pparγ2, Fabp4, AdipoQ) in C57BL/6J mice [
7]. Further research using aP2-Cre–transgenic mice treated with bleomycin showed that loss of intradermal adipocytes leads to dermal fibrosis [
7]. However, despite increasing research interest in the role of dWAT in skin physiology, the current knowledge regarding dWAT homeostasis and the molecular pathways which modulate its physiology is limited. One possible factor involved in dWAT regulation is the epidermal Wnt/β-catenin signaling pathway. Donati et al. demonstrated that 3T3-L1 preadipocytes stimulated by adipogenic media and cultured in keratinocytes conditioned media from K14Cre/CatnbFlox(ex3)/+ mice, in which K14-positive epidermal basal keratinocytes express β-catenin, differentiate into adipocytes more efficiently than those cultured in keratinocyte-conditioned media from wild type mice [
8]. The authors concluded that adipocyte differentiation was induced by epidermal Wnt/β-catenin through secretion of ligands for the Bmp and insulin signaling pathways [
8].
Another factor that may participate in dWAT homeostasis is the transcription factor Foxn1. In the skin, epidermally expressed Foxn1 is responsible for the initiation of the keratinocytes’ terminal differentiation program (suprabasal layer of epidermis) and stimulation of epithelial cell proliferation (basal layer) [
9,
10]. Previous studies have reported the regulatory role of Foxn1 in the reparative (scar-forming) skin wound healing by participation in re-epithelialization and by promoting epithelial to mesenchymal transition (EMT) [
11,
12]. Our recent data demonstrated that Foxn1 may regulate susceptibility to diet-induced obesity. Indeed, activation/nonactivation of Foxn1 may play a role in modulating body weight/metabolic homeostasis [
13]. Moreover, partial inactivation of Foxn1 abrogated adipogenesis in intact and injured skin by possible impact on the expression of adipogenesis-promoting genes Bmp2 and Igf2 [
13].
In the present study, we examined how age and diet (1) affect the adipogenic potential of DFs and (2) modulate the dWAT compartment in the skin. We also investigated the potential role of epidermally expressed Foxn1 on DF adipogenic potential through Bmp2 and Igf2 signaling contribution using the coculture model of keratinocytes and differentiated DFs.
3. Discussion
In recent years, there has been an increasing interest in dermal white adipose tissue (dWAT), a newly recognized fat depot within the dermal skin compartment [
1,
15,
26]. Due to the fact that dWAT is developmentally, biochemically and functionally different from other white adipose tissue/fat depots (i.e., subcutaneous, intraperitoneal) [
2,
26] it is essential to examine and understand its function and role in skin homeostasis. It has been well-established that dWAT is regulated in a manner that is distinct from other fat depots, displaying parallels to hair follicle cycling. During the active growth phase of the hair cycle (anagen), dWAT thickness increases while following the regression stage of the hair cycle (catagen), dWAT thickness decreases considerably [
8,
27,
28]. Furthermore, the population of immature dWAT adipocytes plays a role in proper hair cycling by participation in induction of telogen to anagen transition [
3].
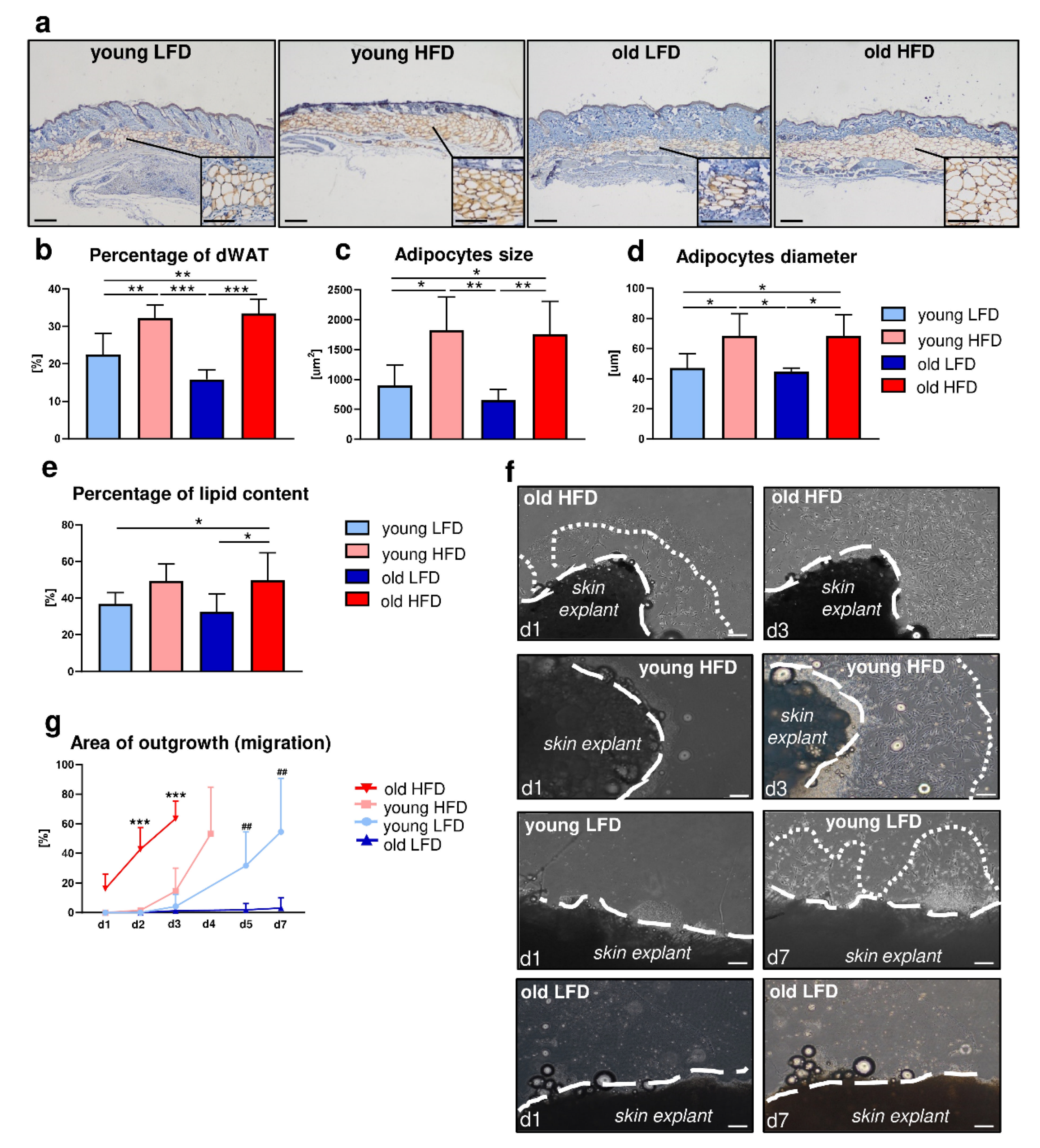
