Vascular Endothelial Growth Factor A and VEGFR-1 Change during Preimplantation in Heifers

 , and
, and
Abstract
1. Introduction
2. Results
2.1. Study A: Cycling versus Pregnant Heifers
2.1.1. The Concentration of P4 and E2 in the Serum of Pregnant and Nonpregnant Heifers
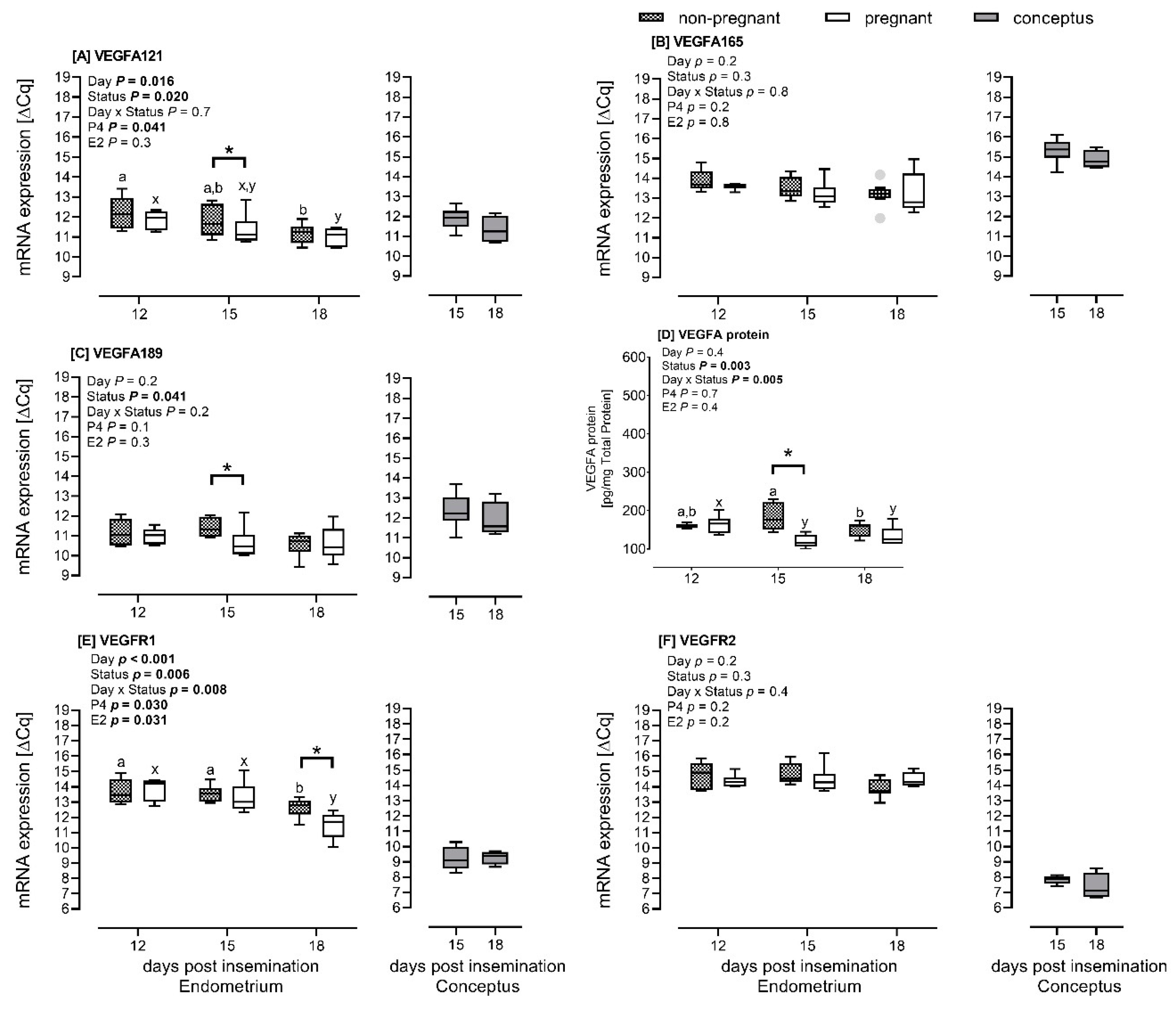
2.1.2. mRNA Expression of VEGFA Isoforms and VEGF Receptors in the Endometrial and Conceptus Tissues
2.1.3. Endometrial VEGFA Protein Abundance
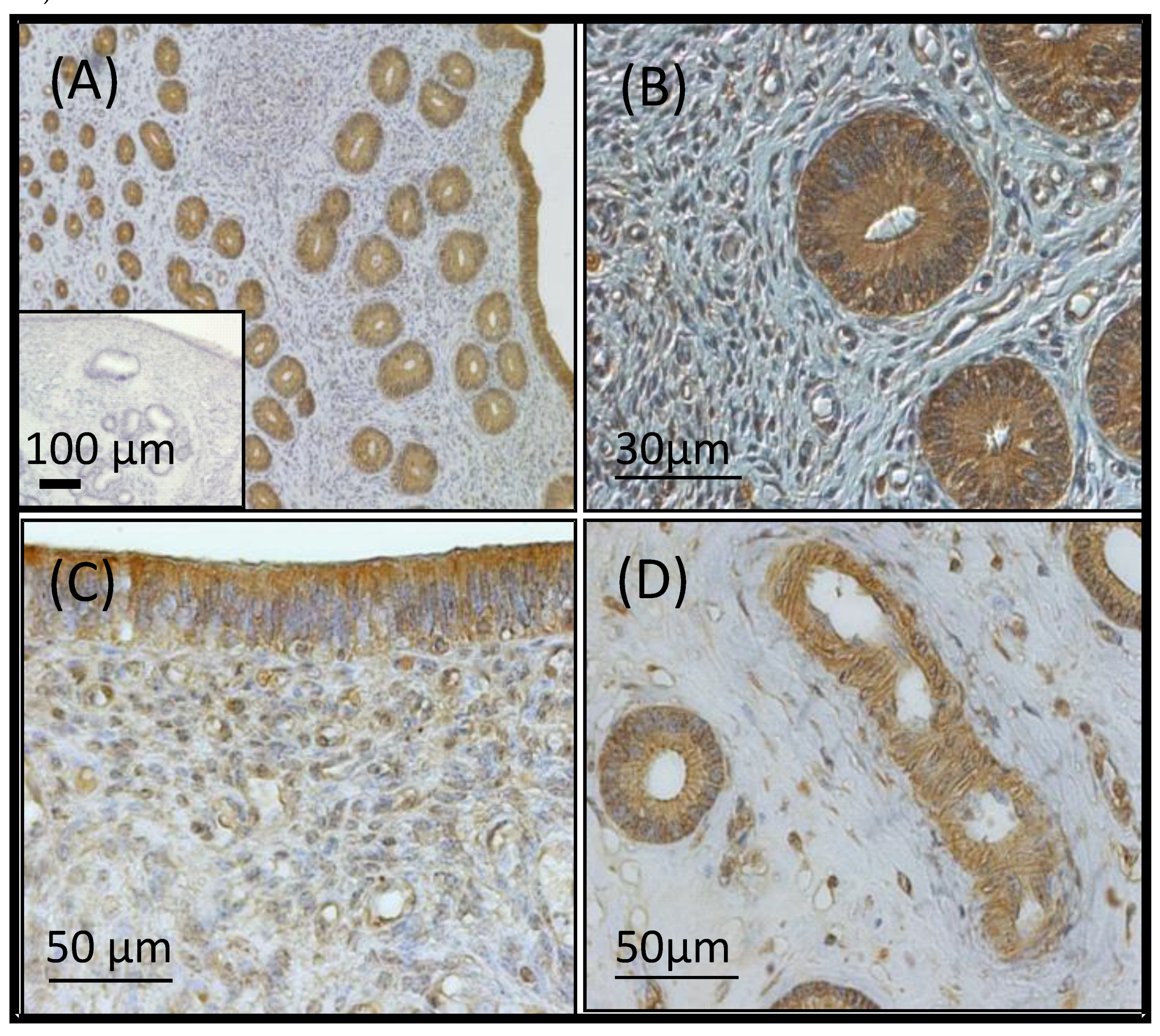
2.1.4. Localisation of VEGFA in the Endometrium
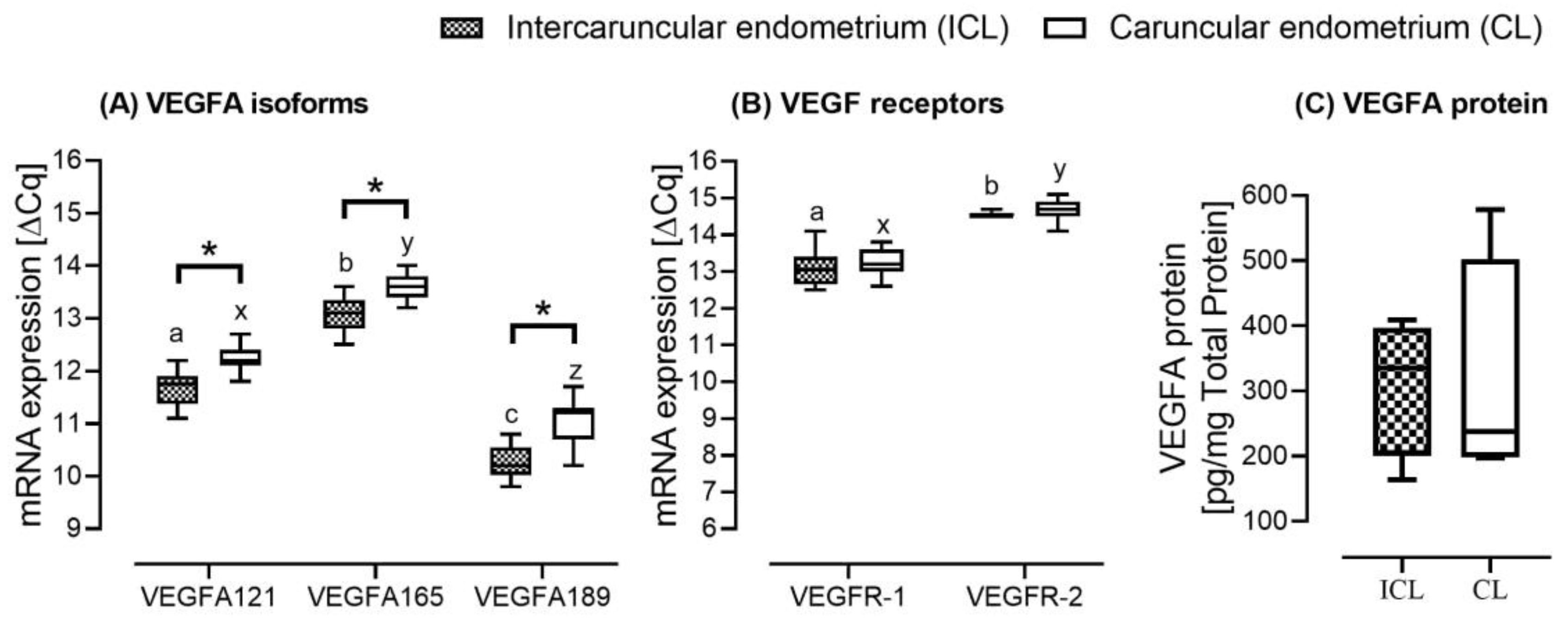
2.2. Study B: Caruncular versus Intercaruncular Endometrium in Pregnant Heifers
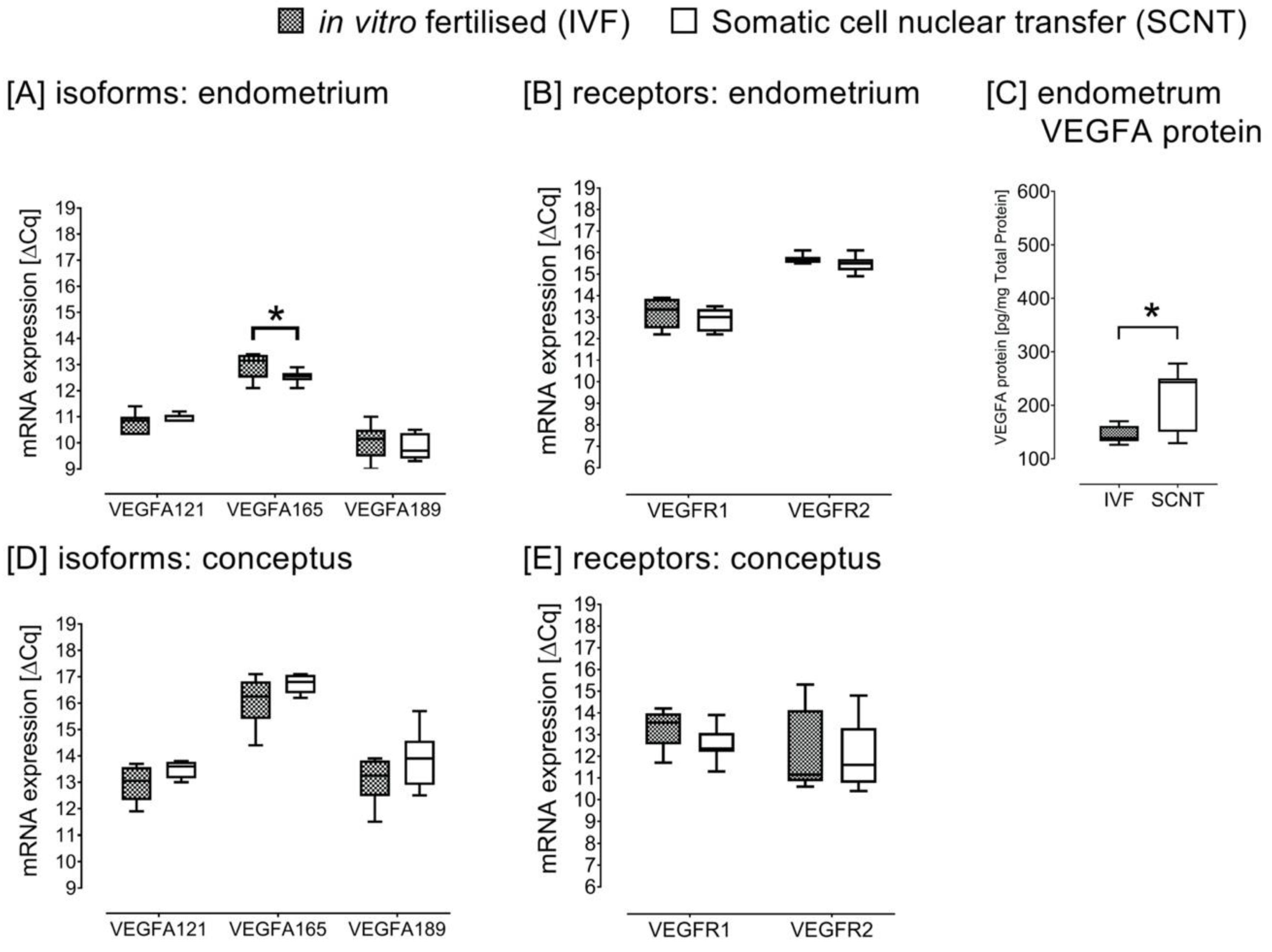
2.3. Study C: SCNT versus IVF Pregnancies
3. Discussion
4. Materials and Methods
4.1. Study A: Cycling versus Pregnant Heifers
4.1.1. Animals and Collection of Samples
4.1.2. Analyses of P4 and E2 Concentration in Serum
4.1.3. Gene Expression Analysis of VEGFA Isoforms and VEGF Receptors in Endometrial and Conceptus Tissues
4.1.4. VEGFA Protein Concentration in Endometrial Homogenates
4.1.5. Immunolocalisation of VEGFA in the Bovine Endometrium
4.2. Study B: Caruncular versus Intercaruncular Endometrium in Pregnant Heifers
4.3. Study C: SCNT versus IVF Pregnancies
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pfarrer, C.D.; Ruziwa, S.D.; Winther, H.; Callesen, H.; Leiser, R.; Schams, D.; Dantzer, V. Localization of vascular endothelial growth factor (VEGF) and its receptors VEGFR-1 and VEGFR-2 in Bovine placentomes from implantation until term. Placenta 2006, 27, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Torry, D.S.; Leavenworth, J.; Chang, M.; Maheshwari, V.; Groesch, K.; Ball, E.R.; Torry, R.J. Angiogenesis in implantation. J. Assist. Reprod. Genet. 2007, 24, 303–315. [Google Scholar] [CrossRef]
