Effect of Acute Sprint Exercise on Myokines and Food Intake Hormones in Young Healthy Men

 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Results
2.1. Exercise Performance
2.2. Appetite Perceptions
2.3. Energy Intake (EI)
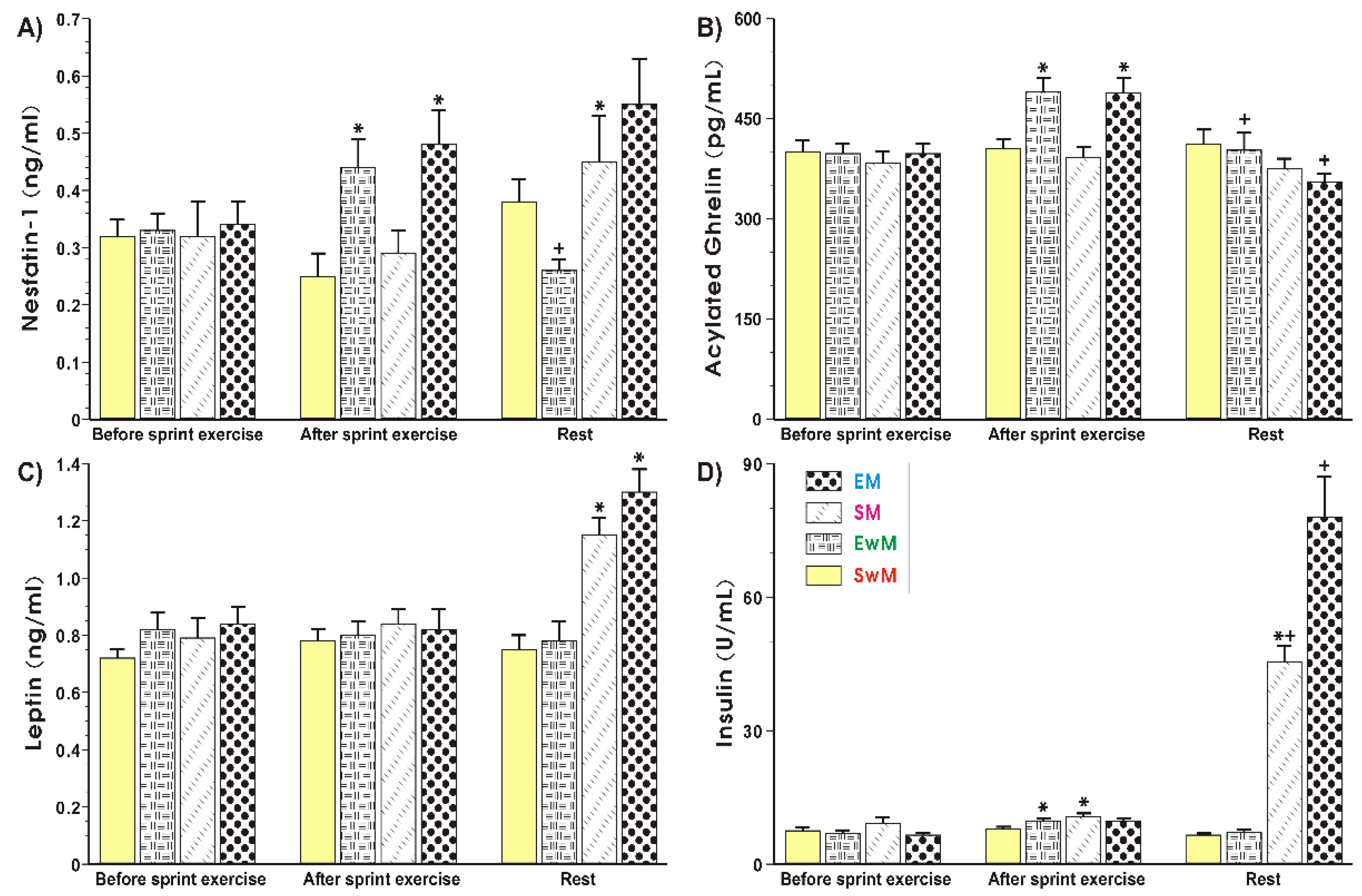
2.4. Effect of Exercise on Plasma Hormones Concentrations
3. Discussion
4. Materials and Methods
4.1. Study Participants
4.2. Study Design and Experimental Protocol
4.3. Exercise with the W-T30
4.4. Visual Analog Scales and Food Intake
4.5. Borg’s Rating of Perceived Exertion Scale (6–20)
4.6. Blood Collection and Biochemical Measurements
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [PubMed]
- Wharton, S.; Lau, D.C.W.; Vallis, M.; Sharma, A.M.; Biertho, L.; Campbell-Scherer, D.; Adamo, K.; Alberga, A.; Bell, R.; Boule, N.; et al. Obesity in adults: A clinical practice guideline. CMAJ 2020, 192, E875–E891. [Google Scholar] [CrossRef] [PubMed]
- Valassi, E.; Scacchi, M.; Cavagnini, F. Neuroendocrine control of food intake. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.M.; Desbrow, B.; Sabapathy, S.; Leveritt, M. Acute exercise and subsequent energy intake. A meta-analysis. Appetite 2013, 63, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Muller, T.D.; Nogueiras, R.; Andermann, M.L.; Andrews, Z.B.; Anker, S.D.; Argente, J.; Batterham, R.L.; Benoit, S.C.; Bowers, C.Y.; Broglio, F.; et al. Ghrelin. Mol. Metab. 2015, 4, 437–460. [Google Scholar] [CrossRef]
- Schubert, M.M.; Sabapathy, S.; Leveritt, M.; Desbrow, B. Acute exercise and hormones related to appetite regulation: A meta-analysis. Sports Med. 2014, 44, 387–403. [Google Scholar] [CrossRef]
- Murphy, K.G.; Dhillo, W.S.; Bloom, S.R. Gut peptides in the regulation of food intake and energy homeostasis. Endocr. Rev. 2006, 27, 719–727. [Google Scholar] [CrossRef]
- Casey, A.; Constantin-Teodosiu, D.; Howell, S.; Hultman, E.; Greenhaff, P.L. Metabolic response of type I and II muscle fibers during repeated bouts of maximal exercise in humans. Am. J. Physiol. 1996, 271, E38–E43. [Google Scholar] [CrossRef]
- Grassi, B.; Rossiter, H.B.; Zoladz, J.A. Skeletal muscle fatigue and decreased efficiency: Two sides of the same coin? Exerc. Sport Sci. Rev. 2015, 43, 75–83. [Google Scholar] [CrossRef]
- Zoladz, J.A.; Szkutnik, Z.; Grassi, B. Metabolic Transitions and Muscle Metabolic Stability: Effects of Exercise Training. In Muscle and Exercise Physiology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 391–422. [Google Scholar]
- Burgomaster, K.A.; Howarth, K.R.; Phillips, S.M.; Rakobowchuk, M.; Macdonald, M.J.; McGee, S.L.; Gibala, M.J. Similar metabolic adaptations during exercise after low volume sprint interval and traditional endurance training in humans. J. Physiol. 2008, 586, 151–160. [Google Scholar] [CrossRef]
