Neuropeptide Y as Alternative Pharmacotherapy for Antidepressant-Resistant Social Fear


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
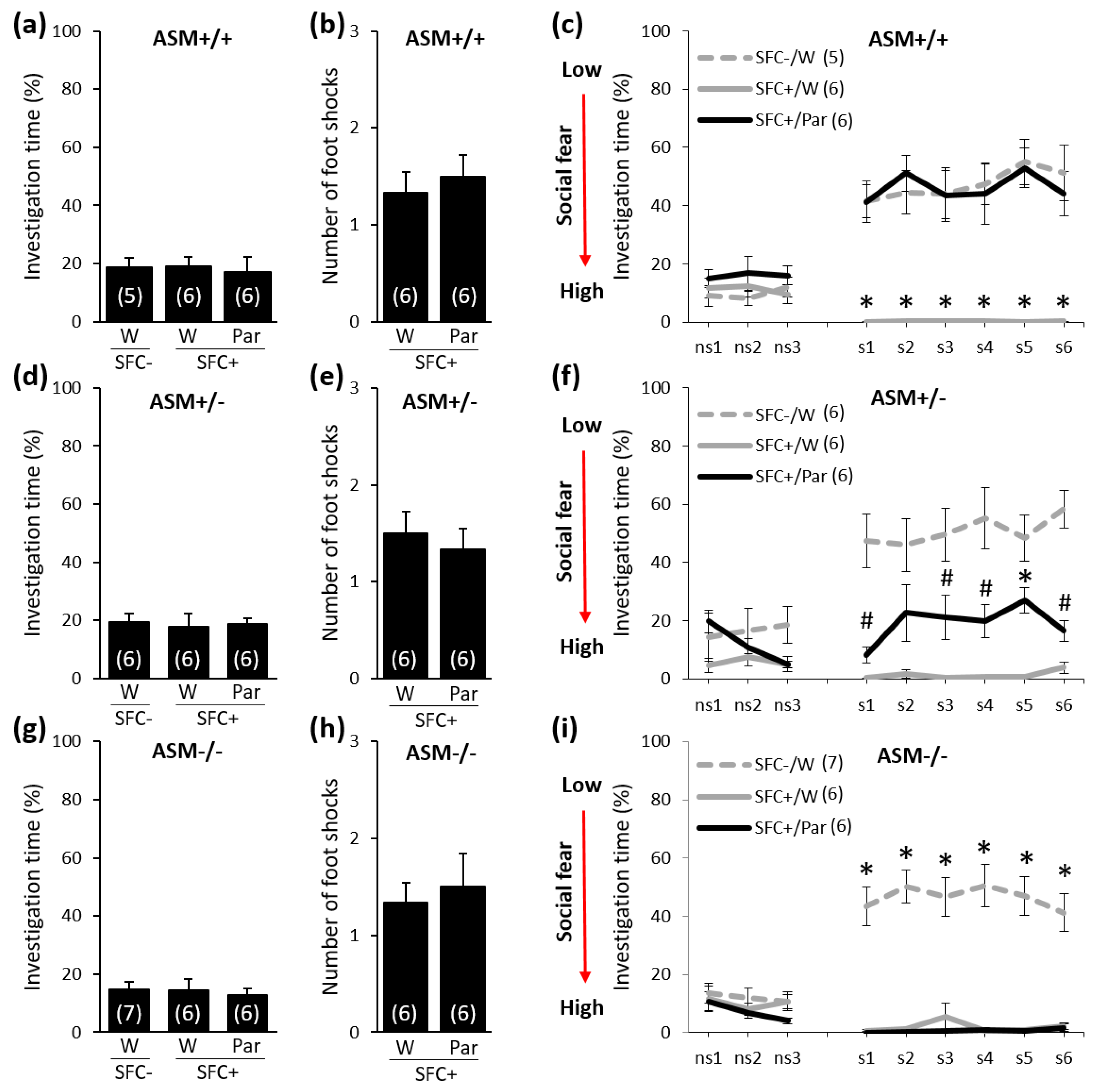
2.1. Chronic Paroxetine Treatment (10 mg/kg/Day for 14 Days) Reduces Social Fear in ASM+/+ and ASM+/− Mice but Not in ASM−/− Mice
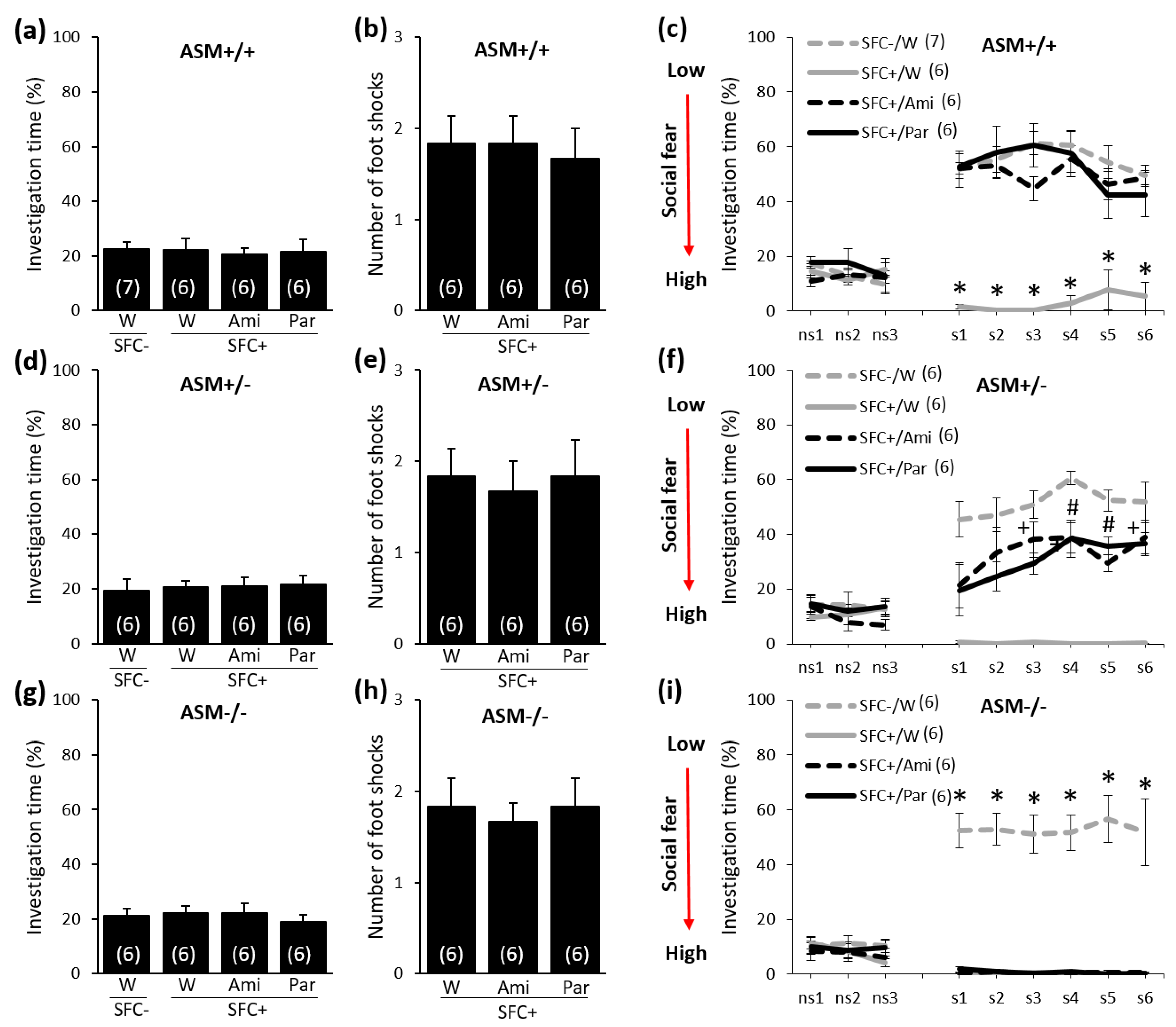
2.2. Chronic Paroxetine (20 mg/kg/Day for 28 Days) or Amitriptyline (180 mg/L for 28 Days) Treatment Reduces Social Fear in ASM+/+ and ASM+/− Mice but Not in ASM−/− Mice
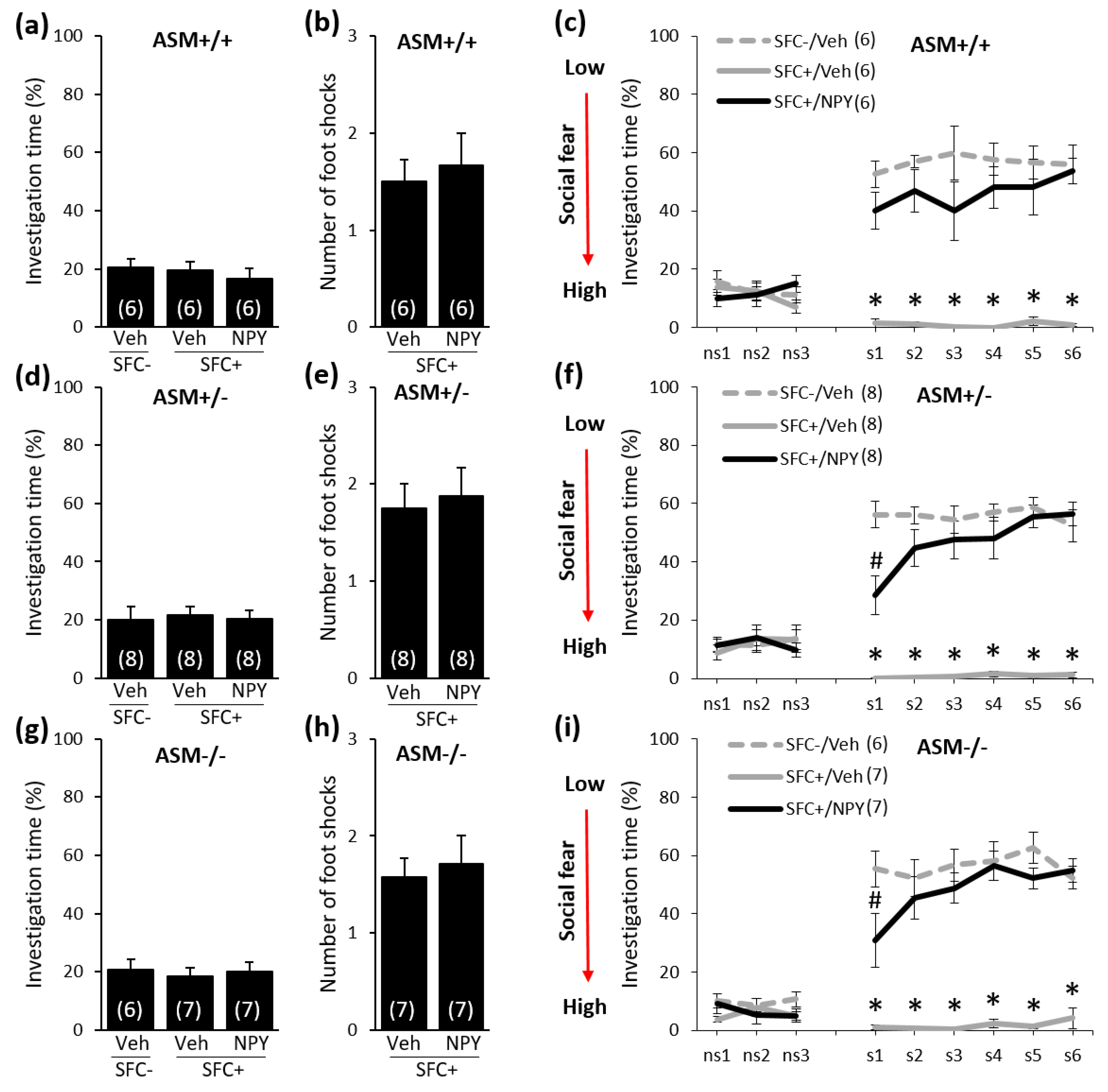
2.3. Acute NPY Treatment Reduces Social Fear in ASM+/+, ASM+/−, and ASM−/− Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Social Fear Conditioning (SFC) Paradigm
4.3. Social Fear Extinction
4.4. Antidepressant Treatment
4.5. Stereotaxic Cannula Implantation
4.6. Intracerebral Infusions
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kessler, R.C.; Petukhova, M.; Sampson, N.A.; Zaslavsky, A.M.; Wittchen, H.U. Twelve-month and lifetime prevalence and lifetime morbid risk of anxiety and mood disorders in the United States. Int. J. Methods Psychiatr. Res. 2012, 21, 169–184. [Google Scholar] [CrossRef] [PubMed]
- McGuire, J.F.; Lewin, A.B.; Storch, E.A. Enhancing exposure therapy for anxiety disorders, obsessive-compulsive disorder and post-traumatic stress disorder. Expert Rev. Neurother. 2014, 14, 893–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, T.; Hattingh, C.J.; Kariuki, C.M.; Tromp, S.A.; van Balkom, A.J.; Ipser, J.C.; Stein, D.J. Pharmacotherapy for social anxiety disorder (SAnD). Cochrane Database Syst. Rev. 2017, 10. [Google Scholar] [CrossRef]
- Blanco, C.; Antia, S.X.; Liebowitz, M.R. Pharmacotherapy of social anxiety disorder. Biol. Psychiatry 2002, 51, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Springer, K.S.; Levy, H.C.; Tolin, D.F. Remission in CBT for adult anxiety disorders: A meta-analysis. Clin. Psychol. Rev. 2018, 61, 1–8. [Google Scholar] [CrossRef] [PubMed]
- de Quidt, M.E.; Emson, P.C. Distribution of neuropeptide Y-like immunoreactivity in the rat central nervous system-II. Immunohistochemical analysis. Neuroscience 1986, 18, 545–618. [Google Scholar] [CrossRef]