- Mackenzie, F.; Ruhrberg, C. Diverse roles for VEGF-A in the nervous system. Development 2012, 139, 1371–1380. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.; Acharya, K.R. Tying the knot: The cystine signature and molecular-recognition processes of the vascular endothelial growth factor family of angiogenic cytokines. FEBS J. 2011, 278, 4304–4322. [Google Scholar] [CrossRef] [PubMed]
- McFee, R.M.; Rozell, T.G.; Cupp, A.S. The balance of proangiogenic and antiangiogenic VEGFA isoforms regulate follicle development. Cell Tissue Res. 2012, 349, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Peach, C.J.; Mignone, V.W.; Arruda, M.A.; Alcobia, D.C.; Hill, S.J.; Kilpatrick, L.E.; Woolard, J. Molecular pharmacology of VEGF-A isoforms: binding and signalling at VEGFR2. Int. J. Mol. Sci. 2018, 19, 1264. [Google Scholar] [CrossRef]
- Bridgett, S.; Dellett, M.; Simpson, D.A. RNA-Sequencing data supports the existence of novel VEGFA splicing events but not of VEGFAxxxb isoforms. Sci. Rep. 2017, 7, 58. [Google Scholar] [CrossRef]
- Sarabipour, S.; Mac-Gabhann, F. VEGF-A121a binding to Neuropilins—A concept revisited. Cell. Adh. Migr. 2018, 12, 204–214. [Google Scholar] [CrossRef]
- Karkkainen, M.J.; Petrova, T.V. Vascular endothelial growth factor receptors in the regulation of angiogenesis and lymphangiogenesis. Oncogene 2000, 19, 5598. [Google Scholar] [CrossRef]
- Mess, A.M.; Carreira, A.C.O.; Marinovic, D.-O.C.; Fratini, P.; Favaron, P.O.; Barreto, R.-D.-S.N.; Pfarrer, C.; Meirelles, F.V. Vascularization and VEGF expression altered in bovine yolk sacs from IVF and NT technologies. Theriogenology 2017, 87, 290–297. [Google Scholar] [CrossRef]