In our study we examined how systematic/environmental changes, i.e., diet (LFD vs. HFD) and age (young vs. old), affect dWAT and its cellular component. We showed that (1) HFD during a period of 8 weeks stimulates dWAT expansion in the context of dWAT area extension in the skin, adipocyte diameter/size and percentage of extracted lipids, regardless of animals’ age (young vs. old mice) and (2) the increase in dWAT content due to HFD is achieved at least partially through hyperthrophy as indicated by the increase in adipocytes size and diameter. This data suggests that dWAT, similar to other fat depots, is regulated by diet as presented previously [
29]. However, evidence from other fat depots, namely comparison of epididymal (eWAT) vs. inguinal (iWAT), revealed that the differences in fat tissue enlargement due to an obesogenic environment was achieved through hypertrophy (eWAT) and/or hyperplasia (iWAT) [
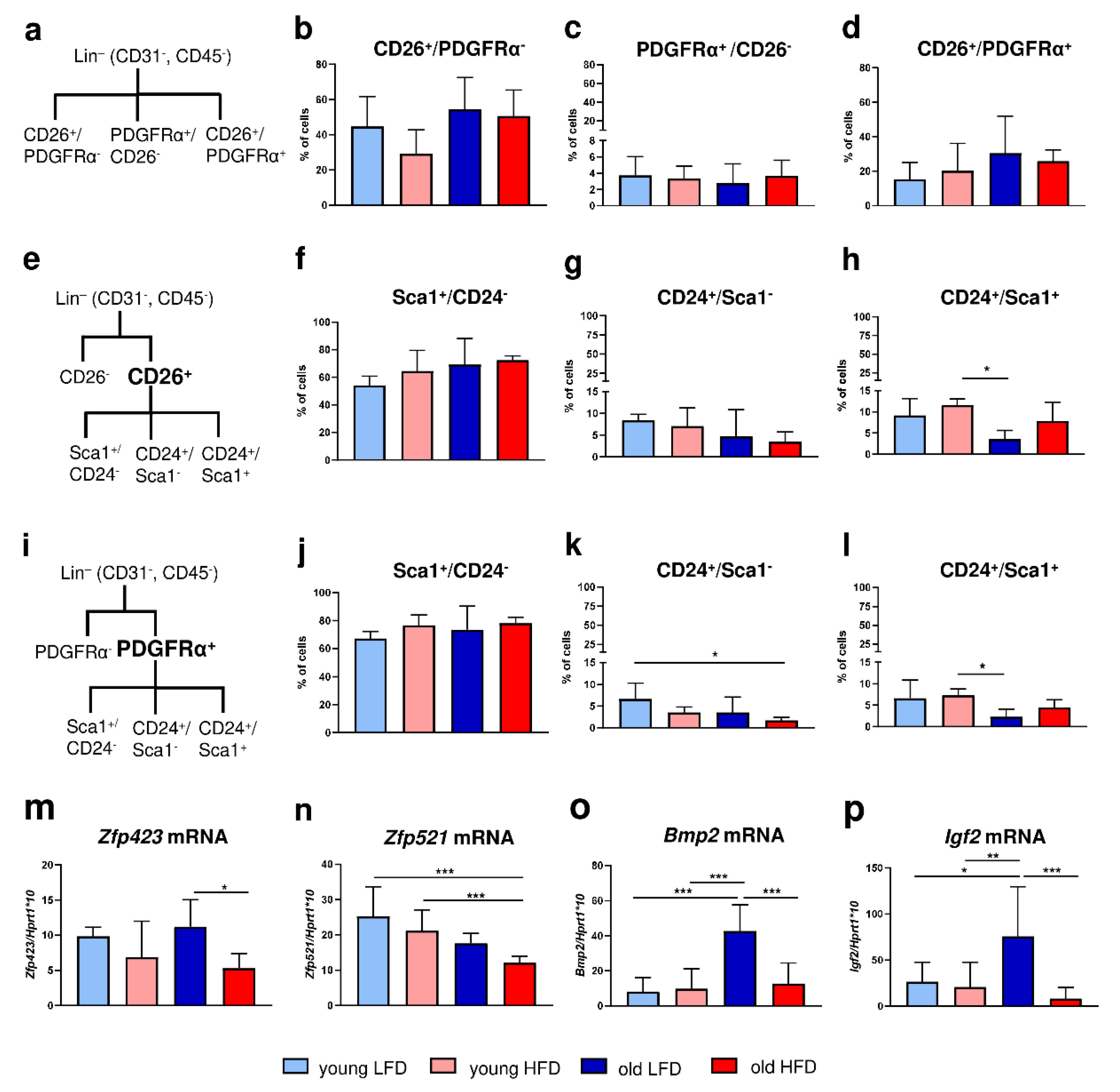
30]. To shed light on changes related to age and/or diet, phenotypic characterization of DFs with adipogenic properties was performed. We distinguished three major skin cell populations: CD26
+ (approx. 60%), CD26
+/PDGFRα
+ (approx. 20%) and PDGFRα
+ (approx. 4%), the latter group associated with the predominant profibrotic nature of DFs. The data also showed that those populations were percentage stable and were not affected by animals’ age or diet. Further analyses revealed that the subpopulation of PDGFRα
+/CD24
+/Sca1
+ DFs, previously recognized as adipocyte progenitors [
17,
18], decreased with animals’ age particularly in the group fed LFD, indicating a reduced capacity to differentiate into mature adipocytes. Interestingly, the detected decrease observed in the CD24
+ cell population due to age seemed to be compensated by the increase in the percentage of Sca1
+ in the old HFD mice. These results are in line with previous findings showing that the loss of adipocyte progenitor cells with aging may affect the wound healing process [
31]. Further evidence of the reduced pool of adipocyte progenitor cells with age is provided by the analysis of the transcription factor
Zfp521 expression. Kang et al. identified Zfp521 as a potent repressor of adipogenesis in vivo and in vitro [
21]. Our data indicate that expression of
Zfp521 decreased with animals’ age correlating with a decrease in the population of CD24
+ cells. Another transcriptional regulator involved in preadipocyte determination is
Zfp423 [
20]. The decrease observed in the expression of
Zfp423 of animals fed HFD may indicate that the pool of cells with adipogenic potential achieved adipogenic differentiation capacity whereas higher mRNA of
Zfp423 levels in DFs isolated from mice fed LFD may indicate that it still contains a population of DFs with adipogenic ability. However, analyses to evaluate whether changes in dWAT expansion involves adipocyte stem cell/progenitor recruitment and if advanced age of animals affects the pool of dWAT progenitors, will require further in vivo study, i.e., involving label-retaining cells (LRC) analysis [
30]. Nevertheless, the present data broadens the complexity of dWAT regulation by showing that in addition to its local regulation by hair cycling, the systemic factors age and diet exhibit considerable impact.