- Lynch, D.R.; Walker, M.W.; Miller, R.J.; Snyder, S.H. Neuropeptide Y receptor binding sites in rat brain: Differential autoradiographic localizations with 125I-peptide YY and 125I-neuropeptide Y imply receptor heterogeneity. J. Neurosci. 1989, 9, 2607–2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blomqvist, A.G.; Herzog, H. Y-receptor subtypes-how many more? Trends Neurosci. 1997, 20, 294–298. [Google Scholar] [CrossRef]
- Colmers, W.F.; Bleakman, D. Effects of neuropeptide Y on the electrical properties of neurons. Trends Neurosci. 1994, 17, 373–379. [Google Scholar] [CrossRef]
- Michalkiewicz, M.; Michalkiewicz, T.; Kreulen, D.L.; McDougall, S.J. Increased blood pressure responses in neuropeptide Y transgenic rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, 417–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magni, P. Hormonal control of the neuropeptide Y system. Curr. Protein Pept. Sci. 2003, 4, 45–57. [Google Scholar] [CrossRef]
- Hökfelt, T.; Stanic, D.; Sanford, S.D.; Gatlin, J.C.; Nilsson, I.; Paratcha, G.; Ledda, F.; Fetissov, S.; Lindfors, C.; Herzog, H.; et al. NPY and its involvement in axon guidance, neurogenesis, and feeding. Nutrition 2008, 24, 860–868. [Google Scholar] [CrossRef]
- Stanley, B.G.; Leibowitz, S.F. Neuropeptide Y: Stimulation of feeding and drinking by injection into the paraventricular nucleus. Life Sci. 1984, 35, 2635–2642. [Google Scholar] [CrossRef]
- Kask, A.; Nguyen, H.P.; Pabst, R.; Von Hörsten, S. Neuropeptide Y Y1 receptor-mediated anxiolysis in the dorsocaudal lateral septum: Functional antagonism of corticotropin-releasing hormone-induced anxiety. Neuroscience 2001, 104, 799–806. [Google Scholar] [CrossRef]
- Sajdyk, T.J.; Vandergriff, M.G.; Gehlert, D.R. Amygdalar neuropeptide Y Y1 receptors mediate the anxiolytic-like actions of neuropeptide Y in the social interaction test. Eur. J. Pharmacol. 1999, 368, 143–147. [Google Scholar] [CrossRef]
