Early-Life Intervention Using Fecal Microbiota Combined with Probiotics Promotes Gut Microbiota Maturation, Regulates Immune System Development, and Alleviates Weaning Stress in Piglets

 ,
,
Abstract
1. Introduction
2. Results
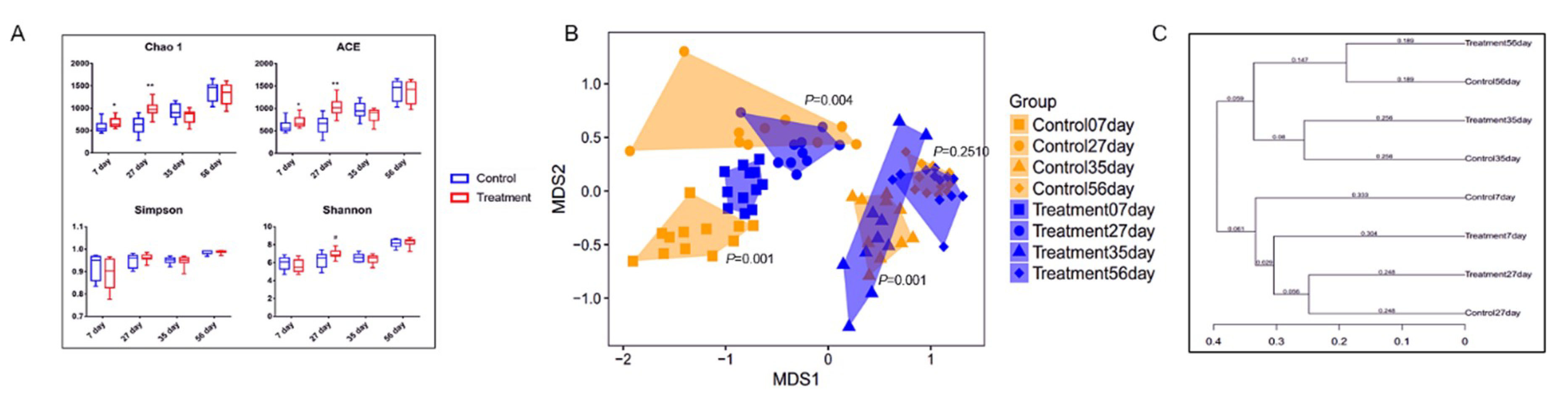
2.1. Bacterial Alpha-Diversity and Beta-Diversity
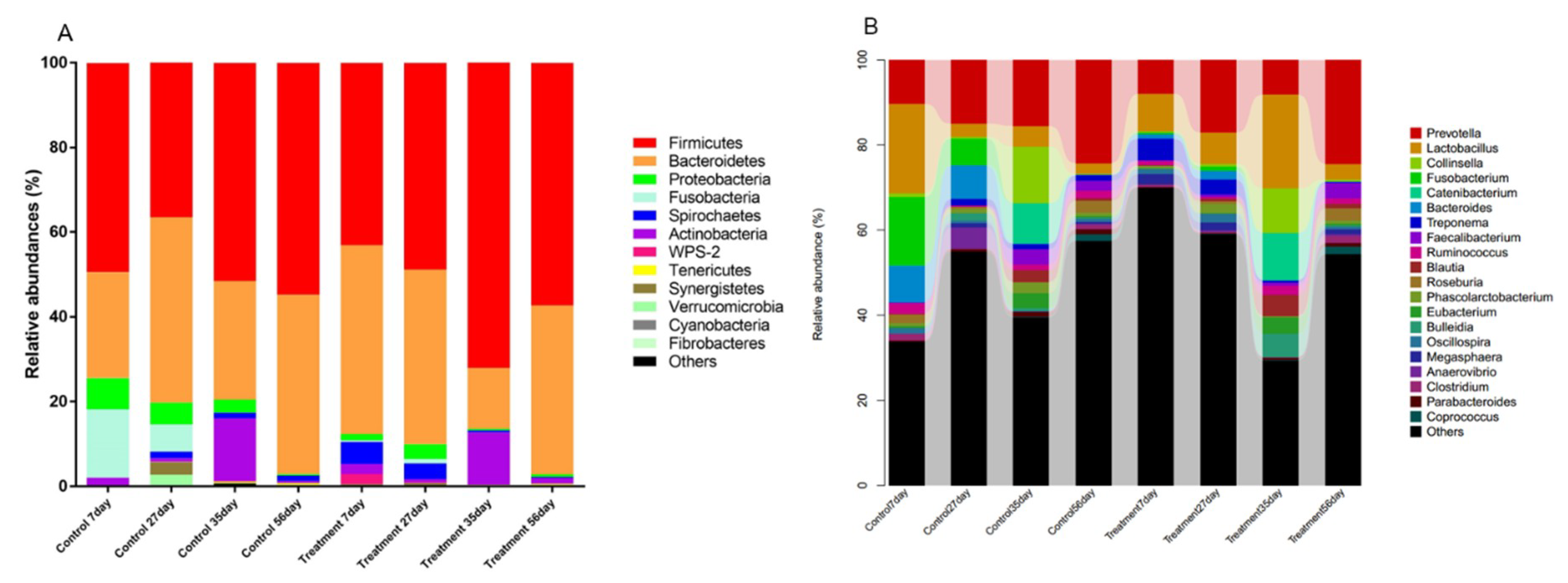
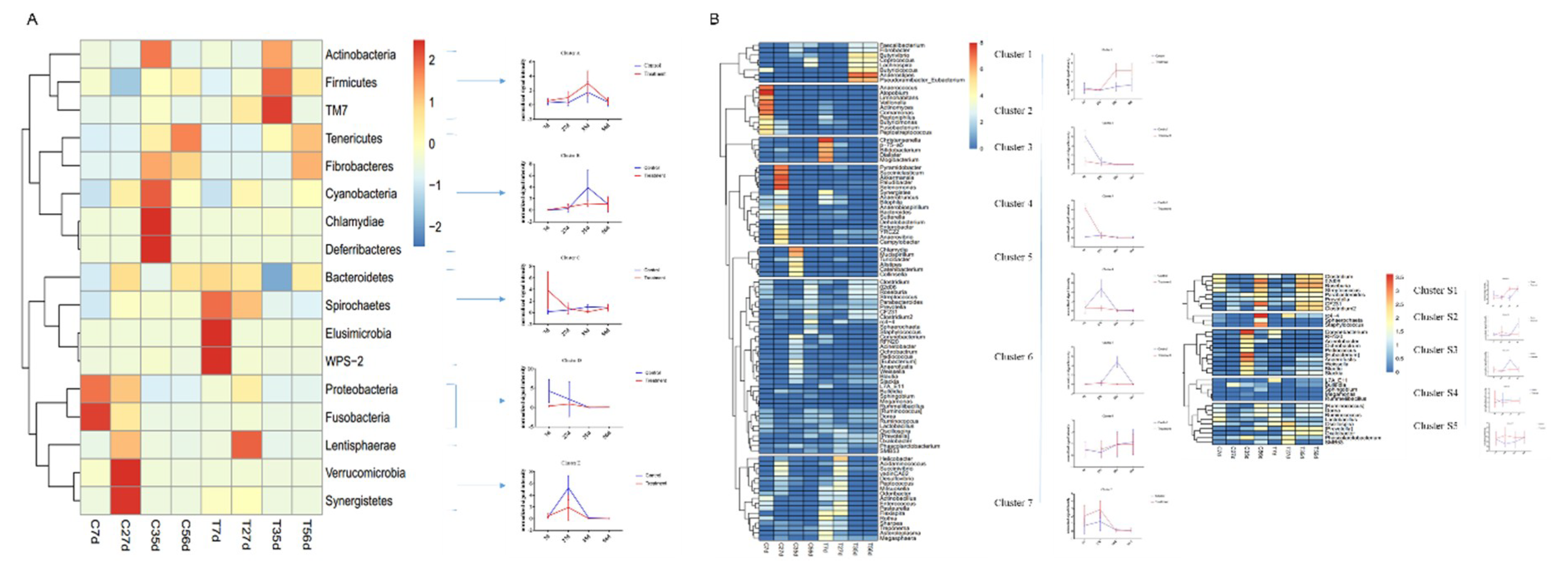
2.2. Early-Life Intervention with FMT-CS Affected the Composition of Fecal Microbiota
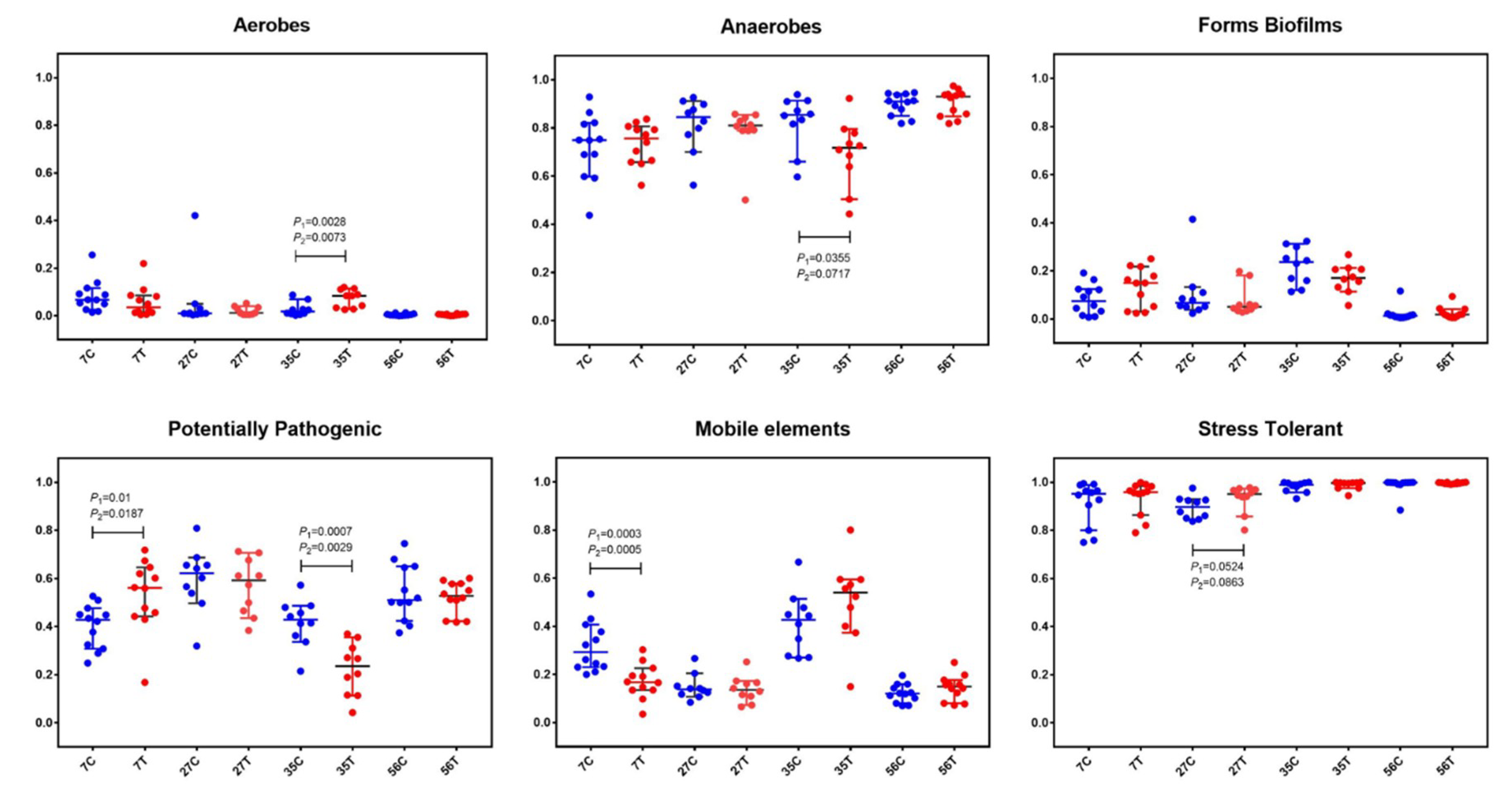
2.3. Early-Life Intervention with FMT-CS Affected the Organism-Level Microbiome Phenotypes
2.4. Early-Life Intervention with FMT-CS Affected the Dominant Bacteria
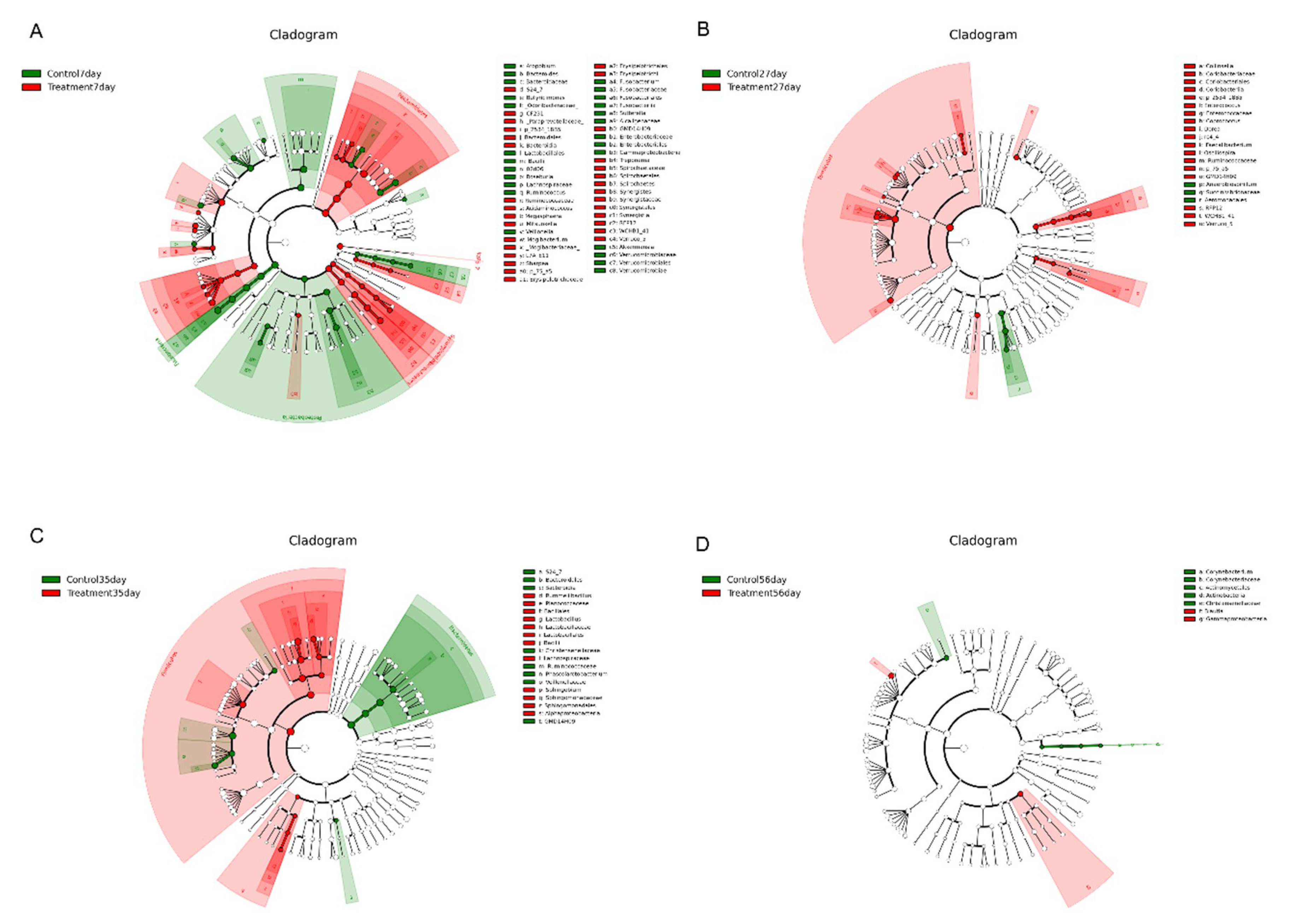
2.5. Early-Life Intervention with FMT-CS Affected the Changing Pattern of Differential Bacteria
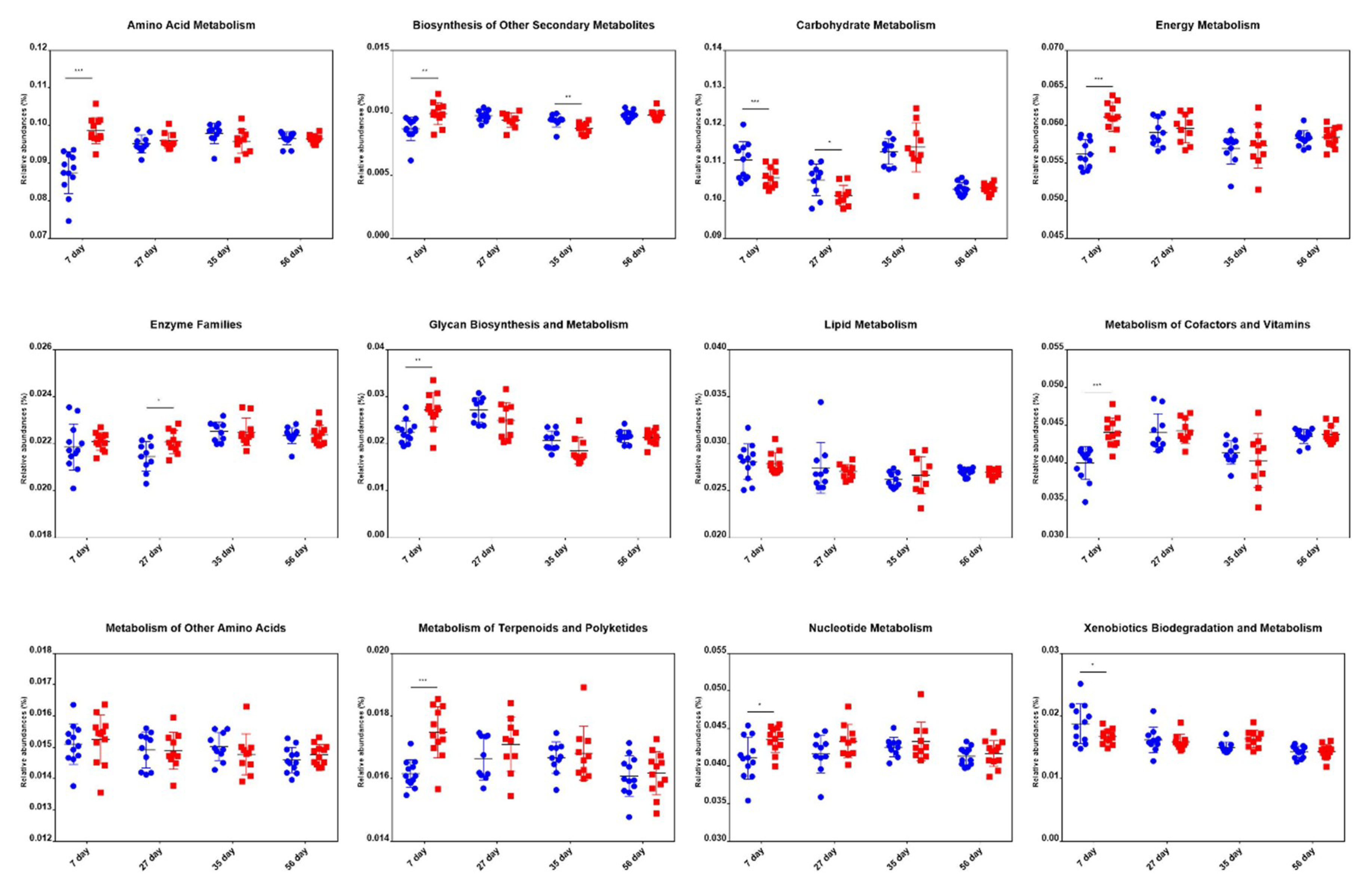
2.6. Comparison of Metabolic Pathway Abundance
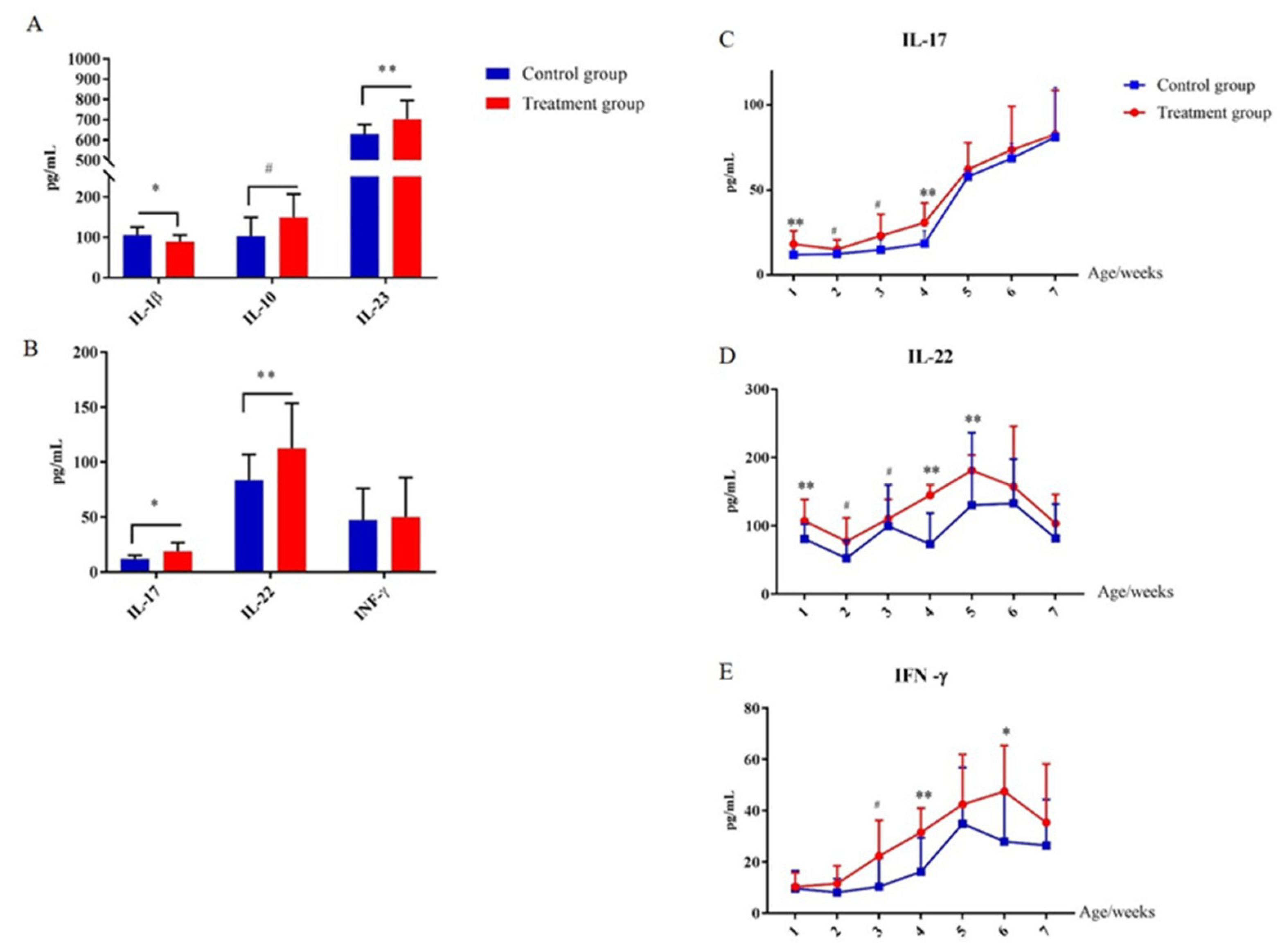
2.7. Early-Life Intervention with FMT-CS Affected Cytokines of the Innate Immune System
2.8. Early-Life Intervention with FMT-CS Affected the Adaptive Immune Homeostasis
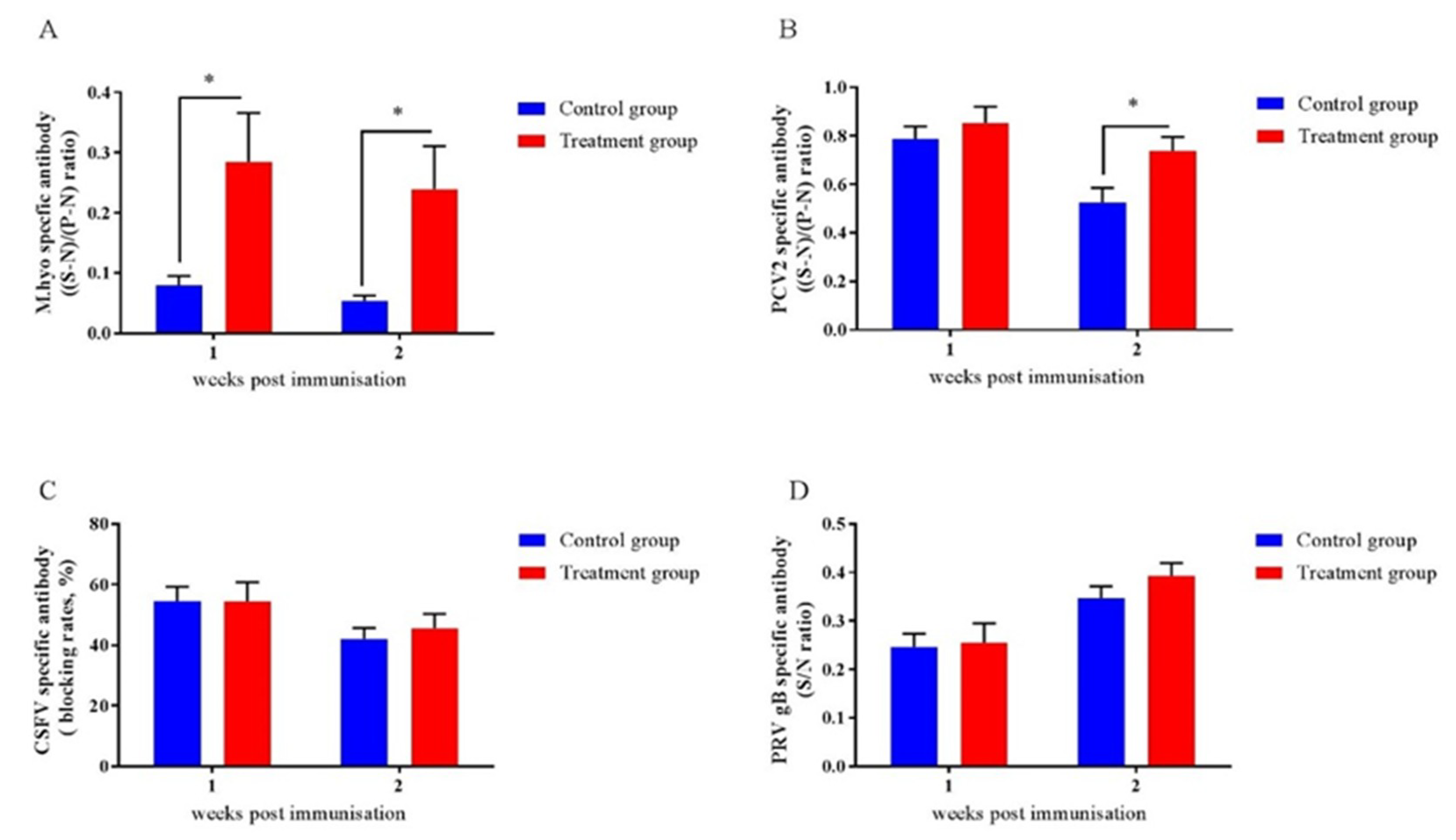
2.9. Early-Life Intervention with FMT-CS Affected the Efficacy of Vaccines
2.10. Early-Life Intervention with FMT-CS Affected Weaning Stress
3. Discussion
3.1. Early-Life Intervention with FMT-CS Promotes the Maturation of Gut Microbiota
3.2. Early-life Intervention with FMT-CS Promotes the Development and Function of Immune System
3.3. Early Intervention with FMT-CS Increases Beneficial Bacteria and Decreases the Opportunistic Pathogens of Piglets
3.4. Early-Life Intervention with FMT-CS Confers Weaning Stress Resistance to Piglets
4. Materials and Methods
4.1. Preparation of Fecal Microbiota Suspension
4.2. Animal Experimental Design
4.3. Immunization
4.4. Sample Collection
4.5. DNA Extraction, 16S rRNA Gene Amplification, and Illumina MiSeq Sequencing
4.6. Analysis of Sequencing Data
4.7. Measurements of Cytokines Related to Innate and Adaptive Immunity Development and Antibody Responses
4.8. Intestinal Barrier Function
4.9. Antioxidant Indices
4.10. Inflammation Levels
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FMT | Fecal microbiota transplantation |
| FMT-CS | FMT combined with C. butyricum and S. boulardii |
| GSH-px | Glutathione peroxidase |
| CAT | Catalase |
| T-AOC | Total antioxidant capacity |
| MDA | Malondialdehyde |
| T-SOD | Total superoxide dismutase |
| D-LA | D-lactate |
| DAO | Diamine oxidase |
| PBSG | Phosphate-buffered saline containing 10% glycerol |
| TBS | Transplanted bacterial suspension |
References
- Brandtzaeg, P. Mucosal immunity: Integration between mother and the breast-fed infant. Vaccine 2003, 21, 3382–3388. [Google Scholar] [CrossRef]
- Georgountzou, A.; Papadopoulos, N.G. Postnatal innate immune development: From birth to adulthood. Front. Immunol. 2017, 8, 957. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Huang, X.; Wang, P.; Yan, Z.; Sun, W.; Zhao, S.; Gun, S. Longitudinal development of the gut microbiota in healthy and diarrheic piglets induced by age-related dietary changes. MicrobiologyOpen 2019. [Google Scholar] [CrossRef] [PubMed]