Previous studies on subcutaneous adipose tissue have indicated that the size of the fat cell was associated with its secretory activity of adipokines and expression of pro- and anti-inflammatory factors [
32,
33]. Moreover, heterogeneity of adipocyte size affects subsequent alternations in lipid and energy metabolism. This is consistent with our results which showed that due to the obesogenic environment the total content of lipids in the skin increased which correlated with intradermal adipocyte size and the general content of dWAT in uninjured skin. Considering the secretory role of dWAT in the wound healing process [
34], understanding mechanisms of age- and diet-dependent changes on lipid content in the skin remains an important issue for future research.
Schmidt and Horsley demonstrated that the population of adipocyte precursor cells and mature adipocytes is necessary for efficient skin wound repair [
5]. The presence of intradermal adipocytes affects the function of DFs by stimulating their migration to the wound bed [
5]. Authors also showed that adipocyte-conditioned medium applied to explants of tail skin enhanced dermal fibroblast migration but had no effect on DF proliferation [
5]. Our present study based on an ex vivo model revealed that HFD accelerated dermal fibroblasts’ migratory abilities and increased content of dWAT in uninjured skin. Interestingly, migration of DFs was significantly enhanced in old animals fed HFD especially in contrast to old animals fed LFD. This may be an additional evidence of compensatory role of HFD on skin homeostasis in older animals. Our previous studies indicated that mice fed HFD had a thicker dWAT layer which may compensate for the loss in age-related thickness of the fibroblast-rich dermis and may have a beneficial effect on the skin wound healing process [
14]. Although several studies specify caloric restriction as a favorable dietary intervention which can delay age-related changes in the structure and metabolism of the skin [
19,
