The Potential Role of Hepatocyte Growth Factor in Degenerative Disorders of the Synovial Joint and Spine

 ,
,
Abstract
:1. Introduction
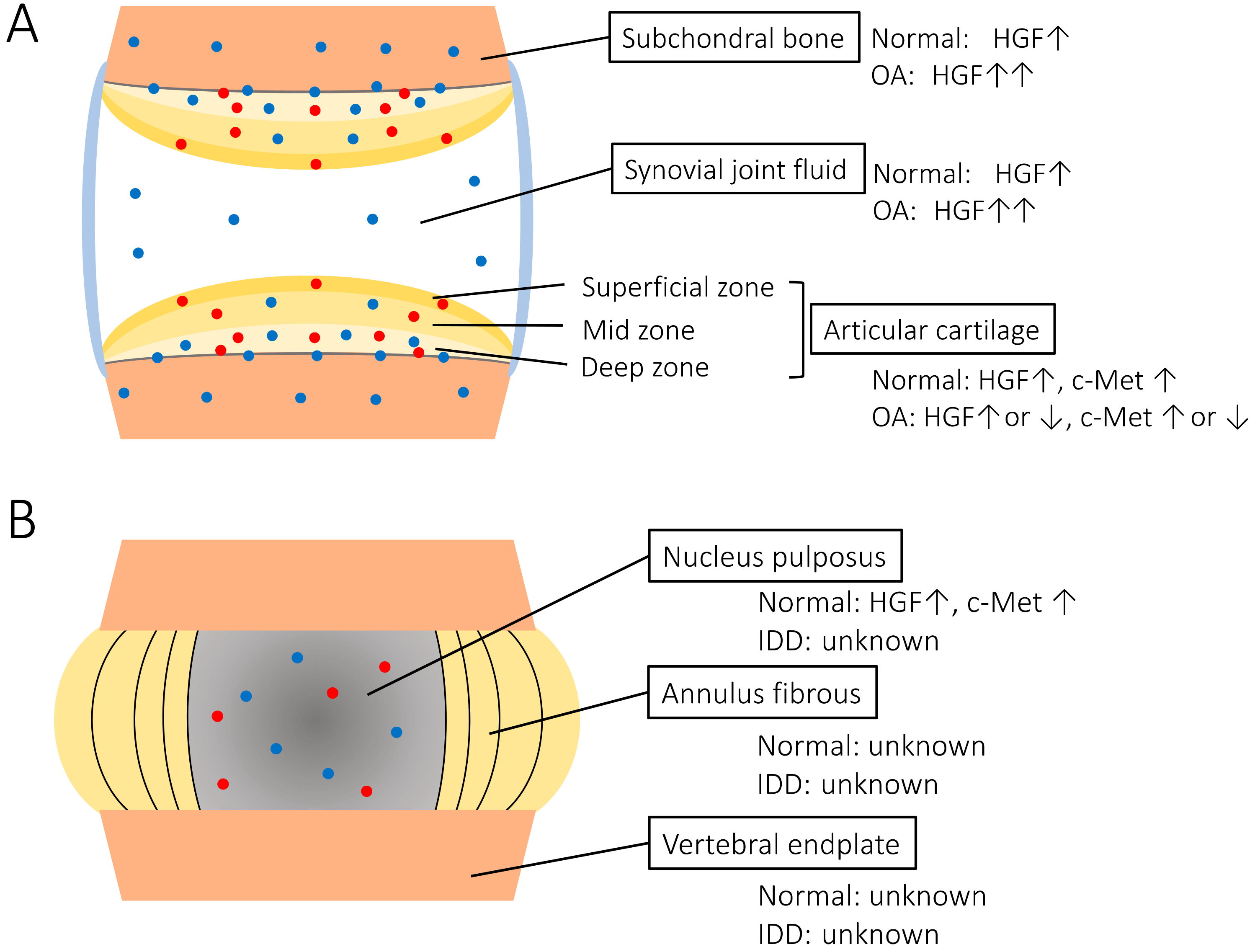
2. HGF/c-Met Expression in the Synovial Joint and Spine
2.1. Synovial Joint
2.1.1. Articular Cartilage and Synovial Joint Fluid
2.1.2. Subchondral Bone
2.2. Spine
Intervertebral Disc
3. HGF/c-Met Activities in the Synovial Joint and Spine
3.1. Articular Cartilage Proliferation and ECM Metabolism
3.2. Bony Remodeling and Osteogenesis in the Articular Joint
3.3. Intervertebral Disc Proliferation and ECM Metabolism
3.4. Neurotrophic Effect of Degenerative Spinal Disorder-Related Pain
4. Recent Advances in Clinical Applications for Degenerative Disorder of the Synovial Joint and Spine
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AF | Annulus fibrosus |
| BMP | Bone morphogenic protein |
| ECM | Extracellular matrix |
| HGF | Hepatocyte growth factor |
| HIF-1α | Hypoxia-inducible factor-1α |
| IDD | Intervertebral disc degeneration |
| MMP | Matrix metalloprotease |
| NP | Nucleus pulposus |
| OA | Osteoarthritis |
| TNF | Tumor necrosis factor |
References
- Neogi, T. The epidemiology and impact of pain in osteoarthritis. Osteoarthritis Cartilage 2013, 21, 1145–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, K.M.; Karppinen, J.; Chan, D.; Ho, D.W.; Song, Y.Q.; Sham, P.; Cheah, K.S.; Leong, J.C.; Luk, K.D. Prevalence and pattern of lumbar magnetic resonance imaging changes in a population study of one thousand forty-three individuals. Spine 2009, 34, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Sakai, D. Future perspectives of cell-based therapy for intervertebral disc disease. Eur. Spine J. 2008, 17 (Suppl. 4), 452–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hustedt, J.W.; Goltzer, O.; Bohl, D.D.; Fraser, J.F.; Lara, N.J.; Spangehl, M.J. Calculating the Cost and Risk of Comorbidities in Total Joint Arthroplasty in the United States. J. Arthroplasty 2017, 32, 355.e1–361.e1. [Google Scholar] [CrossRef]
- Deyo, R.A.; Mirza, S.K.; Martin, B.I.; Kreuter, W.; Goodman, D.C.; Jarvik, J.G. Trends, major medical complications, and charges associated with surgery for lumbar spinal stenosis in older adults. JAMA 2010, 303, 1259–1265. [Google Scholar] [CrossRef] [Green Version]
- Goldring, M.B.; Goldring, S.R. Osteoarthritis. J. Cell Physiol. 2007, 213, 626–634. [Google Scholar] [CrossRef]
- Miller, J.A.; Schmatz, C.; Schultz, A.B. Lumbar disc degeneration: Correlation with age, sex, and spine level in 600 autopsy specimens. Spine 1988, 13, 173–178. [Google Scholar] [CrossRef]
- Kirkwood, T.B. Understanding the odd science of aging. Cell 2005, 120, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Hasty, P.; Campisi, J.; Hoeijmakers, J.; van Steeg, H.; Vijg, J. Aging and genome maintenance: Lessons from the mouse? Science 2003, 299, 1355–1359. [Google Scholar] [CrossRef]