- Little, J.P.; Safdar, A.; Bishop, D.; Tarnopolsky, M.A.; Gibala, M.J. An acute bout of high-intensity interval training increases the nuclear abundance of PGC-1alpha and activates mitochondrial biogenesis in human skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1303–R1310. [Google Scholar] [CrossRef] [PubMed]
- Deighton, K.; Barry, R.; Connon, C.E.; Stensel, D.J. Appetite, gut hormone and energy intake responses to low volume sprint interval and traditional endurance exercise. Eur. J. Appl. Physiol. 2013, 113, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Tache, Y. Minireview: Nesfatin-1—An emerging new player in the brain-gut, endocrine, and metabolic axis. Endocrinology 2011, 152, 4033–4038. [Google Scholar] [CrossRef] [PubMed]
- Weibert, E.; Stengel, A. The X/A-like cell revisited-spotlight on the peripheral effects of NUCB2/nesfatin-1 and ghrelin. J. Physiol. Pharmacol. 2017, 68, 497–520. [Google Scholar] [PubMed]
- Senin, L.L.; Al-Massadi, O.; Barja-Fernandez, S.; Folgueira, C.; Castelao, C.; Tovar, S.A.; Leis, R.; Lago, F.; Baltar, J.; Baamonde, I.; et al. Regulation of NUCB2/nesfatin-1 production in rat’s stomach and adipose tissue is dependent on age, testosterone levels and lactating status. Mol. Cell. Endocrinol. 2015, 411, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Algul, S.; Ozdenk, C.; Ozcelik, O. Variations in leptin, nesfatin-1 and irisin levels induced by aerobic exercise in young trained and untrained male subjects. Biol. Sport 2017, 34, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Ahmadizad, S.; Avansar, A.S.; Ebrahim, K.; Avandi, M.; Ghasemikaram, M. The effects of short-term high-intensity interval training vs. moderate-intensity continuous training on plasma levels of nesfatin-1 and inflammatory markers. Horm. Mol. Biol. Clin. Investig. 2015, 21, 165–173. [Google Scholar] [CrossRef]
- Ghanbari-Niaki, A.; Kraemer, R.R.; Soltani, R. Plasma nesfatin-1 and glucoregulatory hormone responses to two different anaerobic exercise sessions. Eur. J. Appl. Physiol. 2010, 110, 863–868. [Google Scholar] [CrossRef]
- Mohebbi, H.; Nourshahi, M.; Ghasemikaram, M.; Safarimosavi, S. Effects of exercise at individual anaerobic threshold and maximal fat oxidation intensities on plasma levels of nesfatin-1 and metabolic health biomarkers. J. Physiol. Biochem. 2015, 71, 79–88. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Febbraio, M.A. Muscles, exercise and obesity: Skeletal muscle as a secretory organ. Nat. Rev. Endocrinol. 2012, 8, 457–465. [Google Scholar] [CrossRef]