35], the physiological increase in dWAT with age may be beneficial as our previous [
14] and present study indicate.
Despite increasing interest in the role of dWAT in skin physiology, the current knowledge considering dWAT homeostasis and molecular pathways is limited. Donati et al. demonstrated that the epidermal Wnt/β-catenin signaling pathway is a key initiator of the signal transduction cascade that stimulates the process of adipogenesis in the skin [
8]. Furthermore, the supplementation of adipogenic media with Bmp2, Bmp6 and Igf2 proteins also stimulated the process of adipogenesis suggesting that the Wnt/β-catenin pathway promotes adipocyte differentiation by stimulating secretion of ligands for the Bmp (Bmp2/Bmp6) and insulin (Igf2) signaling pathways [
8]. Our recently published data revealed that the transcription factor Foxn1, whose expression is limited to epithelial cells of the skin, may affect skin adiposity. A series of in vivo experiments using transgenic Foxn1::Egfp mice (Foxn1
+/−) and B6 mice (Foxn1
+/+) demonstrated that young and old Foxn1
+/− mice fed HFD gained significantly less body weight than their male B6 counterparts, indicating the resistance of heterozygous Foxn1
+/− mice to diet-induced obesity [
13]. Moreover, expression of adipogenic-related genes (
Cepbα,
Pparγ,
Fabp4,
leptin) was abrogated in injured and intact skin of Foxn1
+/− mice. Interestingly, we observed that with gradual decrease of Foxn1 activation in the skin of Foxn1
+/+, Foxn1
+/− and Foxn1
−/− mice, expression of
Bmp2 and
Igf2 decreased correspondingly [
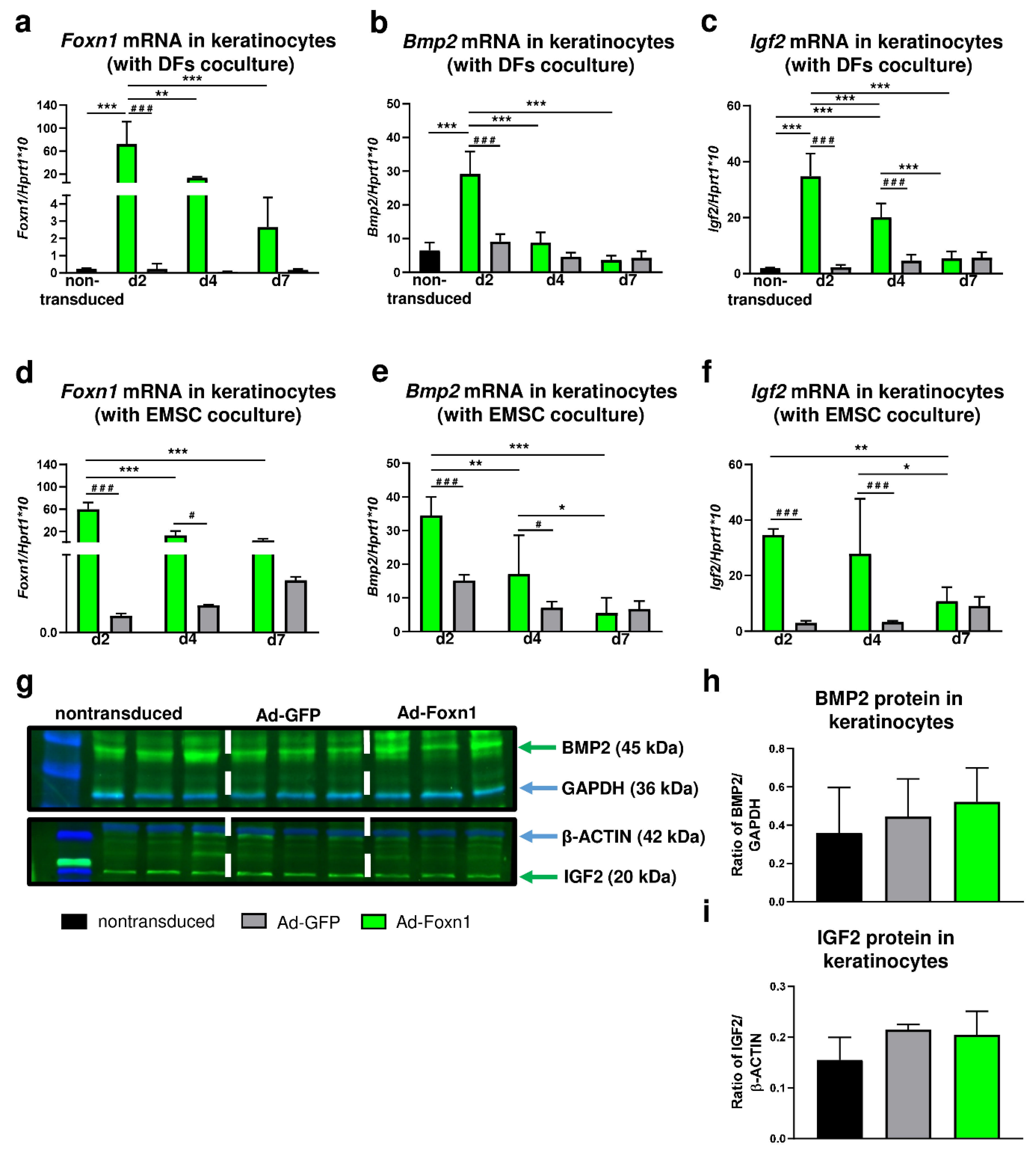
13]. In the present study, we analyzed the expression of
Bmp2 and
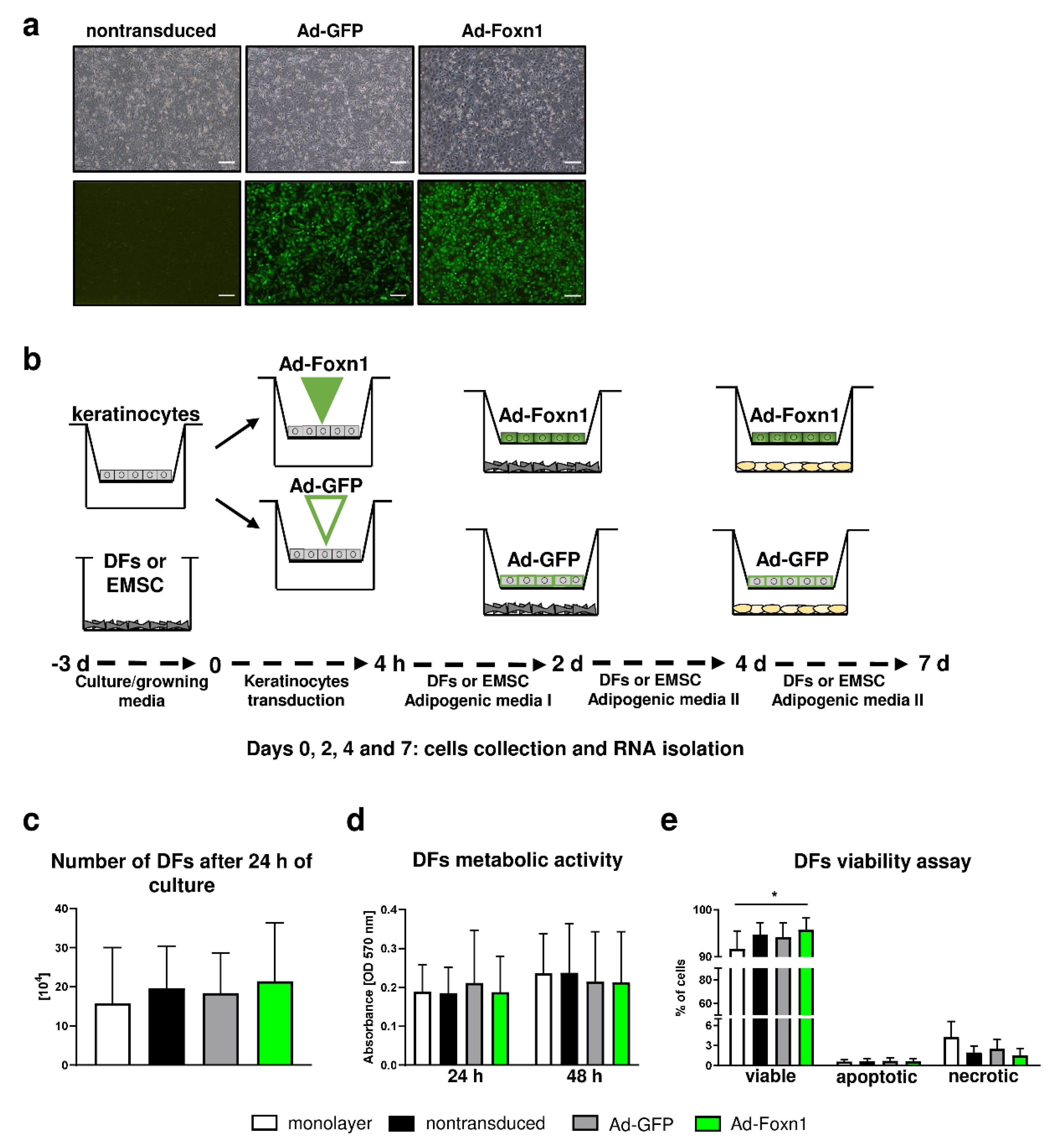
Igf2 in keratinocytes after transduction with Foxn1-GFP-expressing (Ad-Foxn1) or control (Ad-GFP) adenoviruses. In vitro experiments revealed increased expression of
Bmp2 and
Igf2 with overexpression of Foxn1 confirming the relationship between the Foxn1 activity and expression of adipogenesis-promoting genes.
