Functioning of the Photosynthetic Apparatus in Response to Drought Stress in Oat × Maize Addition Lines

 , , ,
, , ,  , , ,
, , , 
Abstract
1. Introduction
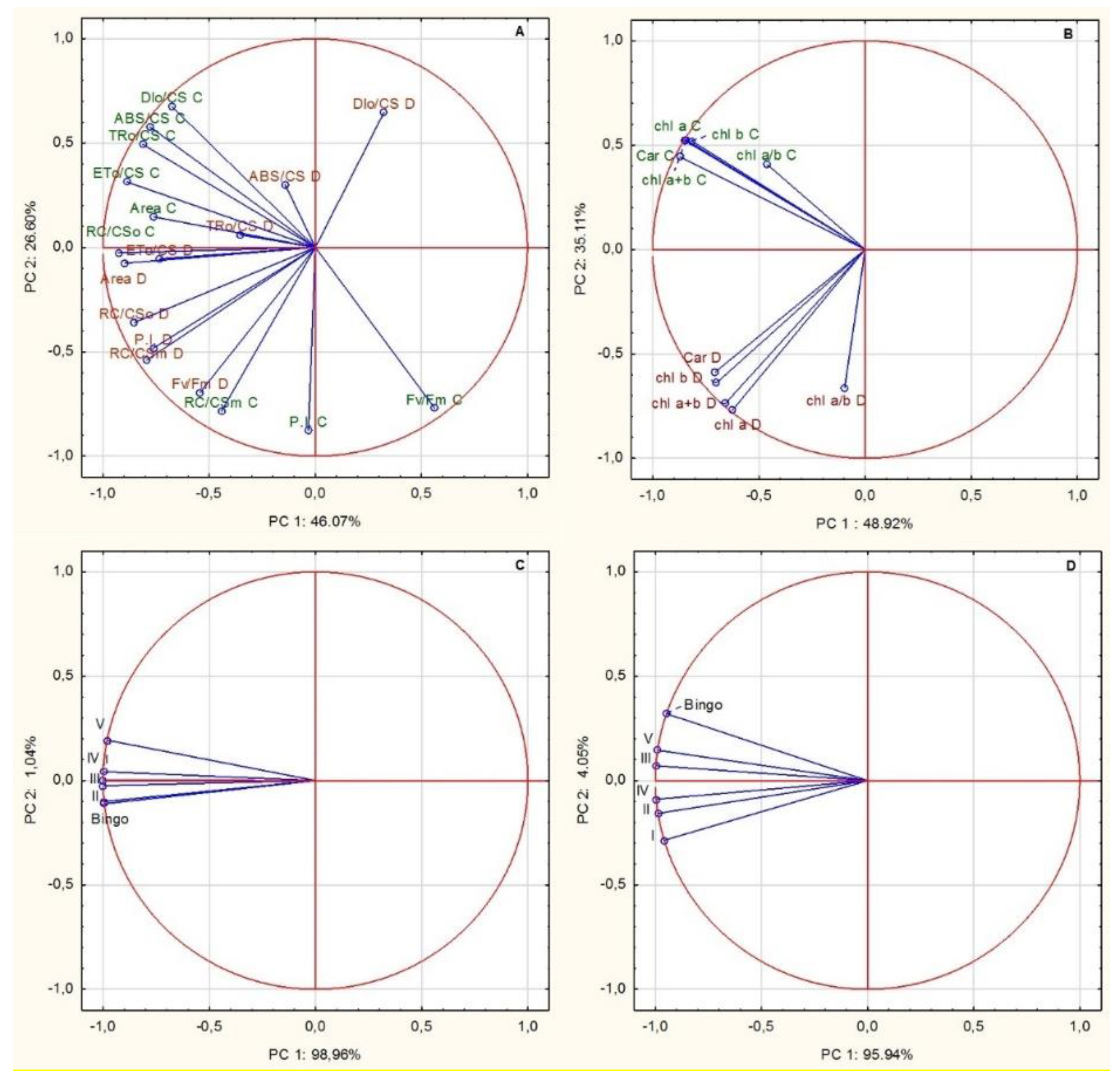
2. Results
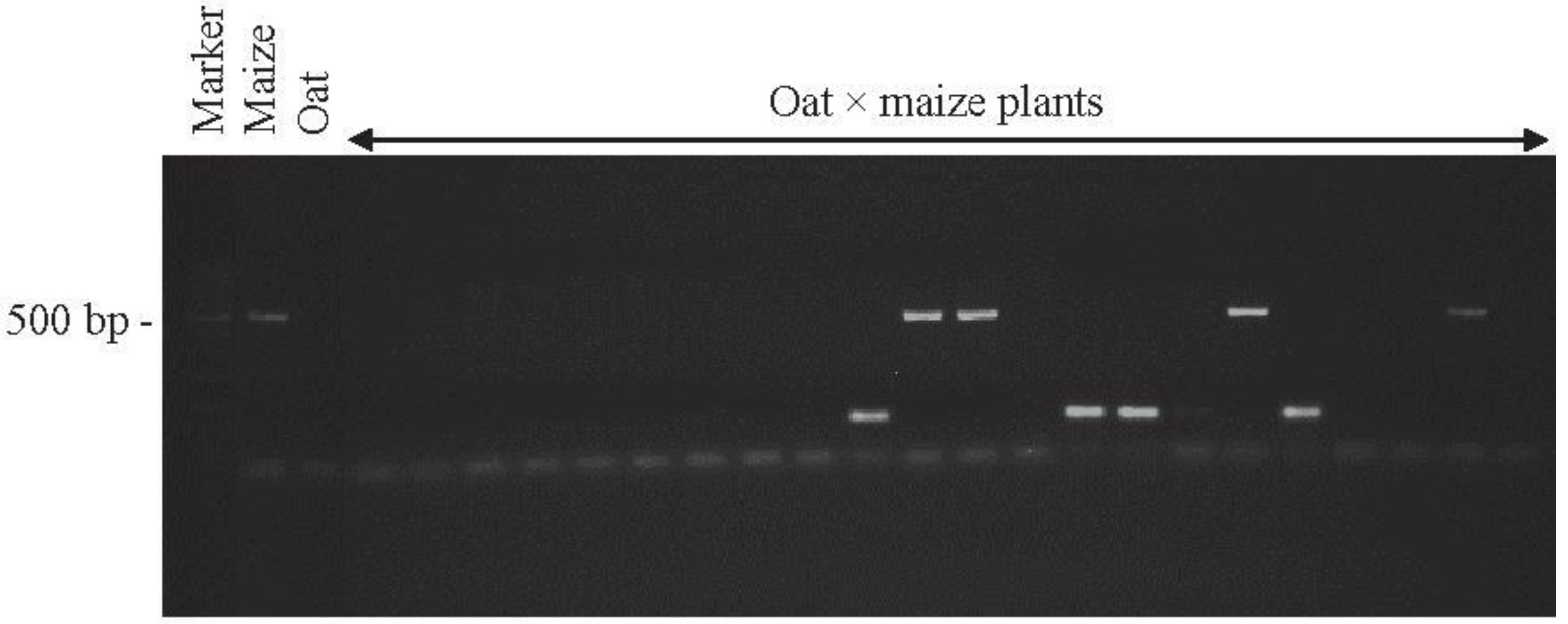
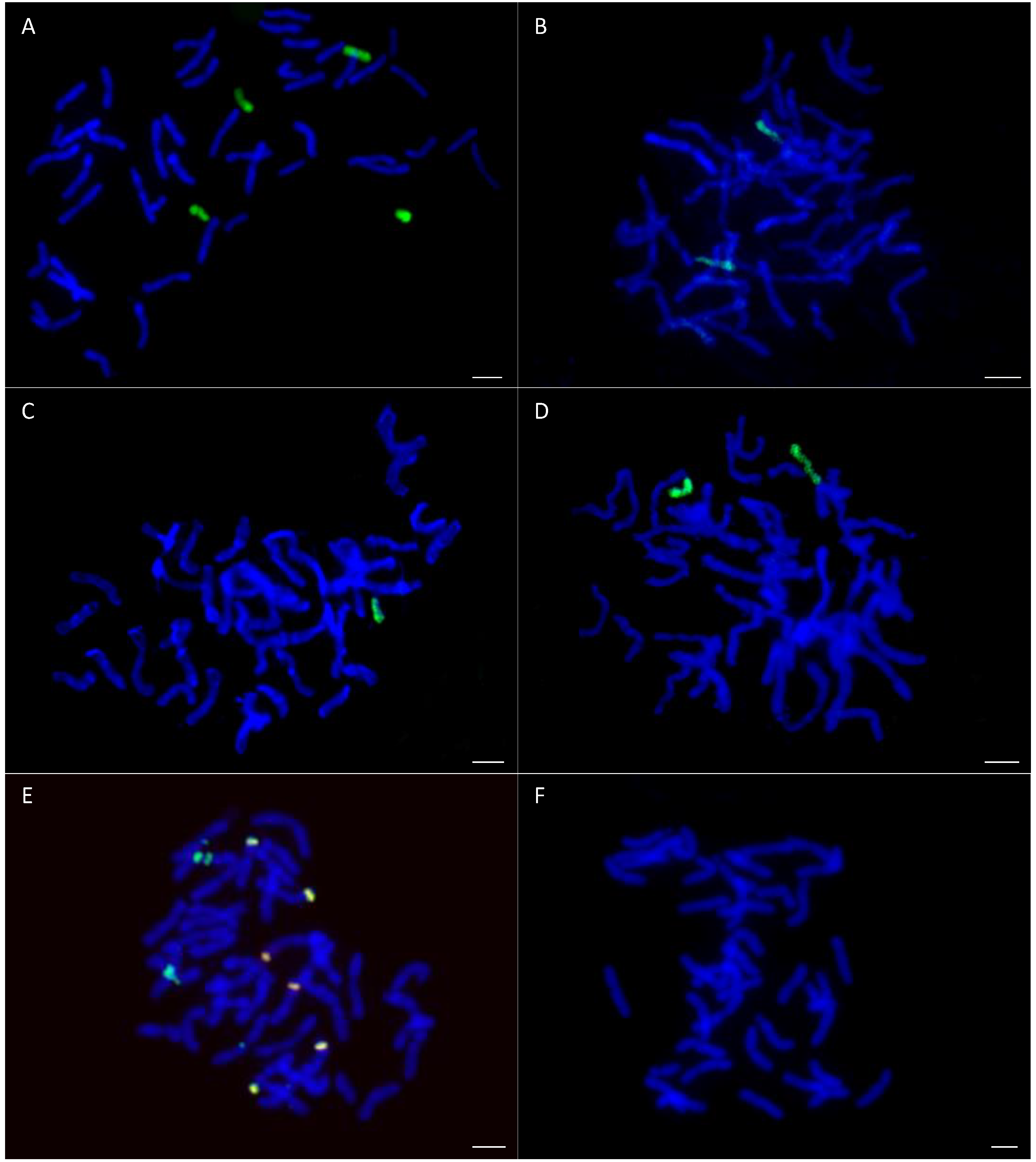
2.1. Detection of OMA Lines
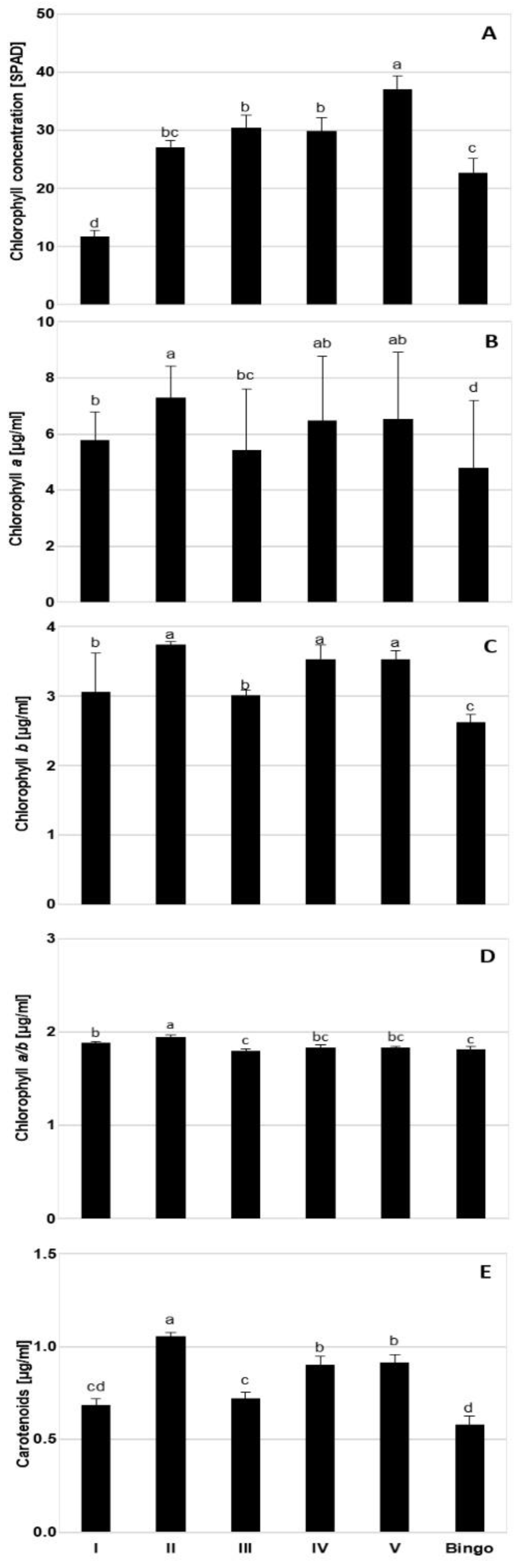
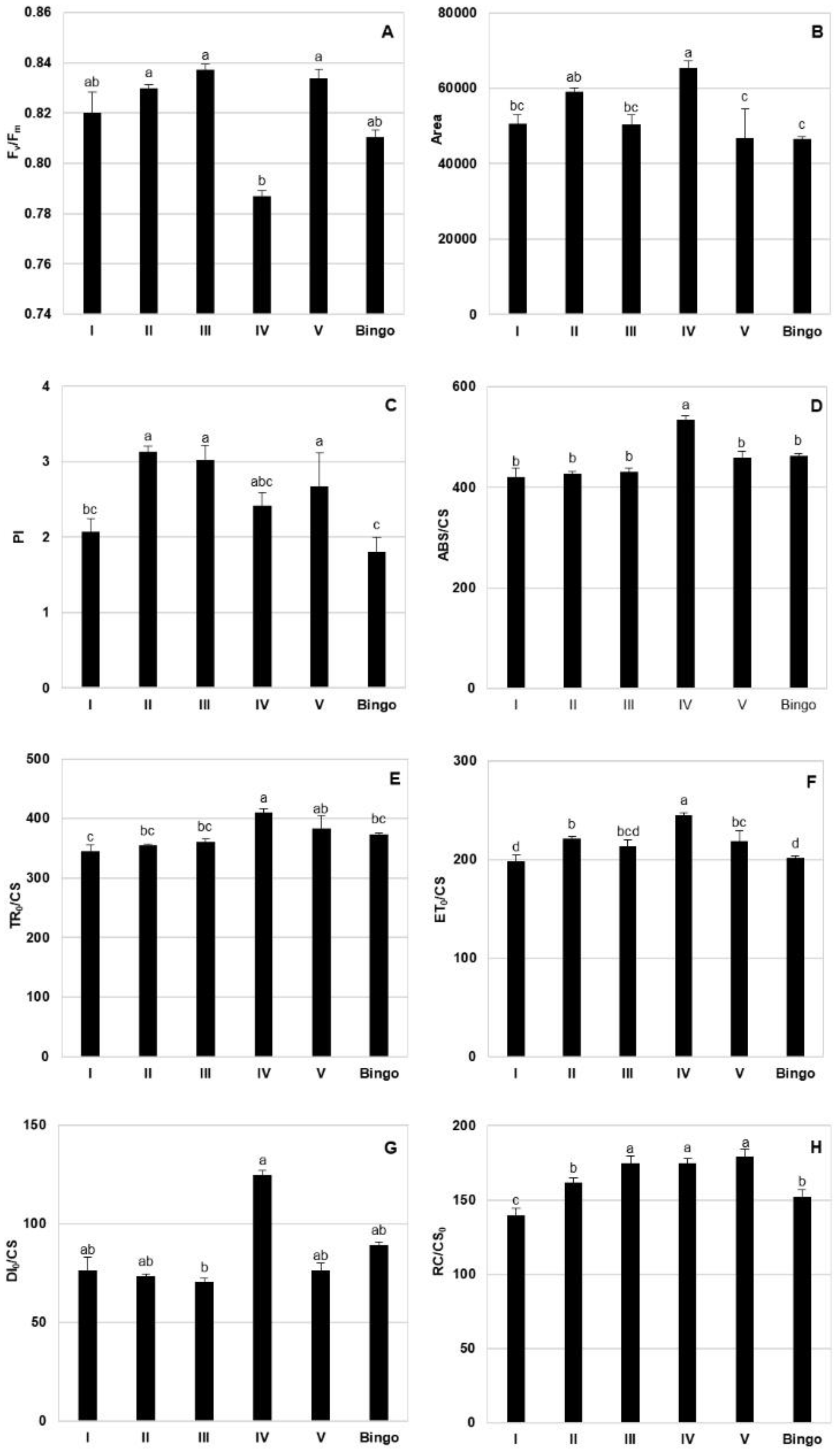
2.2. Comparison of OMA Lines in Control Conditions
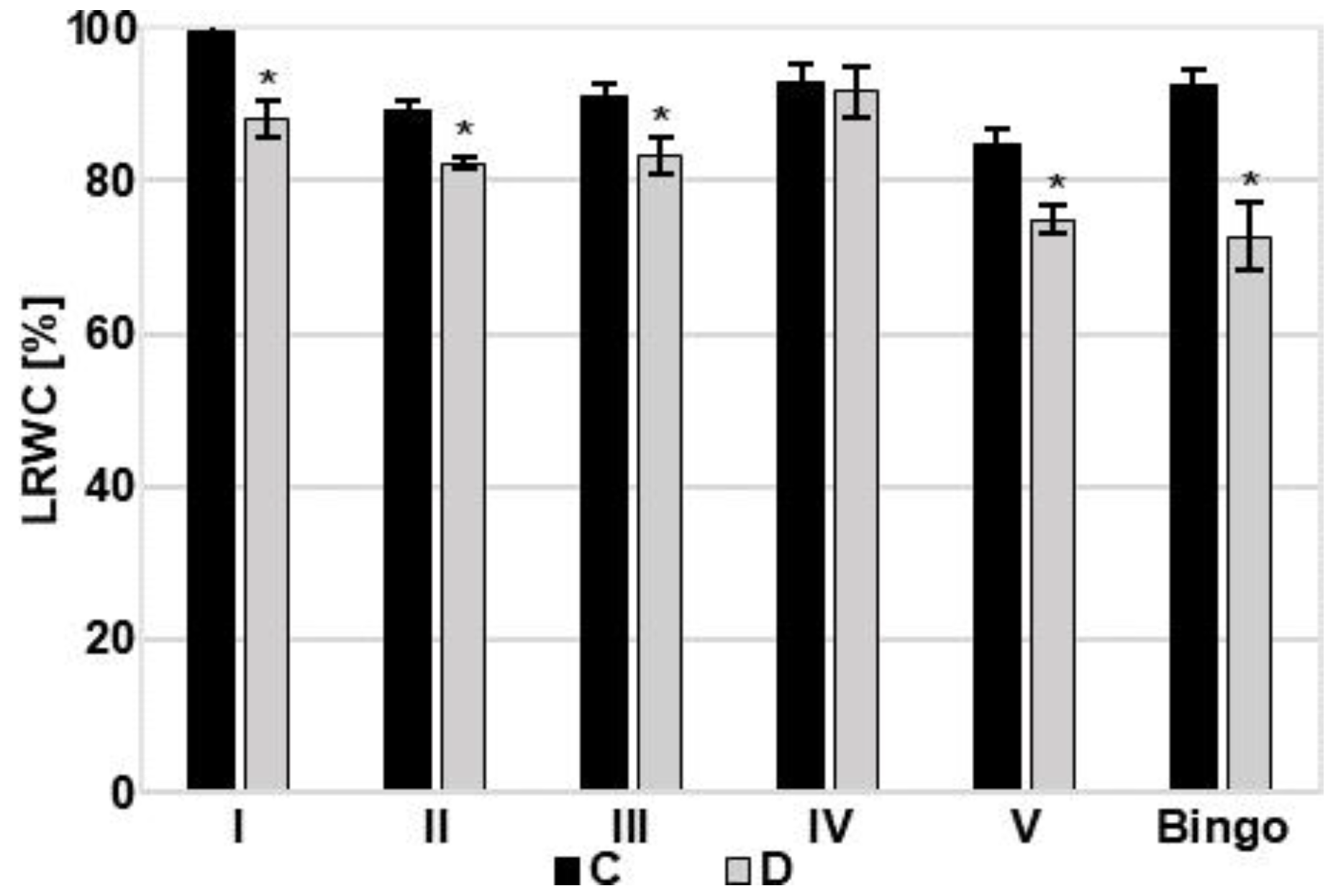
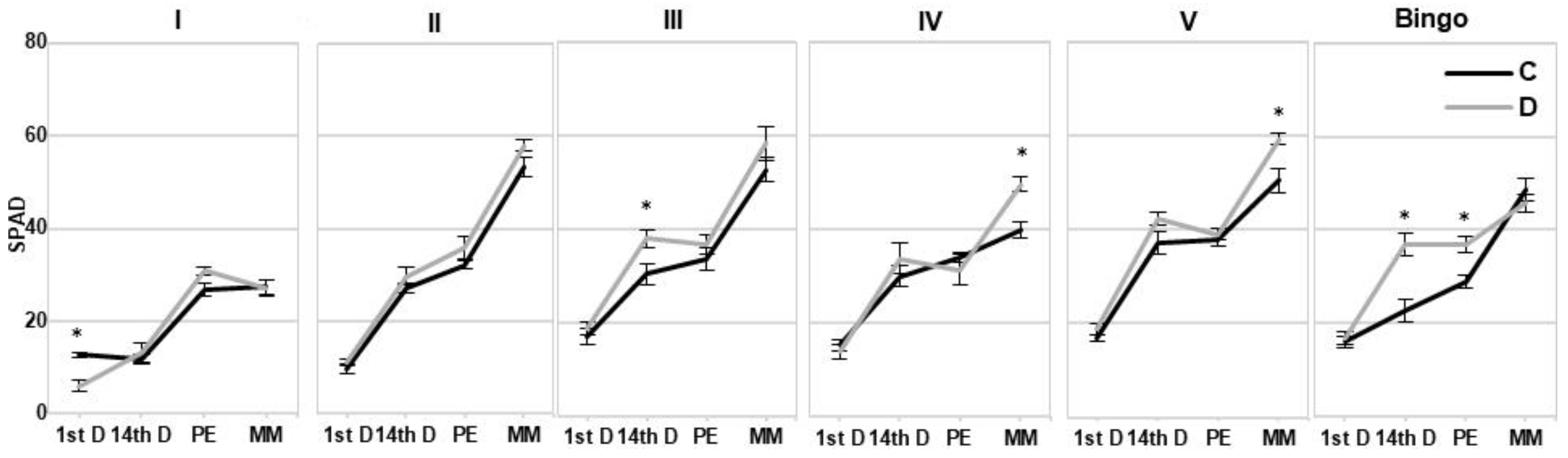
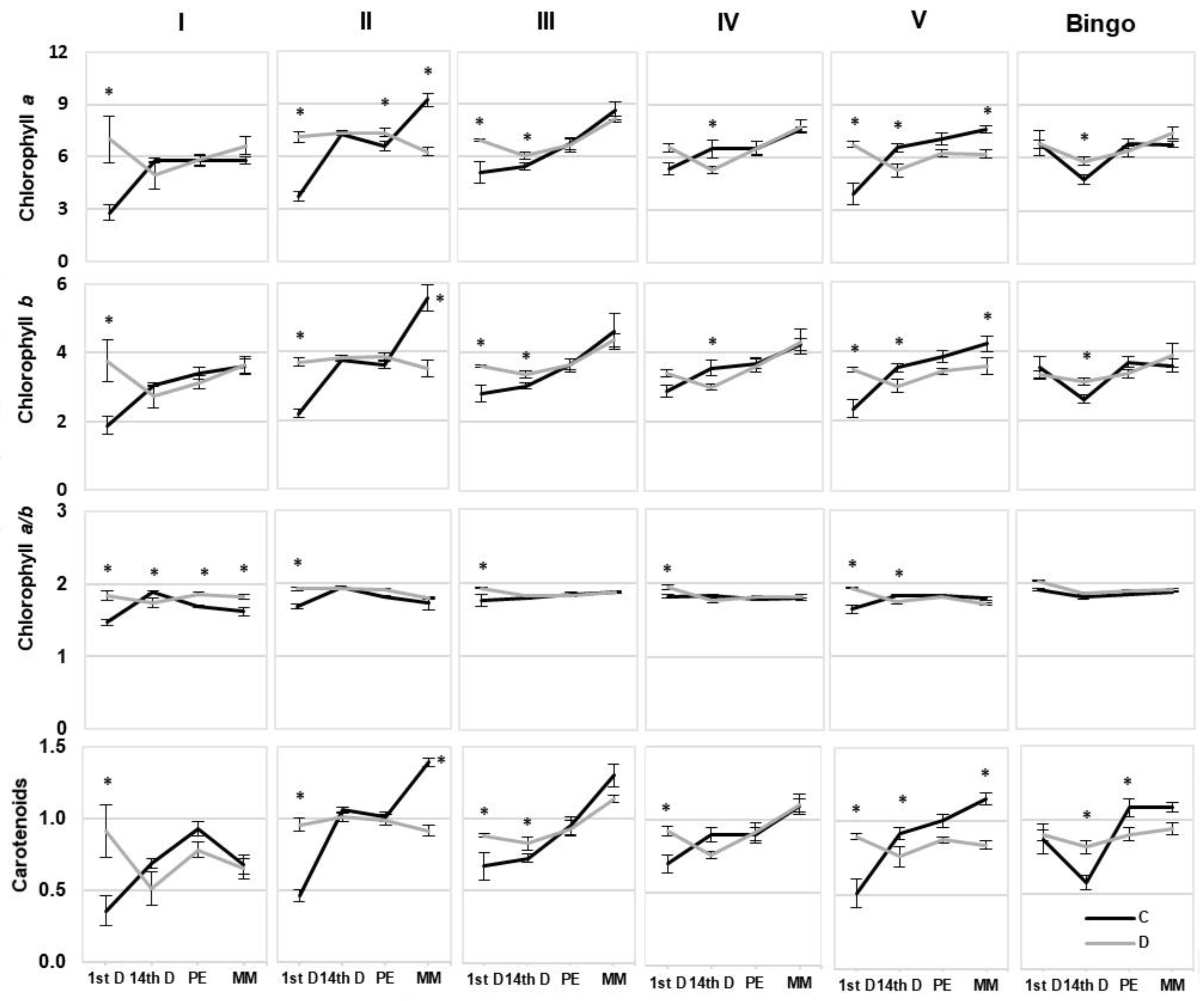
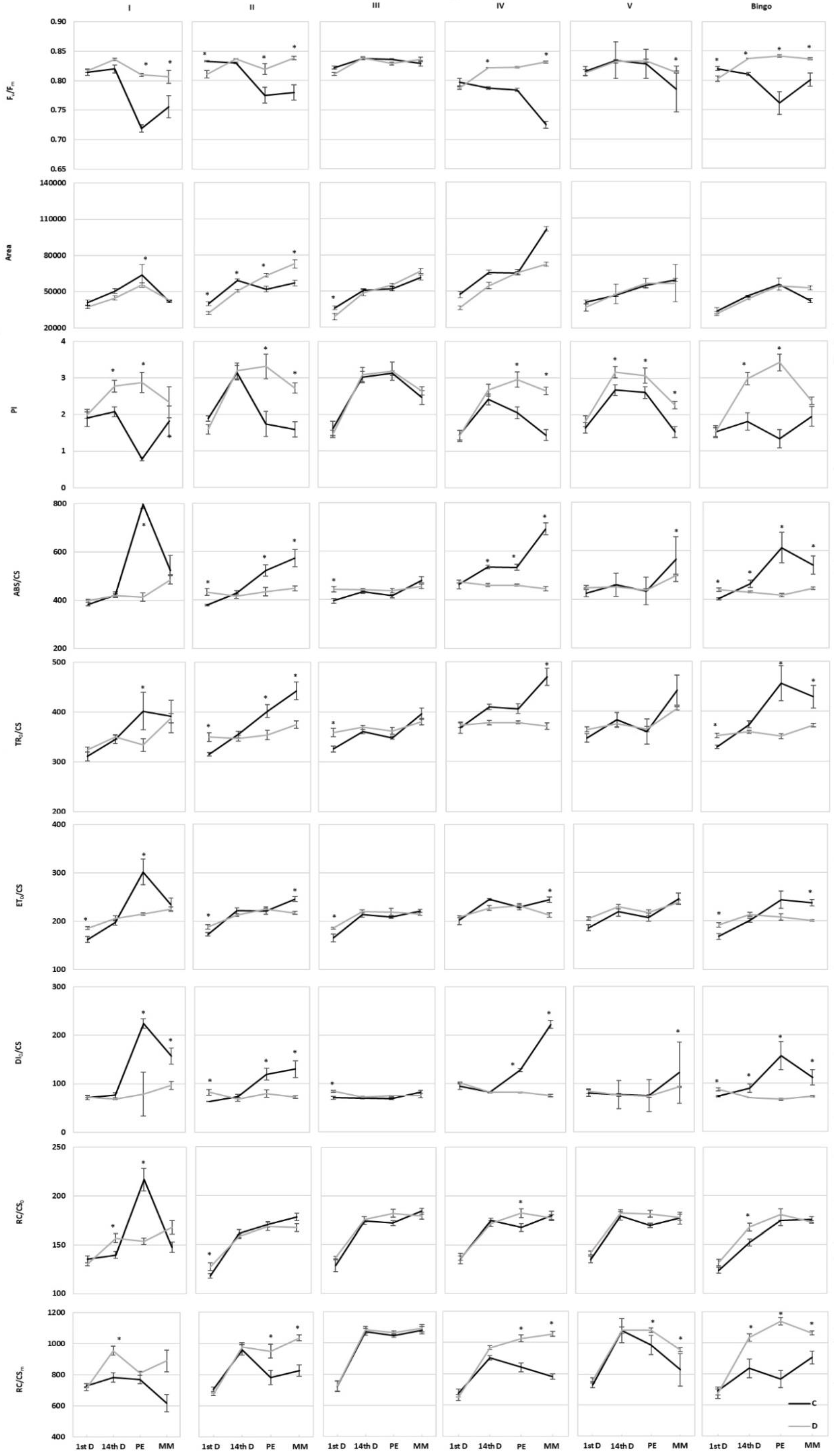
2.3. Assessment of OMA Lines in Drought Condition
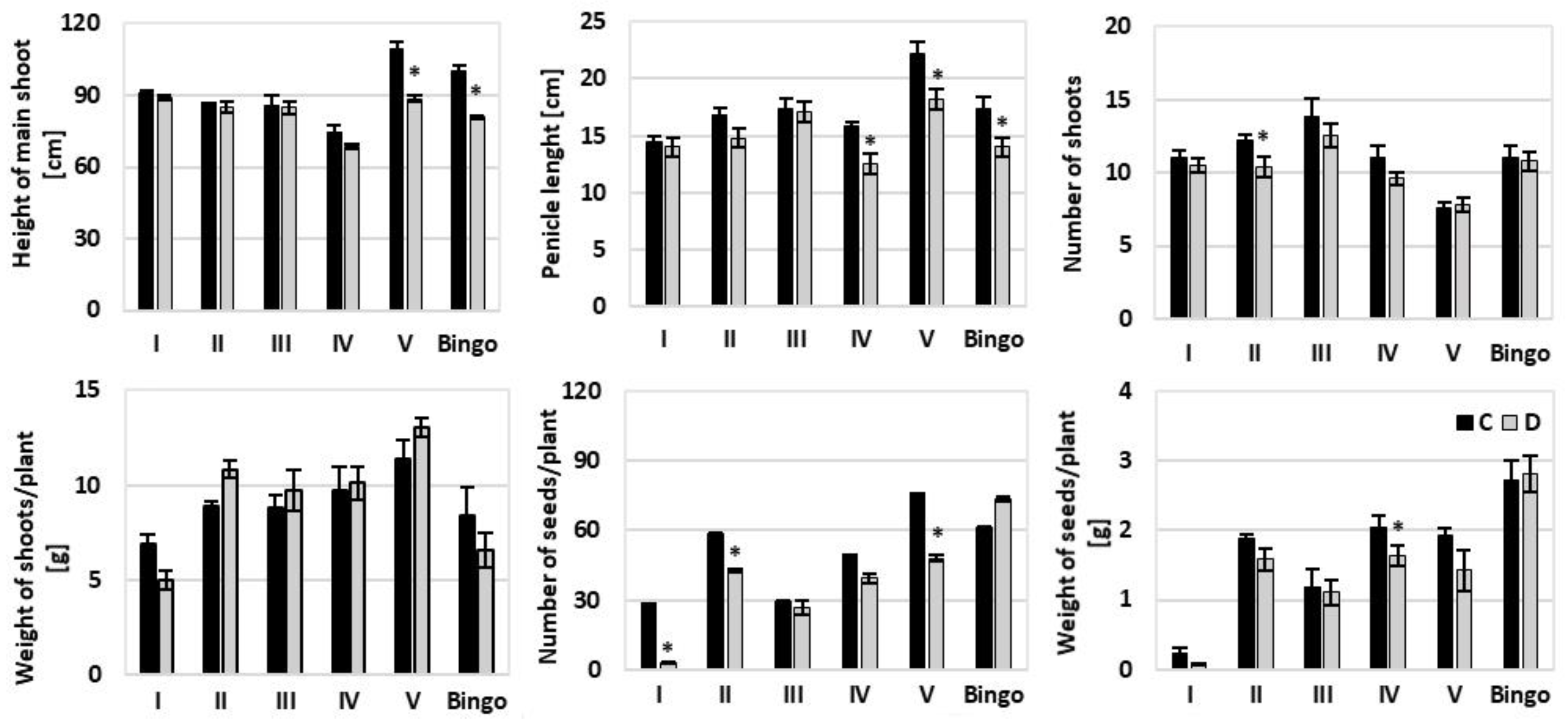
2.4. Morphological Characteristics and Yield Formation
3. Discussion
3.1. Functioning of the OMA Lines Photosynthetic Apparatus in Optimal Hydration
3.2. Functioning of the OMA Line Photosynthetic Apparatus in Drought Conditions
3.3. Morphology and Yield Components of OMA Lines
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Identification of Oat × Maize Hybrids
4.3. Mitotic Chromosome Preparations
4.4. Probe Labelling and in Situ Hybridization
4.5. Maize Chromosome Identification
4.6. Leaf Relative Water Content
4.7. Chlorophyll Concentration in Leaves
4.8. Photosynthetic Pigments
4.9. Chlorophyll A Fluorescence Parameters
4.10. Yield Component Analysis
4.11. Statistical Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABS/CS | Light Energy Absorption |
| Area | Area over the chlorophyll a fluorescence induction curve |
| Car | Carotenoids |
| CF | Chlorophyll Fluorescence |
| Chl | Chlorophyll |
| DI0/CS | Energy dissipated from PSII |
| DM | Dry Mass |
| ET0/CS | Energy used for electron transport |
| FM | Fresh Mass |
| Fv/Fm | Maximum photochemical efficiency of PSII |
| FWC | Field Water Capacity |
| LRWC | Leaf Relative Water Content |
| OMA | Oat × maize chromosome addition line |
| PCA | Principal Components Analysis |
| PC | Principal Component |
| PI | Overall performance index of PSII photochemistry |
| RC/CSm | Number of active reaction centers in the relaxation state of leaf fragment |
| RC/CS0 | Number of active reaction centers in the excited leaf fragment |
| SPAD | Soil Plant Analysis Development |
| TR0/CS | Excitation energy trapped in PSII reaction centers |
References
- Rines, H.W.; Phillips, R.L.; Kynast, R.G.; Okagaki, R.J.; Galatowitsch, M.W.; Huettl, P.A.; Stec, A.O.; Jacobs, M.S.; Suresh, J.; Porter, H.L.; et al. Addition of individual chromosomes of maize inbreds B73 and Mo17 to oat cultivars Starter and Sun II: Maize chromosome retention, transmission, and plant phenotype. Theor. Appl. Genet. 2009, 119, 1255–1264. [Google Scholar] [CrossRef]
- Ananiev, E.V.; Phillips, R.L.; Rines, H.W. Chromosome-specific molecular organization of maize (Zea mays L.) centromeric regions. Proc. Natl. Acad. Sci. USA 1998, 95, 13073–13078. [Google Scholar] [CrossRef]