- Zinkernagel, R.M. Maternal antibodies, childhood infections, and autoimmune diseases. N. Engl. J. Med. 2001, 345, 1331. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, S.; Zheng, J.; Li, W.; Jiang, X.; Zhao, X.; Li, J.; Che, L.; Lin, Y.; Xu, S. Effects of dietary clostridium butyricum supplementation on growth performance, intestinal development, and immune response of weaned piglets challenged with lipopolysaccharide. J. Anim. Sci. Biotechnol. 2018, 9, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Stras, S.F.; Werner, L.; Toothaker, J.M.; Olaloye, O.O.; Oldham, A.L.; McCourt, C.C.; Lee, Y.N.; Rechavi, E.; Shouval, D.S.; Konnikova, L. Maturation of the human intestinal immune system occurs early in fetal development. Dev. Cell 2019, 51, 357–373. [Google Scholar] [CrossRef]
- Renz, H.; Brandtzaeg, P.; Hornef, M. The impact of perinatal immune development on mucosal homeostasis and chronic inflammation. Nat. Rev. Immunol. 2011, 12, 9–23. [Google Scholar] [CrossRef]
- Jain, N.; Walker, W.A. Diet and host-microbial crosstalk in postnatal intestinal immune homeostasis. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 14–25. [Google Scholar] [CrossRef]
- De Gomez, A.M.; Ganal-Vonarburg, S.C.; Fuhrer, T.; Rupp, S.; Uchimura, Y.; Li, H.; Steinert, A.; Heikenwalder, M.; Hapfelmeier, S.; Sauer, U. The maternal microbiota drives early postnatal innate immune development. Science 2016, 351, 1296. [Google Scholar] [CrossRef]
- Hachung, C.; Pamp, S.J.; Hill, J.A.; Surana, N.K.; Edelman, S.M.; Troy, E.B.; Reading, N.C.; Villablanca, E.J.; Sen, W.; Mora, J.R. Gut immune maturation depends on colonization with a host-specific microbiota. Cell 2012, 149, 1578–1593. [Google Scholar]
- Sanos, S.; Bui, V.A.; Oberle, K.; Heners, C.; Johner, C.; Diefenbach, A. Rorgammat and commensal microflora are required for the differentiation of mucosal interleukin 22-producing nkp46+ cells. Nat. Immunol. 2009, 10, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Zhu, A.; Benes, V.; Costea, P.I.; Hercog, R.; Hildebrand, F.; Huerta-Cepas, J.; Nieuwdorp, M.; Salojarvi, J.; Voigt, A.Y.; et al. Durable coexistence of donor and recipient strains after fecal microbiota transplantation. Science 2016, 352, 586–589. [Google Scholar] [CrossRef] [PubMed]
- Litvak, Y.; Baumler, A.J. The founder hypothesis: A basis for microbiota resistance, diversity in taxa carriage, and colonization resistance against pathogens. PLoS Pathog. 2019, 15, e1007563. [Google Scholar] [CrossRef] [PubMed]
- Martinez, I.; Maldonado-Gomez, M.X.; Gomes-Neto, J.C.; Kittana, H.; Ding, H.; Schmaltz, R.; Joglekar, P.; Cardona, R.J.; Marsteller, N.L.; Kembel, S.W.; et al. Experimental evaluation of the importance of colonization history in early-life gut microbiota assembly. Elife 2018, 7, e36521. [Google Scholar] [CrossRef]
- Al Nabhani, Z.; Eberl, G. GAPs in early life facilitate immune tolerance. Sci. Immunol. 2017, 2. [Google Scholar] [CrossRef]
