MicroRNA Biomarkers in IBD—Differential Diagnosis and Prediction of Colitis-Associated Cancer

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. MicroRNA—An Introduction
3. MiRNAs in IBD
4. MiRNA Biomarkers for IBD Diagnosis
5. MiRNAs and CAC
6. MiRNAs as Predictive Biomarkers and in IBD Treatment
7. Circulating miRNAs vs. Tissue miRNAs
8. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- De Souza, H.S.P.; Fiocchi, C. Immunopathogenesis of IBD: Current state of the art. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Bouma, G.; Strober, W. The immunological and genetic basis of inflammatory bowel disease. Nat. Rev. Immunol. 2003, 3, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, B.A.; Gokhale, R.; Cho, J.H. Clinical aspects and pathophysiology of inflammatory bowel disease. Clin. Microbiol. Rev. 2002, 15, 79–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Shen, B. Overview of Diagnosis and Medical Treatment of Inflammatory Bowel Diseases; Academic Press: New York, NY, USA, 2018; ISBN 9780128113882. [Google Scholar]
- Kappelman, M.D.; Grand, R.J. Does inflammatory bowel disease develop in infants? Inflamm. Bowel Dis. 2008, 14 (Suppl. 2), S6–S8. [Google Scholar] [CrossRef] [PubMed]
- Molodecky, N.A.; Soon, I.S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W.; et al. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology 2012, 142, 46–54.e42. [Google Scholar] [CrossRef] [Green Version]
- Burisch, J.; Jess, T.; Martinato, M.; Lakatos, P.L. The burden of inflammatory bowel disease in Europe. J. Crohn’s Colitis 2013, 7, 322–337. [Google Scholar] [CrossRef] [Green Version]
- Lophaven, S.N.; Lynge, E.; Burisch, J. The incidence of inflammatory bowel disease in Denmark 1980–2013: A nationwide cohort study. Aliment. Pharmacol. Ther. 2017, 45, 961–972. [Google Scholar] [CrossRef]
- Hendriksen, C.; Kreiner, S.; Binder, V. Long term prognosis in ulcerative colitis—Based on results from a regional patient group from the county of Copenhagen. Gut 1985, 26, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Loftus, E.V., Jr. Clinical epidemiology of inflammatory bowel disease: Incidence, prevalence, and environmental influences. Gastroenterology 2004, 126, 1504–1517. [Google Scholar] [CrossRef]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef]
- Chapman, C.G.; Pekow, J. The emerging role of miRNAs in inflammatory bowel disease: A review. Therap. Adv. Gastroenterol. 2015, 8, 4–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, J.S. MicroRNAs: How many in inflammatory bowel disease? Curr. Opin. Gastroenterol. 2016, 32, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, J. microRNAs as therapeutic targets in intestinal diseases. ExRNA 2019, 1, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ambros, V. microRNAs: Tiny Regulators with Great Potential. Cell 2001, 107, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans Heterochronic Gene lin-4 Encodes Small RNAs with Antisense Complementarity to &II-14 Rosalind. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function Review. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef]
- Zarjou, A.; Yang, S.; Abraham, E.; Agarwal, A.; Liu, G. Identification of a microRNA signature in renal fibrosis: Role of miR-21. AJP Ren. Physiol. 2011, 301, F793–F801. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, S.; Hanna, J.; Sherman, M.E.; Figueroa, J.; Rimm, D.L. Quantitative assessment of miR34a as an independent prognostic marker in breast cancer. Br. J. Cancer 2015, 112, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Xuan, Y.; Yang, H.; Zhao, L.; Bond Lau, W.; Bonnie, L.; Ning, R.; Yuehong, H.; Tao, Y.; Xia, Z.; Shengtao, Z.; et al. MicroRNAs in colorectal cancer: Small molecules with big functions. Cancer Lett. 2015, 360, 89–105. [Google Scholar] [CrossRef]
- Nielsen, B.S.; Balslev, E.; Poulsen, T.; Nielsen, D.; Møller, T.; Mortensen, C.E.; Holmstrøm, K.; Høgdall, E. miR-21 expression in cancer cells may not predict resistance to adjuvant trastuzumab in primary breast cancer. Front. Oncol. 2014, 4, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Møller, T.; James, J.P.; Holmstrøm, K.; Sørensen, F.B.; Lindebjerg, J.; Nielsen, B.S. Co-Detection of miR-21 and TNF-α mRNA in Budding Cancer Cells in Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjaer-Frifeldt, S.; Hansen, T.F.; Nielsen, B.S.; Joergensen, S.; Lindebjerg, J.; Soerensen, F.B.; dePont Christensen, R.; Jakobsen, A.; Group, D.C.C. The prognostic importance of miR-21 in stage II colon cancer: A population-based study. Br. J. Cancer 2012, 107, 1169–1174. [Google Scholar] [CrossRef] [Green Version]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef]
- Galimberti, D.; Villa, C.; Fenoglio, C.; Serpente, M.; Ghezzi, L.; Cioffi, S.M.G.; Arighi, A.; Fumagalli, G.; Scarpini, E. Circulating miRNAs as potential biomarkers in alzheimer’s disease. J. Alzheimer’s Dis. 2014, 42, 1261–1267. [Google Scholar] [CrossRef]
- Correia, C.N.; Nalpas, N.C.; McLoughlin, K.E.; Browne, J.A.; Gordon, S.V.; MacHugh, D.E.; Shaughnessy, R.G. Circulating microRNAs as potential biomarkers of infectious disease. Front. Immunol. 2017, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alamdari-palangi, V.; Vahedi, F.; Shabaninejad, Z.; Dokeneheifard, S.; Movehedpour, A.; Taheri-Anganeh, M.; Savardashtaki, A. microRNA in inflammatory bowel disease at a glance. Eur. J. Gastroenterol. Hepatol. 2020, 1–9. [Google Scholar] [CrossRef]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A.O.; Landthaler, M.; et al. A Mammalian microRNA Expression Atlas Based on Small RNA Library Sequencing. Cell 2007, 129, 1401–1414. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Zhang, Y.; Zhou, D.; Chen, G.; Li, N. MicroRNAs, intestinal inflammatory and tumor. Bioorg. Med. Chem. Lett. 2019, 29, 2051–2058. [Google Scholar] [CrossRef]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tölle, A.; Jung, M.; Rabenhorst, S.; Kilic, E.; Jung, K.; Weikert, S. Identification of microRNAs in blood and urine as tumour markers for the detection of urinary bladder cancer. Oncol. Rep. 2013, 30, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shachar, S.; Yanai, H.; Horev, H.S.; Elad, H.; Baram, L.; Issakov, O.; Tulchinsky, H.; Pasmanik-Chor, M.; Shomron, N.; Dotan, I. MicroRNAs expression in the ileal pouch of patients with ulcerative colitis is robustly up-regulated and correlates with disease phenotypes. PLoS ONE 2016, 11, e159956. [Google Scholar] [CrossRef]