- Bass, H.W.; Riera-Lizarazu, O.; Ananiev, E.V.; Bordoli, S.J.; Rines, H.W.; Phillips, R.L.; Sedat, J.W.; Agard, D.A.; Cande, W.Z. Evidence for the coincident initiation of homolog pairing and synapsis during the telomere-clustering (bouquet) stage of meiotic prophase. J. Cell Sci. 2000, 113, 1033–1042. [Google Scholar] [PubMed]
- Amarillo, F.I.E.; Bass, H.W. A transgenomic cytogenetic sorghum (Sorghum propinquum) bacterial artificial chromosome fluorescence in situ hybridization map of maize (Zea mays L.) pachytene chromosome 9, evidence for regions of genome hyperexpansion. Genetics 2007, 177, 1509–1526. [Google Scholar] [CrossRef] [PubMed]
- Li, L.J.; Arumuganathan, K.; Rines, H.W.; Phillips, R.L.; Riera-Lizarazu, O.; Sandhu, D.; Zhou, Y.; Gill, K.S. Flow cytometric sorting of maize chromosome 9 from an oat-maize chromosome addition line. Theor. Appl. Genet. 2001, 102, 658–663. [Google Scholar] [CrossRef]
- Muehlbauer, G.J.; Riera-Lizarazu, O.; Kynast, R.G.; Martin, D.; Phillips, R.L.; Rines, H.W. A maize chromosome 3 addition line of oat exhibits expression of the maize homeobox gene liguleless3 and alteration of cell fates. Genome 2000, 43, 1055–1064. [Google Scholar] [CrossRef]
- Kowles, R.V.; Walch, M.D.; Minnerath, J.M.; Bernacchi, C.J.; Stec, A.O.; Rines, H.W.; Phillips, R.L. Expression of C4 photosynthetic enzymes in oat-maize chromosome addition lines. Maydica 2008, 53, 69–78. [Google Scholar]
- Walch, M.D. Expression of Maize Pathogenesis-Related and Photosynthetic Genes in Oat × Maize Addition Lines. Ph.D. Thesis, University of Minnesota, Minneapolis, Saint Paul, MN, USA, 2007. [Google Scholar]
- Kynast, R.G.; Riera-Lizarazu, O.; Vales, M.I.; Okagaki, R.J.; Maquieira, S.B.; Chen, G.; Ananiev, E.V.; Odland, W.E.; Russell, C.D.; Stec, A.O.; et al. A complete set of maize individual chromosome additions to the oat genome. Plant Physiol. 2001, 125, 1216–1227. [Google Scholar] [CrossRef][Green Version]
- Dong, Z.; Yu, J.; Li, H.; Huang, W.; Xu, L.; Zhao, Y.; Zhang, T.; Xu, W.; Jiang, J.; Su, Z.; et al. Transcriptional and epigenetic adaptation of maize chromosomes in Oat-Maize addition lines. Nucleic Acids Res. 2018, 46, 5012–5028. [Google Scholar] [CrossRef]
- Rines, H.W.; Phillips, R.L.; Kynast, R.G.; Okagaki, R.; Odland, W.E.; Stec, A.O.; Jacobs, M.S.; Granath, S.R. Maize chromosome additions and radiation hybrids in oat and their use in dissecting the maize genome. In The Wake of the Double Helix: From the Green Revolution to the Gene Revolution, Proceedings of the International Congress, Bologna, Italy, 27–31 May 2003; University of Bologna: Bologna, Italy, 2005; pp. 427–441. [Google Scholar]
- Jastrzebska, M.; Kostrzewska, M.; Wanic, M. Wpływ deficytu wody i interakcji międzygatunkowych na wybrane parametry fizjologiczne roślin jęczmienia jarego i koniczyny czerwonej. Fragm. Agron. 2016, 33, 44–59. [Google Scholar]