- Redrobe, J.P.; Dumont, Y.; Fournier, A.; Quirion, R. The neuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swimming test. Neuropsychopharmacology 2002, 26, 615–624. [Google Scholar] [CrossRef]
- Enman, N.M.; Sabban, E.L.; McGonigle, P.; Van Bockstaele, E.J. Targeting the neuropeptide Y system in stress-related psychiatric disorders. Neurobiol. Stress 2015, 1, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Broqua, P.; Wettstein, J.G.; Rocher, M.N.; Gauthier-Martin, B.; Junien, J.L. Behavioral effects of neuropeptide Y receptor agonists in the elevated plus-maze and fear-potentiated startle procedures. Behav. Pharmacol. 1995, 6, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, R.M.; Holmes, A.; Heilig, M.; Crawley, J.N. Anxiolytic-like actions of centrally-administered neuropeptide Y, but not galanin, in C57BL/6Jmice. Pharmacol. Biochem. Behav. 2005, 80, 427–436. [Google Scholar] [CrossRef]
- Lach, G.; de Lima, T.C. Role of NPY Y1 receptor on acquisition, consolidation and extinction on contextual fear conditioning: Dissociation between anxiety, locomotion and non-emotional memory behavior. Neurobiol. Learn. Mem. 2013, 103, 26–33. [Google Scholar] [CrossRef]
- Gutman, A.R.; Yang, Y.; Ressler, K.J.; Davis, M. The role of neuropeptide Y in the expression and extinction of fear-potentiated startle. J. Neurosci. 2008, 28, 12682–12690. [Google Scholar] [CrossRef] [Green Version]
- Kornhuber, J.; Zoicas, I. Neuropeptide Y reduces expression of social fear via simultaneous activation of Y1 and Y2 receptors. J. Psychopharmacol. 2019, 33, 1533–1539. [Google Scholar] [CrossRef] [Green Version]
- Toth, I.; Neumann, I.D.; Slattery, D.A. Social fear conditioning: A novel and specific animal model to study social anxiety disorder. Neuropsychopharmacology 2012, 37, 1433–1443. [Google Scholar] [CrossRef] [Green Version]
- Toth, I.; Neumann, I.D.; Slattery, D.A. Social fear conditioning as an animal model of social anxiety disorder. Curr. Protoc. Neurosci. 2013. [Google Scholar] [CrossRef]
- Gulbins, E.; Palmada, M.; Reichel, M.; Lüth, A.; Böhmer, C.; Amato, D.; Müller, C.P.; Tischbirek, C.H.; Groemer, T.; Tabatabai, G.; et al. Acid sphingomyelinase-ceramide system mediates effects of antidepressant drugs. Nat. Med. 2013, 19, 934–938. [Google Scholar] [CrossRef] [Green Version]