- Thorlacius-Ussing, G.; Schnack Nielsen, B.; Andersen, V.; Holmstrøm, K.; Pedersen, A.E. Expression and Localization of miR-21 and miR-126 in Mucosal Tissue from Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2017, 23, 739–752. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Ma, Y.; Shi, C.; Chen, H.; Zhang, H.; Chen, N.; Zhang, P.; Wang, F.; Yang, J.; Yang, J.; et al. Overexpression of miR-21 in patients with ulcerative colitis impairs intestinal epithelial barrier function through targeting the Rho GTPase RhoB. Biochem. Biophys. Res. Commun. 2013, 434, 746–752. [Google Scholar] [CrossRef]
- Nagy, Z.B.; Barták, B.K.; Kalmár, A.; Galamb, O.; Wichmann, B.; Dank, M.; Igaz, P.; Tulassay, Z.; Molnár, B. Comparison of Circulating miRNAs Expression Alterations in Matched Tissue and Plasma Samples During Colorectal Cancer Progression. Pathol. Oncol. Res. 2019, 25, 97–105. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—MicroRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Secreto, C.R.; Knox, T.R.; Ding, W.; Mukhopadhyay, D.; Kay, N.E. Circulating microvesicles in B-cell chronic lymphocytic leukemia can stimulate marrow stromal cells: Implications for disease progression. Blood 2010, 115, 1755–1764. [Google Scholar] [CrossRef] [Green Version]
- Lima, L.G.; Chammas, R.; Monteiro, R.Q.; Moreira, M.E.C.; Barcinski, M.A. Tumor-derived microvesicles modulate the establishment of metastatic melanoma in a phosphatidylserine-dependent manner. Cancer Lett. 2009, 283, 168–175. [Google Scholar] [CrossRef]
- Waters, P.S.; McDermott, A.M.; Wall, D.; Heneghan, H.M.; Miller, N.; Newell, J.; Kerin, M.J.; Dwyer, R.M. Relationship between Circulating and Tissue microRNAs in a Murine Model of Breast Cancer. PLoS ONE 2012, 7, e50459. [Google Scholar] [CrossRef] [PubMed]
- Iborra, M.; Bernuzzi, F.; Correale, C.; Vetrano, S.; Fiorino, G.; Beltrán, B.; Marabita, F.; Locati, M.; Spinelli, A.; Nos, P.; et al. Identification of serum and tissue micro-RNA expression profiles in different stages of inflammatory bowel disease. Clin. Exp. Immunol. 2013, 173, 250–258. [Google Scholar] [CrossRef]
- Kalla, R.; Ventham, N.T.; Kennedy, N.A.; Quintana, J.F.; Nimmo, E.R.; Buck, A.H.; Satsangi, J. MicroRNAs: New players in IBD. Gut 2015, 64, 504–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Zikusoka, M.; Trindade, A.; Dassopoulos, T.; Harris, M.L.; Bayless, T.M.; Brant, S.R.; Chakravarti, S.; Kwon, J.H. MicroRNAs Are Differentially Expressed in Ulcerative Colitis and Alter Expression of Macrophage Inflammatory Peptide-2α. Gastroenterology 2008, 135, 1624–1635.e24. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Kelly, O.B.; Smith, M.I.; Kabakchiev, B.; Silverberg, M.S. Differential miRNA expression in ileal and colonic tissues reveals an altered immunoregulatory molecular profile in individuals with Crohn’s disease versus healthy subjects. J. Crohn’s Colitis 2019, 13, 1459–1469. [Google Scholar] [CrossRef]
- Béres, N.J.; Szabó, D.; Kocsis, D.; Szucs, D.; Kiss, Z.; Müller, K.E.; Lendvai, G.; Kiss, A.; Arató, A.; Sziksz, E.; et al. Role of Altered Expression of MIR-146a, MIR-155, and MIR-122 in Pediatric Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2016. [Google Scholar] [CrossRef] [Green Version]
- Schönauen, K.; Le, N.; Von Arnim, U.; Schulz, C.; Malfertheiner, P.; Link, A. Circulating and Fecal microRNAs as Biomarkers for Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2018, 24, 1547–1557. [Google Scholar] [CrossRef] [PubMed]
- Valmiki, S.; Ahuja, V.; Paul, J. MicroRNA exhibit altered expression in the inflamed colonic mucosa of ulcerative colitis patients. World J. Gastroenterol. 2017, 23, 5324. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Tsao, C. Emerging role of microRNA-21 in cancer (Review). Biomed. Rep. 2016, 5, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.-X.; Ren, L.-H.; Shi, R.-H. Implication of miRNAs for inflammatory bowel disease treatment: Systematic review. World J. Gastrointest. Pathophysiol. 2014, 5, 63. [Google Scholar] [CrossRef]
- Pathak, S.; Grillo, A.R.; Scarpa, M.; Brun, P.; D’Incà, R.; Nai, L.; Banerjee, A.; Cavallo, D.; Barzon, L.; Palù, G.; et al. MiR-155 modulates the inflammatory phenotype of intestinal myofibroblasts by targeting SOCS1 in ulcerative colitis. Exp. Mol. Med. 2015, 47, e164. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-Y.; Kwon, H.Y.; Ha Thi, H.T.; Lee, H.J.; Kim, G.Il.; Hahm, K.-B.; Hong, S. MicroRNA-132 and microRNA-223 control positive feedback circuit by regulating FOXO3a in inflammatory bowel disease. J. Gastroenterol. Hepatol. 2016, 31, 1727–1735. [Google Scholar] [CrossRef] [PubMed]
- Pierdomenico, M.; Cesi, V.; Cucchiara, S.; Vitali, R.; Prete, E.; Costanzo, M.; Aloi, M.; Oliva, S.; Stronati, L. NOD2 Is Regulated By Mir-320 in Physiological Conditions but this Control Is Altered in Inflamed Tissues of Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2016, 22, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Moein, S.; Vaghari-Tabari, M.; Qujeq, D.; Majidinia, M.; Nabavi, S.M.; Yousefi, B. MiRNAs and inflammatory bowel disease: An interesting new story. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef]
- Tian, Y.; Xu, J.; Li, Y.; Zhao, R.; Du, S.; Lv, C.; Wu, W.; Liu, R.; Sheng, X.; Song, Y.; et al. MicroRNA-31 Reduces Inflammatory Signaling and Promotes Regeneration in Colon Epithelium, and Delivery of Mimics in Microspheres Reduces Colitis in Mice. Gastroenterology 2019, 156, 2281–2296.e6. [Google Scholar] [CrossRef]