- Lawlor, D.W.; Tezara, W. Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: A critical evaluation of mechanisms and integration of processes. Ann. Bot. 2009, 103, 561–579. [Google Scholar] [CrossRef] [PubMed]
- Akıncı, Ş.; Lösel, D.M. Plant water-stress response mechanisms. In Water Stress; Rahman, I.M.M., Hasegawa, H., Eds.; InTech: Rijeka, Croatia, 2012; pp. 15–42. [Google Scholar]
- Ji, X.; Shiran, B.; Wan, J.; Lewis, D.C.; Jenkins, C.L.D.; Condon, A.G.; Richards, R.A.; Dolferus, R. Importance of pre-anthesis anther sink strength for maintenance of grain number during reproductive stage water stress in wheat. Plant Cell Environ. 2010, 33, 926–942. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.E.; Chaitanya, K.V. Photosynthesis and antioxidative defense mechanisms in deciphering drought stress tolerance of crop plants. Biol. Plant. 2016, 60, 201–218. [Google Scholar] [CrossRef]
- Yan, K.; Chen, P.; Shao, H.; Shao, C.; Zhao, S.; Brestic, M. Dissection of photosynthetic electron transport process in sweet sorghum under heat stress. PLoS ONE 2013, 8, e62100. [Google Scholar] [CrossRef]
- Peng, Y.; Nguy-Robertson, A.; Arkebauer, T.; Gitelson, A.A. Assessment of canopy chlorophyll content retrieval in maize and soybean: Implications of hysteresis on the development of generic algorithms. Remote Sens. 2017, 9, 226. [Google Scholar] [CrossRef]
- Demirevska, K.; Zasheva, D.; Dimitrov, R.; Simova-Stoilova, L.; Stamenova, M.; Feller, U. Drought stress effects on Rubisco in wheat: Changes in the Rubisco large subunit. Acta Physiol. Plant. 2009, 31, 1129–1138. [Google Scholar] [CrossRef]
- Okagaki, R.J.; Kynast, R.G.; Livingston, S.M.; Russell, C.D.; Rines, H.W.; Phillips, R.L. Mapping maize sequences to chromosomes using oat-maize chromosome addition materials. Plant Physiol. 2001, 125, 1228–1235. [Google Scholar] [CrossRef]
- Riera-Lizarazu, O.; Rines, H.W.; Phillips, R.L. Cytological and molecular characterization of oat × maize partial hybrids. Theor. Appl. Genet. 1996, 93, 123–135. [Google Scholar] [CrossRef]
- Phillips, R.L.; Rines, H.W. Genetic analyses with oat-maize addition and radiation hybrid lines. In Handbook of Maize: Genetics and Genomics; Bennetzen, J.L., Hake, S., Eds.; Springer Science + Business Media: New York, NY, USA, 2009; Volume 2, pp. 523–538. [Google Scholar]
- Cho, S.H.; Garvin, D.F.; Muehlbauer, G.J. Transcriptome analysis and physical mapping of barley genes in wheat-barley chromosome addition lines. Genetics 2006, 172, 1277–1285. [Google Scholar] [CrossRef]
- Noga, A.; Warchol, M.; Czyczylo-Mysza, I.; Marcinska, I.; Dziurka, K.; Warzecha, T.; Skrzypek, E. Chlorophyll a fluorescence parameters in the evaluation of oat DH lines yield components. Cereal Res. Commun. 2017, 45, 665–674. [Google Scholar] [CrossRef]