- El, A.S.; Derrien, M.; Merrifield, C.A.; Levenez, F.; Doré, J.; Boekschoten, M.V.; Dekker, J.; Holmes, E.; Zoetendal, E.G.; Van, B.P. Gut bacteria-host metabolic interplay during conventionalisation of the mouse germfree colon. ISME J. 2013, 7, 743–755. [Google Scholar]
- El, A.S.; Hooiveld, G.; Tremaroli, V.; Bäckhed, F.; Kleerebezem, M. The gut microbiota and mucosal homeostasis: Colonized at birth or at adulthood, does it matter? Gut Microbes 2013, 4, 118–124. [Google Scholar]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef]
- Rey-Santano, C.; Mielgo, V.; Valls-i-Soler, A.; Encinas, E.; Lukas, J.C.; Vozmediano, V.; Suarez, E. Evaluation of fentanyl disposition and effects in newborn piglets as an experimental model for human neonates. PLoS ONE 2014, 9, e90728. [Google Scholar] [CrossRef]
- Cheng, C.S.; Wei, H.K.; Wang, P.; Yu, H.C.; Zhang, X.M.; Jiang, S.W.; Peng, J. Early intervention with faecal microbiota transplantation: An effective means to improve growth performance and the intestinal development of suckling piglets. Anim. Int. J. Anim. Biosci. 2019, 13, 533–541. [Google Scholar] [CrossRef]
- Espey, M.G. Role of oxygen gradients in shaping redox relationships between the human intestine and its microbiota. Free Radic. Biol. Med. 2013, 55, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.G.; Sakamoto, K.; Seo, S.U.; Pickard, J.M.; Gillilland, M.G., 3rd; Pudlo, N.A.; Hoostal, M.; Li, X.; Wang, T.D.; Feehley, T.; et al. Neonatal acquisition of clostridia species protects against colonization by bacterial pathogens. Science 2017, 356, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Hudson, L.E.; McDermott, C.D.; Stewart, T.P.; Hudson, W.H.; Rios, D.; Fasken, M.B.; Corbett, A.H.; Lamb, T.J. Characterization of the probiotic yeast saccharomyces boulardii in the healthy mucosal immune system. PLoS ONE 2016, 11, e0153351. [Google Scholar] [CrossRef]
- Hancox, L.R.; Le Bon, M.; Richards, P.J.; Guillou, D.; Dodd, C.E.; Mellits, K.H. Effect of a single dose of Saccharomyces cerevisiae var. boulardii on the occurrence of porcine neonatal diarrhoea. Anim. Int. J. Anim. Biosci. 2015, 9, 1756–1759. [Google Scholar] [CrossRef] [PubMed]
- Stier, H.; Bischoff, S.C. Saccharomyces boulardii cncm i-745 influences the gut-associated immune system. MMW Fortschr. Med. 2017, 159, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.F.; Tang, Z.S.; Ling, T.; Tao, X.X.; Suo, Q.F.; Xu, X.M. Effects of clostridium butyricum and bifidobacterium on btla expression on cd4 + t cells and lymphocyte differentiation in late preterm infants. Microb. Pathog. 2016, 100, 112–118. [Google Scholar] [CrossRef]
- Ling, Z.; Liu, X.; Cheng, Y.; Luo, Y.; Yuan, L.; Li, L.; Xiang, C. Clostridium butyricum combined with bifidobacterium infantis probiotic mixture restores fecal microbiota and attenuates systemic inflammation in mice with antibiotic-associated diarrhea. Biomed. Res. Int. 2015, 2015, 582048. [Google Scholar] [CrossRef]