- Goni, F.M.; Alonso, A. Sphingomyelinases: Enzymology and membrane activity. FEBS Lett. 2002, 531, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Hurwitz, R.; Ferlinz, K.; Sandhoff, K. The tricyclic antidepressant desipramine causes proteolytic degradation of lysosomal sphingomyelinase in human fibroblasts. Biol. Chem. Hoppe Seyler. 1994, 375, 447–450. [Google Scholar] [CrossRef]
- Kornhuber, J.; Tripal, P.; Reichel, M.; Mühle, C.; Rhein, C.; Muehlbacher, M.; Groemer, T.W.; Gulbins, E. Functional Inhibitors of Acid Sphingomyelinase (FIASMAs): A novel pharmacological group of drugs with broad clinical applications. Cell Physiol. Biochem. 2010, 26, 9–20. [Google Scholar] [CrossRef]
- Sterner, E.Y.; Kalynchuk, L.E. Behavioral and neurobiological consequences of prolonged glucocorticoid exposure in rats: Relevance to depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2010, 34, 777–790. [Google Scholar] [CrossRef]
- Sainsbury, A.; Rohner-Jeanrenaud, F.; Grouzmann, E.; Jeanrenaud, B. Acute intracerebroventricular administration of neuropeptide Y stimulates corticosterone output and feeding but not insulin output in normal rats. Neuroendocrinology 1996, 63, 318–326. [Google Scholar] [CrossRef]
- Yang, Y.L.; Chao, P.K.; Lu, K.T. Systemic and intra-amygdala administration of glucocorticoid agonist and antagonist modulate extinction of conditioned fear. Neuropsychopharmacology 2006, 31, 912–924. [Google Scholar]
- Klemfuss, H.; Southerland, S.; Britton, K.T. Cardiovascular actions of neuropeptide Y and social stress. Peptides 1998, 19, 85–92. [Google Scholar] [CrossRef]
- Tovote, P. Central NPY receptor-mediated alteration of heart rate dynamics in mice during expression of fear conditioned to an auditory cue. Regul. Pept. 2004, 120, 205–214. [Google Scholar] [CrossRef]
- Zoicas, I.; Slattery, D.A.; Neumann, I.D. Brain oxytocin in social fear conditioning and its extinction: Involvement of the lateral septum. Neuropsychopharmacology 2014, 39, 3027–3035. [Google Scholar] [CrossRef] [Green Version]