- Ludlow, M.; Muchow, R. A critical evaluation of traits for improving crop yields in water-limited environments. In Advances in Agronomy; Brady, N.C., Ed.; Academic Press: London, UK, 1990; Volume 43, pp. 107–153. [Google Scholar]
- Clarke, J.M.; McCaig, T.N. Evaluation of techniques for screening for drought resistance in wheat. Crop Sci. 1982, 22, 503–506. [Google Scholar] [CrossRef]
- Almeselmani, M.; Abdullah, F.; Hareri, F.; Naaesan, M.; Ammar, M.A.; ZuherKanbar, O.; Saud, A.A. Effect of drought on different physiological characters and yield component in different varieties of Syrian durum wheat. J. Agric. Sci. 2011, 3, 127. [Google Scholar] [CrossRef]
- Siddique, M.R.B.; Hamid, A.; Islam, M.S. Drought stress effects on water relations of wheat. Bot. Bull. Acad. Sin. 2000, 41, 35–39. [Google Scholar]
- Silva, M.D.A.; Jifon, J.L.; Silva, J.A.G.D.; Sharma, V. Use of physiological parameters as fast tools to screen for drought tolerance in sugarcane. Braz. J. Plant Physiol. 2007, 19, 193–201. [Google Scholar] [CrossRef]
- Keyvan, S. The effects of drought stress on yield, relative water content, proline, soluble carbohydrates and chlorophyll of bread wheat cultivars. J. Anim. Plant Sci. 2010, 8, 1051–1060. [Google Scholar]
- Juzon, K.; Czyczylo-Mysza, I.; Ostrowska, A.; Marcinska, I.; Skrzypek, E. Chlorophyll fluorescence for prediction of yellow lupin (Lupinus luteus L.) and pea (Pisum sativum L.) susceptibility to drought. Photosynthetica 2019, 57, 950–959. [Google Scholar] [CrossRef]
- Hsiao, T.C. Plant responses to water stress. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1973, 24, 519–570. [Google Scholar] [CrossRef]
- Mao, H.; Chen, M.; Su, Y.; Wu, N.; Yuan, M.; Yuan, S.; Brestic, M.; Zivcak, M.; Zhang, H.; Chen, Y. Comparison on photosynthesis and antioxidant defense systems in wheat with different ploidy levels and octoploid Triticale. Int. J. Mol. Sci. 2018, 19, 3006. [Google Scholar] [CrossRef]
- Long, S.P.; Humphries, S.; Falkowski, P.G. Photoinhibition of photosynthesis in nature. Annu. Rev. Plant Biol. 1994, 45, 633–662. [Google Scholar] [CrossRef]
- Dale, J.E. The control of leaf expansion. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1988, 39, 267–295. [Google Scholar] [CrossRef]
- O’Neill, P.M.; Shanahan, J.E.; Schepers, J.S. Use of chlorophyll fluorescence assessments to differentiate corn hybrid response to variable water conditions. Crop Sci. 2006, 46, 681–687. [Google Scholar] [CrossRef]
- Cassol, D.; De Silva, F.S.P.; Falqueto, A.R.; Bacarin, M.A. An evaluation of non-destructive methods to estimate total chlorophyll content. Photosynthetica 2008, 46, 634–636. [Google Scholar] [CrossRef]
- Huseynova, I.M.; Suleymanov, S.Y.; Rustamova, S.M.; Aliyev, J.A. Drought-induced changes in photosynthetic membranes of two wheat (Triticum aestivum L.) cultivars. Biochemistry (Moscow) 2009, 74, 903–909. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.-Y.; Wang, L.-C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Mafakheri, A.; Siosemardeh, A.; Bahramnejad, B.; Struik, P.C.; Sohrabi, E. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 4, 580–585. [Google Scholar]