- Luo, H.; Kimura, K.; Aoki, M.; Hirako, M. Vascular endothelial growth factor (VEGF) promotes the early development of bovine embryo in the presence of cumulus cells. J. Vet. Med. Sci. 2002, 64, 967–971. [Google Scholar] [CrossRef]
- Luo, H.; Kimura, K.; Aoki, M.; Hirako, M. Effect of vascular endothelial growth factor on maturation, fertilization and developmental competence of bovine oocytes. J. Vet. Med. Sci. 2002, 64, 803–806. [Google Scholar] [CrossRef][Green Version]
- Anchordoquy, J.M.; Anchordoquy, J.P.; Testa, J.A.; Sirini, M.Á.; Furnus, C.C. Influence of vascular endothelial growth factor and Cysteamine on in vitro bovine oocyte maturation and subsequent embryo development. Cell Biol. Int. 2015, 39, 1090–1098. [Google Scholar] [CrossRef]
- Ortega-Serrano, P.V.; Guzmán, A.; Hernández–Coronado, C.G.; Castillo-Juárez, H.; Rosales-Torres, A.M. Reduction in the mRNA expression of sVEGFR1 and sVEGFR2 is associated with the selection of dominant follicle in cows. Reprod. Domest. Anim. 2016, 51, 985–991. [Google Scholar] [CrossRef]
- Mansouri-Attia, N.; Aubert, J.; Reinaud, P.; Giraud-Delville, C.; Taghouti, G.; Galio, L.; Everts, R.E.; Degrelle, S.; Richard, C.; Hue, I.; et al. Gene expression profiles of bovine caruncular and intercaruncular endometrium at implantation. Physiol. Genom. 2009, 39, 14–27. [Google Scholar] [CrossRef]
- Bauersachs, S.; Ulbrich, S.E.; Zakhartchenko, V.; Minten, M.; Reichenbach, M.; Reichenbach, H.-D.; Blum, H.; Spencer, T.E.; Wolf, E. The endometrium responds differently to cloned versus fertilized embryos. Proc. Natl. Acad. Sci. USA 2009, 106, 5681. [Google Scholar] [CrossRef] [PubMed]