- Guarente, L. Mitochondria—A nexus for aging, calorie restriction, and sirtuins? Cell 2008, 132, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Nawa, K.; Ichihara, A. Partial purification and characterization of hepatocyte growth factor from serum of hepatectomized rats. Biochem. Biophys. Res. Commun. 1984, 122, 1450–1459. [Google Scholar] [CrossRef]
- Russell, W.E.; McGowan, J.A.; Bucher, N.L. Partial characterization of a hepatocyte growth factor from rat platelets. J. Cell Physiol. 1984, 119, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Sakai, K.; Nakamura, T.; Matsumoto, K. Hepatocyte growth factor twenty years on: Much more than a growth factor. J. Gastroenterol. Hepatol. 2011, 26 (Suppl. 1), 188–202. [Google Scholar] [CrossRef] [Green Version]
- Bottaro, D.P.; Rubin, J.S.; Faletto, D.L.; Chan, A.M.; Kmiecik, T.E.; Vande Woude, G.F.; Aaronson, S.A. Identification of the hepatocyte growth factor receptor as the c-met proto-oncogene product. Science 1991, 251, 802–804. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Funakoshi, H.; Takahashi, H.; Sakai, K. HGF-Met Pathway in Regeneration and Drug Discovery. Biomedicines 2014, 2, 275–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grumbles, R.M.; Howell, D.S.; Wenger, L.; Altman, R.D.; Howard, G.A.; Roos, B.A. Hepatocyte growth factor and its actions in growth plate chondrocytes. Bone 1996, 19, 255–261. [Google Scholar] [CrossRef]
- Pfander, D.; Cramer, T.; Weseloh, G.; Pullig, O.; Schuppan, D.; Bauer, M.; Swoboda, B. Hepatocyte growth factor in human osteoarthritic cartilage. Osteoarthritis Cartilage 1999, 7, 548–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feuerherm, A.J.; Borset, M.; Seidel, C.; Sundan, A.; Leistad, L.; Ostensen, M.; Faxvaag, A. Elevated levels of osteoprotegerin (OPG) and hepatocyte growth factor (HGF) in rheumatoid arthritis. Scand. J. Rheumatol. 2001, 30, 229–234. [Google Scholar]
- Bau, B.; McKenna, L.A.; Soeder, S.; Fan, Z.; Pecht, A.; Aigner, T. Hepatocyte growth factor/scatter factor is not a potent regulator of anabolic and catabolic gene expression in adult human articular chondrocytes. Biochem. Biophys. Res. Commun. 2004, 316, 984–990. [Google Scholar] [CrossRef]
- Koch, A.E.; Halloran, M.M.; Hosaka, S.; Shah, M.R.; Haskell, C.J.; Baker, S.K.; Panos, R.J.; Haines, G.K.; Bennett, G.L.; Pope, R.M.; et al. Hepatocyte growth factor. A cytokine mediating endothelial migration in inflammatory arthritis. Arthritis Rheum. 1996, 39, 1566–1575. [Google Scholar] [CrossRef]
- Nagashima, M.; Hasegawa, J.; Kato, K.; Yamazaki, J.; Nishigai, K.; Ishiwata, T.; Asano, G.; Yoshino, S. Hepatocyte growth factor (HGF), HGF activator, and c-Met in synovial tissues in rheumatoid arthritis and osteoarthritis. J. Rheumatol. 2001, 28, 1772–1778. [Google Scholar] [PubMed]