Considering participation of Foxn1 in the Bmp2 and Igf2 signaling pathways [
13] and the previously described double paracrine mechanism between keratinocytes and fibroblasts [
24], we analyzed DF adipogenic differentiation in a coculture system with Ad-Foxn1- vs. Ad-GFP- (control) transduced keratinocytes. The analysis of adipogenic genes (
Cebpα,
Pparγ,
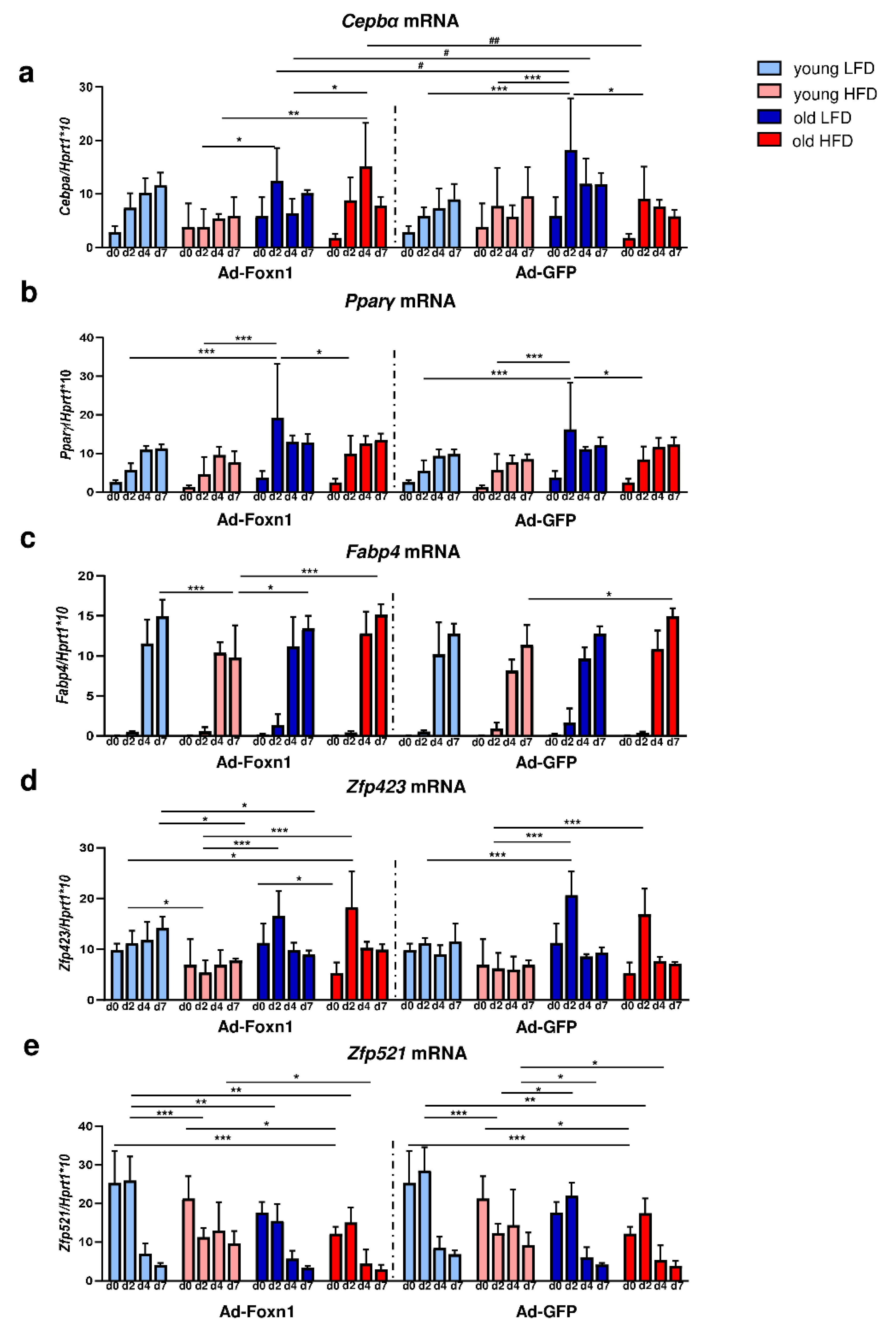
Fabp4) in differentiating DFs showed differences among the groups due to age and diet, especially for markers characteristic for the initial stage of the adipogenesis process (
Cebpα and
Pparγ). Surprisingly, the highest expression of adipogenic genes was observed in DFs isolated from old animals fed LFD. The changes detected in aged DFs may possibly cause the loss of functional identity and the acquisition of pro-adipogenic features. Salzer et al. revealed that during aging DFs displayed reduction in extracellular matrix (ECM) genes i.e., collagens and glycosaminogycans with simultaneously higher expression of markers of the immune response and the adipogenesis process, including lipogenesis and fat cell differentiation [
19].
