Can Gut Microbiota Affect Dry Eye Syndrome?


Abstract
1. Introduction
2. Innate and Adaptive Immune Education by Gut Microbiota and Homeostatic Crosstalk between Microbiota and Host
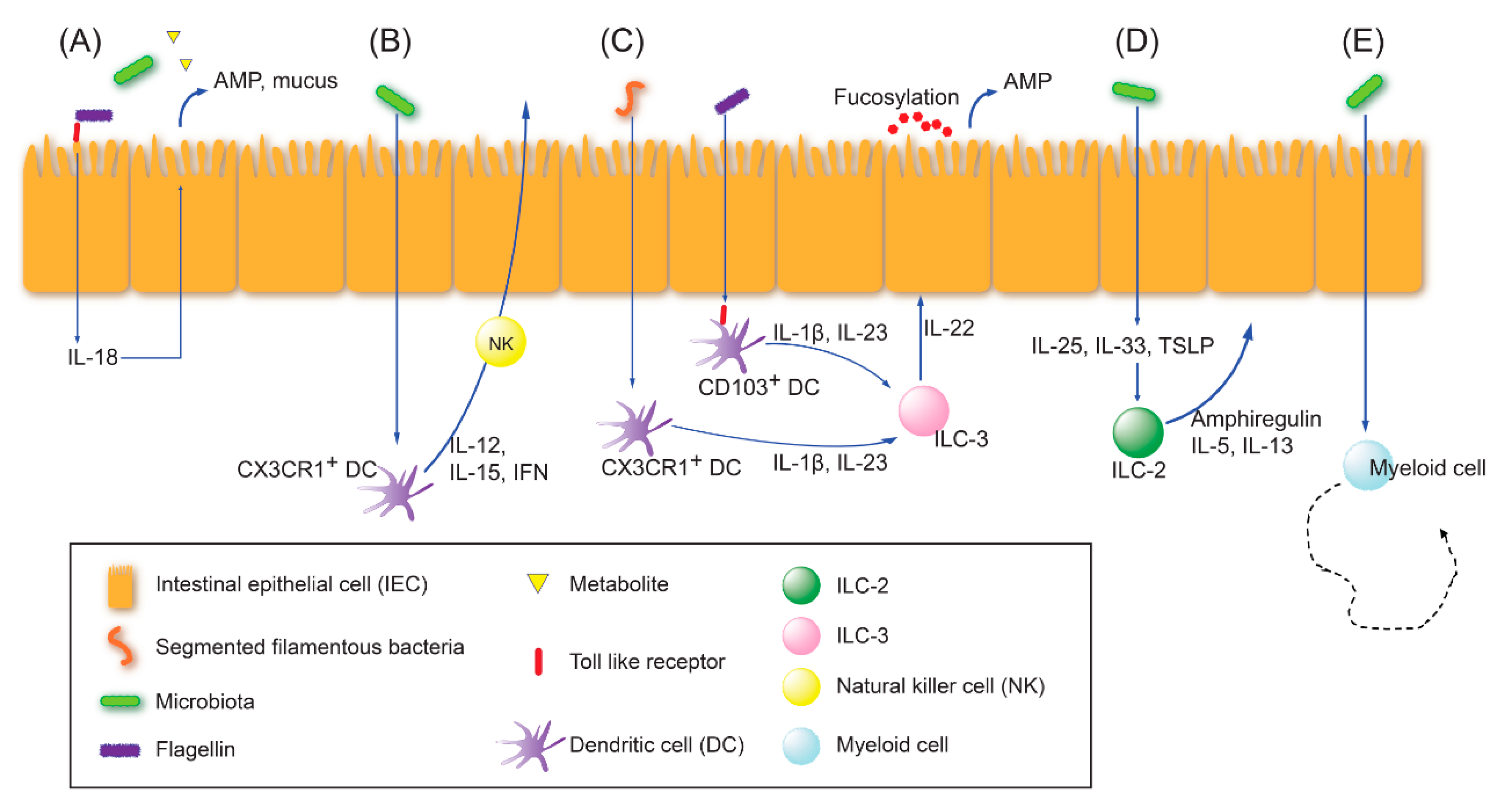
2.1. Innate Immune Education
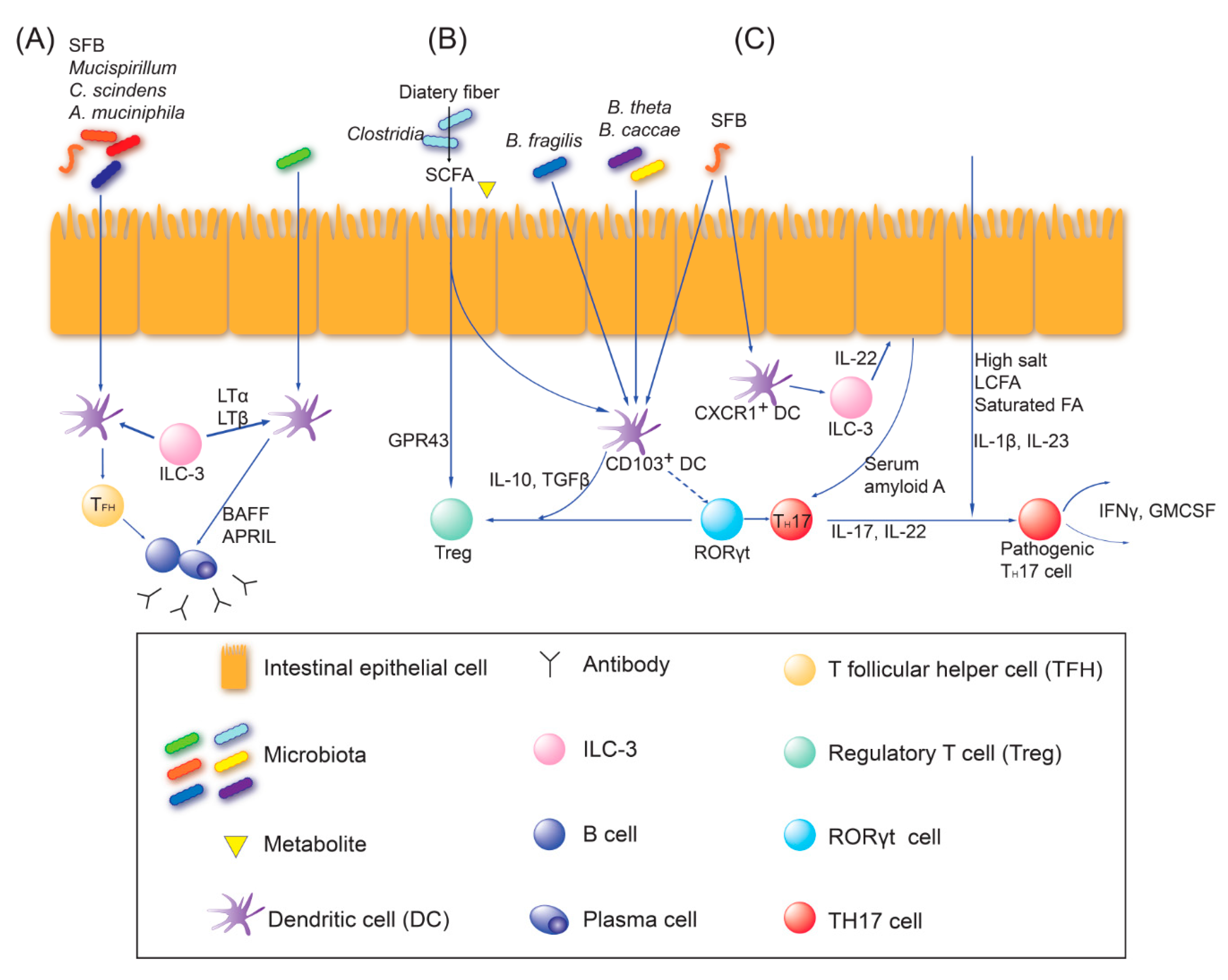
2.2. Adaptive Immune Education
3. Current Knowledge of the Gut Dysbiosis Associated with Non-SS Autoimmune Disease in Human Studies Detected by Metagenomic Sequencing Methods
3.1. Gut Dysbiosis Affects Connective Tissue Disease
3.2. Gut Dysbiosis Affects the Central Nervous System
4. Gut Microbiota–Immune Axis Contributes to the Pathogenesis of Non-Dry Eye Ocular Autoimmune Disease
4.1. Uveitis
4.2. Age-Related Macular Disease (AMD)
4.3. Primary Open Angle Glaucoma (POAG)
4.4. Neuromyelitis Optica Spectrum Disorders (NMOSDs)
4.5. Inflammation of Lacrimal Gland
5. Pathogenesis of Non-Sjögren or Sjögren Syndrome-Related Dry Eye
5.1. Non-Sjögren Dry Eye
5.2. Sjögren Syndrome (SS)-Related Dry Eye
6. Dry Eye Syndrome and Gut Microbiota
6.1. Evidence from Animal Studies
6.2. Evidence from Clinical Studies
6.3. Gut Microbiota Comparison of Dry Eye and Sjögren’s Syndrome
7. Dry Eye and Probiotics
7.1. Prebiotics and Probiotics
7.2. Effects Seen in Animal Studies
7.3. Effects Seen in Clinical Studies
8. The Hypothesis of Gut Dysbiosis–Ocular Surface–Lacrimal Gland Axis Communications
9. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADDE | Aqueous deficient dry eye |
| AMD | Age related macular disease |
| APC | Antigen-presenting cell |
| AQP | Aquaporin |
| DES | Dry eye syndrome |
| EDE | Evaporative dry eye |
| GM-CSF | Granulocyte–macrophage colony-stimulating factor |
| HLA | Human leukocyte antigen |
| IFN | Interferon |
| Ig | Immunoglobulin |
| IL | Interleukin |
| ILC | Innate lymphoid cell |
| KO | Knock-out |
| LCFA | Long-chain fatty acid |
| LG | Lacrimal gland |
| MGD | Meibomian gland dysfunction |
| MMP | Matrix metalloproteinase |
| NK | Natural killer |
| NMOSD | Neuromyelitis optica spectrum disorders |
| NOD | NOD.B10.H2b |
| pDC | Plasmacytoid dendritic cell |
| POAG | Primary open angle glaucoma |
| PRR | Pattern recognition receptor |
| pSS | Primary Sjögren’s syndrome |
| RA | Rheumatoid arthritis |
| SFB | Segmented filamentous bacteria |
| SFCA | Short-chain fatty acid |
| SLE | Systemic lupus erythematosus |
| SS | Sjögren’s syndrome |
| Teff | Effector T cells |
| TFH | T follicular helper |
| TGF | Transforming growth factor |
| TH1 | T helper 1 |
| TH17 | T helper 17 |
| TLR | Toll-like receptor |
| TNF | Tumor necrosis factor |
| Treg | Regulatory T cells |
| VEGF | Vascular endothelial growth factor |
| VKH | Vogt-Koyanagi-Harada |
References
- Gill, S.R.; Pop, M.; DeBoy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.; Garges, S.; Giovanni, M.; McInnes, P.; Wang, L.; Schloss, J.A.; Bonazzi, V.; McEwen, J.E.; Wetterstrand, K.A.; Deal, C.; et al. The NIH Human Microbiome Project. Genome Res. 2009, 19, 2317–2323. [Google Scholar] [PubMed]
- Integrative, H.M.P.; Proctor, L.M.; Creasy, H.H.; Fettweis, J.M.; Lloyd-Price, J.; Mahurkar, A.; Zhou, W.; Buck, G.A.; Snyder, M.P.; Strauss, J.F., III; et al. The Integrative Human Microbiome Project. Nat. Cell Biol. 2019, 569, 641–648. [Google Scholar]
- Kahrstrom, C.T.; Pariente, N.; Weiss, U. Intestinal microbiota in health and disease. Nature 2016, 535, 47. [Google Scholar] [CrossRef]
- Lee, Y.K.; Mazmanian, S.K. Has the microbiota played a critical role in the evolution of the adaptive immune system? Science 2010, 330, 1768–1773. [Google Scholar] [CrossRef]
- Berer, K.; Mues, M.; Koutrolos, M.; Al Rasbi, Z.; Boziki, M.; Johner, C.; Wekerle, H.; Krishnamoorthy, G. Commensal microbiota and myelin autoantigen cooperate to trigger autoimmune demyelination. Nature 2011, 479, 538–541. [Google Scholar] [CrossRef]
- Fleischmann, R.D.; Adams, M.D.; White, O.; Clayton, A.R.; Kirkness, E.F.; Kerlavage, A.R.; Bult, C.J.; Tomb, J.F.; Dougherty, B.A.; Merrick, J.M.; et al. Whole-genome random sequencing and assembly of Haemophilus influenzae Rd. Science 1995, 269, 496–512. [Google Scholar] [CrossRef]
- Relman, D.A. Microbial Genomics and Infectious Diseases. N. Engl. J. Med. 2011, 365, 347–357. [Google Scholar] [CrossRef]
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef]
- Segre, J.A. Microbial growth dynamics and human disease. Science 2015, 349, 1058–1059. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Both, T.; Dalm, V.A.; Van Hagen, P.M.; van Daele, P.L. Reviewing primary Sjögren’s syndrome: Beyond the dryness—From pathophysiology to diagnosis and treatment. Int. J. Med. Sci. 2017, 14, 191–200. [Google Scholar] [CrossRef]
- Kuklinski, E.; Asbell, P.A. Sjogren’s syndrome from the perspective of ophthalmology. Clin. Immunol. 2017, 182, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.; Downie, L.E.; Korb, D.; Benitez-Del-Castillo, J.M.; Dana, R.; Deng, S.X.; Dong, P.N.; Geerling, G.; Hida, R.Y.; Liu, Y.; et al. TFOS DEWS II Management and Therapy Report. Ocul. Surf. 2017, 15, 575–628. [Google Scholar] [CrossRef] [PubMed]
- Craig, J.P.; Nelson, J.D.; Azar, D.T.; Belmonte, C.; Bron, A.; Chauhan, S.K.; De Paiva, C.S.; Gomes, J.A.; Hammitt, K.M.; Jones, L.W.; et al. TFOS DEWS II Report Executive Summary. Ocul. Surf. 2017, 15, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Horai, R.; Caspi, R.R. Microbiome and Autoimmune Uveitis. Front. Immunol. 2019, 10, 232. [Google Scholar] [CrossRef]