- Panarace, M.; Agüero, J.I.; Garrote, M.; Jauregui, G.; Segovia, A.; Cané, L.; Gutiérrez, J.; Marfil, M.; Rigali, F.; Pugliese, M.; et al. How healthy are clones and their progeny: 5 years of field experience. Theriogenology 2007, 67, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Mansouri-Attia, N.; Sandra, O.; Aubert, J.; Degrelle, S.; Everts, R.E.; Giraud-Delville, C.; Heyman, Y.; Galio, L.; Hue, I.; Yang, X.; et al. Endometrium as an early sensor of in vitro embryo manipulation technologies. Proc. Natl. Acad. Sci. USA 2009, 106, 5687–5692. [Google Scholar] [CrossRef]
- Azimi-Nezhad, M. Vascular endothelial growth factor from embryonic status to cardiovascular pathology. Rep. Biochem. Mol. Biol. 2014, 2, 59–69. [Google Scholar] [PubMed]
- Cheung, C.Y. Vascular endothelial growth factor: Possible role in fetal development and placental function. J. Obstet. Gynaecol. Res. 1997, 4, 169–177. [Google Scholar]
- Kaczmarek, M.M.; Waclawik, A.; Blitek, A.; Kowalczyk, A.E.; Schams, D.; Ziecik, A.J. Expression of the vascular endothelial growth factor-receptor system in the porcine endometrium throughout the estrous cycle and early pregnancy. Mol. Reprod. Dev. 2008, 75, 362–372. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.-P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.-G.; Hosoe, M.; Fujii, S.; Kanahara, H.; Sakumoto, R. Temporal expression and localization of vascular endothelial growth factor family members in the bovine uterus during peri-implantation period. Theriogenology 2019, 133, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Tasaki, Y.; Nishimura, R.; Shibaya, M.; Lee, H.Y.; Acosta, T.J.; Okuda, K. Expression of VEGF and its receptors in the bovine endometrium