- Wang, K.; Cao, G.; Zhang, H.; Li, Q.; Yang, C. Effects of Clostridium butyricum and Enterococcus faecalis on growth performance, immune function, intestinal morphology, volatile fatty acids, and intestinal flora in a piglet model. Food Funct 2019, 10, 7844–7854. [Google Scholar] [CrossRef]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16s rrna marker gene sequences. Nat. Biotechnol. 2013, 31, 814. [Google Scholar] [CrossRef]
- Hu, J.; Nie, Y.; Chen, J.; Zhang, Y.; Wang, Z.; Fan, Q.; Yan, X. Gradual changes of gut microbiota in weaned miniature piglets. Front. Microbiol. 2016, 7, 1727. [Google Scholar] [CrossRef]
- Wang, T.; Teng, K.; Liu, Y.; Shi, W.; Zhang, J.; Dong, E.; Zhang, X.; Tao, Y.; Zhong, J. Lactobacillus plantarum pfm 105 promotes intestinal development through modulation of gut microbiota in weaning piglets. Front. Microbiol. 2019, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Geng, S.; Cheng, S.; Li, Y.; Wen, Z.; Ma, X.; Jiang, X.; Wang, Y.; Han, X. Faecal microbiota transplantation reduces susceptibility to epithelial injury and modulates tryptophan metabolism of the microbial community in a piglet model. J. Crohns Colitis 2018, 12, 1359–1374. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef]
- Hu, L.; Geng, S.; Li, Y.; Cheng, S.; Fu, X.; Yue, X.; Han, X. Exogenous fecal microbiota transplantation from local adult pigs to crossbred newborn piglets. Front. Microbiol. 2017, 8, 2663. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Bergsma, N.; Parhiala, R.; Lahtinen, S.; Gueimonde, M.; Finne-Soveri, H.; Strandberg, T.; Pitkala, K.; Salminen, S. Bifidobacterium microbiota and parameters of immune function in elderly subjects. FEMS Immunol. Med. Microbiol. 2008, 53, 18–25. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef]
- Nguyen, Q.N.; Himes, J.E.; Martinez, D.R.; Permar, S.R. The impact of the gut microbiota on humoral immunity to pathogens and vaccination in early infancy. PLoS Pathog. 2016, 12, e1005997. [Google Scholar] [CrossRef]
- Slifierz, M.J.; Friendship, R.M.; Weese, J.S. Longitudinal study of the early-life fecal and nasal microbiotas of the domestic pig. BMC Microbiol. 2015, 15, 184. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, X.; Liu, P.; Zhao, J.; Sun, J.; Guan, W.; Johnston, L.J.; Levesque, C.L.; Fan, P.; He, T.; et al. Dietary clostridium butyricum induces a phased shift in fecal microbiota structure and increases the acetic acid-producing bacteria in a weaned piglet model. J. Agric. Food Chem. 2018, 66, 5157–5166. [Google Scholar] [CrossRef]
- McDonald, E. Vitamins and cofactors: Highlights of esboc 2009. Nat. Chem. Biol. 2009, 5, 530–533. [Google Scholar] [CrossRef] [PubMed]
- Sawa, S.; Cherrier, M.; Lochner, M.; Satoh-Takayama, N.; Fehling, H.J.; Langa, F.; Di Santo, J.P.; Eberl, G. Lineage relationship analysis of rorgammat+ innate lymphoid cells. Science 2010, 330, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.; Horn, K.; Rudloff, I.; Goren, I.; Holdener, M.; Christen, U.; Darsow, N.; Hunfeld, K.P.; Koehl, U.; Kind, P.; et al. Early production of il-22 but not il-17 by peripheral blood mononuclear cells exposed to live borrelia burgdorferi: The role of monocytes and interleukin-1. PLoS Pathog. 2010, 6, e1001144. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate lymphoid cells: 10 years on. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Ando, M.; Kamada, N.; Nagano, Y.; Narushima, S.; Suda, W.; Imaoka, A.; Setoyama, H.; Nagamori, T.; et al. Th17 cell induction by adhesion of microbes to intestinal epithelial cells. Cell 2015, 163, 367–380. [Google Scholar] [CrossRef]