- Pascal, V.; Pozuelo, M.; Borruel, N.; Casellas, F.; Campos, D.; Santiago, A.; Martinez, X.; Varela, E.; Sarrabayrouse, G.; Machiels, K.; et al. A microbial signature for Crohn’s disease. Gut 2017, 66, 813–822. [Google Scholar] [CrossRef]
- Zárate-Bladés, C.R.; Horai, R.; Mattapallil, M.J.; Ajami, N.J.; Wong, M.; Petrosino, J.F.; Itoh, K.; Chan, C.-C.; Caspi, R.R. Gut microbiota as a source of a surrogate antigen that triggers autoimmunity in an immune privileged site. Gut Microbes 2017, 8, 59–66. [Google Scholar] [CrossRef]
- Silverman, G.J. The microbiome in SLE pathogenesis. Nat. Rev. Rheumatol. 2019, 15, 72–74. [Google Scholar] [CrossRef]
- Li, Z.; Zhu, H.; Zhang, L.; Qin, C. The intestinal microbiome and Alzheimer’s disease: A review. Anim. Model Exp. Med. 2018, 1, 180–188. [Google Scholar] [CrossRef]
- Opazo, M.C.; Ortega-Rocha, E.M.; Coronado-Arrázola, I.; Bonifaz, L.C.; Boudin, H.; Neunlist, M.; Bueno, S.M.; Kalergis, A.M.; Riedel, C.A. Intestinal Microbiota Influences Non-intestinal Related Autoimmune Diseases. Front. Microbiol. 2018, 9, 432. [Google Scholar] [CrossRef] [PubMed]
- Rujillo-Vargas, C.M.; Schaefer, L.; Alam, J.; Pflugfelder, S.C.; Britton, R.A.; De Paiva, C.S. The gut-eye-lacrimal gland-microbiome axis in Sjögren Syndrome. Ocul. Surf. 2020, 18, 335–344. [Google Scholar] [CrossRef]
- Fabbiano, S.; Suárez-Zamorano, N.; Trajkovski, M. Host–Microbiota Mutualism in Metabolic Diseases. Front. Endocrinol. 2017, 8, 267. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Muller, P.A.; Schneeberger, M.; Matheis, F.; Wang, P.; Kerner, Z.; Ilanges, A.; Pellegrino, K.; Del Mármol, J.; Castro, T.B.R.; Furuichi, M.; et al. Microbiota modulate sympathetic neurons via a gut-brain circuit. Nat. Cell Biol. 2020, 583, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Elson, C.O. Adaptive immune education by gut microbiota antigens. Immunology 2018, 154, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nat. Cell Biol. 2016, 535, 65–74. [Google Scholar] [CrossRef]
- Jiao, Y.; Wu, L.; Huntington, N.D.; Zhang, X. Crosstalk Between Gut Microbiota and Innate Immunity and Its Implication in Autoimmune Diseases. Front. Immunol. 2020, 11, 282. [Google Scholar] [CrossRef]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nat. Cell Biol. 2016, 535, 75–84. [Google Scholar] [CrossRef]
- Zheng, Y.; Valdez, P.A.; Danilenko, D.M.; Hu, Y.; Sa, S.M.; Gong, Q.; Abbas, A.R.; Modrusan, Z.; Ghilardi, N.; De Sauvage, F.J.; et al. Interleukin-22 mediates early host defense against attaching and effacing bacterial pathogens. Nat. Med. 2008, 14, 282–289. [Google Scholar] [CrossRef]
- Zhang, D.; Chen, G.; Manwani, D.; Mortha, A.; Xu, C.; Faith, J.J.; Burk, R.D.; Kunisaki, Y.; Jang, J.-E.; Scheiermann, C.; et al. Neutrophil ageing is regulated by the microbiome. Nat. Cell Biol. 2015, 525, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.A.; Siracusa, M.C.; Abt, M.C.; Kim, B.S.; Kobuley, D.; Kubo, M.; Kambayashi, T.; LaRosa, D.F.; Renner, E.D.; Orange, J.S.; et al. Commensal bacteria—Derived signals regulate basophil hematopoiesis and allergic inflammation. Nat. Med. 2012, 18, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhu, L.; Qin, S. Gut Microbiota Modulation on Intestinal Mucosal Adaptive Immunity. J. Immunol. Res. 2019, 2019, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Guzman-Bautista, E.R.; Suzuki, K.; Asami, S.; Fagarasan, S. Bacteria-immune cells dialog and the homeostasis of the systems. Curr. Opin. Immunol. 2020, 66, 82–89. [Google Scholar] [CrossRef] [PubMed]
- van Der Meulen, T.A.; Harmsen, H.J.; Vila, A.V.; Kurilshikov, A.; Liefers, S.C.; Zhernakova, A.; Fu, J.; Wijmenga, C.; Weersma, R.K.; de Leeuw, K.; et al. Shared gut, but distinct oral microbiota composition in primary Sjögren’s syndrome and systemic lupus erythematosus. J. Autoimmun. 2019, 97, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lei, L. Interleukin-35 regulates the balance of Th17 and Treg responses during the pathogenesis of connective tissue diseases. Int. J. Rheum. Dis. 2020, 1–7. [Google Scholar] [CrossRef]
- Jiang, J.; Zhao, M.; Chang, C.; Wu, H.; Lu, Q. Type I Interferons in the Pathogenesis and Treatment of Autoimmune Diseases. Clin. Rev. Allergy Immunol. 2020, 59, 248–272. [Google Scholar] [CrossRef]
- Scher, J.U.; Sczesnak, A.; Longman, R.S.; Segata, N.; Ubeda, C.; Bielski, C.; Rostron, T.; Cerundolo, V.; Pamer, E.G.; Abramson, S.B.; et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. eLife 2013, 2, e01202. [Google Scholar] [CrossRef]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. 2016, 8, 1–14. [Google Scholar] [CrossRef]
- Kim, J.-W.; Kwok, S.-K.; Choe, J.-Y.; Park, S.-H. Recent Advances in Our Understanding of the Link between the Intestinal Microbiota and Systemic Lupus Erythematosus. Int. J. Mol. Sci. 2019, 20, 4871. [Google Scholar] [CrossRef] [PubMed]
- Azzouz, D.; Omarbekova, A.; Heguy, A.; Schwudke, D.; Gisch, N.; Rovin, B.H.; Caricchio, R.; Buyon, J.P.; Alekseyenko, A.V.; Silverman, G.J. Lupus nephritis is linked to disease-activity associated expansions and immunity to a gut commensal. Ann. Rheum. Dis. 2019, 78, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Wang, H.; Xu, S.; Zhuang, Y.; An, J.; Su, C.; Xia, Y.; Chen, J.; Xu, Z.Z.; Liu, Q.; et al. Alteration in gut microbiota is associated with dysregulation of cytokines and glucocorticoid therapy in systemic lupus erythematosus. Gut Microbes 2020, 11, 1758–1773. [Google Scholar] [CrossRef]
- Vaahtovuo, J.; Munukka, E.; Korkeamäki, M.; Luukkainen, R.; Toivanen, P. Fecal microbiota in early rheumatoid arthritis. J. Rheumatol. 2008, 35, 1500–1505. [Google Scholar]