- Lu, Z.J.; Wu, J.J.; Jiang, W.L.; Xiao, J.H.; Tao, K.Z.; Ma, L.; Zheng, P.; Wan, R.; Wang, X.P. MicroRNA-155 promotes the pathogenesis of experimental colitis by repressing SHIP-1 expression. World J. Gastroenterol. 2017, 23, 976–985. [Google Scholar] [CrossRef]
- Oliveira, D.N.P.; Carlsen, A.L.; Heegaard, N.H.H.; Prahm, K.P.; Christensen, I.J.; Høgdall, C.K.; Høgdall, E.V. Diagnostic plasma miRNA-profiles for ovarian cancer in patients with pelvic mass. PLoS ONE 2019, 14, e225249. [Google Scholar] [CrossRef]
- Thakral, S.; Ghoshal, K. miR-122 is a unique molecule with great potential in diagnosis, prognosis of liver disease, and therapy both as miRNA mimic and antimir. Curr. Gene Ther. 2015, 15, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Lin, Y.; Zhan, M.; Mann, D.L.; Stass, S.A.; Jiang, F. Differential miRNA expressions in peripheral blood mononuclear cells for diagnosis of lung cancer. HHS Public Interes. 2016, 95, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Coskun, M.; Bjerrum, J.T.; Seidelin, J.B.; Nielsen, O.H. MicroRNAs in inflammatory bowel disease—Pathogenesis, diagnostics and therapeutics. World J. Gastroenterol. 2012, 18, 4629–4634. [Google Scholar] [CrossRef]
- El-Khoury, V.; Pierson, S.; Kaoma, T.; Bernardin, F.; Berchem, G. Assessing cellular and circulating miRNA recovery: The impact of the RNA isolation method and the quantity of input material. Sci. Rep. 2015, 6, 19529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Cao, D.L.; Zhao, L.X.; Han, Y.; Zhang, Y.L. MicroRNA-146a-5p attenuates visceral hypersensitivity through targeting chemokine CCL8 in the spinal cord in a mouse model of colitis. Brain Res. Bull. 2018, 139, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, S.; Qiu, Y.; He, Y.; Chen, B.; Mao, R.; Cui, Y.; Zeng, Z.; Chen, M. Upregulation of miR-665 promotes apoptosis and colitis in inflammatory bowel disease by repressing the endoplasmic reticulum stress components XBP1 and ORMDL3. Cell Death Dis. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijhuis, A.; Curciarello, R.; Mehta, S.; Feakins, R.; Bishop, C.L.; Lindsay, J.O.; Silver, A. MCL-1 is modulated in Crohn’s disease fibrosis by miR-29b via IL-6 and IL-8. Cell Tissue Res. 2017, 368, 325–335. [Google Scholar] [CrossRef] [Green Version]
- Zidar, N.; Boštjančič, E.; Jerala, M.; Kojc, N.; Drobne, D.; Štabuc, B.; Glavač, D. Down-regulation of microRNAs of the miR-200 family and up-regulation of Snail and Slug in inflammatory bowel diseases—Hallmark of epithelial−mesenchymal transition. J. Cell. Mol. Med. 2016, 20, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Chen, S.; Hu, W. MicroRNA-141 Is Involved in Ulcerative Colitis Pathogenesis via Aiming at CXCL5. J. Interferon Cytokine Res. 2017, 37, 415–420. [Google Scholar] [CrossRef]
- Koukos, G.; Polytarchou, C.; Kaplan, J.L.; Morley-Fletcher, A.; Gras-Miralles, B.; Kokkotou, E.; Baril-Dore, M.; Pothoulakis, C.; Winter, H.S.; Iliopoulos, D. MicroRNA-124 regulates STAT3 expression and is down-regulated in colon tissues of pediatric patients with ulcerative colitis. Gastroenterology 2013, 145, 842. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Zhang, X.; Su, J.; Zhang, Y.; Zhou, W.; Zhou, J.; Wang, C.; Liang, H.; Chen, X.; Shi, R.; et al. MiR-19b downregulates intestinal SOCS3 to reduce intestinal inflammation in Crohn’s disease. Sci. Rep. 2015, 5, 10397. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Luo, Y.; Cong, Z.; Mu, Y.; Qiu, Y.; Zhong, M. MicroRNA-590-5p Inhibits Intestinal Inflammation by Targeting YAP. J. Crohn’s Colitis 2018, 12, 993–1004. [Google Scholar] [CrossRef]
- Bai, J.; Yu, J.; Wang, J.; Xue, B.; He, N.; Tian, Y.; Yang, L.; Wang, Y.; Wang, Y.; Tang, Q. DNA Methylation of miR-122 Aggravates Oxidative Stress in Colitis Targeting SELENBP1 Partially by p65NF-κB Signaling. Oxidative Med. Cell. Longev. 2019, 2019, 5294105. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; He, C.; Liu, C.; Cao, A.T.; Xue, X.; Evans-Marin, H.L.; Sun, M.; Fang, L.; Yao, S.; Pinchuk, I.V.; et al. miR-10a inhibits dendritic cell activation and Th1/Th17 cell immune responses in IBD. Gut 2015, 64, 1755–1764. [Google Scholar] [CrossRef]
- Tang, W.-J.; Peng, K.-Y.; Tang, Z.-F.; Wang, Y.-H.; Xue, A.-J.; Huang, Y. MicroRNA-15a—Cell division cycle 42 signaling pathway in pathogenesis of pediatric inflammatory bowel disease. World J. Gastroenterol. 2018, 24, 5234–5245. [Google Scholar] [CrossRef] [PubMed]
- Szcs, D.; Béres, N.J.; Rokonay, R.; Boros, K.; Borka, K.; Kiss, Z.; Arató, A.; Szabó, A.J.; Vannay, Á.; Sziksz, E.; et al. Increased duodenal expression of miR-146a and -155 in pediatric Crohn’s disease. World J. Gastroenterol. 2016, 22, 6027–6035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Li, Y.; Li, L.; Yu, Q.; Chao, K.; Zhou, G.; Qiu, Y.; Feng, R.; Huang, S.; He, Y.; et al. Circulating microRNA146b-5p is superior to C-reactive protein as a novel biomarker for monitoring inflammatory bowel disease. Aliment. Pharmacol. Ther. 2019, 49, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; He, Q.; Guo, Z.; Xiong, F.; Li, Y.; Pan, Y.; Gao, C.; Li, L.; He, C. MicroRNA-425 facilitates pathogenic Th17 cell differentiation by targeting forkhead box O1 (Foxo1) and is associated with inflammatory bowel disease. Biochem. Biophys. Res. Commun. 2018, 496, 352–358. [Google Scholar] [CrossRef]
- He, C.; Shi, Y.; Wu, R.; Sun, M.; Fang, L.; Wu, W.; Liu, C.; Tang, M.; Li, Z.; Wang, P.; et al. MIR-301a promotes intestinal mucosal inflammation through induction of IL-17A and TNF-α in IBD. Gut 2016, 65, 1938–1950. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Chen, D.; Zheng, R.H.; Zhang, H.; Chen, Y.P.; Zun, X. MiRNA-133a-UCP2 pathway regulates inflammatory bowel disease progress by influencing inflammation, oxidative stress and energy metabolism. World J. Gastroenterol. 2017, 23, 76–86. [Google Scholar] [CrossRef]