throughout the estrous cycle: Effects of VEGF on prostaglandin production in endometrial cells. J. Reprod. Dev. 2010, 56, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, B.; Mao, W.; Li, Q.; Fu, C.; Zhang, N.; Zhang, Y.; Gao, L.; Shen, Y.; Cao, J. The effect of prostaglandin E2 receptor (PTGER2) activation on growth factor expression and cell proliferation in bovine endometrial explants. Prostaglandins Leukot. Essent. Fat. Acid. 2017, 122, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Cooke, N.T.F.; Pennington, A.K.; Yang, Q.; Ealy, D.A. Several fibroblast growth factors are expressed during pre-attachment bovine conceptus development and regulate interferon-tau expression from trophectoderm. Reproduction 2009, 37, 259–269. [Google Scholar] [CrossRef]
- Boudjenah, R.; Molina-Gomes, D.; Wainer, R.; de Mazancourt, P.; Selva, J.; Vialard, F. The vascular endothelial growth factor (VEGF) +405 G/C polymorphism and its relationship with recurrent implantation failure in women in an IVF programme with ICSI. J. Assist. Reprod. Genet. 2012, 29, 1415–1420. [Google Scholar] [CrossRef]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef]
- Bloise, E.; Feuer, S.K.; Rinaudo, P.F. Comparative intrauterine development and placental function of ART concepti: Implications for human reproductive medicine and animal breeding. Hum. Reprod. 2014, 20, 822–839. [Google Scholar] [CrossRef]
- Ulbrich, S.E.; Schulke, K.; Groebner, A.E.; Reichenbach, H.D.; Angioni, C.; Geisslinger, G.; Meyer, H.H. Quantitative characterization of prostaglandins in the uterus of early pregnant cattle. Reproduction 2009, 138, 371–382. [Google Scholar] [CrossRef]