- He, Z.; Shao, T.; Li, H.; Xie, Z.; Wen, C. Alterations of the gut microbiome in Chinese patients with systemic lupus erythematosus. Gut Pathog. 2016, 8, 64. [Google Scholar] [CrossRef]
- Hevia, A.; Milani, C.; López, P.; Cuervo, A.; Arboleya, S.; Duranti, S.; Turroni, F.; González, S.; Suárez, A.; Gueimonde, M.; et al. Intestinal Dysbiosis Associated with Systemic Lupus Erythematosus. mBio 2014, 5, e01548-14. [Google Scholar] [CrossRef]
- Luo, X.M.; Edwards, M.R.; Mu, Q.; Yu, Y.; Vieson, M.D.; Reilly, C.M.; Ahmed, S.A.; Bankole, A.A. Gut Microbiota in Human Systemic Lupus Erythematosus and a Mouse Model of Lupus. Appl. Environ. Microbiol. 2017, 84, e02288-17. [Google Scholar] [CrossRef]
- Zegarra-Ruiz, D.F.; El Beidaq, A.; Iñiguez, A.J.; Di Ricco, M.L.; Vieira, S.M.; Ruff, W.E.; Mubiru, D.; Fine, R.L.; Sterpka, J.; Greiling, T.M.; et al. A Diet-Sensitive Commensal Lactobacillus Strain Mediates TLR7-Dependent Systemic Autoimmunity. Cell Host Microbe 2019, 25, 113–127. [Google Scholar] [CrossRef]
- Rudbane, S.M.A.; Rahmdel, S.; Abdollahzadeh, S.M.; Zare, M.; Bazrafshan, A.; Mazloomi, S.M. The efficacy of probiotic supplementation in rheumatoid arthritis: A meta-analysis of randomized, controlled trials. Inflammopharmacology 2018, 26, 67–76. [Google Scholar] [CrossRef]
- de la Visitación, N.; Robles-Vera, I.; Toral, M.; Duarte, J. Protective Effects of Probiotic Consumption in Cardiovascular Disease in Systemic Lupus Erythematosus. Nutrients 2019, 11, 2676. [Google Scholar] [CrossRef]
- Johnson, K.V.-A.; Foster, K.R. Why does the microbiome affect behaviour? Nat. Rev. Genet. 2018, 16, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.R.; Osadchiy, V.; Kalani, A.; Mayer, E.A. The Brain-Gut-Microbiome Axis. Cell Mol. Gastroenterol. Hepatol. 2018, 6, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Holzer, P.; Farzi, A. Neuropeptides and the Microbiota-Gut-Brain Axis. Adv. Exp. Med. Biol. 2014, 817, 195–219. [Google Scholar] [PubMed]
- Hajjo, H.; Geva-Zatorsky, N. Gut microbiota—Host interactions now also brain-immune axis. Curr. Opin. Neurobiol. 2020, 62, 53–59. [Google Scholar] [CrossRef]
- Barbosa, R.S.D.; Vieira-Coelho, M.A. Probiotics and prebiotics: Focus on psychiatric disorders—A systematic review. Nutr. Rev. 2019, 78, 437–450. [Google Scholar] [CrossRef]
- Gong, H.; Zhang, S.; Li, Q.; Zuo, C.; Gao, X.; Zheng, B.; Lin, M. Gut microbiota compositional profile and serum metabolic phenotype in patients with primary open-angle glaucoma. Exp. Eye Res. 2020, 191, 107921. [Google Scholar] [CrossRef]
- Ye, Z.; Wu, C.; Zhang, N.; Du, L.; Cao, Q.; Huang, X.; Tang, J.; Wang, Q.; Li, F.; Zhou, C.; et al. Altered gut microbiome composition in patients with Vogt-Koyanagi-Harada disease. Gut Microbes 2020, 11, 539–555. [Google Scholar] [CrossRef]
- Rowan, S.; Jiang, S.; Korem, T.; Szymanski, J.; Chang, M.-L.; Szelog, J.; Cassalman, C.; Dasuri, K.; McGuire, C.; Nagai, R.; et al. Involvement of a gut–retina axis in protection against dietary glycemia-induced age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2017, 114, E4472–E4481. [Google Scholar] [CrossRef]
- Horai, R.; Zarateblades, C.R.; Dillenburg-Pilla, P.; Chen, J.; Kielczewski, J.L.; Silver, P.B.; Jittayasothorn, Y.; Chan, C.-C.; Yamane, H.; Honda, K.; et al. Microbiota-Dependent Activation of an Autoreactive T Cell Receptor Provokes Autoimmunity in an Immunologically Privileged Site. Immunity 2015, 43, 343–353. [Google Scholar] [CrossRef]
- Huang, X.; Ye, Z.; Cao, Q.; Su, G.; Wang, Q.; Deng, J.; Zhou, C.; Kijlstra, A.; Yang, P. Gut Microbiota Composition and Fecal Metabolic Phenotype in Patients With Acute Anterior Uveitis. Investig. Opthalmology Vis. Sci. 2018, 59, 1523–1531. [Google Scholar] [CrossRef]
- Ye, Z.; Zhang, N.; Wu, C.; Zhang, X.; Wang, Q.; Huang, X.; Du, L.; Cao, Q.; Tang, J.; Zhou, C.; et al. A metagenomic study of the gut microbiome in Behcet’s disease. Microbiome 2018, 6, 1–13. [Google Scholar] [CrossRef]
- Shimizu, J.; Kubota, T.; Takada, E.; Takai, K.; Fujiwara, N.; Arimitsu, N.; Ueda, Y.; Wakisaka, S.; Suzuki, T.; Suzuki, N. Relative abundance of Megamonas hypermegale and Butyrivibrio species decreased in the intestine and its possible association with the T cell aberration by metabolite alteration in patients with Behcet’s disease (210 characters). Clin. Rheumatol. 2019, 38, 1437–1445. [Google Scholar] [CrossRef] [PubMed]
- Janowitz, C.; Nakamura, Y.K.; Metea, C.; Gligor, A.; Yu, W.; Karstens, L.; Rosenbaum, J.T.; Asquith, M.; Lin, P. Disruption of Intestinal Homeostasis and Intestinal Microbiota During Experimental Autoimmune Uveitis. Investig. Opthalmology Vis. Sci. 2019, 60, 420–429. [Google Scholar] [CrossRef]
- Du, Z.; Wang, Q.; Huang, X.; Yi, S.; Mei, S.; Yuan, G.; Su, G.; Cao, Q.; Zhou, C.; Wang, Y.; et al. Effect of berberine on spleen transcriptome and gut microbiota composition in experimental autoimmune uveitis. Int. Immunopharmacol. 2020, 81, 106270. [Google Scholar] [CrossRef]
- Chakravarthy, S.K.; Jayasudha, R.; Prashanthi, G.S.; Ali, M.H.; Sharma, S.; Tyagi, M.; Shivaji, S. Dysbiosis in the Gut Bacterial Microbiome of Patients with Uveitis, an Inflammatory Disease of the Eye. Indian J. Microbiol. 2018, 58, 457–469. [Google Scholar] [CrossRef]
- Zinkernagel, M.S.; Zysset-Burri, D.C.; Keller, I.; Berger, L.E.; Leichtle, A.B.; Largiadèr, C.R.; Fiedler, G.M.; Wolf, S. Association of the Intestinal Microbiome with the Development of Neovascular Age-Related Macular Degeneration. Sci. Rep. 2017, 7, 40826. [Google Scholar] [CrossRef]