- Sano, T.; Huang, W.; Hall, J.A.; Yang, Y.; Chen, A.; Gavzy, S.J.; Lee, J.Y.; Ziel, J.W.; Miraldi, E.R.; Domingos, A.I.; et al. An il-23r/il-22 circuit regulates epithelial serum amyloid a to promote local effector th17 responses. Cell 2015, 163, 381–393. [Google Scholar] [CrossRef]
- Mortha, A.; Chudnovskiy, A.; Hashimoto, D.; Bogunovic, M.; Spencer, S.P.; Belkaid, Y.; Merad, M. Microbiota-dependent crosstalk between macrophages and ilc3 promotes intestinal homeostasis. Science 2014, 343, 1249288. [Google Scholar] [CrossRef]
- Celi, P.; Verlhac, V.; Pérez Calvo, E.; Schmeisser, J.; Kluenter, A.-M. Biomarkers of gastrointestinal functionality in animal nutrition and health. Anim. Feed Sci. Technol. 2019, 250, 9–31. [Google Scholar] [CrossRef]
- Harris, V.C.; Haak, B.W.; Handley, S.A.; Jiang, B.; Velasquez, D.E.; Hykes, B.L., Jr.; Droit, L.; Berbers, G.A.M.; Kemper, E.M.; van Leeuwen, E.M.M.; et al. Effect of antibiotic-mediated microbiome modulation on rotavirus vaccine immunogenicity: A human, randomized-control proof-of-concept trial. Cell Host Microbe 2018, 24, 197–207. [Google Scholar] [CrossRef]
- Lynn, M.A.; Tumes, D.J.; Choo, J.M.; Sribnaia, A.; Blake, S.J.; Leong, L.E.X.; Young, G.P.; Marshall, H.S.; Wesselingh, S.L.; Rogers, G.B.; et al. Early-life antibiotic-driven dysbiosis leads to dysregulated vaccine immune responses in mice. Cell Host Microbe 2018, 23, 653–660. [Google Scholar] [CrossRef]
- Luo, Q.; Cui, H.; Peng, X.; Fang, J.; Zuo, Z.; Deng, J.; Liu, J.; Deng, Y. Intestinal iga(+) cell numbers as well as iga, igg, and igm contents correlate with mucosal humoral immunity of broilers during supplementation with high fluorine in the diets. Biol. Trace Elem. Res. 2013, 154, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, R.; Kim, H.B. The intestinal microbiome of the pig. Anim. Health Res. Rev. 2012, 13, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Brunse, A.; Martin, L.; Rasmussen, T.S.; Christensen, L.; Skovsted Cilieborg, M.; Wiese, M.; Khakimov, B.; Pieper, R.; Nielsen, D.S.; Sangild, P.T. Effect of fecal microbiota transplantation route of administration on gut colonization and host response in preterm pigs. ISME J. 2019, 13, 720. [Google Scholar] [CrossRef]
- Reetta, S.; Susana, F.; Eero, M.; Jonna, J.; De Vos, W.M.; Perttu, A. Fecal transplantation treatment of antibiotic-induced, noninfectious colitis and long-term microbiota follow-up. Case Rep. Med. 2014, 2014, 913867. [Google Scholar]