- Zoicas, I.; Neumann, I.D. Maternal separation facilitates extinction of social fear in adult male mice. Behav. Brain Res. 2016, 297, 323–328. [Google Scholar] [CrossRef]
- Zoicas, I.; Menon, R.; Neumann, I.D. Neuropeptide S reduces fear and avoidance of con-specifics induced by social fear conditioning and social defeat, respectively. Neuropharmacology 2016, 108, 284–291. [Google Scholar] [CrossRef]
- Slattery, D.A.; Neumann, I.D.; Flor, P.J.; Zoicas, I. Pharmacological modulation of metabotropic glutamate receptor subtype 5 and 7 impairs extinction of social fear in a time-point-dependent manner. Behav. Brain Res. 2017, 328, 57–61. [Google Scholar] [CrossRef]
- Menon, R.; Grund, T.; Zoicas, I.; Althammer, F.; Fiedler, D.; Biermeier, V.; Bosch, O.J.; Hiraoka, Y.; Nishimori, K.; Eliava, M.; et al. Oxytocin signaling in the lateral septum prevents social fear during lactation. Curr. Biol. 2018, 28, 1066–1078.e6. [Google Scholar] [CrossRef] [Green Version]
- Kornhuber, J.; Huber, S.E.; Zoicas, I. Effects of conditioned social fear on ethanol drinking and vice-versa in male mice. Psychopharmacology 2019, 236, 2059–2067. [Google Scholar] [CrossRef]
- Michalikova, S.; van Rensburg, R.; Chazot, P.L.; Ennaceur, A. Anxiety responses in Balb/c, c57 and CD-1 mice exposed to a novel open space test. Behav. Brain Res. 2010, 207, 402–417. [Google Scholar] [CrossRef]
- Benatti, C.; Alboni, S.; Montanari, C.; Caggia, F.; Tascedda, F.; Brunello, N.; Blom, J.M.C. Central effects of a local inflammation in three commonly used mouse strains with a different anxious phenotype. Behav. Brain Res. 2011, 224, 23–34. [Google Scholar] [CrossRef]
- Painsipp, E.; Köfer, M.J.; Sinner, F.; Holzer, P. Prolonged depression-like behavior caused by immune challenge: Influence of mouse strain and social environment. PLoS ONE 2011, 6, e20719. [Google Scholar] [CrossRef] [Green Version]
- de Sá-Calçada, D.; Roque, S.; Branco, C.; Monteiro, S.; Cerqueira-Rodrigues, B.; Sousa, N.; Palha, J.A.; Correia-Neves, M. Exploring female mice interstrain differences relevant for models of depression. Front. Behav. Neurosci. 2015, 9, 335. [Google Scholar] [CrossRef]