- Legård, G.E.; Pedersen, B.K. Muscle as an Endocrine Organ. In Muscle and Exercise Physiology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 285–307. [Google Scholar]
- Bilski, J.; Mazur-Bialy, A.I.; Wierdak, M.; Brzozowski, T. The impact of physical activity and nutrition on inflammatory bowel disease: The potential role of cross talk between adipose tissue and skeletal muscle. J. Physiol. Pharmacol. 2013, 64, 143–155. [Google Scholar] [PubMed]
- Bilski, J.; Mazur-Bialy, A.I.; Brzozowski, B.; Magierowski, M.; Jasnos, K.; Krzysiek-Maczka, G.; Urbanczyk, K.; Ptak-Belowska, A.; Zwolinska-Wcislo, M.; Mach, T.; et al. Moderate Exercise Training Attenuates the Severity of Experimental Rodent Colitis: The Importance of Crosstalk between Adipose Tissue and Skeletal Muscles. Mediators Inflamm. 2015, 2015, 605071. [Google Scholar] [CrossRef] [PubMed]
- Almada, C.; Cataldo, L.R.; Smalley, S.V.; Diaz, E.; Serrano, A.; Hodgson, M.I.; Santos, J.L. Plasma levels of interleukin-6 and interleukin-18 after an acute physical exercise: Relation with post-exercise energy intake in twins. J. Physiol. Biochem. 2013, 69, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Islam, H.; Townsend, L.K.; McKie, G.L.; Medeiros, P.J.; Gurd, B.J.; Hazell, T.J. Potential involvement of lactate and interleukin-6 in the appetite-regulatory hormonal response to an acute exercise bout. J. Appl. Physiol. (1985) 2017, 123, 614–623. [Google Scholar] [CrossRef]
- Butt, Z.D.; Hackett, J.D.; Volkoff, H. Irisin in goldfish (Carassius auratus): Effects of irisin injections on feeding behavior and expression of appetite regulators, uncoupling proteins and lipoprotein lipase, and fasting-induced changes in FNDC5 expression. Peptides 2017, 90, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Kohler, R.M.; Rundell, K.W.; Evans, T.M.; Levine, A.M. Peak power during repeated wingate trials: Implications for testing. J. Strength Cond. Res. 2010, 24, 370–374. [Google Scholar] [PubMed]
- Alis, R.; Risso-Ballester, J.; Sanchis-Gomar, F.; Primo-Carrau, C.; Lozano-Calve, S.; Dipalo, M.; Aloe, R.; Blesa, J.-R.; Romagnoli, M.; Lippi, G. Hemoconcentration Induced By Exercise. Revisiting The Dill & Costill Equation: 3273 Board# 34 May 30, 930 AM-1100 AM. Med. Sci. Sports Exerc. 2015, 47, 874. [Google Scholar]
- Zoladz, J.A.; Rademaker, A.C.; Sargeant, A.J. Human muscle power generating capability during cycling at different pedalling rates. Exp. Physiol. 2000, 85, 117–124. [Google Scholar]
- Sargeant, A.J. Structural and functional determinants of human muscle power. Exp. Physiol. 2007, 92, 323–331. [Google Scholar] [CrossRef]