The differences in the process of DF differentiation due to Ad-Foxn1 presence in keratinocytes has been exclusively detected for
Cebpα expression (see
Figure 5a). This mild effect of Foxn1 on DF adipogenesis in our present in vitro study may indicate that this model is not sufficient to mirror the in vivo conditions. However, considering the robust expression of Foxn1 in hair follicles and interfollicular epidermis [
9,
36,
37], changes in the adiposity in Foxn1
+/+, Foxn1
+/− and Foxn1
−/− mice [
13], the stimulatory effect of Foxn1 in keratinocytes on the release of adipogenic signaling factors Bmp2, Igf2 (present data) and the decline in the skin Foxn1 expression in very old mice [
13], it strongly suggests a role of Foxn1 in dWAT physiology.
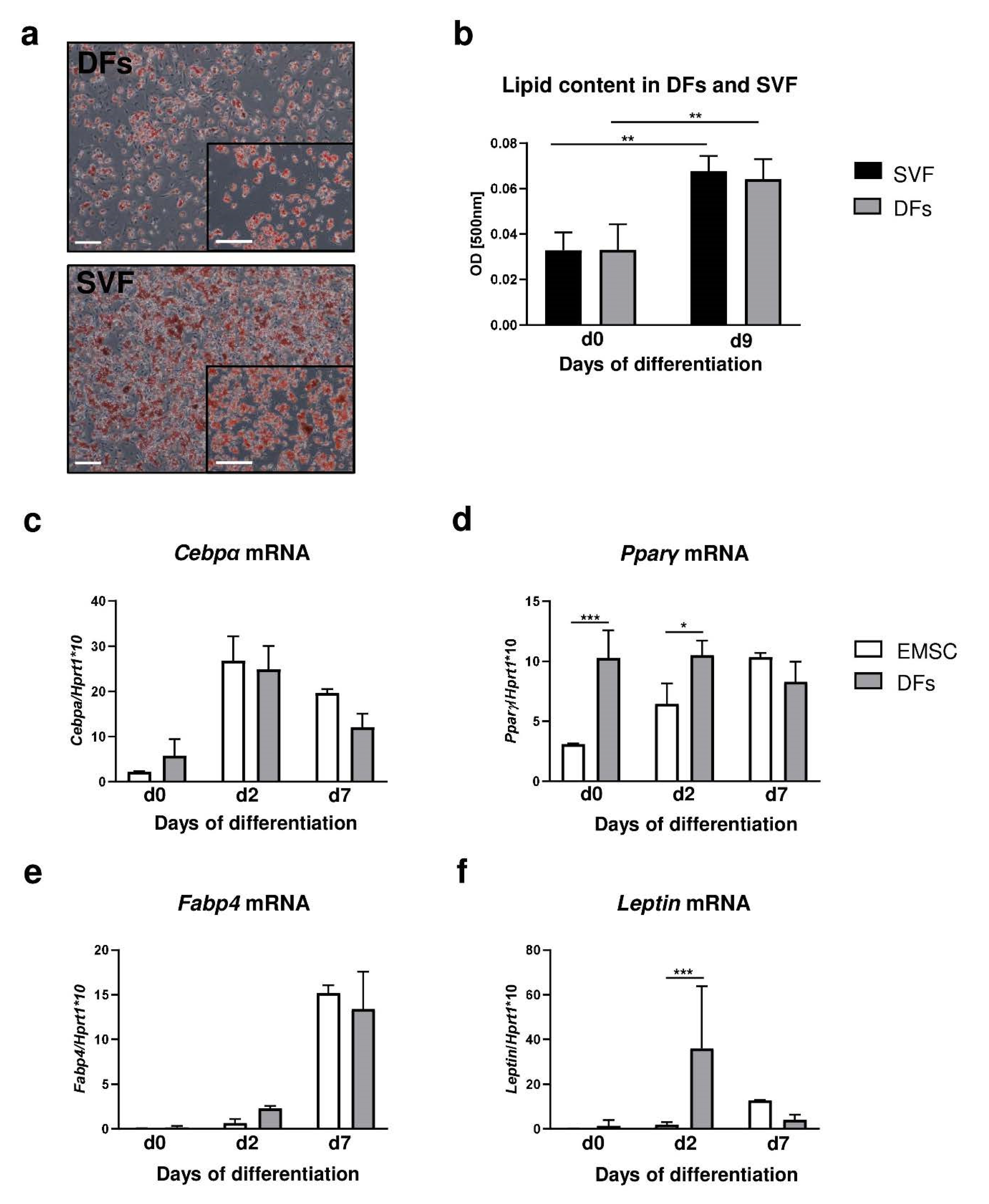
Collectively, our data indicate that diet and age regulate skin homeostasis influencing dWAT morphology and functionality. DFs display the adipogenic potential of differentiation that is comparable to that of SVF of fat depot and EMSC models. Furthermore, we showed that age and diet affect the pool of DFs with adipogenic properties. We also propose that the transcription factor Foxn1 expressed in epidermis has a stimulatory effect on the adipogenic signaling of Bmp2 and Igf2.
4. Materials and Methods
4.1. Animals
The studies were performed on B6 mice. All mice were obtained from colonies established at the animal facility of the Institute of Animal Reproduction and Food Research, The Polish Academy of Sciences. B6 mice were originally purchased from the Jackson Laboratory (Bar Harbor, ME, USA). Animals were bred and housed in a temperature- and humidity-controlled room (22 °C ± 2 °C and 35–65%, respectively) with a 12-h light/12-h dark cycle at the Institute of Animal Reproduction and Food Research, Polish Academy of Sciences, Olsztyn, Poland.
Young (2 months) and old (18 months old) mice were divided into two experimental groups fed (1) a low-fat diet (11% kcal from fat, PicoLab Rodent Diet 20 5053) and (2) a high-fat diet (58% kcal from fat, TestDiet AIN-76A; LabDiet) for 8 weeks. Skin samples from the back of mice were collected postmortem for cell isolation, ex vivo explants or histological assays.