- Din, J.; Khan, S.U.; Ali, I.; Gurmani, A.R. Physiological and agronomic response of canola varieties to drought stress. J. Anim. Plant Sci. 2011, 21, 78–82. [Google Scholar]
- Hamada, A.M.; Al-Hakimi, A.M.A. Salicylic acid versus salinity-drought-induced stress on wheat seedlings. Rostlinna Vyroba 2001, 47, 444–450. [Google Scholar]
- Pirzad, A.; Shakiba, M.R.; Zehtab-Salmasi, S.; Mohammadi, S.A.; Darvishzadeh, R.; Samadi, A. Effect of water stress on leaf relative water content, chlorophyll, proline and soluble carbohydrates in Matricaria chamomilla L. J. Med. Plant Res. 2011, 5, 2483–2488. [Google Scholar]
- Ashraf, M.; Karim, F. Screening of some cultivars/lines of black gram (Vigna mungo L. Hepper) for resistance to water stress. J. Trop. Agric. 1991, 68, 57–62. [Google Scholar]
- Campbell, P.K.E.; Huemmrich, K.F.; Middleton, E.M.; Ward, L.A.; Julitta, T.; Daughtry, C.S.T.; Burkart, A.; Russ, A.L.; Kustas, W.P. Diurnal and seasonal variations in chlorophyll fluorescence associated with photosynthesis at leaf and canopy scales. Remote Sens. 2019, 11, 448. [Google Scholar] [CrossRef]
- Jain, M.; Tiwary, S.; Gadre, R. Sorbitol-induced changes in various growth and biochemici parameters in maize. Plant Soil Environ. 2010, 56, 263–267. [Google Scholar] [CrossRef]
- Ashraf, M.Y.; Azmi, A.R.; Khan, A.H.; Ala, S.A. Effect of water stress on total phenol, peroxidase activity and chlorophyll contents in wheat (Triticum aestivum L.). Acta Physiol. Plant. 1994, 16, 185–191. [Google Scholar]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Rohacek, K. Chlorophyll fluorescence parameters: The definitions, photosynthetic meaning, and mutual relationships. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Dabrowski, P.; Baczewska-Dabrowska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M.; Wojcik-Jagla, M.; Pawluskiewicz, B.; Baba, W.; Brestic, M. Exploration of chlorophyll a fluorescence and plant gas exchange parameters as indicators of drought tolerance in perennial ryegrass. Sensors 2019, 19, 2736. [Google Scholar] [CrossRef]
- Li, R.-H.; Guo, P.-G.; Michael, B.; Stefania, G.; Salvatore, C. Evaluation of chlorophyll content and fluorescence parameters as indicators of drought tolerance in barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Kalaji, M.; Guo, P. Chlorophyll fluorescence: A useful tool in barley plant breeding programs. In Photochemistry Research Progress; Sánchez, A., Gutierrez, S.J., Eds.; Nova Science Publishers: New York, NY, USA, 2008; Volume 29, pp. 439–463. [Google Scholar]
- Tsonev, T.; Wahbi, S.; Sun, P.; Sorrentino, G.; Centritto, M. Gas exchange, water relations photochemical reflectance index in stress and recovery and their relationships with Quercus ilex plants during water stress and recovery. Int. J. Agric. Biol. 2014, 16, 335–341. [Google Scholar]