- Tsofliou, F.; Pitsiladis, Y.P.; Malkova, D.; Wallace, A.M.; Lean, M.E. Moderate physical activity permits acute coupling between serum leptin and appetite-satiety measures in obese women. Int. J. Obes. Relat. Metab. Disord. 2003, 27, 1332–1339. [Google Scholar]
- Larson-Meyer, D.E.; Palm, S.; Bansal, A.; Austin, K.J.; Hart, A.M.; Alexander, B.M. Influence of running and walking on hormonal regulators of appetite in women. J. Obes. 2012, 2012, 730409. [Google Scholar] [CrossRef] [PubMed]
- Date, Y.; Murakami, N.; Toshinai, K.; Matsukura, S.; Niijima, A.; Matsuo, H.; Kangawa, K.; Nakazato, M. The role of the gastric afferent vagal nerve in ghrelin-induced feeding and growth hormone secretion in rats. Gastroenterology 2002, 123, 1120–1128. [Google Scholar] [PubMed]
- Chen, H.Y.; Trumbauer, M.E.; Chen, A.S.; Weingarth, D.T.; Adams, J.R.; Frazier, E.G.; Shen, Z.; Marsh, D.J.; Feighner, S.D.; Guan, X.M.; et al. Orexigenic action of peripheral ghrelin is mediated by neuropeptide Y and agouti-related protein. Endocrinology 2004, 145, 2607–2612. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.J.; Fullick, S.; Gregson, W.; Clarke, N.; Doran, D.; MacLaren, D.; Atkinson, G. Paradoxical post-exercise responses of acylated ghrelin and leptin during a simulated night shift. Chronobiol. Int. 2010, 27, 590–605. [Google Scholar] [CrossRef]
- Hagobian, T.A.; Yamashiro, M.; Hinkel-Lipsker, J.; Streder, K.; Evero, N.; Hackney, T. Effects of acute exercise on appetite hormones and ad libitum energy intake in men and women. Appl. Physiol. Nutr. Metab. 2013, 38, 66–72. [Google Scholar] [CrossRef]
- Unick, J.L.; Otto, A.D.; Goodpaster, B.H.; Helsel, D.L.; Pellegrini, C.A.; Jakicic, J.M. Acute effect of walking on energy intake in overweight/obese women. Appetite 2010, 55, 413–419. [Google Scholar] [CrossRef]
- Broom, D.R.; Miyashita, M.; Wasse, L.K.; Pulsford, R.; King, J.A.; Thackray, A.E.; Stensel, D.J. Acute effect of exercise intensity and duration on acylated ghrelin and hunger in men. J. Endocrinol. 2017, 232, 411–422. [Google Scholar] [CrossRef]
- Mani, B.K.; Castorena, C.M.; Osborne-Lawrence, S.; Vijayaraghavan, P.; Metzger, N.P.; Elmquist, J.K.; Zigman, J.M. Ghrelin mediates exercise endurance and the feeding response post-exercise. Mol. Metab. 2018, 9, 114–130. [Google Scholar] [CrossRef]
- Wasse, L.K.; Sunderland, C.; King, J.A.; Miyashita, M.; Stensel, D.J. The influence of vigorous running and cycling exercise on hunger perceptions and plasma acylated ghrelin concentrations in lean young men. Appl. Physiol. Nutr. Metab. 2013, 38, 1–6. [Google Scholar] [CrossRef]
- Erdmann, J.; Tahbaz, R.; Lippl, F.; Wagenpfeil, S.; Schusdziarra, V. Plasma ghrelin levels during exercise-effects of intensity and duration. Regul. Pept. 2007, 143, 127–135. [Google Scholar] [CrossRef]
