Effect of Seminal Plasma Protein Fractions on Stallion Sperm Cryopreservation

 ,
,
Abstract

1. Introduction
2. Results
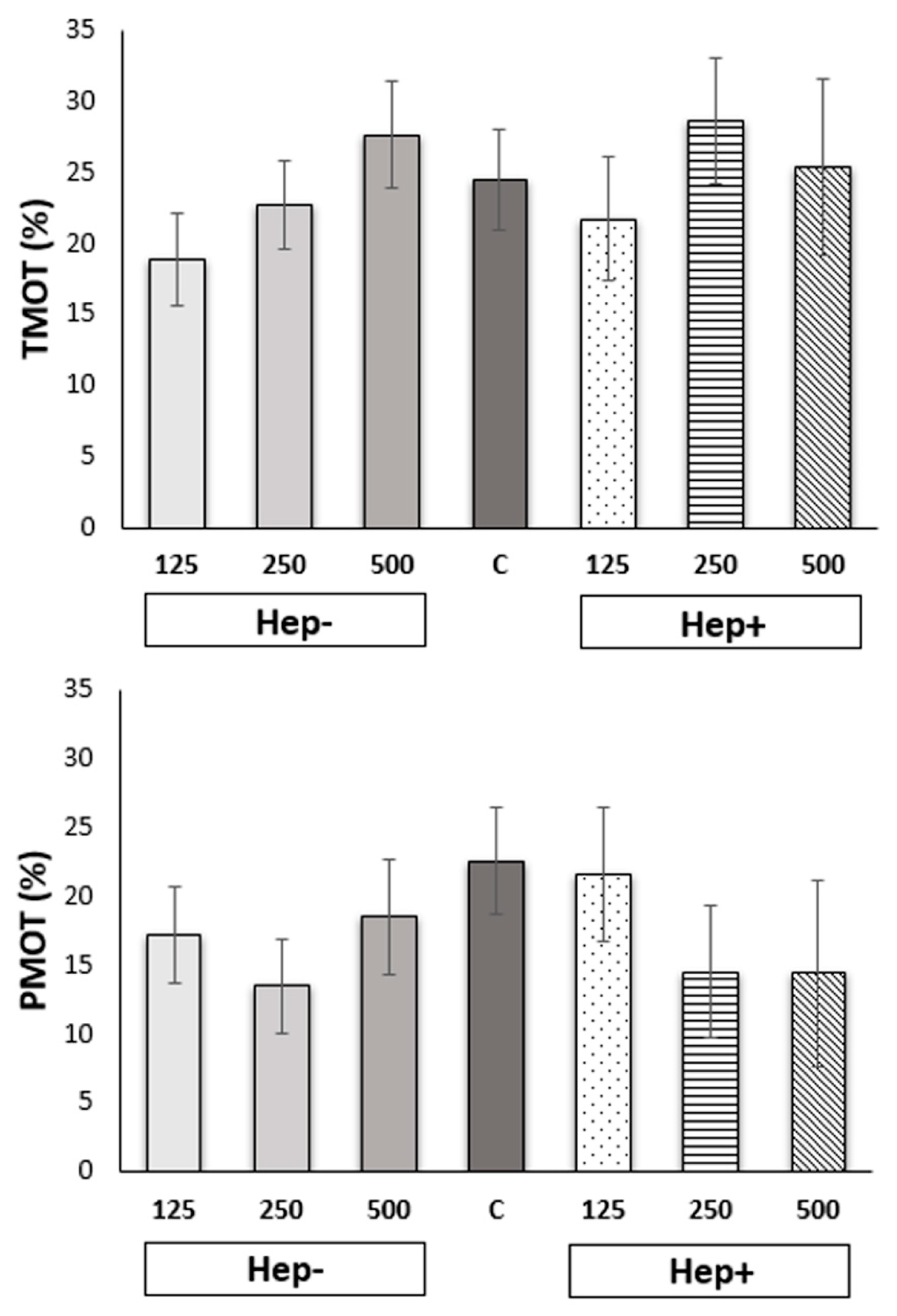
2.1. Effect of SP Protein Fractions on Stallion Sperm Motility and Kinematic Parameters
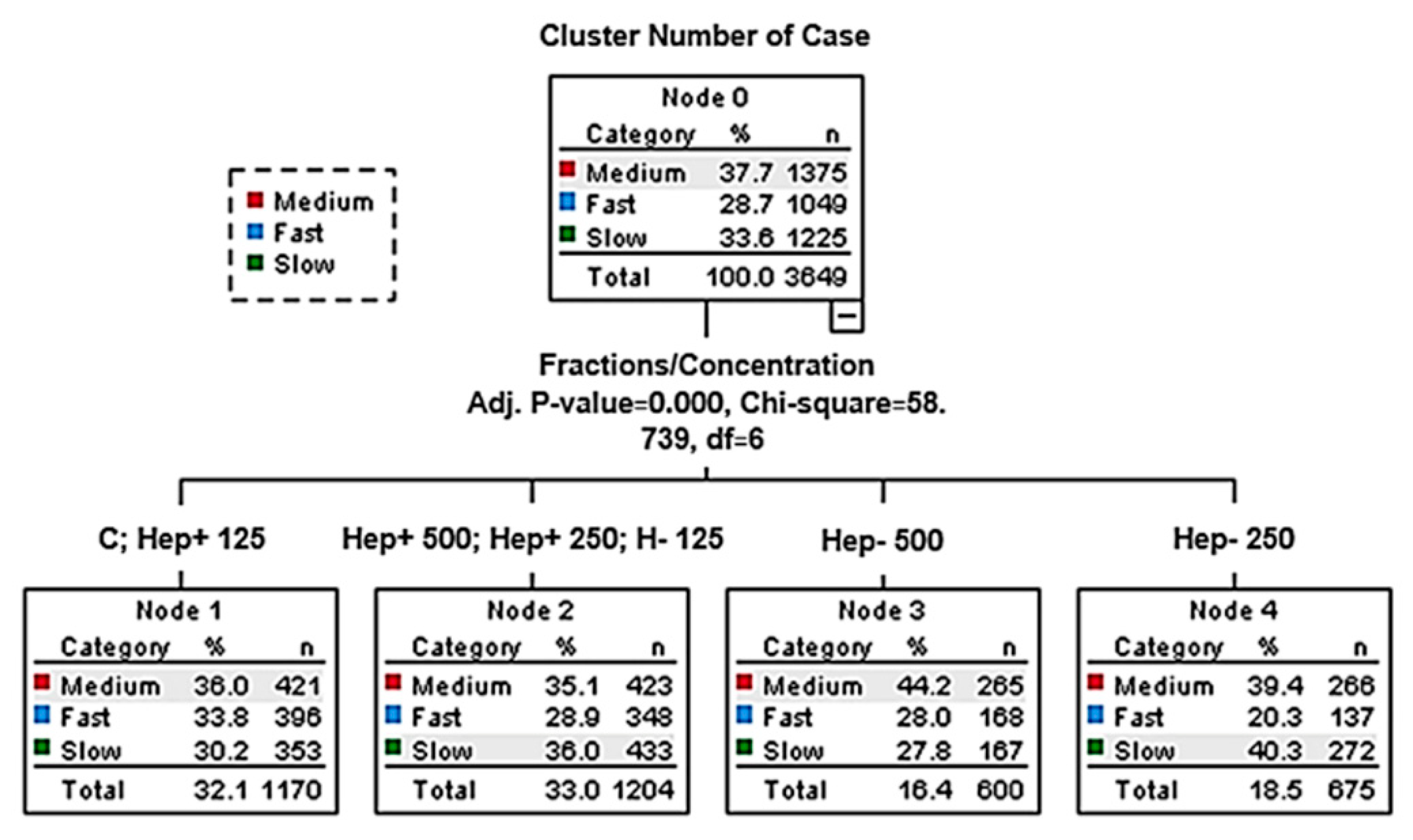
2.2. Effect of SP Protein Fractions on the Distribution of Motile Spermatozoa to Subpopulations
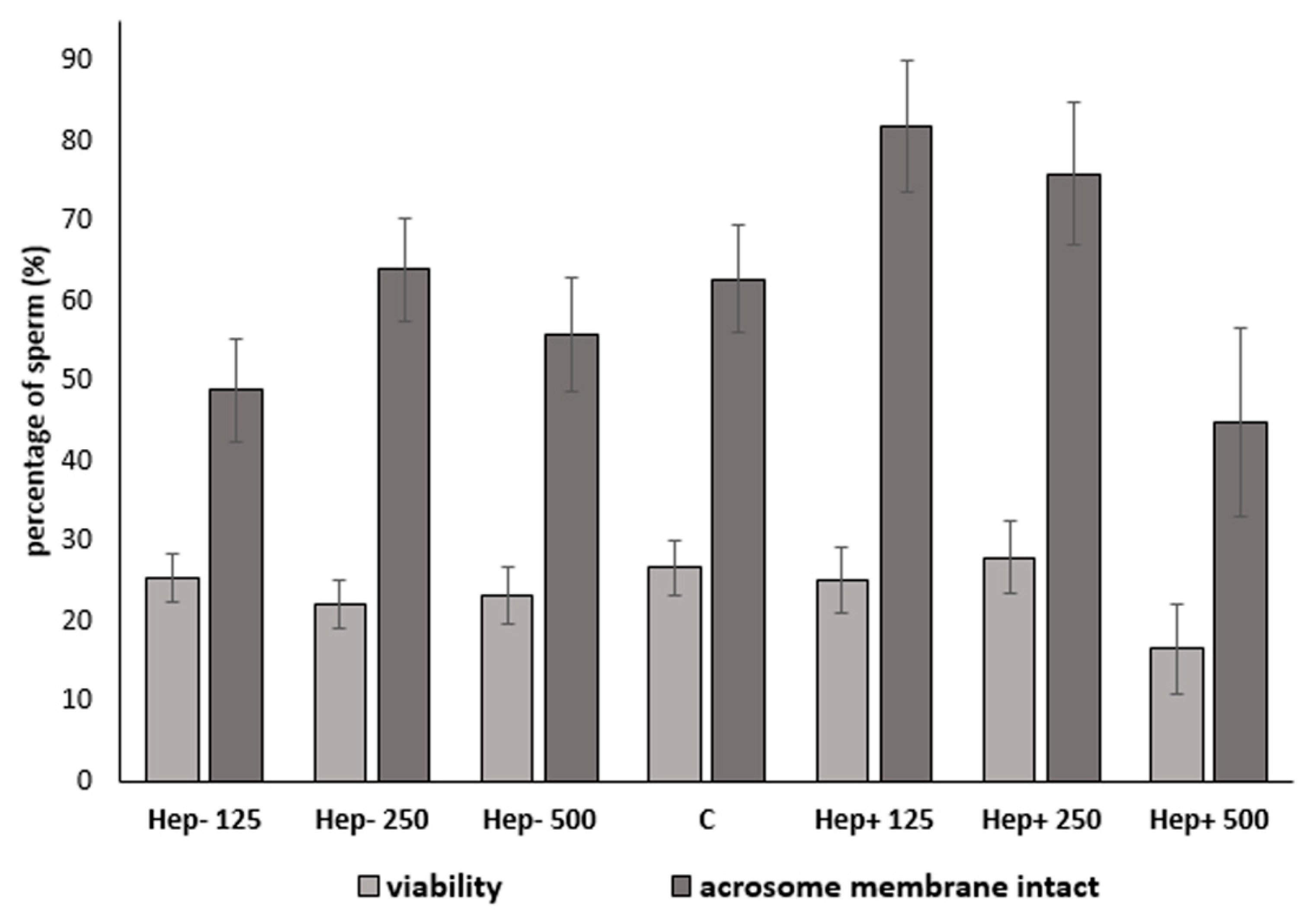
2.3. The Effect of SP Protein Fractions on Viability and Acrosomal Integrity
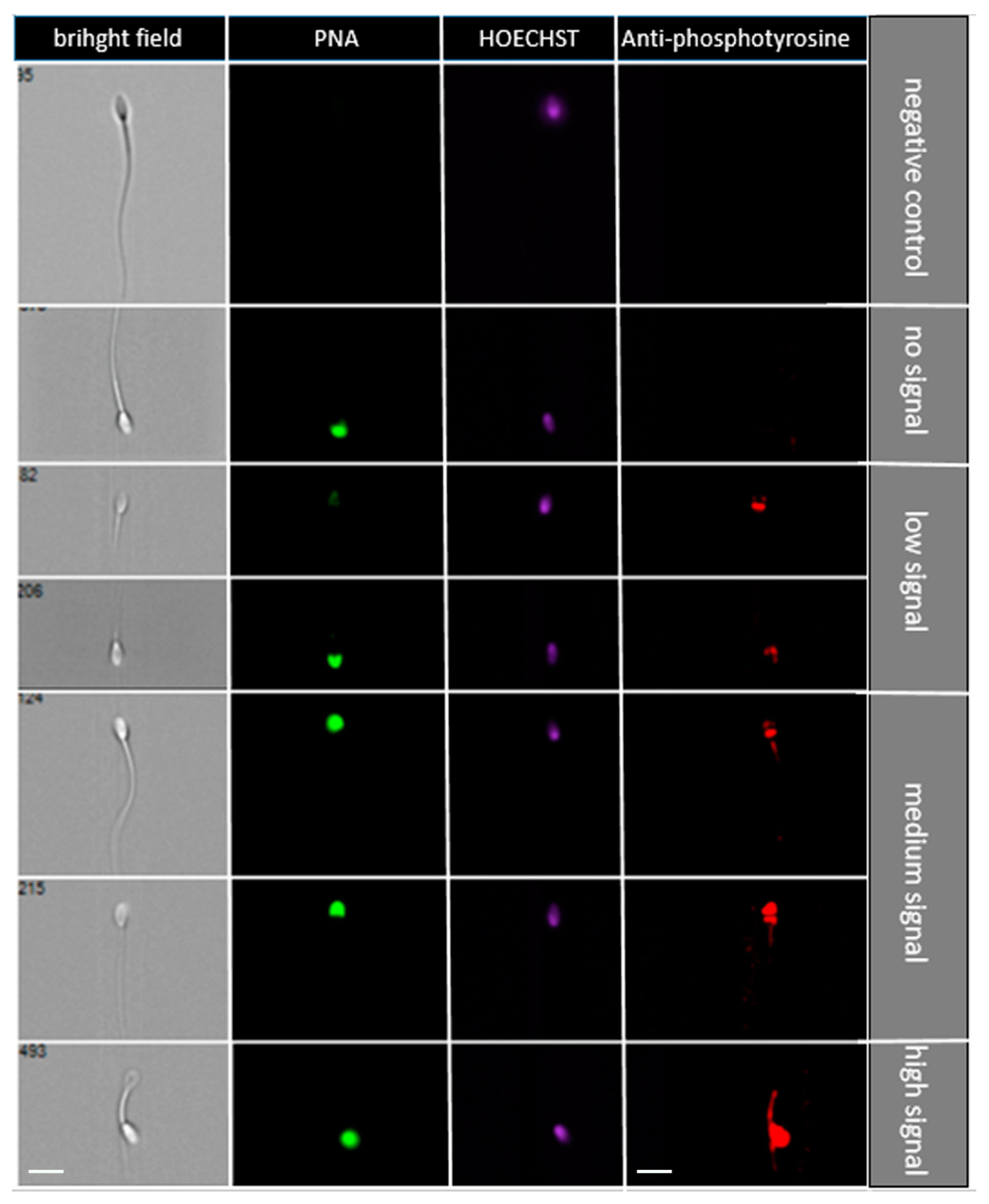
2.4. The Effect of SP Protein Fractions on Sperm Protein Phosphorylation
3. Discussion
4. Materials and Methods
4.1. Processing of Seminal Plasma
4.2. Isolation of Seminal Plasma Protein Fractions
4.3. Sperm Collection and Freezing
4.4. Sperm Quality Parameters
4.4.1. Motility Assessment
4.4.2. Viability and Acrosomal Integrity
4.4.3. Sperm Phosphorylation Rate
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vidament, M.; Dupere, A.M.; Julienne, P.; Evain, A.; Noue, P.; Palmer, E. Equine frozen semen: Freezability and fertility field results. Theriogenology 1997, 48, 907–917. [Google Scholar] [CrossRef]