- O’Toole, P.W.; Cooney, J.C. Probiotic bacteria influence the composition and function of the intestinal microbiota. Interdiscip. Perspect. Infect. Dis. 2008, 2008, 175285. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, C.; Lv, W.; Chao, L.; Li, Z.; Shi, D.; Guo, S. Structural modulation of gut microbiota during alleviation of suckling piglets diarrhoea with herbal formula. Evid. Based Complement. Altern. Med. 2017, 2017, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Correa, R.O.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar]
- Konstantinov, S.R.; Awati, A.A.; Williams, B.A.; Miller, B.G.; Jones, P.; Stokes, C.R.; Akkermans, A.D.L.; Smidt, H.; De Vos, W.M. Post-natal development of the porcine microbiota composition and activities. Environ. Microbiol. 2006, 8, 1191–1199. [Google Scholar] [CrossRef]
- Daly, K.; Darby, A.C.; Hall, N.; Nau, A.; Bravo, D.; Shirazi-Beechey, S.P. Dietary supplementation with lactose or artificial sweetener enhances swine gut lactobacillus population abundance. Br. J. Nutr. 2014, 111 (Suppl. 1), S30. [Google Scholar] [CrossRef]
- Jiao, J.; Wu, J.; Zhou, C.; Tang, S.; Wang, M.; Tan, Z. Composition of ileal bacterial community in grazing goats varies across non-rumination, transition and rumination stages of life. Front. Microbiol. 2016, 7, 1364. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Shan, X.; Cai, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan modulates the gut microbiota in mice by increasing the abundance of lactobacillus and ruminococcaceae. Food Funct. 2016, 7, 3224. [Google Scholar] [CrossRef] [PubMed]
- Vujkovic-Cvijin, I.; Swainson, L.; Chu, S.; Ortiz, A.; Santee, C.; Petriello, A.; Dunham, R.; Fadrosh, D.; Lin, D.; Faruqi, A. Gut-resident lactobacillus abundance associates with ido1 inhibition and th17 dynamics in siv-infected macaques. Cell Rep. 2015, 13, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Ma, L.; Nie, Y.; Chen, J.; Zheng, W.; Wang, X.; Xie, C.; Zheng, Z.; Wang, Z.; Yang, T.; et al. A microbiota-derived bacteriocin targets the host to confer diarrhea resistance in early-weaned piglets. Cell Host Microbe 2018, 24, 817–832. [Google Scholar] [CrossRef] [PubMed]
- Fukudome, I.; Kobayashi, M.; Dabanaka, K.; Maeda, H.; Okamoto, K.; Okabayashi, T.; Baba, R.; Kumagai, N.; Oba, K.; Fujita, M. Diamine oxidase as a marker of intestinal mucosal injury and the effect of soluble dietary fiber on gastrointestinal tract toxicity after intravenous 5-fluorouracil treatment in rats. Med. Mol. Morphol. 2014, 47, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Rokkas, T.; Vaja, S.; Murphy, G.M.; Dowling, R.H. Postheparin plasma diamine oxidase in health and intestinal disease. Gastroenterology 1990, 98, 1493–1501. [Google Scholar] [CrossRef]
- Xun, W.; Shi, L.; Zhou, H.; Hou, G.; Cao, T.; Zhao, C. Effects of curcumin on growth performance, jejunal mucosal membrane integrity, morphology and immune status in weaned piglets challenged with enterotoxigenic escherichia coli. Int. Immunopharmacol. 2015, 27, 46–52. [Google Scholar] [CrossRef]
- Odamaki, T.; Kato, K.; Sugahara, H. Effect of probiotic yoghurt on animal-based diet-induced change in gut microbiota an open, randomised, parallel-group study. Benef. Microbes 2016, 7, 1–12. [Google Scholar] [CrossRef]