- Coskun, M. miR-20b, miR-98, miR-125b-1*, and let-7e* as new potential diagnostic biomarkers in ulcerative colitis. World J. Gastroenterol. 2013, 19, 4289. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Ge, W.; Ma, Y.; Xie, G.; Wang, W.; Han, L.; Bian, B.; Li, L.; Shen, L. miR-155 regulates IL-10-producing CD24hiCD27+ B cells and impairs their function in patients with Crohn’s disease. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Fang, K.; Law, I.K.M.; Padua, D.; Sideri, A.; Huang, V.; Kevil, C.G.; Iliopoulos, D.; Pothoulakis, C. MicroRNA-31-3p Is Involved in Substance P (SP)-Associated Inflammation in Human Colonic Epithelial Cells and Experimental Colitis. Am. J. Pathol. 2018, 188, 586–599. [Google Scholar] [CrossRef] [Green Version]
- Takagi, T.; Naito, Y.; Mizushima, K.; Hirata, I.; Yagi, N.; Tomatsuri, N.; Ando, T.; Oyamada, Y.; Isozaki, Y.; Hongo, H.; et al. Increased expression of microRNA in the inflamed colonic mucosa of patients with active ulcerative colitis. J. Gastroenterol. Hepatol. (Aust.) 2010, 25, S129–S133. [Google Scholar] [CrossRef]
- Zhang, H.; Li, W. MicroRNA-15 Activates NF-κB pathway via down regulating expression of adenosine A2 receptor in ulcerative colitis. Cell. Physiol. Biochem. 2018, 51, 1932–1944. [Google Scholar] [CrossRef] [PubMed]
- Pekow, J.R.; Dougherty, U.; Mustafi, R.; Zhu, H.; Kocherginsky, M.; Rubin, D.T.; Hanauer, S.B.; Hart, J.; Chang, E.B.; Fichera, A.; et al. MiR-143 and miR-145 are downregulated in ulcerative colitis: Putative regulators of inflammation and protooncogenes. Inflamm. Bowel Dis. 2012, 18, 94–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minacapelli, C.D.; Bajpai, M.; Geng, X.; Van Gurp, J.; Poplin, E.; Amenta, P.S.; Brant, S.R.; Das, K.M. miR-206 as a Biomarker for Response to Mesalamine Treatment in Ulcerative Colitis. Inflamm. Bowel Dis. 2019, 25. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Chen, X.; Chen, Q.; Shi, L.; Liang, H.; Zhou, Z.; Liu, Q.; Pang, W.; Hou, D.; Wang, C.; et al. MicroRNA-193a-3p reduces intestinal inflammation in response to microbiota via down-regulation of colonic PepT1. J. Biol. Chem. 2015, 290, 16099–16115. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; She, S.; Li, D.; Liu, Z.; Yang, X.; Zeng, Z.; Liu, F. Role of miR-19a targeting TNF-α in mediating ulcerative colitis. Scand. J. Gastroenterol. 2013, 48, 815–824. [Google Scholar] [CrossRef]
- Lu, X.; Yu, Y.; Tan, S. The role of the miR-21-5p-mediated inflammatory pathway in ulcerative colitis. Exp. Ther. Med. 2019, 19, 981–989. [Google Scholar] [CrossRef]
- Chen, Y.; Ge, W.; Xu, L.; Qu, C.; Zhu, M.; Zhang, W.; Xiao, Y. miR-200b is involved in intestinal fibrosis of Crohn’s disease. Int. J. Mol. Med. 2012, 29, 601–606. [Google Scholar] [CrossRef] [Green Version]
- Olaru, A.V.; Selaru, F.M.; Mori, Y.; Vazquez, C.; David, S.; Paun, B.; Cheng, Y.; Jin, Z.; Yang, J.; Agarwal, R.; et al. Dynamic changes in the expression of MicroRNA-31 during inflammatory bowel disease-associated neoplastic transformation. Inflamm. Bowel Dis. 2011, 17, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Bian, Z.; Li, L.; Cui, J.; Zhang, H.; Liu, Y.; Zhang, C.-Y.; Zen, K. Role of miR-150-targeting c-Myb in colonic epithelial disruption during dextran sulphate sodium-induced murine experimental colitis and human ulcerative colitis. J. Pathol. 2011, 225, 544–553. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, C.; Liu, Y.; Tang, L.; Zheng, M.; Xu, C.; Song, J.; Meng, X. MiR-122 targets NOD2 to decrease intestinal epithelial cell injury in Crohn’s disease. Biochem. Biophys. Res. Commun. 2013, 438, 133–139. [Google Scholar] [CrossRef]
- Huang, Z.; Shi, T.; Zhou, Q.; Shi, S.; Zhao, R.; Shi, H.; Dong, L.; Zhang, C.; Zeng, K.; Chen, J.; et al. MIR-141 regulates colonic leukocytic trafficking by targeting CXCL12β during murine colitis and human crohn’s disease. Gut 2014, 63, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.T.; Dalmasso, G.; Yan, Y.; Laroui, H.; Dahan, S.; Mayer, L.; Sitaraman, S.V.; Merlin, D. MicroRNA-7 modulates CD98 expression during intestinal epithelial cell differentiation. J. Biol. Chem. 2010, 285, 1479–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nata, T.; Fujiya, M.; Ueno, N.; Moriichi, K.; Konishi, H.; Tanabe, H.; Ohtake, T.; Ikuta, K.; Kohgo, Y. MicroRNA-146b improves intestinal injury in mouse colitis by activating nuclear factor-κB and improving epithelial barrier function. J. Gene Med. 2013, 15, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Liang, Y.; Yang, J.; Xia, Y.; Chen, H.; Han, H.; Yang, Y.; Wu, W.; Gao, R.; Qin, H. MicroRNA-21 Knockout Improve the Survival Rate in DSS Induced Fatal Colitis through Protecting against Inflammation and Tissue Injury. PLoS ONE 2013, 8, e66814. [Google Scholar] [CrossRef] [Green Version]
- Pekow, J.; Meckel, K.; Dougherty, U.; Haider, H.I.; Deng, Z.; Hart, J.; Rubin, D.T.; Bissonnette, M. Increased mucosal expression of miR-215 precedes the development of neoplasia in patients with long-standing ulcerative colitis. Oncotarget 2018, 9, 20709–20720. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Dong, Y.W.; Song, Y.N.; Xiao, J.H.; Guo, X.Y.; Jiang, W.L.; Lu, L.G. MicroRNA-449a is a potential predictor of colitis-associated colorectal cancer progression. Oncol. Rep. 2018, 40, 1684–1694. [Google Scholar] [CrossRef] [Green Version]
- Lou, C.; Li, Y. Functional role of microRNA-135a in colitis. J. Inflamm. 2018, 15, 7. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Li, Y.; Shao, T.; Zhao, Z.; Wang, Y.; Wu, A.; Chen, H.; Li, S.; Jiang, C.; Xu, J.; et al. Integrating analysis reveals microRNA-mediated pathway crosstalk among Crohn’s disease, ulcerative colitis and colorectal cancer. Mol. Biosyst. 2014, 10, 2317–2328. [Google Scholar] [CrossRef]
- Li, W.; Han, W.; Zhao, X.; Wang, H. [Changes of expression of miR-155 in colitis-associated colonic carcinogenesis]. Zhonghua Zhong Liu Za Zhi 2014, 36, 257–262. [Google Scholar]
- Eichele, D.D.; Kharbanda, K.K. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef]
- Keubler, L.M.; Buettner, M.; Häger, C.; Bleich, A. A multihit model: Colitis lessons from the interleukin-10-deficient mouse. Inflamm. Bowel Dis. 2015, 21, 1967–1975. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, E.; Kanneganti, M.; Mino-Kenudson, M. Animal models of colitis-associated carcinogenesis. J. Biomed. Biotechnol. 2011, 2011. [Google Scholar] [CrossRef] [Green Version]