- Shi, Z.; Qiu, Y.; Wang, J.; Fang, Y.; Zhang, Y.; Chen, H.; Du, Q.; Zhao, Z.; Yan, C.; Yang, M.; et al. Dysbiosis of gut microbiota in patients with neuromyelitis optica spectrum disorders: A cross sectional study. J. Neuroimmunol. 2020, 339, 577126. [Google Scholar] [CrossRef]
- Gong, J.; Qiu, W.; Zeng, Q.; Liu, X.; Sun, X.; Li, H.; Yang, Y.; Wu, A.; Bao, J.; Wang, Y.; et al. Lack of short-chain fatty acids and overgrowth of opportunistic pathogens define dysbiosis of neuromyelitis optica spectrum disorders: A Chinese pilot study. Mult. Scler. J. 2018, 25, 1316–1325. [Google Scholar] [CrossRef]
- Cree, B.A.C.; Spencer, C.M.; Varrin-Doyer, M.; Baranzini, S.E.; Zamvil, S.S. Gut microbiome analysis in neuromyelitis optica reveals overabundance of Clostridium perfringens. Ann. Neurol. 2016, 80, 443–447. [Google Scholar] [CrossRef]
- Lin, P.; Bach, M.; Asquith, M.; Lee, A.Y.; Akileswaran, L.; Stauffer, P.; Davin, S.; Pan, Y.; Cambronne, E.D.; Dorris, M.; et al. HLA-B27 and Human beta2-Microglobulin Affect the Gut Microbiota of Transgenic Rats. PLoS ONE 2014, 9, e105684. [Google Scholar]
- Andriessen, E.M.; Wilson, A.M.; Mawambo, G.; Dejda, A.; Miloudi, K.; Sennlaub, F.; Sapieha, P. Gut microbiota influences pathological angiogenesis in obesity-driven choroidal neovascularization. EMBO Mol. Med. 2016, 8, 1366–1379. [Google Scholar] [CrossRef]
- Kugadas, A.; Wright, Q.; Geddes-McAlister, J.; Gadjeva, M. Role of Microbiota in Strengthening Ocular Mucosal Barrier Function Through Secretory IgA. Investig. Opthalmology Vis. Sci. 2017, 58, 4593–4600. [Google Scholar] [CrossRef]
- Kodati, S.; Sen, H.N. Uveitis and the gut microbiota. Best Pract. Res. Clin. Rheumatol. 2019, 33, 101500. [Google Scholar] [CrossRef]
- Horai, R.; Sen, H.N.; Caspi, R.R. Commensal microbiota as a potential trigger of autoimmune uveitis. Expert Rev. Clin. Immunol. 2017, 13, 291–293. [Google Scholar] [CrossRef]
- Nakamura, Y.K.; Metea, C.; Karstens, L.; Asquith, M.; Gruner, H.; Moscibrocki, C.; Lee, I.; Brislawn, C.J.; Jansson, J.K.; Rosenbaum, J.T.; et al. Gut Microbial Alterations Associated With Protection From Autoimmune Uveitis. Investig. Opthalmol. Vis. Sci. 2016, 57, 3747–3758. [Google Scholar] [CrossRef]
- Du, L.; Kijlstra, A.; Yang, P. Vogt-Koyanagi-Harada disease: Novel insights into pathophysiology, diagnosis and treatment. Prog. Retin. Eye Res. 2016, 52, 84–111. [Google Scholar] [CrossRef]
- Liang, L.; Tan, X.; Zhou, Q.; Tian, Y.; Kijlstra, A.; Yang, P. TLR3 and TLR4 But not TLR2 are Involved in Vogt-Koyanagi-Harada Disease by Triggering Proinflammatory Cytokines Production Through Promoting the Production of Mitochondrial Reactive Oxygen Species. Curr. Mol. Med. 2015, 15, 529–542. [Google Scholar] [CrossRef]
- Rinninella, E.; Mele, M.C.; Merendino, N.; Cintoni, M.; Anselmi, G.; Caporossi, A.; Gasbarrini, A.; Minnella, A.M. The Role of Diet, Micronutrients and the Gut Microbiota in Age-Related Macular Degeneration: New Perspectives from the Gut–Retina Axis. Nutrients 2018, 10, 1677. [Google Scholar] [CrossRef]
- Lin, P. Importance of the intestinal microbiota in ocular inflammatory diseases: A review. Clin. Exp. Ophthalmology 2019, 47, 418–422. [Google Scholar] [CrossRef]
- Gill, T.; Asquith, M.; Rosenbaum, J.T.; Colbert, R.A. The intestinal microbiome in spondyloarthritis. Curr. Opin. Rheumatol. 2015, 27, 319–325. [Google Scholar] [CrossRef]
- Mancino, R.; Martucci, A.; Cesareo, M.; Giannini, C.; Corasaniti, M.T.; Bagetta, G.; Nucci, C. Glaucoma and Alzheimer Disease: One Age-Related Neurodegenerative Disease of the Brain. Curr. Neuropharmacol. 2018, 16, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Skrzypecki, J.; Żera, T.; Ufnal, M. Butyrate, a Gut Bacterial Metabolite, Lowers Intraocular Pressure in Normotensive But Not in Hypertensive Rats. J. Glaucoma 2018, 27, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Ratelade, J.; Verkman, A. Neuromyelitis optica: Aquaporin-4 based pathogenesis mechanisms and new therapies. Int. J. Biochem. Cell Biol. 2012, 44, 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zaheer, M.; Bian, F.; Quach, D.; Swennes, A.G.; Britton, R.A.; Pflugfelder, S.C.; De Paiva, C.S. Sjögren-Like Lacrimal Keratoconjunctivitis in Germ-Free Mice. Int. J. Mol. Sci. 2018, 19, 565. [Google Scholar] [CrossRef]
- Yanagisawa, N.; Ueshiba, H.; Abe, Y.; Kato, H.; Higuchi, T.; Yagi, J. Outer Membrane Protein of Gut Commensal Microorganism Induces Autoantibody Production and Extra-Intestinal Gland Inflammation in Mice. Int. J. Mol. Sci. 2018, 19, 3241. [Google Scholar] [CrossRef]
- Choi, S.H.; Oh, J.W.; Ryu, J.S.; Kim, H.M.; Im, S.-H.; Kim, K.P.; Kim, M.K. IRT5 Probiotics Changes Immune Modulatory Protein Expression in the Extraorbital Lacrimal Glands of an Autoimmune Dry Eye Mouse Model. Investig. Opthalmology Vis. Sci. 2020, 61, 42. [Google Scholar] [CrossRef]
- Bron, A.J.; De Paiva, C.S.; Chauhan, S.K.; Bonini, S.; Gabison, E.E.; Jain, S.; Knop, E.; Markoulli, M.; Ogawa, Y.; Perez, V.; et al. TFOS DEWS II pathophysiology report. Ocul. Surf. 2017, 15, 438–510. [Google Scholar] [CrossRef]
- Chauhan, S.K.; Dana, R. Role of Th17 cells in the immunopathogenesis of dry eye disease. Mucosal Immunol. 2009, 2, 375–376. [Google Scholar] [CrossRef]
- de Paiva, C.S.; Chotikavanich, S.; Pangelinan, S.B.; Pitcher, J.D.; Fang, B.; Zheng, X.; Ma, P.; Farley, W.J.; Siemasko, K.F.; Niederkorn, J.Y.; et al. IL-17 disrupts corneal barrier following desiccating stress. Mucosal Immunol. 2009, 2, 243–253. [Google Scholar] [CrossRef]
- Albert, L.J.; Inman, R.D. Molecular Mimicry and Autoimmunity. N. Engl. J. Med. 1999, 341, 2068–2074. [Google Scholar] [CrossRef]