- Anderson, J.L.; Edney, R.J.; Whelan, K. Systematic review: Faecal microbiota transplantation in the management of inflammatory bowel disease. Aliment. Pharmacol. Ther. 2012, 36, 503–516. [Google Scholar] [CrossRef]
- Cammarota, G.; Ianiro, G.; Gasbarrini, A. Fecal microbiota transplantation for the treatment of clostridium difficile infection: A systematic review. J. Clin. Gastroenterol. 2014, 48, 693. [Google Scholar] [CrossRef]
- Quraishi, M.N.; Widlak, M.; Bhala, N.; Moore, D.; Price, M.; Sharma, N.; Iqbal, T.H. Systematic review with meta-analysis: The efficacy of faecal microbiota transplantation for the treatment of recurrent and refractory clostridium difficile infection. Aliment. Pharmacol. Ther. 2017, 46, 479. [Google Scholar] [CrossRef]
- Ping, Z.; Bing, Y.; Jun, H.; Gang, T.; Yuheng, L.; Xiangbing, M.; Keying, Z.; Lianqiang, C.; Daiwen, C. Protective effects of dietary arginine supplementation against oxidative stress in weaned piglets. Br. J. Nutr. 2013, 109, 2253–2260. [Google Scholar]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut microbiota dysbiosis in postweaning piglets: Understanding the keys to health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, X.; Shang, Q.; Gao, Z.; Hao, F.; Guo, H.; Guo, C. Fecal microbiota transplantation (fmt) could reverse the severity of experimental necrotizing enterocolitis (nec) via oxidative stress modulation. Free Radic. Biol. Med. 2017, 108, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.R.; Borre, Y.; Brien, C.O.; Patterson, E.; Aidy, S.E.; Deane, J.; Kennedy, P.J.; Beers, S.; Scott, K.; Moloney, G. Transferring the blues: Depression-associated gut microbiota induces neurobehavioural changes in the rat. J. Psychiatr. Res. 2016, 82, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Agata, K.; Lidia, C.; Jerzy, S.; Aleksandra, S.; Katarzyna, K.; Irena, F. Exogenous il-2 controls the balance in th1, th17, and treg cell distribution in patients with progressive rheumatoid arthritis treated with tnf-alpha inhibitors. Inflammation 2015, 38, 765–774. [Google Scholar]
- Shameli, A.; Yamanouchi, J.; Tsai, S.; Yang, Y.; Clemente-Casares, X.; Moore, A.; Serra, P.; Santamaria, P. Il-2 promotes the function of memory-like autoregulatory cd8+ t cells but suppresses their development via foxp3+ treg cells. Eur. J. Immunol. 2013, 43, 394–403. [Google Scholar] [CrossRef]
- Pang, X.; Hua, X.; Yang, Q.; Ding, D.; Che, C.; Cui, L.; Jia, W.; Bucheli, P.; Zhao, L. Inter-species transplantation of gut microbiota from human to pigs. ISME J. 2007, 1, 156–162. [Google Scholar] [CrossRef]
- Ward, T.; Larson, J.; Meulemans, J.; Hillmann, B.; Lynch, J.; Sidiropoulos, D.; Spear, J.; Caporaso, G.; Ran, B.; Knight, R. Bugbase predicts organism level microbiome phenotypes. BioRxiv 2017, 133462. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Control | Treatment | SEM | p-Value |
|---|---|---|---|---|
| Day 14 | ||||
| Fecal sIgA, μg/g | 128.24 b | 168.53 a | 1.35 | < 0.0001 |
| Plasma IgG, mg/mL | 19.8 b | 26.94 a | 1.47 | 0.0135 |