- Mizoguchi, A.; Mizoguchi, E. Animal models of IBD: Linkage to human disease. Curr. Opin. Pharmacol. 2010, 10, 578–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindi, M.; Riddell, R.H. Indeterminate colitis. J. Clin. Pathol. 2004, 57, 1233–1244. [Google Scholar] [CrossRef]
- Tontini, G.E.; Vecchi, M.; Pastorelli, L.; Neurath, M.F.; Neumann, H. Differential diagnosis in inflammatory bowel disease colitis: State of the art and future perspectives. World J. Gastroenterol. 2015, 21, 21–46. [Google Scholar] [CrossRef]
- Birimberg-Schwartz, L.; Zucker, D.M.; Akriv, A.; Cucchiara, S.; Cameron, F.L.; Wilson, D.C.; Łazowska, I.; Yianni, L.; Paul, S.P.; Romano, C.; et al. Development and validation of diagnostic criteria for IBD subtypes including IBdunclassified in children: A multicentre study from the pediatric IBD porto group of ESPGHAN. J. Crohn’s Colitis 2017, 1078–1084. [Google Scholar] [CrossRef]
- Schaefer, J.S.; Attumi, T.; Opekun, A.R.; Abraham, B.; Hou, J.; Shelby, H.; Graham, D.Y.; Streckfus, C.; Klein, J.R. MicroRNA signatures differentiate Crohn’s disease from ulcerative colitis. BMC Immunol. 2015, 16, 5. [Google Scholar] [CrossRef] [Green Version]
- Peck, B.C.E.; Weiser, M.; Lee, S.E.; Gipson, G.R.; Iyer, V.B.; Sartor, R.B.; Herfarth, H.H.; Long, M.D.; Hansen, J.J.; Isaacs, K.L.; et al. MicroRNAs classify different disease behavior phenotypes of Crohn’s disease and may have prognostic utility. Inflamm. Bowel Dis. 2015, 21, 2178–2187. [Google Scholar] [CrossRef] [Green Version]
- Kantono, M.; Guo, B. Inflammasomes and Cancer: The Dynamic Role of the Inflammasome in Tumor Development. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Khalafalla, F.G.; Khan, M.W. Inflammation and Epithelial-Mesenchymal Transition in Pancreatic Ductal Adenocarcinoma: Fighting Against Multiple Opponents. Cancer Growth Metastasis 2017, 10. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalapour, S.; Karin, M. Cruel to Be Kind: Epithelial, Microbial, and Immune Cell Interactions in Gastrointestinal Cancers. Annu. Rev. Immunol. 2020, 38, 649–671. [Google Scholar] [CrossRef] [PubMed]
- Todoric, J.; Karin, M. The Fire within: Cell-Autonomous Mechanisms in Inflammation-Driven Cancer. Cancer Cell 2019, 35, 714–720. [Google Scholar] [CrossRef] [PubMed]
- Malham, M.; Jakobsen, C.; Paerregaard, A.; Virta, L.J.; Kolho, K.L.; Wewer, V. The incidence of cancer and mortality in paediatric onset inflammatory bowel disease in Denmark and Finland during a 23-year period: A population-based study. Aliment. Pharmacol. Ther. 2019, 50, 33–39. [Google Scholar] [CrossRef]
- Chen, R.; Lai, L.A.; Brentnall, T.A.; Pan, S. Biomarkers for colitis-associated colorectal cancer. World J. Gastroenterol. 2016, 22, 7882–7891. [Google Scholar] [CrossRef]
- Duricova, D.; Pedersen, N.; Elkjaer, M.; Gamborg, M.; Munkholm, P.; Jess, T. Overall and cause-specific mortality in Crohn’s disease: A meta-analysis of population-based studies. Inflamm. Bowel Dis. 2010, 16, 347–353. [Google Scholar] [CrossRef]
- Jess, T.; Gamborg, M.; Matzen, P.; Munkholm, P.; Sorensen, T.I. Increased Risk of Intestinal Cancer in Crohn’s Disease: A Meta-Analysis of Population-Based Cohort Studies. Am. J. Gastroenterol. 2005, 100, 2724–2729. [Google Scholar] [CrossRef]
- Foersch, S.; Neurath, M.F. Colitis-associated neoplasia: Molecular basis and clinical translation. Cell. Mol. Life Sci. 2014, 71, 3523–3535. [Google Scholar] [CrossRef]
- Romano, M.; De Francesco, F.; Zarantonello, L.; Ruffolo, C.; Ferraro, G.A.; Zanus, G.; Giordano, A.; Bassi, N.; Cillo, U. From inflammation to cancer in inflammatory bowel disease: Molecular perspectives. Anticancer Res. 2016, 36, 1447–1460. [Google Scholar]
- Galandiuk, S.; Rodriguezjusto, M.; Jeffery, R.; Nicholson, A.M.; Cheng, Y.; Oukrif, D.; Elia, G.; Leedham, S.J.; McDonald, S.A.C.; Wright, N.A.; et al. Field cancerization in the intestinal epithelium of patients with Crohn’s ileocolitis. Gastroenterology 2012, 142, 855. [Google Scholar] [CrossRef] [Green Version]
- Risques, R.A.; Lai, L.A.; Himmetoglu, C.; Ebaee, A.; Li, L.; Feng, Z.; Bronner, M.P.; Al-Lahham, B.; Kowdley, K.V.; Lindor, K.D.; et al. Ulcerative colitis-associated colorectal cancer arises in a field of short telomeres, senescence, and inflammation. Cancer Res. 2011, 71, 1669–1679. [Google Scholar] [CrossRef] [Green Version]
- Karvellas, C.J.; Fedorak, R.N.; Hanson, J.; Wong, C.K.W. Increased risk of colorectal cancer in ulcerative colitis patients diagnosed after 40 years of age. Can. J. Gastroenterol. 2007, 21, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Den Low, E.N.; Mokhtar, N.M.; Wong, Z.; Raja Ali, R.A. Colonic mucosal transcriptomic changes in patients with long-duration ulcerative colitis revealed colitis-associated cancer pathways. J. Crohn’s Colitis 2019, 13, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, T.M.; Brückner, M.; Eveslage, M.; Tepasse, P.; Pott, F.; Thoennissen, N.H.; Hengst, K.; Ross, M.; Bettenworth, D. The Risk of Colorectal Cancer in Patients with Ulcerative Colitis. Dig. Dis. Sci. 2015, 60, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Josse, C.; Bours, V. MicroRNAs and inflammation in colorectal cancer. Adv. Exp. Med. Biol. 2016, 937, 53–69. [Google Scholar] [CrossRef]
- Ranjha, R.; Aggarwal, S.; Bopanna, S.; Ahuja, V.; Paul, J. Site-specific MicroRNA expression may lead to different subtypes in ulcerative colitis. PLoS ONE 2015, 10, e0142869. [Google Scholar] [CrossRef]
- Itzkowitz, S.H.; Yio, X. Inflammation and cancer—IV. Colorectal cancer in inflammatory bowel disease: The role of inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G7–G17. [Google Scholar] [CrossRef]
- He, C.; Yu, T.; Shi, Y.; Ma, C.; Yang, W.; Fang, L.; Sun, M.; Wu, W.; Xiao, F.; Guo, F.; et al. MicroRNA 301A Promotes Intestinal Inflammation and Colitis-Associated Cancer Development by Inhibiting BTG1. Gastroenterology 2017, 152, 1434–1448.e15. [Google Scholar] [CrossRef]