- Ivanov, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfarb, K.C.; Santee, C.A.; Lynch, S.V.; et al. Induction of Intestinal Th17 Cells by Segmented Filamentous Bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Stern, M.E.; Schaumburg, C.S.; Pflugfelder, S.C. Dry Eye as a Mucosal Autoimmune Disease. Int. Rev. Immunol. 2013, 32, 19–41. [Google Scholar] [CrossRef] [PubMed]
- Perez, V.L.; Pflugfelder, S.C.; Zhang, S.; Shojaei, A.; Haque, R. Lifitegrast, a Novel Integrin Antagonist for Treatment of Dry Eye Disease. Ocul. Surf. 2016, 14, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Ma, J.-F.; Chang, C.; Xu, T.; Gao, C.-Y.; Gershwin, M.E.; Lian, Z.-X. Immunobiology of T Cells in Sjögren’s Syndrome. Clin. Rev. Allergy Immunol. 2020, 1–21. [Google Scholar] [CrossRef]
- Ainola, M.; Porola, P.; Takakubo, Y.; Przybyla, B.; Kouri, V.P.; Tolvanen, T.A.; Hänninen, A.; Nordström, D.C. Activation of plasmacytoid dendritic cells by apoptotic particles—Mechanism for the loss of immunological tolerance in Sjögren’s syndrome. Clin. Exp. Immunol. 2018, 191, 301–310. [Google Scholar] [CrossRef]
- Verstappen, G.M.; Corneth, O.B.; Bootsma, H.; Kroese, F.G. Th17 cells in primary Sjögren’s syndrome: Pathogenicity and plasticity. J. Autoimmun. 2018, 87, 16–25. [Google Scholar] [CrossRef]
- Bacman, S.; Sterin-Borda, L.; Camusso, J.J.; Arana, R.; Hübscher, O.; Borda, E. Circulating antibodies against rat parotid gland M3 muscarinic receptors in primary Sjögren’s syndrome. Clin. Exp. Immunol. 1996, 104, 454–459. [Google Scholar] [CrossRef]
- Baudouin, C.; Messmer, E.M.; Aragona, P.; Geerling, G.; Akova, Y.A.; Benítez-Del-Castillo, J.; Boboridis, K.G.; Merayo-Lloves, J.; Rolando, M.; Labetoulle, M. Revisiting the vicious circle of dry eye disease: A focus on the pathophysiology of meibomian gland dysfunction. Br. J. Ophthalmol. 2016, 100, 300–306. [Google Scholar] [CrossRef]
- Yoon, C.H.; Ryu, J.S.; Hwang, H.S.; Kim, M.K. Comparative Analysis of Age-Related Changes in Lacrimal Glands and Meibomian Glands of a C57BL/6 Male Mouse Model. Int. J. Mol. Sci. 2020, 21, 4169. [Google Scholar] [CrossRef]
- Dogru, M.; Kojima, T.; Şimşek, C.; Tsubota, K. Potential Role of Oxidative Stress in Ocular Surface Inflammation and Dry Eye Disease. Investig. Opthalmology Vis. Sci. 2018, 59, DES163–DES168. [Google Scholar] [CrossRef]
- Uchino, Y.; Kawakita, T.; Miyazawa, M.; Ishii, T.; Onouchi, H.; Yasuda, K.; Ogawa, Y.; Shimmura, S.; Ishii, N.; Tsubota, K. Oxidative Stress Induced Inflammation Initiates Functional Decline of Tear Production. PLoS ONE 2012, 7, e45805. [Google Scholar] [CrossRef] [PubMed]
- Jester, J.V.; Parfitt, G.J.; Brown, D.J. Meibomian gland dysfunction: Hyperkeratinization or atrophy? BMC Ophthalmol. 2015, 15 (Suppl. S1), 3–11. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Nandoskar, P.; Lu, M.; Thomas, P.; Trousdale, M.D.; Wang, Y. Changes of Aquaporins in the Lacrimal Glands of a Rabbit Model of Sjögren’s Syndrome. Curr. Eye Res. 2011, 36, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Delporte, C. Aquaporins in Secretory Glands and their Role in Sjögren’s Syndrome. In Aquaporins; Beitz, E., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 185–201. [Google Scholar]
- Soyfoo, M.S.; Chivasso, C.; Perret, J.; Delporte, C. Involvement of Aquaporins in the Pathogenesis, Diagnosis and Treatment of Sjögren’s Syndrome. Int. J. Mol. Sci. 2018, 19, 3392. [Google Scholar] [CrossRef] [PubMed]
- van Deursen, J.M. The role of senescent cells in ageing. Nat. Cell Biol. 2014, 509, 439–446. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nat. Cell Biol. 2013, 500, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc. Natl. Acad. Sci. USA 2010, 107, 12204–12209. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic Treg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef]
- Barr, J.Y.; Wang, X.; Meyerholz, D.K.; Lieberman, S.M. CD8 T cells contribute to lacrimal gland pathology in the nonobese diabetic mouse model of Sjögren syndrome. Immunol. Cell Biol. 2017, 95, 684–694. [Google Scholar] [CrossRef]
- Brkic, Z.; Corneth, O.; van Helden-Meeuwsen, C.G.; Dolhain, R.J.; Maria, N.I.; Paulissen, S.M.J.; Davelaar, N.; Van Hamburg, J.P.; van Daele, P.L.; Dalm, V.A.; et al. T-helper 17 cell cytokines and interferon type I: Partners in crime in systemic lupus erythematosus? Arthritis Res. Ther. 2014, 16, R62. [Google Scholar] [CrossRef]
- Pernis, A.B. Th17 cells in rheumatoid arthritis and systemic lupus erythematosus. J. Intern. Med. 2009, 265, 644–652. [Google Scholar] [CrossRef] [PubMed]
- de Paiva, C.S.; Jones, D.B.; Stern, M.E.; Bian, F.; Moore, Q.L.; Corbiere, S.; Streckfus, C.F.; Hutchinson, D.S.; Ajami, N.J.; Petrosino, J.F.; et al. Altered Mucosal Microbiome Diversity and Disease Severity in Sjögren Syndrome. Sci. Rep. 2016, 6, 23561. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Schaefer, L.; Bian, F.; Yu, Z.; Pflugfelder, S.C.; Britton, R.A.; De Paiva, C.S. Dysbiosis Modulates Ocular Surface Inflammatory Response to Liposaccharide. Investig. Opthalmology Vis. Sci. 2019, 60, 4224–4233. [Google Scholar] [CrossRef]
- Zaheer, M.; Wang, C.; Bian, F.; Yu, Z.; Hernandez, H.; De Souza, R.G.; Simmons, K.T.; Schady, D.; Swennes, A.G.; Pflugfelder, S.C.; et al. Protective role of commensal bacteria in Sjögren Syndrome. J. Autoimmun. 2018, 93, 45–56. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, J.; Bu, J.; Tang, L.-Y.; Yang, Y.; Ouyang, W.; Lin, X.; Liu, Z.; Huang, C.; Quantock, A.J.; et al. High-fat diet induces dry eye-like ocular surface damages in murine. Ocul. Surf. 2020, 18, 267–276. [Google Scholar] [CrossRef]