- Samper, J.C.; Morris, C.A. Current methods for stallion semen cryopreservation: A survey. Theriogenology 1998, 49, 895–903. [Google Scholar] [CrossRef]
- Loomis, P.R. The equine frozen semen industry. Anim. Reprod. Sci. 2001, 68, 191–200. [Google Scholar] [CrossRef]
- Love, C.C.; Brinsko, S.P.; Rigby, S.L.; Thompson, J.A.; Blanchard, T.L.; Varner, D.D. Relationship of seminal plasma level and extender type to sperm motility and DNA integrity. Theriogenology 2005, 63, 1584–1591. [Google Scholar] [CrossRef] [PubMed]
- Topfer-Petersen, E.; Ekhlasi-Hundrieser, M.; Kirchhoff, C.; Leeb, T.; Sieme, H. The role of stallion seminal proteins in fertilisation. Anim. Reprod. Sci. 2005, 89, 159–170. [Google Scholar] [CrossRef]
- Centurion, F.; Vazquez, J.M.; Calvete, J.J.; Roca, J.; Sanz, L.; Parrilla, I.; Garcia, E.M.; Martinez, E.A. Influence of porcine spermadhesins on the susceptibility of boar spermatozoa to high dilution. Biol. Reprod. 2003, 69, 640–646. [Google Scholar] [CrossRef]
- Lessard, C.; Parent, S.; Leclerc, P.; Bailey, J.L.; Sullivan, R. Cryopreservation alters the levels of the bull sperm surface protein P25b. J. Androl. 2000, 21, 700–707. [Google Scholar] [PubMed]