- Groebner, A.E.; Schulke, K.; Unterseer, S.; Reichenbach, H.D.; Reichenbach, M.; Büttner, M.; Wolf, E.; Meyer, H.H.D.; Ulbrich, S.E. Enhanced proapoptotic gene expression of XAF1, CASP8 and TNFSF10 in the bovine endometrium during early pregnancy is not correlated with augmented apoptosis. Placenta 2010, 31, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Prakash, B.S.; Meyer, H.H.; Schallenberger, E.; van de Weil, D.F. Development of a sensitive enzymeimmunoassay (EIA) for progesterone determination in unextracted bovine plasma using the second antibody technique. J. Steroid Biochem. 1987, 28, 623–627. [Google Scholar] [CrossRef]
- Giller, K.; Drews, B.; Berard, J.; Kienberger, H.; Schmicke, M.; Frank, J.; Spanier, B.; Daniel, H.; Geisslinger, G.; Ulbrich, S.E. Bovine embryo elongation is altered due to maternal fatty acid supplementation. Biol. Reprod. 2018, 99, 600–610. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day | Status | Animals | Progesterone [ng/mL] | Estradiol-17β [pg/mL] | ||
|---|---|---|---|---|---|---|
| Mean ± SEM | p-Value | Mean ± SEM | p-Value | |||
| Day 12 | nonpregnant | 6 | 8.40 ± 1.02 | 0.7 | 3.51 ± 0.61 | 0.7 |
| pregnant | 5 | 7.83 ± 0.93 | 3.23 ± 0.56 | |||
| Day 15 | nonpregnant | 7 | 6.63 ± 0.86 | 0.04 | 1.92 ± 0.52 | 0.3 |
| pregnant | 6 | 9.37 ± 0.93 | 2.81 ± 0.56 | |||
| Day 18 | nonpregnant | 8 | 6.80 ± 0.80 | 0.003 | 1.20 ± 0.48 | 0.2 |
| pregnant | 5 | 10.91 ± 1.02 | 2.32 ± 0.61 | |||
| Primer | Sequence | Fragment Length [bp] | Accession Number | |
|---|---|---|---|---|
| Polyubiquitin | for | AGATCCAGGATAAGGAAGGCAT | 198 | NM_174133 |
| rev | GCTCCACCTCCAGGGTGAT | |||
| VEGFA-121 | for | CCGTCCCATTGAGACCCTG | 208 | AB455252 |
| rev | CGGCTTGTCACAATTTTTCTTGTC | |||
| VEGFA-165 | for | CCGTCCCATTGAGACCCTG | 278 | NM_174216 |