- Signs, S.A.; Fisher, R.C.; Tran, U.; Chakrabarti, S.; Sarvestani, S.K.; Xiang, S.; Liska, D.; Roche, V.; Lai, W.; Gittleman, H.R.; et al. Stromal miR-20a controls paracrine CXCL8 secretion in colitis and colon cancer. Oncotarget 2018, 9, 13048–13059. [Google Scholar] [CrossRef] [Green Version]
- Krichevsky, A.M.; Gabriely, G. miR-21: A small multi-faceted RNA. J. Cell. Mol. Med. 2008, 13, 39–53. [Google Scholar] [CrossRef]
- Bautista-Sánchez, D.; Arriaga-Canon, C.; Pedroza-Torres, A.; De La Rosa-Velázquez, I.A.; González-Barrios, R.; Contreras-Espinosa, L.; Montiel-Manríquez, R.; Castro-Hernández, C.; Fragoso-Ontiveros, V.; Álvarez-Gómez, R.M.; et al. The Promising Role of miR-21 as a Cancer Biomarker and Its Importance in RNA-Based Therapeutics. Mol. Ther. Nucleic Acids 2020, 20, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, B.S.; Jørgensen, S.; Fog, J.U.; Søkilde, R.; Christensen, I.J.; Hansen, U.; Brünner, N.; Baker, A.; Møller, S.; Nielsen, H.J. High levels of microRNA-21 in the stroma of colorectal cancers predict short disease-free survival in stage II colon cancer patients. Clin. Exp. Metastasis 2011, 28, 27–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, K.; Fassan, M.; Mescoli, C.; Pizzi, M.; Balistreri, M.; Albertoni, L.; Pucciarelli, S.; Scarpa, M.; Sturniolo, G.C.; Angriman, I.; et al. PDCD4/miR-21 dysregulation in inflammatory bowel disease-associated carcinogenesis. Virchows Arch. 2013, 462, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Frankel, L.B.; Christoffersen, N.R.; Jacobsen, A.; Lindow, M.; Krogh, A.; Lund, A.H. Programmed cell death 4 (PDCD4) is an important functional target of the microRNA miR-21 in breast cancer cells. J. Biol. Chem. 2008, 283, 1026–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, E.C.; Tarnawski, A.S. PTEN regulatory functions in tumor suppression and cell biology. Med. Sci. Monit. 2004, 10, RA235-41. [Google Scholar]
- Cho, W.C.S. OncomiRs: The discovery and progress of microRNAs in cancers. Mol. Cancer 2007, 6, 60. [Google Scholar] [CrossRef] [Green Version]
- Ando, Y.; Yang, G.-X.; Kenny, T.P.; Kawata, K.; Zhang, W.; Huang, W.; Leung, P.S.C.; Lian, Z.-X.; Okazaki, K.; Ansari, A.A.; et al. Overexpression of microRNA-21 is associated with elevated pro- inflammatory cytokines in dominant-negative TGF-β receptor type II mouse. J. Autoimmun. 2013, 41, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Hu, X.; Chen, B.; Chen, X.; Zhao, L.; Chen, Z.; Liu, F.; Liu, Z. miR-155 targets Est-1 and induces ulcerative colitis via the IL-23/17/6-mediated Th17 pathway. Pathol. Res. Pract. 2017, 213, 1289–1295. [Google Scholar] [CrossRef]
- Paraskevi, A.; Theodoropoulos, G.; Papaconstantinou, I.; Mantzaris, G.; Nikiteas, N.; Gazouli, M. Circulating MicroRNA in inflammatory bowel disease. J. Crohn’s Colitis 2012, 6, 900–904. [Google Scholar] [CrossRef]
- Kara, M.; Yumrutas, O.; Ozcan, O.; Celik, O.I.; Bozgeyik, E.; Bozgeyik, I.; Tasdemir, S. Differential expressions of cancer-associated genes and their regulatory miRNAs in colorectal carcinoma. Gene 2015, 567, 81–86. [Google Scholar] [CrossRef]
- Shibuya, H.; Iinuma, H.; Shimada, R.; Horiuchi, A.; Watanabe, T. Clinicopathological and prognostic value of microRNA-21 and microRNA-155 in colorectal cancer. Oncology 2011, 79, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.-c.; Fan, Y.-s.; Chen, H.-b.; Zhao, D.-w. Investigation of microRNA-155 as a serum diagnostic and prognostic biomarker for colorectal cancer. Tumor Biol. 2014, 36, 1619–1625. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, Z.; Xiang, J.; Gu, X. MicroRNA-155 acts as a tumor suppressor in colorectal cancer by targeting CTHRC1 in vitro. Oncol. Lett. 2018, 15, 5561–5568. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, F.; Li, H.; Fan, H.; Wu, H.; Dong, Y.; Chu, S.; Tan, C.; Wang, Q.; He, H.; et al. MiR-155 contributes to intestinal barrier dysfunction in DSS-induced mice colitis via targeting HIF-1α/TFF-3 axis. Aging (Albany N.Y.) 2020, 12, 14966–14977. [Google Scholar] [CrossRef]
- Karimi, N.; Feizi, M.A.H.; Safaralizadeh, R.; Hashemzadeh, S.; Baradaran, B.; Shokouhi, B.; Teimourian, S. Serum overexpression of miR-301a and miR-23a in patients with colorectal cancer. J. Chin. Med. Assoc. 2019, 82, 215–220. [Google Scholar] [CrossRef]
- Loftus, E.V.; Kane, S.V.; Bjorkman, D. Systematic review: Short-term adverse effects of 5-aminosalicylic acid agents in the treatment of ulcerative colitis. Aliment. Pharmacol. Ther. 2004, 19, 179–189. [Google Scholar] [CrossRef]
- Scarpa, M.; Castagliuolo, I.; Castoro, C.; Pozza, A.; Scarpa, M.; Kotsafti, A.; Angriman, I. Inflammatory colonic carcinogenesis: A review on pathogenesis and immunosurveillance mechanisms in ulcerative colitis. World J. Gastroenterol. 2014, 20, 6774–6785. [Google Scholar] [CrossRef]
- Williams, C.; Panaccione, R.; Ghosh, S.; Rioux, K. Optimizing clinical use of mesalazine (5-aminosalicylic acid) in inflammatory bowel disease. Ther. Adv. Gastroenterol. 2011, 4, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Olesen, C.M.; Coskun, M.; Peyrin-Biroulet, L.; Nielsen, O.H. Mechanisms behind efficacy of tumor necrosis factor inhibitors in inflammatory bowel diseases. Pharmacol. Ther. 2016, 159, 110–119. [Google Scholar] [CrossRef]
- Espinosa Morales, R.; Díaz Borjón, A.; Barile Fabris, L.A.; Esquivel Valerio, J.A.; Medrano Ramírez, G.; Arce Salinas, C.A.; Barreira Mercado, E.R.; Cardiel Ríos, M.H.; Díaz Jouanen, E.; Flores Murrieta, F.J.; et al. Long-term infliximab maintenance therapy for ulcerative colitis: The ACT-1 and -2 extension studies. Inflamm. Bowel Dis. 2012. [Google Scholar] [CrossRef]