- Yukioka, K.; Inaba, M.; Furumitsu, Y.; Yukioka, M.; Nishino, T.; Goto, H.; Nishizawa, Y.; Morii, H. Levels of hepatocyte growth factor in synovial fluid and serum of patients with rheumatoid arthritis and release of hepatocyte growth factor by rheumatoid synovial fluid cells. J. Rheumatol. 1994, 21, 2184–2189. [Google Scholar] [PubMed]
- Dankbar, B.; Neugebauer, K.; Wunrau, C.; Tibesku, C.O.; Skwara, A.; Pap, T.; Fuchs-Winkelmann, S. Hepatocyte growth factor induction of macrophage chemoattractant protein-1 and osteophyte-inducing factors in osteoarthritis. J. Orthop. Res. 2007, 25, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Guevremont, M.; Martel-Pelletier, J.; Massicotte, F.; Tardif, G.; Pelletier, J.P.; Ranger, P.; Lajeunesse, D.; Reboul, P. Human adult chondrocytes express hepatocyte growth factor (HGF) isoforms but not HgF: Potential implication of osteoblasts on the presence of HGF in cartilage. J. Bone Miner. Res. 2003, 18, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Abed, E.; Bouvard, B.; Martineau, X.; Jouzeau, J.Y.; Reboul, P.; Lajeunesse, D. Elevated hepatocyte growth factor levels in osteoarthritis osteoblasts contribute to their altered response to bone morphogenetic protein-2 and reduced mineralization capacity. Bone 2015, 75, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Grano, M.; Galimi, F.; Zambonin, G.; Colucci, S.; Cottone, E.; Zallone, A.Z.; Comoglio, P.M. Hepatocyte growth factor is a coupling factor for osteoclasts and osteoblasts in vitro. Proc. Natl. Acad. Sci. USA 1996, 93, 7644–7648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisch, R.N.; Curtis, K.M.; Aenlle, K.K.; Howard, G.A. Hepatocyte growth factor and alternative splice variants—Expression, regulation and implications in osteogenesis and bone health and repair. Expert Opin. Ther. Targets 2016, 20, 1087–1098. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, I.M.; Vresilovic, E.J.; Risbud, M.V. Is the spinal motion segment a diarthrodial polyaxial joint: What a nice nucleus like you doing in a joint like this? Bone 2012, 50, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Humzah, M.D.; Soames, R.W. Human intervertebral disc: Structure and function. Anat. Rec. 1988, 220, 337–356. [Google Scholar] [CrossRef]
- Ishibashi, H.; Tonomura, H.; Ikeda, T.; Nagae, M.; Sakata, M.; Fujiwara, H.; Tanida, T.; Mastuda, K.; Kawata, M.; Kubo, T. Hepatocyte growth factor/c-met promotes proliferation, suppresses apoptosis, and improves matrix metabolism in rabbit nucleus pulposus cells in vitro. J. Orthop. Res. 2016, 34, 709–716. [Google Scholar] [CrossRef] [Green Version]