The experimental animal procedures performed in these studies were approved 29 April 2015 by the Ethics Committee of The University of Warmia and Mazury (Olsztyn, Poland), No. 22/2015.
4.2. Histology
Immunohistochemical detection of perilipin (1:200, ab3526, Abcam Cambridge, MA, USA) was performed on formalin-fixed samples that were processed, embedded in paraffin blocks and sectioned at 5 μm. Heat-induced antigen retrieval followed by ABC detection (Vectastain ABC kit, Vector Laboratories, Inc., Burlingame, CA, USA) was performed using the protocols described previously [
11]. Peroxidase activity was revealed using 3.3-diaminobenzidine (Sigma-Aldrich, St. Louis, MO, USA) as a substrate. Slides were counterstained with hematoxylin and visualized using an Olympus microscope (BX43, Olympus, Tokyo, Japan).
Measurements of percentage of the adipocyte area in the skin, and diameter and size of intradermal adipocytes were performed using ImageJ software version v1.52a (National Institutes of Health, Bethesda, MD, USA) (number of mice n = 5 per group).
4.3. Lipid Extraction
Lipid extraction according to the Folch method with modifications described previously [
38,
39] was performed on uninjured skin collected from young, old, fed LFD or HFD mice. Briefly, tissues were weighed and homogenized with a 2:1 mixture of chloroform–methanol (4 mL of mixture per 0.05 g of tissue) (
n = 4 per group). After homogenization, mixtures were centrifuged at 15,000×
g for 10 min and supernatants were transferred to the new tubes with addition of 0.8 mL of distilled water, vortexed and centrifuged for 15 min (2500×
g). Upper phases were removed and lower phases with lipid content were evaporated under a nitrogen stream at 37 °C for 35 min. Percentage of lipid content was calculated by dividing weight of lipids after evaporation by weight of tissue and multiplying by 100.
4.4. Ex Vivo DF Migration Assay
Skin explants collected postmortem from young or old B6 mice fed LFD or HFD were cut into small (1–2 mm
2) samples, seeded in 60-mm Petri dishes (10 samples per plate) and cultured in DMEM/F-12 medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 15% of fetal bovine serum (FBS; Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA) and antibiotics (Penicillin/Streptomycin, Sigma-Aldrich, St. Louis, MO, USA). Every second day media was removed and replaced with a fresh equivalent volume. Dermal fibroblast migration was photo documented daily and evaluated by the DF area of outgrowth from skin explant using ImageJ (National Institutes of Health, Bethesda, MD, USA) similar to the previously described method for keratinocytes migration [
40].
4.5. Flow Cytometry Analysis
EMSC and DFs isolated from young or old B6 mice fed LFD or HFD (
n = 3–5) were characterized by expression of cell surface markers using flow cytometry analysis. Briefly, cells (
p = 1) seeded in a 60-mm culture dish were cultured in DMEM/F12 culture medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 15% fetal bovine serum (FBS, Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA) with antibiotics (Penicilin/Streptomycin, Sigma-Aldrich, St. Louis, MO, USA). Confluent cells were washed with PBS, trypsinized and counted (Countess, Life Technology, Thermo Fisher Scientific, Waltham, MA, USA). For the analysis, approximately 0.6–1.5 × 10
6 cells from each set of cells were incubated for 30 min with fluorochrome-labeled antibodies to specific surface antigens or their isotype controls (
Supplementary Table S7) and fixed with 1% PFA for 20 min. Cells were analyzed the same day using a BD LSRFortessa cell analyzer flow cytometer and BD FACSDiva TM v6.2 software (Becton Dickinson, Franklin Lakes, NJ, USA).
4.6. Cell Isolation and Culture
Skin samples were mechanically processed and digested in collagenase type 1 (3.68 mg/mL; Sigma-Aldrich, St. Louis, MO, USA) for 80 min at 37 °C in an incubator shaker. Isolates were filtered with a cell strainer with a pore diameter of 100 μm and centrifuged for 5 min at 1300 rpm at room temperature. Cells were plated in 60- or 100-mm Petri dishes in DMEM/F-12 medium (Sigma-Aldrich, St. Louis, MO, USA) with 15% fetal bovine serum (FBS, Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA) and gentamicin/amphotericin solution (Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA). Confluent fibroblasts were trypsinized (0.05%; Sigma-Aldrich, St. Louis, MO, USA) and frozen for further experiments. Ear mesenchymal stem cells (EMSC) and stromal vascular fraction (SVF) of inguinal fat depot were isolated and cultured as described before [
23].
Skin tissues isolated from newborn B6 mice were incubated overnight in dispase solution (6 U/mL; Life Technologies) at 4 °C to separate epidermis from dermis. Keratinocytes were isolated from separated epidermis by a series of trypsin digestions as previously described [
41]. Cells were counted and seeded in cell culture inserts with a 0.4-μm pore size membrane (0.5 × 10
6 per insert). Keratinocytes were seeded in DMEM/F-12 medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% FBS (Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA), 0.2% Primocin (InvivoGen, San Diego, CA, USA) and 120 μM β-mercaptoethanol (Sigma-Aldrich, St. Louis, MO, USA). After 24 h of culture, seeding medium was switched to keratinocyte-specific medium (CellnTec, Bern, Switzerland).
4.7. Keratinocyte Transduction with Adenoviruses
Keratinocyte transduction was performed accordingly to a previously described protocol [
41]. Briefly, keratinocytes were seeded in inserts (Corning
® BioCoat™ Control Inserts with a 0.4-µm PET Membrane; Corning, Tewksbury, MA, USA) were transduced with Foxn1-GFP-expressing (Ad-Foxn1) or control (Ad-GFP) adenoviruses at 70% of confluency. After 4 h, 1 mL of CnT Basal Medium with supplements A, B, C (CELLnTEC, Bern, Switzerland) were added per each insert and coculture of keratinocytes-DFs or keratinocytes-EMSC models were set up (day 0). The keratinocytes medium was changed every 2 days of culture.