| Plasma IgM, mg/mL | 15.48 b | 22.46 a | 1.1 | < 0.0001 |
| Day 21 | ||||
| Fecal sIgA, μg/g | 114.19 b | 145.92 a | 1.8 | 0.0242 |
| Plasma IgG, mg/mL | 20.56 b | 26.28 a | 1.08 | 0.0052 |
| Plasma IgM, mg/mL | 11.1 b | 17.86 a | 1.12 | 0.0004 |
| Day 35 | ||||
| Fecal sIgA, μg/g | 132.09 b | 148.78 a | 3.25 | 0.0071 |
| Plasma IgG, mg/mL | 13.74 b | 24.08 a | 1.22 | < 0.0001 |
| Plasma IgM, mg/mL | 10.95 b | 18.03 a | 0.95 | < 0.0001 |
| Day 56 | ||||
| Fecal sIgA, μg/g | 102.24 b | 128.12 a | 3.85 | 0.0001 |
| Plasma IgG, mg/mL | 16.92 b | 19.05 a | 0.41 | 0.0058 |
| Plasma IgM, mg/mL | 9.91 b | 13.69 a | 0.58 | < 0.0001 |
| Items 1 | Biomarkers | Control | Treatment | SEM | p-Value |
|---|---|---|---|---|---|
| Antioxidant Indices | CAT, U/mL | 25.14 b | 33.22 a | 1.81 | 0.0218 |
| GSH-px, U/mL | 368.38 | 469.15 | 26.12 | 0.0514 | |
| MDA, nmol/mL | 7.81 a | 5.30 b | 0.53 | 0.0147 | |
| T-AOC, U/mL | 36.22 b | 48.70 a | 2.94 | 0.0302 | |
| T-SOD, U/mL | 14.58 b | 35.59 a | 1.44 | 0.0271 | |
| Intestinal Barrier Biomarkers | DAO, ng/mL | 374.18 a | 296.8 b | 17.513 | 0.0115 |
| D-LA, umol/mL | 880.16 a | 631.27 b | 19.84 | 0.0004 | |
| Inflammation Levels | IFN-γ, pg/mL | 70.99 | 59.43 | 2.72 | 0.0530 |
| TNF-α, pg/mL | 443.13 a | 245.89 b | 9.42 | < 0.0001 | |
| IL-6, pg/mL | 1152.04 a | 900.18 b | 34.09 | 0.0044 | |
| IL-2, pg/mL | 610.79 | 729.64 | 36.86 | 0.1640 | |
| IL-10, pg/mL | 374.69 | 493.06 | 27.96 | 0.0713 | |
| Cortisol, ng/mL | 272.73 a | 210.05 b | 8.59 | 0.0022 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, Q.; Wu, X.; Pan, Y.; Wang, L.; Cui, C.; Guo, Y.; Zhu, L.; Peng, J.; Wei, H. Early-Life Intervention Using Fecal Microbiota Combined with Probiotics Promotes Gut Microbiota Maturation, Regulates Immune System Development, and Alleviates Weaning Stress in Piglets. Int. J. Mol. Sci. 2020, 21, 503. https://doi.org/10.3390/ijms21020503
Xiang Q, Wu X, Pan Y, Wang L, Cui C, Guo Y, Zhu L, Peng J, Wei H. Early-Life Intervention Using Fecal Microbiota Combined with Probiotics Promotes Gut Microbiota Maturation, Regulates Immune System Development, and Alleviates Weaning Stress in Piglets. International Journal of Molecular Sciences. 2020; 21(2):503. https://doi.org/10.3390/ijms21020503
Chicago/Turabian StyleXiang, Quanhang, Xiaoyu Wu, Ye Pan, Liu Wang, Chenbin Cui, Yuwei Guo, Lingling Zhu, Jian Peng, and Hongkui Wei. 2020. "Early-Life Intervention Using Fecal Microbiota Combined with Probiotics Promotes Gut Microbiota Maturation, Regulates Immune System Development, and Alleviates Weaning Stress in Piglets" International Journal of Molecular Sciences 21, no. 2: 503. https://doi.org/10.3390/ijms21020503
APA StyleXiang, Q., Wu, X., Pan, Y., Wang, L., Cui, C., Guo, Y., Zhu, L., Peng, J., & Wei, H. (2020). Early-Life Intervention Using Fecal Microbiota Combined with Probiotics Promotes Gut Microbiota Maturation, Regulates Immune System Development, and Alleviates Weaning Stress in Piglets. International Journal of Molecular Sciences, 21(2), 503. https://doi.org/10.3390/ijms21020503