- Takebayashi, T.; Iwamoto, M.; Jikko, A.; Matsumura, T.; Enomoto-Iwamoto, M.; Myoukai, F.; Koyama, E.; Yamaai, T.; Matsumoto, K.; Nakamura, T.; et al. Hepatocyte growth factor/scatter factor modulates cell motility, proliferation, and proteoglycan synthesis of chondrocytes. J. Cell Biol. 1995, 129, 1411–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakitani, S.; Imoto, K.; Kimura, T.; Ochi, T.; Matsumoto, K.; Nakamura, T. Hepatocyte growth factor facilitates cartilage repair. Full thickness articular cartilage defect studied in rabbit knees. Acta Orthop. Scand. 1997, 68, 474–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tibesku, C.O.; Daniilidis, K.; Szuwart, T.; Jahn, U.R.; Schlegel, P.M.; Fuchs-Winkelmann, S. Influence of hepatocyte growth factor on autologous osteochondral transplants in an animal model. Arch. Orthop. Trauma Surg. 2011, 131, 1145–1151. [Google Scholar] [CrossRef] [PubMed]
- Reboul, P.; Pelletier, J.P.; Tardif, G.; Benderdour, M.; Ranger, P.; Bottaro, D.P.; Martel-Pelletier, J. Hepatocyte growth factor induction of collagenase 3 production in human osteoarthritic cartilage: Involvement of the stress-activated protein kinase/c-Jun N-terminal kinase pathway and a sensitive p38 mitogen-activated protein kinase inhibitor cascade. Arthritis Rheum. 2001, 44, 73–84. [Google Scholar] [CrossRef]
- von der Mark, K.; Kirsch, T.; Nerlich, A.; Kuss, A.; Weseloh, G.; Gluckert, K.; Stoss, H. Type X collagen synthesis in human osteoarthritic cartilage. Indication of chondrocyte hypertrophy. Arthritis Rheum. 1992, 35, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Terai, H.; Nomura-Furuwatari, C.; Mizuno, S.; Matsumoto, K.; Nakamura, T.; Takaoka, K. Hepatocyte growth factor contributes to fracture repair by upregulating the expression of BMP receptors. J. Bone Miner. Res. 2005, 20, 1723–1730. [Google Scholar] [CrossRef]
- Burr, D.B.; Radin, E.L. Microfractures and microcracks in subchondral bone: Are they relevant to osteoarthrosis? Rheum. Dis. Clin. N. Am. 2003, 29, 675–685. [Google Scholar] [CrossRef]
- Itsuji, T.; Tonomura, H.; Ishibashi, H.; Mikami, Y.; Nagae, M.; Takatori, R.; Tanida, T.; Matsuda, K.I.; Tanaka, M.; Kubo, T. Hepatocyte growth factor regulates HIF-1alpha-induced nucleus pulposus cell proliferation through MAPK-, PI3K/Akt-, and STAT3-mediated signaling. J. Orthop. Res. 2020. [Google Scholar] [CrossRef]
- Zhang, F.; Zhao, X.; Shen, H.; Zhang, C. Molecular mechanisms of cell death in intervertebral disc degeneration (Review). Int. J. Mol. Med. 2016, 37, 1439–1448. [Google Scholar] [CrossRef] [Green Version]
- Shamji, M.F.; Setton, L.A.; Jarvis, W.; So, S.; Chen, J.; Jing, L.; Bullock, R.; Isaacs, R.E.; Brown, C.; Richardson, W.J. Proinflammatory cytokine expression profile in degenerated and herniated human intervertebral disc tissues. Arthritis Rheum. 2010, 62, 1974–1982. [Google Scholar]