- Nielsen, O.H.; Ainsworth, M.A. Tumor necrosis factor inhibitors for inflammatory bowel disease. N. Engl. J. Med. 2013, 369, 754–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Horin, S.; Chowers, Y. Review article: Loss of response to anti-TNF treatments in Crohn’s disease. Aliment. Pharmacol. Ther. 2011, 33, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Morilla, I.; Uzzan, M.; Laharie, D.; Cazals-Hatem, D.; Denost, Q.; Daniel, F.; Belleannee, G.; Bouhnik, Y.; Wainrib, G.; Panis, Y.; et al. Colonic MicroRNA Profiles, Identified by a Deep Learning Algorithm, That Predict Responses to Therapy of Patients With Acute Severe Ulcerative Colitis. Clin. Gastroenterol. Hepatol. 2019, 17, 905–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandborn, W.J.; Gasink, C.; Gao, L.L.; Blank, M.A.; Johanns, J.; Guzzo, C.; Sands, B.E.; Hanauer, S.B.; Targan, S.; Rutgeerts, P.; et al. Ustekinumab induction and maintenance therapy in refractory Crohn’s disease. N. Engl. J. Med. 2012, 367, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.J.; Noh, J.H.; Kim, J.K.; Eun, J.W.; Jung, K.H.; Kim, M.G.; Chang, Y.G.; Shen, Q.; Kim, S.-J.; Park, W.S.; et al. MicroRNA-29c functions as a tumor suppressor by direct targeting oncogenic SIRT1 in hepatocellular carcinoma. Oncogene 2014, 33, 2557–2567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedyk, E.R.; Wyant, T.; Yang, L.L.; Csizmadia, V.; Burke, K.; Yang, H.; Kadambi, V.J. Exclusive antagonism of the α4β7 integrin by vedolizumab confirms the gut-selectivity of this pathway in primates. Inflamm. Bowel Dis. 2012, 18, 2107–2119. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.A.; Yamakuchi, M.; Ferlito, M.; Mendell, J.T.; Lowenstein, C.J. MicroRNA-126 regulates endothelial expression of vascular cell adhesion molecule 1. Proc. Natl. Acad. Sci. USA 2008, 105, 1516–1521. [Google Scholar] [CrossRef] [Green Version]
- Sandborn, W.J.; Ghosh, S.; Panes, J.; Vranic, I.; Su, C.; Rousell, S.; Niezychowski, W. Tofacitinib, an Oral Janus Kinase Inhibitor, in Active Ulcerative Colitis. N. Engl. J. Med. 2012, 367, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Magro, F.; Estevinho, M.M. Is tofacitinib a game-changing drug for ulcerative colitis? United Eur. Gastroenterol. J. 2020, 8, 755–763. [Google Scholar] [CrossRef]
- Kim, H.; Banerjee, N.; Sirven, M.A.; Minamoto, Y.; Markel, M.E.; Suchodolski, J.S.; Talcott, S.T.; Mertens-Talcott, S.U. Pomegranate polyphenolics reduce inflammation and ulceration in intestinal colitis—Involvement of the miR-145/p70S6K1/HIF1α axis in vivo and in vitro. J. Nutr. Biochem. 2017, 43, 107–115. [Google Scholar] [CrossRef]
- Maschmeyer, P.; Petkau, G.; Siracusa, F.; Zimmermann, J.; Zügel, F.; Kühl, A.A.; Lehmann, K.; Schimmelpfennig, S.; Weber, M.; Haftmann, C.; et al. Selective targeting of pro-inflammatory Th1 cells by microRNA-148a-specific antagomirs in vivo. J. Autoimmun. 2018, 89, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Whiteoak, S.R.; Claridge, A.; Balendran, C.A.; Harris, R.J.; Gwiggner, M.; Bondanese, V.P.; Erlandsson, F.; Hansen, M.B.; Cummings, J.R.F.; Sanchez-Elsner, T. MicroRNA-31 Targets Thymic Stromal Lymphopoietin in Mucosal Infiltrated CD4+ T Cells: A Role in Achieving Mucosal Healing in Ulcerative Colitis? Inflamm. Bowel Dis. 2018, 24, 2377–2385. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.Q.; Wang, J.; Chen, G.X.; Zhang, G.Q.; Zhang, D.Y.; Cai, Y.Y. Overexpression of microRNA-495 improves the intestinal mucosal barrier function by targeting STAT3 via inhibition of the JAK/STAT3 signaling pathway in a mouse model of ulcerative colitis. Pathol. Res. Pract. 2018, 214, 151–162. [Google Scholar] [CrossRef]
- Jørgensen, S.; Baker, A.; Møller, S.; Nielsen, B.S. Robust one-day in situ hybridization protocol for detection of microRNAs in paraffin samples using LNA probes. Methods 2010, 52, 375–381. [Google Scholar] [CrossRef]
- Nielsen, B.S.; Holmstrøm, K. Target Identification and Validation in Drug Discovery; Moll, J., Colombo, R., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2013; Volume 986, ISBN 978-1-62703-310-7. [Google Scholar]
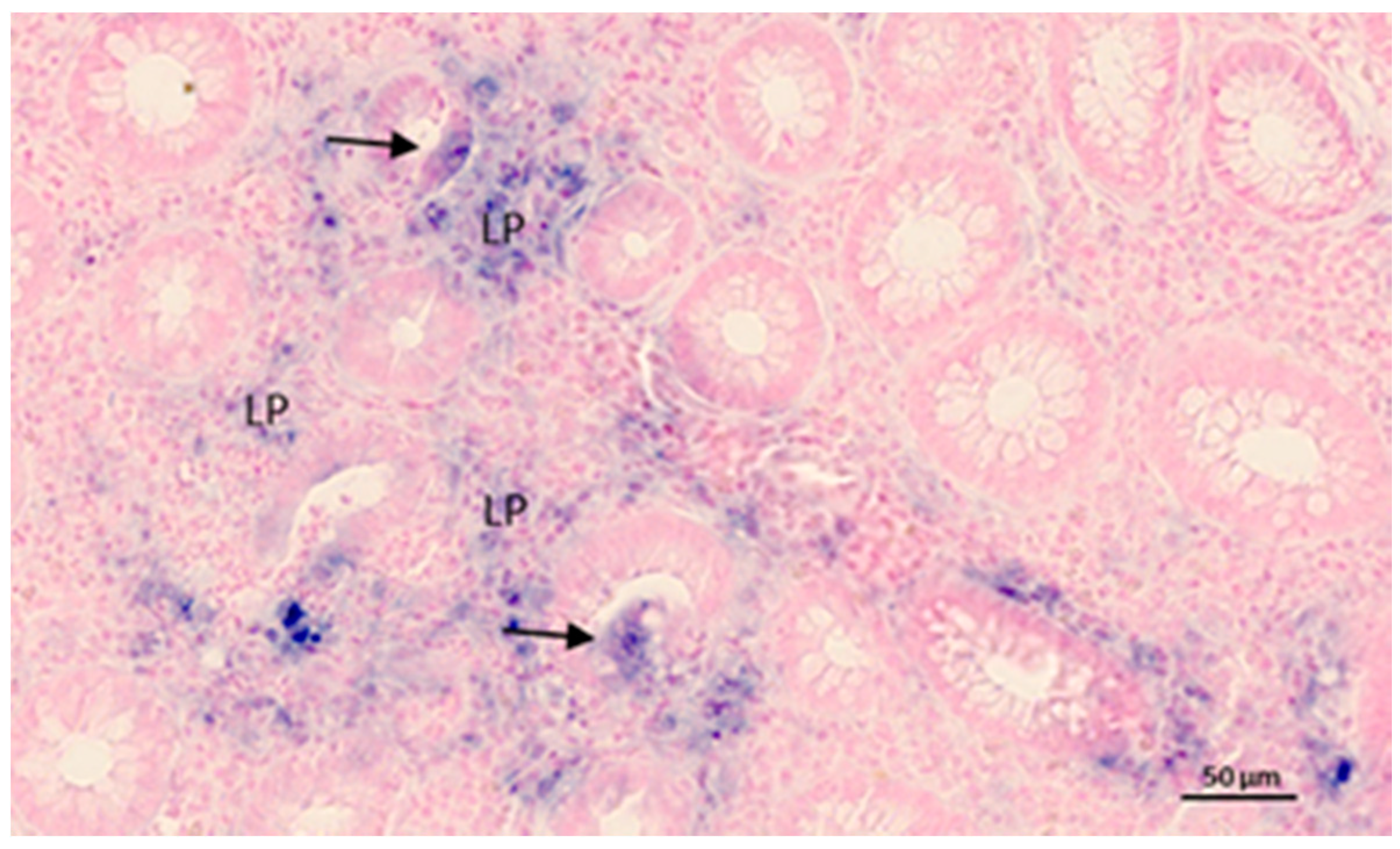

{kind=link}
| # | MiRNAs | Disease Subtype | Sample Type | Techniques Used | Outcome | Reference |
|---|---|---|---|---|---|---|
| 1 | miR-16, miR-29a, miR-199a-5p, miR-363-3p, miR-340, miR-532-3p, miRplus-1271, miR-140-3p, miR-127-3p, miR-196b, miR-877, miR-150 | CD, UD, HC | Serum, Biopsy | RT-qPCR, Microarray | Mixed outcomes | [42] |
| 2 | miR-223-3p, miR-31-5p | CD, HC | Biopsy | Nano string | Mir-223-3p expression showed age and sex effects and miR-31-5p expression was driven by location | [45] |
| 3 | miR-29b | CD | Fibroblasts | RT-qPCR | MCL-1 is modulated in CD fibrosis by miR-29b via IL-6 and IL-8 | [64] |
| 4 | miR-141, miR-200a, miR-200b, miR-200c | UC, CD | Biopsy | RT-qPCR | All investigated miRNAs were significantly down regulated in CD, and 3 of them were downregulated in UC in comparison to the normal or the least affected mucosa. | [65] |
| 5 | miR-141 | UC, HC | Biopsy | Microarray, RT-qPCR | MiR-141 plays a role in the bowel inflammation of individuals with active UC via down regulation of CXCL5 expression. | [66] |
| 6 | miR-124 | UC, HC | Biopsy | RT-qPCR | MiR-124 regulates the expression of STAT3. Reduced levels of miR-124 in colon tissues of children with active UC appear to increase expression and activity of STAT3. | [67] |
| 7 | miR-19b | CD, HC | Biopsy, Cell culture | RT-qPCR, ISH | MiR-19b suppresses the inflammation and prevents the pathogenesis of CD. | [68] |
| 8 | miR-590-5p | CD, HC | Human and mice tissues | RT-qPCR | Decreased miR-590-5p levels in CD. | [69] |
| 9 | miR-122 | CD, HC | Biopsy | RT-qPCR, Sequencing | Significant increase of miR-122 expression in cells treated with 5′-AZA. | [70] |
| 10 | miR-10a | CD, UC, HC | Biopsy | RT-qPCR | Dendritic cell activation and Th1/Th17 cell immune responses were inhibited via miR-10a in IBD. | [71] |
| 11 | miR-192 | CD, UC, HC | Biopsy | RT-qPCR, Microarray, ISH | MiR-192 with decreased expression in active UC. | [44] |
| 12 | miR-15a | CD, UC, HC | Biopsy, Cell cultures | RT-qPCR | MiR-15a negatively regulates epithelial junctions through Cdc42 in Caco-2 cells | [72] |
| 13 | miR-146a, miR-155 | CD | Biopsy | RT-qPCR | MiR-146a and -155 shows increased duodenal expression in pediatric CD. | [73] |
| 14 | miR-146b-5p | CD, UC, HC | Serum | RT-qPCR | Higher expression of serum miR-146b-5p in patients with CD and UC than in HC. | [74] |