- Nass, R.; Gaylinn, B.D.; Thorner, M.O. The role of ghrelin in GH secretion and GH disorders. Mol. Cell. Endocrinol. 2011, 340, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Stokes, K.A.; Sykes, D.; Gilbert, K.L.; Chen, J.W.; Frystyk, J. Brief, high intensity exercise alters serum ghrelin and growth hormone concentrations but not IGF-I, IGF-II or IGF-I bioactivity. Growth Horm. IGF Res. 2010, 20, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, A.; Inui, A.; Yuzuriha, H.; Ueno, N.; Katsuura, G.; Fujimiya, M.; Fujino, M.A.; Niijima, A.; Meguid, M.M.; Kasuga, M. Characterization of the effects of pancreatic polypeptide in the regulation of energy balance. Gastroenterology 2003, 124, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Oh, I.S.; Hashimoto, K.; Nakata, M.; Yamamoto, S.; Yoshida, N.; Eguchi, H.; Kato, I.; Inoue, K.; Satoh, T.; et al. Peripheral administration of nesfatin-1 reduces food intake in mice: The leptin-independent mechanism. Endocrinology 2009, 150, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Pal, M.; Febbraio, M.A.; Whitham, M. From cytokine to myokine: The emerging role of interleukin-6 in metabolic regulation. Immunol. Cell Biol. 2014, 92, 331–339. [Google Scholar] [CrossRef]
- Schele, E.; Benrick, A.; Grahnemo, L.; Egecioglu, E.; Anesten, F.; Palsdottir, V.; Jansson, J.O. Inter-relation between interleukin (IL)-1, IL-6 and body fat regulating circuits of the hypothalamic arcuate nucleus. J. Neuroendocrinol. 2013, 25, 580–589. [Google Scholar] [CrossRef]
- Hazell, T.J.; Islam, H.; Townsend, L.K.; Schmale, M.S.; Copeland, J.L. Effects of exercise intensity on plasma concentrations of appetite-regulating hormones: Potential mechanisms. Appetite 2016, 98, 80–88. [Google Scholar] [CrossRef]
- Huh, J.Y.; Panagiotou, G.; Mougios, V.; Brinkoetter, M.; Vamvini, M.T.; Schneider, B.E.; Mantzoros, C.S. FNDC5 and irisin in humans: I. Predictors of circulating concentrations in serum and plasma and II. mRNA expression and circulating concentrations in response to weight loss and exercise. Metabolism 2012, 61, 1725–1738. [Google Scholar] [CrossRef]
- Schultes, B.; Schmid, S.M.; Wilms, B.; Jauch-Chara, K.; Oltmanns, K.M.; Hallschmid, M. Lactate infusion during euglycemia but not hypoglycemia reduces subsequent food intake in healthy men. Appetite 2012, 58, 818–821. [Google Scholar] [CrossRef]
- Sutton, J.R.; Young, J.D.; Lazarus, L.; Hickie, J.B.; Maksvytis, J. The hormonal response to physical exercise. Australas. Ann. Med. 1969, 18, 84–90. [Google Scholar] [CrossRef]