- Zou, F.; Jiang, J.; Lu, F.; Ma, X.; Xia, X.; Wang, L.; Wang, H. Efficacy of intradiscal hepatocyte growth factor injection for the treatment of intervertebral disc degeneration. Mol. Med. Rep. 2013, 8, 118–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzer, A.C.; Aprill, C.N.; Derby, R.; Fortin, J.; Kine, G.; Bogduk, N. The prevalence and clinical features of internal disc disruption in patients with chronic low back pain. Spine 1995, 20, 1878–1883. [Google Scholar] [CrossRef]
- Manchikanti, L.; Manchikanti, K.N.; Cash, K.A.; Singh, V.; Giordano, J. Age-related prevalence of facet-joint involvement in chronic neck and low back pain. Pain Physician 2008, 11, 67–75. [Google Scholar] [PubMed]
- Maigne, J.Y.; Aivaliklis, A.; Pfefer, F. Results of sacroiliac joint double block and value of sacroiliac pain provocation tests in 54 patients with low back pain. Spine 1996, 21, 1889–1892. [Google Scholar] [CrossRef] [PubMed]
- Panjabi, M.M. A hypothesis of chronic back pain: Ligament subfailure injuries lead to muscle control dysfunction. Eur. Spine J. 2006, 15, 668–676. [Google Scholar] [CrossRef] [Green Version]
- Malik, K.M.; Cohen, S.P.; Walega, D.R.; Benzon, H.T. Diagnostic criteria and treatment of discogenic pain: A systematic review of recent clinical literature. Spine J. 2013, 13, 1675–1689. [Google Scholar] [CrossRef]
- Ohtori, S.; Miyagi, M.; Inoue, G. Sensory nerve ingrowth, cytokines, and instability of discogenic low back pain: A review. Spine Surg. Relat. Res. 2018, 2, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Fujii, K.; Yamazaki, M.; Kang, J.D.; Risbud, M.V.; Cho, S.K.; Qureshi, S.A.; Hecht, A.C.; Iatridis, J.C. Discogenic Back Pain: Literature Review of Definition, Diagnosis, and Treatment. JBMR Plus 2019, 3, e10180. [Google Scholar] [CrossRef] [Green Version]
- Kirkaldy-Willis, W.H.; Wedge, J.H.; Yong-Hing, K.; Reilly, J. Pathology and pathogenesis of lumbar spondylosis and stenosis. Spine 1978, 3, 319–328. [Google Scholar] [CrossRef]
- Weber, K.T.; Satoh, S.; Alipui, D.O.; Virojanapa, J.; Levine, M.; Sison, C.; Quraishi, S.; Bloom, O.; Chahine, N.O. Exploratory study for identifying systemic biomarkers that correlate with pain response in patients with intervertebral disc disorders. Immunol. Res. 2015, 63, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Karamouzian, S.; Eskandary, H.; Faramarzee, M.; Saba, M.; Safizade, H.; Ghadipasha, M.; Malekpoor, A.R.; Ohadi, A. Frequency of lumbar intervertebral disc calcification and angiogenesis, and their correlation with clinical, surgical, and magnetic resonance imaging findings. Spine 2010, 35, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, N.; Yamanaka, H.; Fukuoka, T.; Dai, Y.; Obata, K.; Mashimo, T.; Noguchi, K. Expression of HGF and cMet in the peripheral nervous system of adult rats following sciatic nerve injury. Neuroreport 2001, 12, 1403–1407. [Google Scholar] [CrossRef]