4.8. DF/SVF/EMSC Adipogenic Differentiation
Adipogenic differentiation was performed as previously described [
42]. Cells (DFs, SVFs or EMSC) were seeded in a 6-well plate and maintained in DMEM/F12 supplemented with 15% FBS and penicillin/streptomycin (Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA), until confluence (day 0). The adipogenic differentiation process in DFs, SVFs, or EMSC was induced by changing media to Adipogenic Medium I containing DMEM/F12, 5% FBS, 1% Penicillin/Streptomycin, 0.5 mM isobutylmethylxanthine (IBMX, Sigma-Aldrich, St. Louis, MO, USA), 1.7 μM insulin (Sigma-Aldrich, St. Louis, MO, USA) and 1 μM dexamethasone (Sigma-Aldrich, St. Louis, MO, USA). At day 2 of differentiation, the medium was replaced by Adipogenic Medium II containing DMEM/F12 supplemented with 5% FBS, 1% Penicillin/Streptomycin, 17 nM insulin and 5 μM rosiglitazone (Sigma-Aldrich, St. Louis, MO, USA). Cells (DFs/EMSC/keratinocytes) were harvested at indicated time points: day 0, 2, 4, 7 for RNA isolation.
4.9. Oil Red O Staining
DFs or SVF cells were washed with PBS, fixed in 10% buffered formalin for 1 h and stained with a 60% solution of Oil Red O (Sigma-Aldrich, St. Louis, MO, USA) for 10 min as described previously [
43]. Cells were washed five times with water and visualized under a microscope (IX51, Olympus, Tokyo, Japan) equipped with an Olympus digital camera (XC50, Olympus, Tokyo, Japan). Lipid accumulation was measured by extracting Oil Red O from dye-retaining cells with isopropanol, and determining absorbance at 500 nm.
4.10. Cell Metabolic Activity Measurement and Viability Assay
DFs isolated from young LFD animals were cultured in monolayer condition or in coculture with keratinocytes nontransduced or transduced with Ad-Foxn1 or Ad-GFP (n = 5 per condition).
DF cell metabolic activity was measured using MTT colorimetric method as described previously [
43]. Briefly, after 24 h and 48 h, sterile MTT solution (3-[4,5-dimethylthiazol- 2-yl]-2,5-diphenyltetrazolium bromide; Sigma-Aldrich, St. Louis, MO, USA) was added to each well and incubated for 4 h. Next, the medium was removed and formazan crystals were dissolved in DMSO for 1 h. Absorbance was measured at 570 nm using a microplate reader (ASYS Hitech GmbH, UVM340, Biogenet; Biochrom Ltd., Cambridge, UK) and MicroWin 2000 software version V4.0 for Windows XP (Siemens, Monachium, Germany).
For the viability assay after 24 h of culture, cells were trypsinized, counted using the trypan blue exclusion method and resuspended in Annexin Binding Buffer (Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA) with addition of Annexin V Alexa Fluor 350 Conjugate (Life Technologies) and propidium iodide (Life Technologies). Samples were analyzed using flow cytometry with BD LSRFortessa and BD FACSDiva TM v6.2 software (Becton Dickinson, Franklin Lakes, NJ, USA).
4.11. RNA Isolation and Quantitative Real-Time Reverse Transcriptase PCR
Real-time RT-qPCR was performed as described previously [
11]. Total RNA was isolated using the TRIzol Reagent (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA). RNA (500 ng) was reverse transcribed with the High-Capacity cDNA Reverse Transcription Kit with RNase Inhibitor (Applied Biosystems, Thermo Fisher Scientific) according to the manufacturer’s instructions. The mRNA levels of adipogenic-related genes were measured with Single Tube TaqMan Gene Expression Assays (Life Technologies, Thermo Fisher Scientific) (
Supplementary Table S8). The levels of gene expression were quantified relative to the level of
Hprt1.
4.12. Protein Isolation and Western Blot Analysis
Cell lysate collected from nontransduced, transduced with Ad-Foxn1 or Ad-GFP keratinocytes were prepared in 200 μL RIPA buffer containing protease inhibitor cocktail (PhosStop, Roche; Protease Inhibitor, Sigma-Aldrich, St. Louis, MO, USA). Thirty milligrams of protein per sample separated on 12% SDS/PAGE gels and transferred onto polyvinylidene difluoride membranes (Merck Millipore, Burlington, MA, USA) were incubated with anti-BMP2 (1:1000, ab14933, Abcam, Cambridge, MA, USA), anti-IGF2 (1:800, GTX 129110, GeneTex, Irvine, CA, USA), anti-GAPDH (1:1500, ab 9484, Abcam, Cambridge, MA, USA) and anti-Actb (1:1000, ab 8226, Abcam, Cambridge, MA, USA) antibodies followed by fluorescent secondary antibodies (Alexa Fluor 680, 1:10,000, goat anti-mouse, A10038, Life Technologies, Thermo Fisher Scientific and Irdye 800, 1:10,000, 611-132-122, Rockland, Limerick, PA, USA). Bands were visualized using the ChemiDoc™ Imaging System (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Densitometric analysis was performed using Image Lab™ Software version 6.0 (Bio-Rad Laboratories, Inc.) (n = 3 samples per experimental group).
4.13. Statistical Analysis
Linear models were used in order to analyze the impact of different variables on parameters, in which interaction between these covariates was taken into account as well. Based on such models, lsmeans were calculated (least-squared means—means calculated based on model’s coefficients) and compared. For each model, the adjusted Rsquared is provided as the measure of goodness-of-fit. The significance level was set to 0.05. Calculations were performed in R (ver. 3.5.3) using packages: emmeans (ver. 1.4.1) and tidyverse (ver. 1.2.1).
Statistical analysis of DF flow cytometric characteristics, EMSC and DF adipogenic differentiation was performed with GraphPad Prism, Version 6.0 (GraphPad Software, La Jolla, CA, USA). The data were checked for normality using the Shapiro–Wilk test. One-way analysis of variance with posthoc Tukey’s test or two-way analysis of variance were used. Data are expressed as mean ± standard deviation (SD). A value of p < 0.05 was considered statistically significant.

 ,
, .png)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