- Fox, J.; Rioux, B.; Goulet, E.; Johanssen, N.; Swift, D.; Bouchard, D.; Loewen, H.; Sénéchal, M. Effect of an acute exercise bout on immediate post-exercise irisin concentration in adults: A meta-analysis. Scand. J. Med. Sci. Sports 2018, 28, 16–28. [Google Scholar] [PubMed]
- Blundell, J.; de Graaf, C.; Hulshof, T.; Jebb, S.; Livingstone, B.; Lluch, A.; Mela, D.; Salah, S.; Schuring, E.; van der Knaap, H.; et al. Appetite control: Methodological aspects of the evaluation of foods. Obes. Rev. 2010, 11, 251–270. [Google Scholar] [CrossRef] [PubMed]
- Bilski, J.; Jaworek, J.; Pokorski, J.; Nitecki, J.; Nitecka, E.; Pokorska, J.; Mazur-Bialy, A.; Szklarczyk, J. Effects of time of day and the Wingate test on appetite perceptions, food intake and plasma levels of adipokines. J. Physiol. Pharmacol. 2016, 67, 667–676. [Google Scholar]
- Borg, G.A. Perceived exertion: A note on “history” and methods. Med. Sci. Sports 1973, 5, 90–93. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | MPO5-s (W·kg·BM−1) | MPO30-s (W·kg·BM−1) | Plasma Lactate (mmol·L−1) | HR (bt·min−1) | RPE (6–20) |
|---|---|---|---|---|---|
| Rest | – | – | 1.07 ± 0.04 | 71.9 ± 9.1 | – |
| Exercise | 10.84 ± 0.95 | 8.1 ± 0.67 | 12.4 ± 0.09 * | 179.5 ± 15.8 * | 17 ± 1.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilski, J.; Mazur-Bialy, A.I.; Surmiak, M.; Hubalewska-Mazgaj, M.; Pokorski, J.; Nitecki, J.; Nitecka, E.; Pokorska, J.; Targosz, A.; Ptak-Belowska, A.; et al. Effect of Acute Sprint Exercise on Myokines and Food Intake Hormones in Young Healthy Men. Int. J. Mol. Sci. 2020, 21, 8848. https://doi.org/10.3390/ijms21228848
Bilski J, Mazur-Bialy AI, Surmiak M, Hubalewska-Mazgaj M, Pokorski J, Nitecki J, Nitecka E, Pokorska J, Targosz A, Ptak-Belowska A, et al. Effect of Acute Sprint Exercise on Myokines and Food Intake Hormones in Young Healthy Men. International Journal of Molecular Sciences. 2020; 21(22):8848. https://doi.org/10.3390/ijms21228848
Chicago/Turabian StyleBilski, Jan, Agnieszka Irena Mazur-Bialy, Marcin Surmiak, Magdalena Hubalewska-Mazgaj, Janusz Pokorski, Jacek Nitecki, Ewa Nitecka, Joanna Pokorska, Aneta Targosz, Agata Ptak-Belowska, and et al. 2020. "Effect of Acute Sprint Exercise on Myokines and Food Intake Hormones in Young Healthy Men" International Journal of Molecular Sciences 21, no. 22: 8848. https://doi.org/10.3390/ijms21228848
APA StyleBilski, J., Mazur-Bialy, A. I., Surmiak, M., Hubalewska-Mazgaj, M., Pokorski, J., Nitecki, J., Nitecka, E., Pokorska, J., Targosz, A., Ptak-Belowska, A., A. Zoladz, J., & Brzozowski, T. (2020). Effect of Acute Sprint Exercise on Myokines and Food Intake Hormones in Young Healthy Men. International Journal of Molecular Sciences, 21(22), 8848. https://doi.org/10.3390/ijms21228848