- Ko, K.R.; Lee, J.; Lee, D.; Nho, B.; Kim, S. Hepatocyte Growth Factor (HGF) Promotes Peripheral Nerve Regeneration by Activating Repair Schwann Cells. Sci. Rep. 2018, 8, 8316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, F.; Hilton, M.; Davies, A.M. Cooperation between HGF and CNTF in promoting the survival and growth of sensory and parasympathetic neurons. Mol. Cell Neurosci. 2000, 15, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Kuniyoshi, K.; Iwakura, N.; Matsuura, Y.; Suzuki, T.; Takahashi, K.; Ohtori, S. Vein wrapping for chronic nerve constriction injury in a rat model: Study showing increases in VEGF and HGF production and prevention of pain-associated behaviors and nerve damage. J. Bone Joint Surg. Am. 2014, 96, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Ajroud-Driss, S.; Christiansen, M.; Allen, J.A.; Kessler, J.A. Phase 1/2 open-label dose-escalation study of plasmid DNA expressing two isoforms of hepatocyte growth factor in patients with painful diabetic peripheral neuropathy. Mol. Ther. 2013, 21, 1279–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brucker, P.U.; Izzo, N.J.; Chu, C.R. Tonic activation of hypoxia-inducible factor 1alpha in avascular articular cartilage and implications for metabolic homeostasis. Arthritis Rheum. 2005, 52, 3181–3191. [Google Scholar] [CrossRef]
- Risbud, M.V.; Schipani, E.; Shapiro, I.M. Hypoxic regulation of nucleus pulposus cell survival: From niche to notch. Am. J. Pathol. 2010, 176, 1577–1583. [Google Scholar] [CrossRef]
- Fernandez-Torres, J.; Zamudio-Cuevas, Y.; Martinez-Nava, G.A.; Lopez-Reyes, A.G. Hypoxia-Inducible Factors (HIFs) in the articular cartilage: A systematic review. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2800–2810. [Google Scholar]
- Chaudhary, P.; Babu, G.S.; Sobti, R.C.; Gupta, S.K. HGF regulate HTR-8/SVneo trophoblastic cells migration/invasion under hypoxic conditions through increased HIF-1alpha expression via MAPK and PI3K pathways. J. Cell Commun. Signal. 2019, 13, 503–521. [Google Scholar] [CrossRef]
- Yu, F.; Lin, Y.; Zhan, T.; Chen, L.; Guo, S. HGF expression induced by HIF-1alpha promote the proliferation and tube formation of endothelial progenitor cells. Cell Biol. Int. 2015, 39, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Ashinsky, B.G.; Bonnevie, E.D.; Mandalapu, S.A.; Pickup, S.; Wang, C.; Han, L.; Mauck, R.L.; Smith, H.E.; Gullbrand, S.E. Intervertebral Disc Degeneration Is Associated With Aberrant Endplate Remodeling and Reduced Small Molecule Transport. J. Bone Miner. Res. 2020, 35, 1572–1581. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.R.; Yoo, J.J.; Kim, H.A. Therapeutics in Osteoarthritis Based on an Understanding of Its Molecular Pathogenesis. Int. J. Mol. Sci. 2018, 19, 674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, K.; Chen, S.; Li, Z.; Deng, X.; Huang, D.; Xiong, L.; Shao, Z. Mechanisms of endogenous repair failure during intervertebral disc degeneration. Osteoarthritis Cartilage 2019, 27, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Rider, S.M.; Mizuno, S.; Kang, J.D. Molecular Mechanisms of Intervertebral Disc Degeneration. Spine Surg. Relat. Res. 2019, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Study | Cell Type and Animal Model | Effects |