- Marcinska, I.; Czyczylo-Mysza, I.; Skrzypek, E.; Grzesiak, M.T.; Popielarska-Konieczna, M.; Warchol, M.; Grzesiak, S. Application of photochemical parameters and several indices based on phenotypical traits to assess intraspecific variation of oat (Avena sativa L.) tolerance to drought. Acta Physiol. Plant. 2017, 39. [Google Scholar] [CrossRef]
- Liang, Y.; Zhang, K.; Zhao, L.; Liu, B.; Meng, Q.; Tian, J.; Zhao, S. Identification of chromosome regions conferring dry matter accumulation and photosynthesis in wheat (Triticum aestivum L.). Euphytica 2010, 171, 145–156. [Google Scholar] [CrossRef]
- Qiu, N.; Lu, C. Enhanced tolerance of photosynthesis against high temperature damage in salt-adapted halophyte Atriplex centralasiatica plants. Plant Cell Environ. 2003, 26, 1137–1145. [Google Scholar] [CrossRef]
- Ohashi, Y.; Nakayama, N.; Saneoka, H.; Fujita, K. Effects of drought stress on photosynthetic gas exchange, chlorophyll fluorescence and stem diameter of soybean plants. Biol. Plant. 2006, 50, 138–141. [Google Scholar] [CrossRef]
- Antonkiewicz, J.; Rapacz, M. Assessment of photosynthetic activity of plants grown on stubble sediments and furnace ash. Zesz. Probl. Post. Nauk Rol. 2006, 509, 187–196. (In Polish) [Google Scholar]
- Souza, R.P.; Machado, E.C.; Silva, J.A.B.; Lagoa, A.; Silveira, J.A.G. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environ. Exp. Bot. 2004, 51, 45–56. [Google Scholar] [CrossRef]
- Fghire, R.; Anaya, F.; Ali, O.I.; Benlhabib, O.; Ragab, R.; Wahbi, S. Physiological and photosynthetic response of quinoa to drought stress. Chil. J. Agric. Res. 2015, 75, 174–183. [Google Scholar] [CrossRef]
- Czyczylo-Mysza, I.; Tyrka, M.; Marcinska, I.; Skrzypek, E.; Karbarz, M.; Dziurka, M.; Hura, T.; Dziurka, K.; Quarrie, S.A. Quantitative trait loci for leaf chlorophyll fluorescence parameters, chlorophyll and carotenoid contents in relation to biomass and yield in bread wheat and their chromosome deletion bin assignments. Mol. Breed. 2013, 32, 189–210. [Google Scholar] [CrossRef]
- Tolley, B.; Sage, T.; Langdale, J.; Hibberd, J. Individual maize chromosomes in the C3 plant oat can increase bundle sheath cell size and vein density. Plant Physiol. 2012, 159, 1418–1427. [Google Scholar] [CrossRef]
- Yang, X.; Chen, X.; Ge, Q.; Li, B.; Tong, Y.; Li, Z.; Kuang, T.; Lu, C. Characterization of photosynthesis of flag leaves in a wheat hybrid and its parents grown under field conditions. J. Plant Physiol. 2007, 164, 318–326. [Google Scholar] [CrossRef]
- Langdale, J.A. C-4 cycles: Past, present, and future research on C-4 photosynthesis. Plant Cell 2011, 23, 3879–3892. [Google Scholar] [CrossRef]