- Szymula, A.; Rosenthal, J.; Szczerba, B.M.; Bagavant, H.; Fu, S.M.; Deshmukh, U.S. T cell epitope mimicry between Sjogren’s syndrome Antigen A (SSA)/Ro60 and oral, gut, skin and vaginal bacteria. Clin. Immunol. 2014, 152, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.; Choi, S.H.; Yoon, C.H.; Kim, M.K. Gut dysbiosis is prevailing in Sjögren’s syndrome and is related to dry eye severity. PLoS ONE 2020, 15, e0229029. [Google Scholar] [CrossRef] [PubMed]
- Mendez, R.; Watane, A.; Farhangi, M.; Cavuoto, K.M.; Leith, T.; Budree, S.; Galor, A.; Banerjee, S. Gut microbial dysbiosis in individuals with Sjögren’s syndrome. Microb. Cell Factories 2020, 19, 1–13. [Google Scholar] [CrossRef]
- Mandl, T.; Marsal, J.; Olsson, P.; Ohlsson, B.; Andréasson, K. Severe intestinal dysbiosis is prevalent in primary Sjögren’s syndrome and is associated with systemic disease activity. Arthritis Res. 2017, 19, 237. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.-L.; Lin, T.-L.; Chang, C.-J.; Wu, T.-R.; Lai, W.-F.; Lu, C.-C.; Lai, H.-C. Probiotics, prebiotics and amelioration of diseases. J. Biomed. Sci. 2019, 26, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Alookaran, J.J.; Rhoads, J.M. Probiotics in Autoimmune and Inflammatory Disorders. Nutrients 2018, 10, 1537. [Google Scholar] [CrossRef]
- Kawashima, M.; Nakamura, S.; Izuta, Y.; Inoue, S.; Tsubota, K. Dietary Supplementation with a Combination of Lactoferrin, Fish Oil, and Enterococcus faecium WB2000 for Treating Dry Eye: A Rat Model and Human Clinical Study. Ocul. Surf. 2016, 14, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Choi, S.H.; Kim, Y.J.; Jeong, H.J.; Ryu, J.S.; Lee, H.J.; Kim, T.W.; Im, S.-H.; Oh, J.Y.; Kim, M.K. Clinical Effect of IRT-5 Probiotics on Immune Modulation of Autoimmunity or Alloimmunity in the Eye. Nutrients 2017, 9, 1166. [Google Scholar] [CrossRef]
- Chisari, G.; Chisari, E.M.; Ozyalcin, E.; Borzì, A.M.; Chisari, C.G. Aging Eye Microbiota in Dry Eye Syndrome in Patients Treated with Enterococcus faecium and Saccharomyces boulardii. Curr. Clin. Pharmacol. 2018, 12, 99–105. [Google Scholar] [CrossRef]
- Chisari, G.; Chisari, E.M.; Francaviglia, A.; Chisari, C.G. The mixture of bifidobacterium associated with fructo-oligosaccharides reduces the damage of the ocular surface. La Clin. Ter. 2017, 168, e181–e185. [Google Scholar]
- Kawashima, M.; Tsuno, S.; Matsumoto, M.; Tsubota, K. Hydrogen-producing milk to prevent reduction in tear stability in persons using visual display terminals. Ocul. Surf. 2019, 17, 714–721. [Google Scholar] [CrossRef]
- Hansen, C.H.F.; Larsen, C.S.; Petersson, H.O.; Zachariassen, L.F.; Vegge, A.; Lauridsen, C.; Kot, W.; Krych, Ł.; Nielsen, D.S.; Hansen, A.K. Targeting gut microbiota and barrier function with prebiotics to alleviate autoimmune manifestations in NOD mice. Diabetologia 2019, 62, 1689–1700. [Google Scholar] [CrossRef]
- Ghattargi, V.; Gaikwad, M.A.; Meti, B.S.; Nimonkar, Y.S.; Dixit, K.; Prakash, O.; Shouche, Y.S.; Pawar, S.P.; Dhotre, D. Comparative genome analysis reveals key genetic factors associated with probiotic property in Enterococcus faecium strains. BMC Genom. 2018, 19, 652. [Google Scholar] [CrossRef]
- Azad, M.; Kalam, A.; Sarker, M.; Li, T.; Yin, J. Probiotic Species in the Modulation of Gut Microbiota: An Overview. Biomed Res. Int. 2018, 2018, 9478630. [Google Scholar] [CrossRef] [PubMed]
- Papizadeh, M.; Rohani, M.; Nahrevanian, H.; Javadi, A.; Pourshafie, M.R. Probiotic characters of Bifidobacterium and Lactobacillus are a result of the ongoing gene acquisition and genome minimization evolutionary trends. Microb. Pathog. 2017, 111, 118–131. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, G.; Smith, A.J.; Thakker, B.; Kinsella, J. Yoghurt biotherapy: Contraindicated in immunosuppressed patients? Postgrad. Med. J. 2002, 78, 366–367. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pflugfelder, S.C.; Corrales, R.M.; De Paiva, C.S. T helper cytokines in dry eye disease. Exp. Eye Res. 2013, 117, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Clemente, J.C.; Manasson, J.; Scher, J.U. The role of the gut microbiome in systemic inflammatory disease. BMJ 2018, 360, j5145. [Google Scholar] [CrossRef]
- Asano, Y.; Hiramoto, T.; Nishino, R.; Aiba, Y.; Kimura, T.; Yoshihara, K.; Koga, Y.; Sudo, N. Critical role of gut microbiota in the production of biologically active, free catecholamines in the gut lumen of mice. Am. J. Physiol. Liver Physiol. 2012, 303, G1288–G1295. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Author, Year | Disease | Class | Increased | Decreased | (+/−) Disease Correlation |
|---|---|---|---|---|---|
| Scher, 2013 [39] | RA | Species | P. copri | (+) P. copri | |
| Genus | Prevotella | Bacteroides | (+) Prevotella | ||
| Chen, 2016 [40] | Genus | Collinsella, Eggerthella, Faecalibacterium | (+) Collinsella | ||
| Vaahtovuo, 2008 [44] | Genus /species | Bacteroides, Bifidobacteria Porphyromonas, Prevotella, /B. fragilis, C. coccoides E. rectale, | |||
| He, 2016 [45] | SLE | Genus | Eggerthella, Eubacterium, Flavonifractor, Incertae sedis, Klebsiella, Prevotella, Rhodococcus | Dialister, Pseudobutyrivibrio | |
| Hevia, 2014 [46] | Genus | Bacteroides spp | |||
| Guo, 2020 [43] | Genus | Bacteroides, Bilophila, Coprococcus, Parabacteroides, Prevotella, Succinivibrio | Dialister, Gemmiger | (+) Bacteroides, Succinivibrio, Bilophila, Parabateroides (−) Gemmiger, Dialister | |
| Luo, 2018 [47] | Genus | Blautia | Odoribacter | ||