| rev | GCCCACAGGGATTTTCTTGC | |||
| VEGFA-189 | for | CCGTCCCATTGAGACCCTG | 297 | AB450824 |
| rev | TGCCCTTTGCCCTTTCCTC | |||
| VEGFR-1 | for | CACCAAGAGCGACGTGTG | 351 | X94263 |
| rev | AAGAAGTCCTCGGAGAAGGC | |||
| VEGFR-2 | for | AGACTGGTTCTGGCCCAAC | 379 | X94298 |
| rev | GAAGCCTTTCTGGCTGTC | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiumia, D.; Hankele, A.-K.; Groebner, A.E.; Schulke, K.; Reichenbach, H.-D.; Giller, K.; Zakhartchenko, V.; Bauersachs, S.; Ulbrich, S.E. Vascular Endothelial Growth Factor A and VEGFR-1 Change during Preimplantation in Heifers. Int. J. Mol. Sci. 2020, 21, 544. https://doi.org/10.3390/ijms21020544
Chiumia D, Hankele A-K, Groebner AE, Schulke K, Reichenbach H-D, Giller K, Zakhartchenko V, Bauersachs S, Ulbrich SE. Vascular Endothelial Growth Factor A and VEGFR-1 Change during Preimplantation in Heifers. International Journal of Molecular Sciences. 2020; 21(2):544. https://doi.org/10.3390/ijms21020544
Chicago/Turabian StyleChiumia, Daniel, Anna-Katharina Hankele, Anna E. Groebner, Katy Schulke, Horst-Dieter Reichenbach, Katrin Giller, Valeri Zakhartchenko, Stefan Bauersachs, and Susanne E. Ulbrich. 2020. "Vascular Endothelial Growth Factor A and VEGFR-1 Change during Preimplantation in Heifers" International Journal of Molecular Sciences 21, no. 2: 544. https://doi.org/10.3390/ijms21020544
APA StyleChiumia, D., Hankele, A.-K., Groebner, A. E., Schulke, K., Reichenbach, H.-D., Giller, K., Zakhartchenko, V., Bauersachs, S., & Ulbrich, S. E. (2020). Vascular Endothelial Growth Factor A and VEGFR-1 Change during Preimplantation in Heifers. International Journal of Molecular Sciences, 21(2), 544. https://doi.org/10.3390/ijms21020544