- Skrzypek, E.; Warzecha, T.; Noga, A.; Warchol, M.; Czyczylo-Mysza, I.; Dziurka, K.; Marcinska, I.; Kaploniak, K.; Sutkowska, A.; Nita, Z.; et al. Complex characterization of oat (Avena sativa L.) lines obtained by wide crossing with maize (Zea mays L.). PeerJ 2018, 6, e5107. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, G.; Hasterok, R. BAC ‘landing’ on chromosomes of Brachypodium distachyon for comparative genome alignment. Nat. Protoc. 2007, 2, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Idziak-Helmcke, D.; Warzecha, T.; Sowa, M.; Warchoł, M.; Dziurka, K.; Czyczyło-Mysza, I.; Skrzypek, E. 3-D nucleus architecture in oat × maize addition lines. Int. J. Mol. Sci. 2020, 21, 4280. [Google Scholar] [CrossRef] [PubMed]
- Ober, E.S.; Le Bloa, M.; Clark, C.J.A.; Royal, A.; Jaggard, K.W.; Pidgeon, J.D. Evaluation of physiological traits as indirect selection criteria for drought tolerance in sugar beet. Field Crop Res. 2005, 91, 231–249. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Wang, B.; Ma, M.; Lu, H.; Meng, Q.; Li, G.; Yang, X. Photosynthesis, sucrose metabolism, and starch accumulation in two NILs of winter wheat. Photosynth. Res. 2015, 126, 363–373. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype/Cultivar | Origin | No. of Maize Chromosome/Chromatin Added to Oat Genome | Chromosome ID |
|---|---|---|---|
| I | DC 06011-6 × POB 722 | 4 | 1, 2 |
| II | STH 9511 × Bingo | 2 | 2 |
| III | STH 8-50 × Canyon | 1 | 3 |
| IV | STH 9110 × Contender | 2 | 5 |
| V | STH 93-61 × DC 2112/07 | Chromatin | 6 |
| Bingo | - | - | - |
| Source of Variance | Trait | F |
|---|---|---|
| Total chlorophyll content | SPAD | 18.762 *** |
| Photosynthetic pigment content | Chla | 10.692 *** |
| Chlb | 13.243 *** | |
| Chla/b | 6.120 *** | |
| Car | 21.941 *** | |
| Chlorophyll a fluorescence parameters | Fv/Fm | 2.250 ns |
| Area | 4.226 ** | |
| PI | 4.786 ** | |
| ABS/CS | 3.307 * | |
| TR0/CS | 5.004 *** | |
| ET0/CS | 7.308 *** | |
| DI0/CS | 1.800 ns | |
| RC/CS0 | 13.360 *** | |
| RC/CSm | 5.979 * |
| Trait/Interaction | Degrees of Freedom | LRWC | SPAD | Chla | Chlb | Chla/b | Car |
|---|---|---|---|---|---|---|---|
| Genotype | 5 (k − 1) | 1.837 ns | 54.415 *** | 9.746 *** | 8.389 *** | 19.605 *** | 16.285 *** |
| Treatment | 1 (t − 1) | 6.568 * | 28.666 *** | 9.892 ** | 1.549 ns | 50.340 *** | 0.073 ns |
| Term | 3 (n − 1) | - | 539.706 *** | 46.231 *** | 71.463 *** | 1.921 ns | 52.673 *** |
| Genotype × Treatment | 5 (k − 1) × (t − 1) | 0.898 ns | 1.375 ns | 2.158 ns | 1.537 ns | 3.595 *** | 0.832 ns |
| Genotype × Term | 5 (k − 1) × (n − 1) | - | 9.252 *** | 4.751 *** | 3.540 ** | 6.135 *** | 5.587 *** |
| Treatment × Term | 1 (t − 1) × (n − 1) | - | 5.803 *** | 33.310 *** | 28.344 *** | 34.415 *** | 36.859 *** |
| Genotype × Treatment × Term | 5 (k − 1) × (t − 1) × (n − 1) | - | 2.331 ** | 6.411 *** | 7.329 *** | 3.105 *** | 4.861 *** |
| Trait/Interaction | Fv/Fm | Area | PI | ABS/CS | TR0/CS | ET0/CS | DI0/CS | RC/CS0 | RC/CSm |