| 15 | miR-425 | CD, UC, HC | Biopsy, PBMC | RT-qPCR | Increased expression of miR-425 in IBD. | [75] |
| 16 | miR-301a | IBD | PBMC, Biopsy | RT-qPCR | MiR-301a promotes intestinal mucosal inflammation via induction of IL-17a and TNF in IBD. | [76] |
| 17 | miR-125b, miR-155, miR-223 and miR-138 | UC | Biopsy | RT-qPCR, Microarray | Differential expression of miR-223, miR-125b, miR-138, and miR-155 in the inflamed mucosa compared to non-inflamed mucosa and controls. | [48] |
| 18 | miR-16, miR-21, miR-155, and miR-223 | CD, UC, HC | Serum, Feces | RT-qPCR | Differential expression of miR-16, miR-155, miR-21, and miR-223 in IBD. | [47] |
| 19 | miR-21 | UC, HC | Biopsy | RT-qPCR, ISH | Over expression of miR-21 in UC. | [36] |
| 20 | miR-133a | IBD | Mice Tissue | RT-qPCR | MiR-133a-UCP2 pathway participates in IBD by altering downstream inflammation, oxidative stress, and markers of energy metabolism. | [77] |
| 21 | miR-20b, miR-98, miR-125b-1, let-7e | CD, UC, HC | Biopsy | RT-qPCR, Microarray | MiR-20b, miR-98, miR-125b-1, and let-7e are deregulated in patients with UC. | [78] |
| 22 | miR-155 | CD, HC | PBMC | RT-qPCR, Transfection | MiR-155 regulates IL-10-producing CD24 CD27+ B Cells. | [79] |
| 23 | miR-21, miR-126 | CD, UC, HC | Biopsy | RT-qPCR, qISH | Endothelial expression of miR-126 are increased in UC. MiR-21 is expressed in subsets of monocytes/macrophages and T cells. | [35] |
| 24 | miR-31 | CD, UC, HC | Cell culture, Biopsy | RT-qPCR, ISH, Transfection | Expression of miR-31-3p in human colonic epithelial cells. | [80] |
| 25 | miR-21, miR-155 | UC, HC | Biopsy | RT-qPCR | MiR-21 and miR-155 was highly expressed in UC. | [81] |
| 26 | miR-15 | UC, HC, IBS | Biopsy | RT-qPCR | MiR-15 activates NF-κB Pathway in UC. | [82] |
| 27 | miR-143, miR-145 | UC, HC | Biopsy | RT-qPCR, ISH | MiR-143 and miR-145 are down regulated in UC. | [83] |
| 28 | miR-206 | UC, HC | Cell culture, Biopsy | RT-qPCR, | MiR-206 as a biomarker for response to mesalamine treatment in UC. | [84] |
| 29 | miR-193a-3p | UC, HC | Cell culture, Biopsy | RT-qPCR, ISH | MiR-193a-3p reduces intestinal inflammation in response to microbiota. | [85] |
| 30 | miR-19a | UC, HC | Biopsy, mice tissue | RT-qPCR | Reduced expression of miR-19a in human colon tissue with UC and in DSS-treated mice colitis. | [86] |
| 31 | miR-21-5p | UC, HC | Sera, rat tissue | RT-qPCR, Transfection | MiR-21-5p was down regulated in the sera and colon tissue of UC compared with healthy people and the control group. | [87] |
| 32 | miR-200b | CD, HC | Biopsy, Serum. Cell culture | RT-qPCR | MiR-200b is involved in intestinal fibrosis of CD. | [88] |
| 33 | miR-155 | Colitis | Mice tissue, cell culture | RT-qPCR, Transfection | MiR-155 promotes the pathogenesis of experimental colitis by repressing SHIP-1 expression. | [57] |
| 34 | miR-31 | IBD, CAC, CRC | Biopsy | RT-qPCR, Microarray, Transfection | MiR-31 expression levels as a marker for disease progression and to discriminate distinct pathological entities that co-exist in IBD. | [89] |
| 35 | miR-150 | UC, HC | murine model | RT-qPCR | MiR-150 was elevated and c-Myb were down regulated in human colon with active UC compared to HC. | [90] |
| 36 | miR-122 | CD | Cell culture | RT-qPCR, Transfection | MiR-122 reduces the expression of pro-inflammatory cytokines (TNF and IFN-γ) and promotes the release of anti-inflammatory cytokines (IL-4 and IL-10). | [91] |
| 37 | miR-141 | CD | Murine models, Biopsy | Microarray, RT-qPCR | MiR-141 regulates colonic leukocytic trafficking by targeting CXCL12β during murine colitis and human CD. | [92] |
| 38 | miR-7 | CD, HC | Cell culture, Biopsy | Transfection, RT-qPCR | MiR-7 modulates CD98 expression during intestinal epithelial cell differentiation. | [93] |
| 39 | miR-146b | IBD | IL-10 deficient mouse | Microarray, Transfection, DSS induced colitis in vivo | MiR-146b improves intestinal injury in mouse colitis. | [94] |
| 40 | miR-21 | IBD | IL-10 deficient mouse, Biopsy | DSS-induced Experimental Colitis, RT-qPCR, ISH | MiR-21 is overexpressed in intestinal inflammation and tissue injury. | [95] |
| 41 | miR-215 | UC, CAC | Biopsy | Nano string | MiR-215 discriminates patients who progressed to neoplasia as early as 5 years prior to the diagnosis of neoplasia | [96] |
| 42 | miR-449a | HC, CAC | DSS animal model biopsy | RT-qPCR, ISH | MiR-449a expression decreased gradually during the progression of CAC | [97] |
| 43 | miR-135a | CAC | DSS mouse model biopsy | ISH, RT-qPCR | MiR-135a in colonic cells were suppressed and up-regulating miR-135a inhibited apoptosis and inflammation of colonic epithelial cells | [98] |
| 44 | miR-146a, miR-155, miR-122 | CD, UC, HC | Biopsy | RT-qPCR | Altered expression of all three miRNAs in colonic mucosa of children with IBD | [46] |
| 45 | miR-146a, miR-335, miR-26b and miR-124 | CD, UC, CRC | Genome-wide expression profiles | Bioinformatics | MiR-146a, miR-335, miR-26b and miR-124 were identified in CD, UC, and CRC samples | [99] |
| 46 | miR-155 | CAC, HC | AOM and DSS mouse model biopsy | Microarray, RT-qPCR | MiR-155 is upregulated in and relates to CAC | [100] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
James, J.P.; Riis, L.B.; Malham, M.; Høgdall, E.; Langholz, E.; Nielsen, B.S. MicroRNA Biomarkers in IBD—Differential Diagnosis and Prediction of Colitis-Associated Cancer. Int. J. Mol. Sci. 2020, 21, 7893. https://doi.org/10.3390/ijms21217893
James JP, Riis LB, Malham M, Høgdall E, Langholz E, Nielsen BS. MicroRNA Biomarkers in IBD—Differential Diagnosis and Prediction of Colitis-Associated Cancer. International Journal of Molecular Sciences. 2020; 21(21):7893. https://doi.org/10.3390/ijms21217893
Chicago/Turabian StyleJames, Jaslin P., Lene Buhl Riis, Mikkel Malham, Estrid Høgdall, Ebbe Langholz, and Boye S Nielsen. 2020. "MicroRNA Biomarkers in IBD—Differential Diagnosis and Prediction of Colitis-Associated Cancer" International Journal of Molecular Sciences 21, no. 21: 7893. https://doi.org/10.3390/ijms21217893