- Faure, A.; Pittaras, E.; Nosjean, A.; Chabout, J.; Cressant, A.; Granon, S. Social behaviors and acoustic vocalizations in different strains of mice. Behav. Brain Res. 2017, 320, 383–390. [Google Scholar] [CrossRef]
- Hsieh, L.S.; Wen, J.H.; Miyares, L.; Lombroso, P.J.; Bordey, A. Outbred CD1 mice are as suitable as inbred C57BL/6J mice in performing social tasks. Neurosci. Lett. 2017, 637, 142–147. [Google Scholar] [CrossRef] [Green Version]
- Kalinichenko, L.S.; Mühle, C.; Eulenburg, V.; Praetner, M.; Reichel, M.; Gulbins, E.; Kornhuber, J.; Müller, C.P. Enhanced alcohol preference and anxiolytic alcohol effects in Niemann-Pick disease model in mice. Front. Neurol. 2019, 10, 731. [Google Scholar] [CrossRef] [Green Version]
- Zoicas, I.; Reichel, M.; Gulbins, E.; Kornhuber, J. Role of acid sphingomyelinase in the regulation of social behavior and memory. PLoS ONE 2016, 11, e0162498. [Google Scholar] [CrossRef]
- Kornhuber, J.; Zoicas, I. Neuropeptide Y prolongs non-social memory and differentially affects acquisition, consolidation, and retrieval of non-social and social memory in male mice. Sci. Rep. 2017, 7, 6821. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kornhuber, J.; Zoicas, I. Neuropeptide Y as Alternative Pharmacotherapy for Antidepressant-Resistant Social Fear. Int. J. Mol. Sci. 2020, 21, 8220. https://doi.org/10.3390/ijms21218220
Kornhuber J, Zoicas I. Neuropeptide Y as Alternative Pharmacotherapy for Antidepressant-Resistant Social Fear. International Journal of Molecular Sciences. 2020; 21(21):8220. https://doi.org/10.3390/ijms21218220
Chicago/Turabian StyleKornhuber, Johannes, and Iulia Zoicas. 2020. "Neuropeptide Y as Alternative Pharmacotherapy for Antidepressant-Resistant Social Fear" International Journal of Molecular Sciences 21, no. 21: 8220. https://doi.org/10.3390/ijms21218220
APA StyleKornhuber, J., & Zoicas, I. (2020). Neuropeptide Y as Alternative Pharmacotherapy for Antidepressant-Resistant Social Fear. International Journal of Molecular Sciences, 21(21), 8220. https://doi.org/10.3390/ijms21218220