| van der Meulen, 2019 [36] | Genus | Alistipes, Bacteroides, Proteobacteria | |||
| Species | B.ovatus, B. theta, B. uniformis, B. vulgatus | ||||
| Zegarra-Ruiz, 2019 [48] | Genus | Lactobacillus | |||
| Azzouz, 2019 [42] | Species | Ruminococcus gnavus | (+) Ruminococcus gnavus |
| Author, Year | Analysis | Diversity | Taxa | Interpretation | ||
|---|---|---|---|---|---|---|
| Classes | Increased | Decreased | ||||
| Human study | ||||||
| Kalyana, 2018 [65] | Uveitis | α↓ β+ | Family | Lachnospiraceae, Ruminococcaceae | ↓Anti-inflammatory bacteria ↓Butyrate producing bacteria ↑Proinflammatory bacteria | |
| Genus | Bacteroides, Blautia, Clostridium, Coprococcus, Dialister, Dorea, Faecalibacterium, Lachnospira, Odoribacter, Oscillospira, Megasphaera, Mitsuokella,Roseburia, Ruminococcus | |||||
| Species | Prevotella copri | Akkermansia municiphala, Bifidobacterium adolescentis, Faecalibacterium prausnitzii, Veillonella dispar | ||||
| Huang, 2018 [60] | Uveitis | α→ β+ | Class | Negativicutes | (+) Roseburia ∝ linoleic acid | |
| Order | Oceanospirillales, Selenomonadales | |||||
| Family | Clostridiales_Incertae Sedis XI, Halomonadaceae | |||||
| Genus | Veillonella | Roseburia | ||||
| Ye, 2020 [57] a | VKH | α→ β- | Genus | Azospirillum | ↓Butyrate or lactate producing bacteria ↑Gram(−) bacteria | |
| Species | Paraprevotella clara | Azospirillum spp., Bifidobacterium spp., Clostridium spp. | ||||
| Ye, 2018 [61] a | Behcet’s disease | α: NA β: NA | Phylum | Actinobacteria, Proteobacteria | ↓Butyrate producing bacteria ↑Opportunistic bacteria | |
| Species | Actinomyces spp., Bilophila spp., Corynebacterium spp., Fusobacterium spp., Parabacteroides spp. , Paraprevotella spp. , Stenotrophomonas spp. | Clostridium spp. | ||||
| Shimizu, 2019 [62] | Behcet’s disease | α→ β+ | Species | Acidaminococcus spp., Bifidobacterium bifidum, Eggerthella lenta, Lactobacillus iners, Lactobacillus mucosae, Lactobacillus salivarius, Streptococcus spp. | Butyrivibrio species, Filifactor species, Streptococcus infantis , Megamonas hypermegale | ↓SCFAs producing bacteria |
| Zinkernagel, 2017 [66] a | NAMD | α: NA β+ | Family | Oscillospiraceae | ↑ Inflammatory bacteria | |
| Genus | Anaerotruncus | |||||
| Species | Eubacterium ventriosum, Ruminococcus torques | Bacteroides eggerthii | ||||
| Gong, 2020 [56] | POAG | α→ β+ | Family | Enterobacteriaceae, Prevotellaceae | (−) Faecalibacterium ∝ VF-MD (+) Streptococcus ∝ RNFLT | |
| Genus | Megamonas | |||||
| Species | Escherichia coli | Bacteroides plebeius | ||||
| Shi, 2020 [67] | NMOSD | α→ β+ | Genus | Flavonifractor, Streptococcus | Blautia, Coprococcus, Faecalibacterium, Fusicatenibacter Lachnospiracea_incertae_sedis, Prevotella, Romboutsia, Roseburia, | ↑Pathogenic bacteria ↓Commensal bacteria |
| Gong, 2019 [68] | NMOSD | α→ β+ | Genus | Shigella, Streptococcus | Faecalibacterium,Lachnospira Roseburia | ↓SCFAs producing bacteria ↑Pathogenic bacteria (+) Streptococcus disease severity |
| Species | Streptococcusspp. (S. oralis, S. salivarius, S. parasanguinis, S. pneumonia, and S. mitis) | |||||
| Cree, 2016 [69] | NMOSD | α: NA β+ | Species | Clostridium perfringens | ||
| Mouse study | ||||||
| Janowitz, 2019 [63] | EAU | α↓ β+ | Phylum | Firmicutes | Bacteroidetes, Proteobacteria | ↑F/B ratio (−) α-diversity ∝ uveitis |
| Class | Bacteroidia | |||||
| Genus | Anaeroplasma, Clostridium, Lactobacillus, Parabacteroides, Prevotella | Desulfovibrio, Ruminococcus | ||||
| Du, 2020 [64] | EAU b | α→ β+ | Class | Bacilli | ||
| Family | Anaeroplasmataceae, Lactobacillaceae | Muribaculaceae, Ruminococcaceae | ↑Lactobacillaceae, increasing type I IFN ↓Treg-enhancing Bacteria | |||
| Order | Anaeroplasmatales, Lactobacillales | |||||
| Genus | Anaeroplasma, Lactobacillus | Akkermansia, Bacteroides,Oscillibacter | ||||
| Species | Lactobacillus gasseri, Lactobacillus intestinalis | Bacteroides sartorii, Parabacteroides goldsteinii | ||||
| Lin, 2014 [70] | AS | α: NA β+ | Genus | Paraprevotella | ↑RA-causing Bacteria | |
| Species | Bacteroides vulgatus | Akkermansia muciniphila | ||||
| Andriessen, 2016 [71] | AMD | α→ β+ | Phylum | Firmicutes,Proteobacteria | Bacteroidetes | ↑F/B ratio |
| Rowan, 2017 [58] | AMD | α↑ β+ | Phylum | Firmicutes | Bacteroidetes | ↑F/B ratio (+) Firmicutes, Clostridia ∝ phenotype (−) Bacteroidales, Erysipelotrichi ∝ phenotype |
| Class | Clostridia | Erysipelotrichi | ||||
| Order | Bacteroidales | |||||
| Kugadas, 2017 [72] | SW vs. B6 | α↓ β: NA | Genus | Bacteroides,Dysgonomonas, Prevotella | (+) Bacteroides acidifaciens ∝ LG IgA transcription | |
| Non- Sjögren Dry Eye | Sjögren’s Syndrome-Related Dry Eye | |
|---|---|---|
| Immunologic | IL-1↑, IL-6↑,TNF-α↑ MMPs↑ Neutrophil↑, NK cell↑, Macrophage↑ TH1/ IFN-γ↑ TH17/IL-17↑ [87,88,89,90,91,92,93,94] | Type I IFN (α, β)↑ TNF-α↑, IL-2, IL-7↑ BAFF↑ TH1/ IFN-γ↑, TH17/ IL-17↑ Cytotoxic CD8+ T cell↑ B cell↑, Autoantibody↑ pDC in gland↑ [12,87,95,96,97] |
| Non-immunologic | Hyperkeratinisation of MG orifice Atrophy Oxidative stress Senescence [87,98,99,100,101,102] | Hormone AQP dysfunction? [87,103,104,105] |
| Author, Year | Treatment | Subjects | Representative Gut Microbiota | Change in OS/LG/dLN |
|---|---|---|---|---|
| de Paiva, 2016 [113] | Antibiotics a (24 days) | DS B6 | Number of OTUs↓ α↓ β+ ↓ Alistipes, Allobaculum, Bacteroides, Blautia, Clostridium, Desulfovibrio, Intestinimonas, Lactobacillus (Genera) ↑ Enterobacter, Escherichia/Shigella, Parasutterella, Pseudomonas, Staphylococcus (Genera) | Goblet cell density↓ Corneal staining↑ CD4+ T cell↑(OS) IFN-γ↑(OS) IL-13↓(OS) |