|---|---|---|
| Takebayashi et al. (1995) | In vitro Rabbit, rat-chondrocytes | Increased cell proliferation Promotion of proteoglycan synthesis |
| Wakitani et al. (1997) | In vivo Rabbit-full thickness defect | Repaired with closely resembling hyaline cartilage |
| Reboul et al. (2001) | In vitro Human chondrocytes | Increased both the expression and synthesis of collagenase 3 |
| Bau et al. (2004) | In vitro Human chondrocytes | No change in proteoglycan expression and synthesis No influence in the expression of collagenase and aggrecan degrading enzyme |
| Tibesku et al. (2011) | In vivo Sheep-osteochondral transplantation | Increased cellularity of the transplanted graft Not diminished the fissures in the marginal zone of the graft |
| Study | Cell Type and Animal Model | Effects |
|---|---|---|
| Zou et al. (2013) | In vivo Rat-needle puncture model | Increase in NP water content Increased type II collagen staining in the NP Increased BMP-2-positive cells in the AF |
| Ishibashi et al. (2016) | In vitro Rabbit-NP and AF cells | Increased cell proliferation Inhibition of apoptosis induced by reactive oxygen species Suppression of MMP expression stimulated by TNF-α |
| Itsuji et al. (2020) | In vitro Rabbit-NP cells | Increased cell proliferation under hypoxia Upregulation of HIF-1α |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tonomura, H.; Nagae, M.; Takatori, R.; Ishibashi, H.; Itsuji, T.; Takahashi, K. The Potential Role of Hepatocyte Growth Factor in Degenerative Disorders of the Synovial Joint and Spine. Int. J. Mol. Sci. 2020, 21, 8717. https://doi.org/10.3390/ijms21228717
Tonomura H, Nagae M, Takatori R, Ishibashi H, Itsuji T, Takahashi K. The Potential Role of Hepatocyte Growth Factor in Degenerative Disorders of the Synovial Joint and Spine. International Journal of Molecular Sciences. 2020; 21(22):8717. https://doi.org/10.3390/ijms21228717
Chicago/Turabian StyleTonomura, Hitoshi, Masateru Nagae, Ryota Takatori, Hidenobu Ishibashi, Tomonori Itsuji, and Kenji Takahashi. 2020. "The Potential Role of Hepatocyte Growth Factor in Degenerative Disorders of the Synovial Joint and Spine" International Journal of Molecular Sciences 21, no. 22: 8717. https://doi.org/10.3390/ijms21228717
APA StyleTonomura, H., Nagae, M., Takatori, R., Ishibashi, H., Itsuji, T., & Takahashi, K. (2020). The Potential Role of Hepatocyte Growth Factor in Degenerative Disorders of the Synovial Joint and Spine. International Journal of Molecular Sciences, 21(22), 8717. https://doi.org/10.3390/ijms21228717