|---|---|---|---|---|---|---|---|---|---|
| Genotype | 12.326 *** | 29.475 *** | 6.750 *** | 7.438 *** | 6.853 *** | 10.090 *** | 8.317 *** | 15.872 *** | 26.191 *** |
| Treatment | 51.637 *** | 5.143 * | 86.553 *** | 50.093 *** | 52.487 *** | 5.193 * | 44.822 *** | 0.581 ns | 71.659 *** |
| Term | 11.938 *** | 111.585 *** | 65.181 *** | 26.533 *** | 49.481 *** | 115.331 *** | 12.478 *** | 335.934 *** | 150.815 *** |
| Genotype × Treatment | 5.798 *** | 6.012 *** | 4.742 *** | 6.833 *** | 7.777 *** | 4.684 *** | 6.085 *** | 3.817 *** | 5.412 *** |
| Genotype × Term | 2.841 *** | 6.401 *** | 2.896 *** | 2.856 *** | 4.181 *** | 4.769 *** | 2.118 ** | 5.566 *** | 5.929 *** |
| Treatment × Term | 17.652 *** | 2.904 * | 16.365 *** | 22.345 *** | 29.979 *** | 22.270 *** | 15.271 *** | 3.851 ** | 13.236 *** |
| Genotype × Treatment × Term | 2.381 ** | 2.876 *** | 2.408 ** | 3.062 *** | 3.964 *** | 3.319 *** | 2.347 ** | 4.095 *** | 2.397 ** |
| Source of Variation | Degrees of Freedom | Height of Main Shoot | Panicle Length | Number of Shoots | Weight of Shoots/Plant | Number of Seeds/Plant | Weight of Seeds/Plant |
|---|---|---|---|---|---|---|---|
| Genotype | 5 (k − 1) | 34.699 *** | 21.807 *** | 14.157 *** | 39.379 *** | 3.030 * | 13.536 *** |
| Treatment | 1 (t − 1) | 49.988 *** | 31.642 *** | 4.823 * | 26.811 *** | 18.039 *** | 0.121 ns |
| Genotype × Treatment | 5 (k − 1) × (t − 1) | 7.548 *** | 2.603 *** | 0.715 ns | 8.836 *** | 0.397 ns | 2.831 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Juzoń, K.; Idziak-Helmcke, D.; Rojek-Jelonek, M.; Warzecha, T.; Warchoł, M.; Czyczyło-Mysza, I.; Dziurka, K.; Skrzypek, E. Functioning of the Photosynthetic Apparatus in Response to Drought Stress in Oat × Maize Addition Lines. Int. J. Mol. Sci. 2020, 21, 6958. https://doi.org/10.3390/ijms21186958
Juzoń K, Idziak-Helmcke D, Rojek-Jelonek M, Warzecha T, Warchoł M, Czyczyło-Mysza I, Dziurka K, Skrzypek E. Functioning of the Photosynthetic Apparatus in Response to Drought Stress in Oat × Maize Addition Lines. International Journal of Molecular Sciences. 2020; 21(18):6958. https://doi.org/10.3390/ijms21186958
Chicago/Turabian StyleJuzoń, Katarzyna, Dominika Idziak-Helmcke, Magdalena Rojek-Jelonek, Tomasz Warzecha, Marzena Warchoł, Ilona Czyczyło-Mysza, Kinga Dziurka, and Edyta Skrzypek. 2020. "Functioning of the Photosynthetic Apparatus in Response to Drought Stress in Oat × Maize Addition Lines" International Journal of Molecular Sciences 21, no. 18: 6958. https://doi.org/10.3390/ijms21186958
APA StyleJuzoń, K., Idziak-Helmcke, D., Rojek-Jelonek, M., Warzecha, T., Warchoł, M., Czyczyło-Mysza, I., Dziurka, K., & Skrzypek, E. (2020). Functioning of the Photosynthetic Apparatus in Response to Drought Stress in Oat × Maize Addition Lines. International Journal of Molecular Sciences, 21(18), 6958. https://doi.org/10.3390/ijms21186958