| Wang, 2018 [84] | Germ-free | B6 | Goblet cell density↓ Corneal staining↑ Tear EGF↓ CD8+ & CD4+ T cell↑(LG) TH1+ cell↑(LG) IL-12+CD11b+CD11c+ cell↑(OS, dLN) MHC II, IFN-γ, IL-12 & Caspase 3↑(LG) | |
| Zaheer, 2018 [115] | Germ-free | CD25KO | Goblet cell density↓ Corneal staining↑ TH1+ cell↑(LG, dLN) B220+ cell↑(LG) IL-12+CD11c+MHC−- cell↑(LG, dLN, OS) IFN-γ & IL-12↑(LG) | |
| Antibiotics a (28 days) | CD25KO | N/A | IFN-γ & IL-12↑(LG) | |
| Wang, 2019 [114] | Antibiotics a (14 days) | B6 | α↓ β+ ↓Akkermansia, Bacteroidales, Bacteroides, Lachnospiraceae, Oscillibacter, Parasutterella, Ruminococcaceae, Ruminiclostridium (Genera) ↑Bacillus, Curtobacterium, Escherichia/Shigella, Firmicutes, Lactococcus, Megasphaera, Staphylococcus (Genera) | Serum LPS↑ MHC II+CD11c+CD11b+ cell↑(dLN) Inflammatory response b to topical LPS↑(cornea) |
| Germ-free | B6 | Inflammatory response c to topical LPS↑(OS) | ||
| Wu, 2020 [116] | High-fat diet d | B6 | N/A | Goblet cell density↓ Corneal staining↑ Tear secretion↓ Squamous metaplasia↑(OS) Oxidative stress/apoptosis↑(OS) |
| Author, Year | α | Class | Increased | Decreased | (+/−) Disease Correlation |
|---|---|---|---|---|---|
| pSS | |||||
| de Paiva, 2016 [113] | ↓ | Genus | Anaerostipes, Bifidobacterium, Bilophila, Blautia, Escherichia/Shigella, Lachnospira, Moryella, Pseudobutyrivibrio, Streptococcus | Bacteroides, Faecalibacterium, Haemophilus, Odoribacter, Parabacteroides, Prevotella | (−) Diversity ∝ ocular/systemic disease index |
| Mandl, 2017 [120] | N/A | Genus | N/A | Alistipes, Bifidobacterium | (+) Dysbiosis a ∝ disease activity /F-calprotectin (−) Dysbiosis a ∝ complementemia |
| Species | N/A | Faecalibacterium prausnitzii | |||
| van der Meulen, 2019 [36] | ↓ | Phylum | Bacteroidetes, Proteobacteria | Firmicutes/Bacteroidetes ratio | (+) Clostridium sensu stricto ∝ anti-La/SSB antibody |
| Genus | Alistipes, Bacteroides, Barnesiella, Lachnosclostridium, Lachnospira, Parasutterella | Actinomyces, Clostridium sensu stricto, Enterorhabdus, Romboutsia, Senegalimassilia, Slackia, Turicibacter | |||
| Species | Bacteroides ovatus, Bacteroides uniformis, Bacteroides vulgatus | ||||
| Moon, 2020 [118] | - | Phylum | Bacteriodetes | Firmicutes/Bacteroidetes ratio, Actinobacteria | b (−) Prevotella ∝ tear secretion (+) Actinobacteria ∝ TBUT (−) Prevotella ∝ TBUT |
| Class | Clostridia | ||||
| Genus | Alistipes, Odoribacter, Prevotella, Veillonella | Agathobacter, Bifidobacterium, Blautia, Dorea | |||
| Species | Bifidobacterium longum, Eubacterium hallii | ||||
| DES c | |||||
| Moon, 2020 [118] | - | Genus | Veillonella | Subdoligranulum | b |
| pSS & non-SS d | |||||
| Mendez, 2020 [119] | - | Phylum | Actinobacteria, Bacteroidetes, Proteobacteria | Firmicutes | (+) Eubacteriaceae, Eggerthellaceae, ∝ DEQ5 (−) Ruminococcaceae ∝ DEQ5 (−) Akkermanciaceae ∝ tear secretion |
| Order | Clostridiales | ||||
| Family | Actinomycetaceae, Akkermanciaceae, Coriobacteriaceae, Eggerthellaceae, Eubacteriaceae, Lactobacillaceae | Lachnospiraceae, Ruminococcaceae | |||
| Genus | Megasphaera, Parabacteroides, Prevotella | Bacteroides, Faecalibacterium, Veillonella | |||
| Author, Year | Tx | Tx Period | Subjects | Representative Gut Microbiota | Change in OS/LG/dLN |
|---|---|---|---|---|---|
| Rodent Study | |||||
| Kawashima, 2016 [125] | Fish oil, lactoferrin, zinc, vitamin C, lutein, vitamin E, γ-aminobutanoic acid & E. faecium WB2000 | 2 days | DS rats | N/A | Tear secretion↑ ROS↓(LG) |
| Kim, 2017 [126] | L. casei, L. acidophilus, L. reuteri, B. bifidum & S. thermophiles | 3 weeks | NOD.B10.H2 b | N/A | Tear secretion↑ Corneal staining↓ Inflammation foci a↓(LG) CD8+IFN-γhi T cell↓(dLN) Treg cell↑(dLN) |
| Choi, 2020 [86] | L. casei, L. acidophilus, L. reuteri, B. bifidum & S. thermophiles | 3 weeks | NOD.B10.H2 b | ↑Lactobacillus helveticus, L. hamsteri, L. reuteri, L. casei, L. brantae, L. amylovorous, Akkermansia municipila, Aerococcus viridans, B. bifidum, Streptococcus salivarius ↓Lactobacillus intestinalis | Tear secretion↑ Corneal staining↓ Immune response genes b↓(LG) IL-10↑(OS) IL-1b↓(OS) |
| Human Study | |||||
| Kawashima, 2016 [125] | Fish oil, lactoferrin, zinc, vitamin C, lutein, vitamin E, γ-aminobutanoic acid & E. faecium WB2000 | 8 weeks | DES c | N/A | Scored subjective symptoms d↓ Tear secretion↑ |
| Chisari, 2017 [127] | S. boulardii MUCL 53837 & E. faecium LMG S-28935 | 30 days | DES e | N/A | Subjective dry eye symptoms f↓ TBUT↑ Tear secretion↑ |
| Chisari, 2017 [128] | B. lactis DSM 25566 & B. bifido DSM 25565 | 30 days | DES e | N/A | Tear secretion↑ TBUT↑ |
| Kawashima, 2019 [129] | Hydrogen-producing milk | 3 weeks | DES c | N/A | TBUT↑ (♀) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, J.; Yoon, C.H.; Choi, S.H.; Kim, M.K. Can Gut Microbiota Affect Dry Eye Syndrome? Int. J. Mol. Sci. 2020, 21, 8443. https://doi.org/10.3390/ijms21228443
Moon J, Yoon CH, Choi SH, Kim MK. Can Gut Microbiota Affect Dry Eye Syndrome? International Journal of Molecular Sciences. 2020; 21(22):8443. https://doi.org/10.3390/ijms21228443
Chicago/Turabian StyleMoon, Jayoon, Chang Ho Yoon, Se Hyun Choi, and Mee Kum Kim. 2020. "Can Gut Microbiota Affect Dry Eye Syndrome?" International Journal of Molecular Sciences 21, no. 22: 8443. https://doi.org/10.3390/ijms21228443
APA StyleMoon, J., Yoon, C. H., Choi, S. H., & Kim, M. K. (2020). Can Gut Microbiota Affect Dry Eye Syndrome? International Journal of Molecular Sciences, 21(22), 8443. https://doi.org/10.3390/ijms21228443