- Morris, G.J.; Acton, E.; Murray, B.J.; Fonseca, F. Freezing injury: The special case of the sperm cell. Cryobiology 2012, 64, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Deleeuw, F.E.; Chen, H.C.; Colenbrander, B.; Verkleij, A.J. Cold-Induced ultrastructural-changes in bull and boar sperm plasma-membranes. Cryobiology 1990, 27, 171–183. [Google Scholar] [CrossRef]
- Bailey, J.L.; Bilodeau, J.F.; Cormier, N. Semen cryopreservation in domestic animals: A damaging and capacitating phenomenon. J. Androl. 2000, 21, 1–7. [Google Scholar] [PubMed]
- Srivastava, N.; Srivastava, S.K.; Ghosh, S.K.; Jerome, A.; Das, G.K.; Mehrotra, S. Sequestration of PDC-109 Protein by Specific Antibodies and Egg Yolk Cryoprotects Bull Spermatozoa. Reprod. Domest. Anim. 2013, 48, 724–731. [Google Scholar] [CrossRef] [PubMed]
- D’Amours, O.; Frenette, G.; Fortier, M.; Leclerc, P.; Sullivan, R. Proteomic comparison of detergent-extracted sperm proteins from bulls with different fertility indexes. Reproduction 2010, 139, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, L.P.; Harshan, H.M.; Majumdar, A.C. Seminal plasma non-heparin binding proteins (NHBP) reduce the cryoinjury to buffalo cauda epididymal spermatozoa induced by heparin binding proteins (HBP). Anim. Reprod. Sci. 2008, 104, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Singh, L.P.; Harshan, H.M.; Ansari, M.R. Effect of egg yolk and seminal plasma heparin binding protein interaction on the freezability of buffalo cauda epididymal spermatozoa. Anim. Reprod. Sci. 2007, 99, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Harshan, H.M.; Singh, L.P.; Arangasamy, A.; Ansari, M.R.; Kumar, S. Effect of buffalo seminal plasma heparin binding protein (HBP) on freezability and in vitro fertility of buffalo cauda spermatozoa. Anim. Reprod. Sci. 2006, 93, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Vasquez, R.; Cesari, A.; Greco, M.B.; Cano, A.; Hozbor, F. Extenders modify the seminal plasma ability to minimize freeze-thaw damage on ram sperm. Reprod. Domest. Anim. 2019, 54, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Aurich, J.E.; Kuhne, A.; Hoppe, H.; Aurich, C. Seminal plasma affects membrane integrity and motility of equine spermatozoa after cryopreservation. Theriogenology 1996, 46, 791–797. [Google Scholar] [CrossRef]
- Al-Essawe, E.M.; Johannisson, A.; Wulf, M.; Aurich, C.; Morrell, J.M. Improved cryosurvival of stallion spermatozoa after colloid centrifugation is independent of the addition of seminal plasma. Cryobiology 2018, 81, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.I.; Squires, E.L.; Graham, J.K. Effect of seminal plasma on the cryopreservation of equine spermatozoa. Theriogenology 2005, 63, 2372–2381. [Google Scholar] [CrossRef] [PubMed]
- Arruda, R.P.; Andrade, A.F.C.; Raphael, C.F.; Peres, K.R.; Nascimento, J.; Martins, S.; Bianconi, L.L. Effects of addition of seminal plasma on lifespan of frozen-thawed equine spermatozoa. Anim. Reprod. Sci. 2008, 107, 307–308. [Google Scholar] [CrossRef]
- Neuhauser, S.; Dorfel, S.; Handler, J. Motility of frozen-thawed stallion epididymal sperm after the addition of homologous seminal plasma from stallions with different semen quality. Reprod. Domest. Anim. 2014, 49, 83–84. [Google Scholar]
- Sichtar, J.; Bubenickova, F.; Sirohi, J.; Simonik, O. How to Increase Post-Thaw Semen Quality in Poor Freezing Stallions: Preliminary Results of the Promising Role of Seminal Plasma Added after Thawing. Animals 2019, 9, 414. [Google Scholar] [CrossRef] [PubMed]
- De Andrade, A.F.C.; Zaffalon, F.G.; Celeghini, E.C.C.; Nascimento, J.; Tarrago, O.F.B.; Martins, S.; Alonso, M.A.; Arruda, R.P. Addition of Seminal Plasma to Post-thawing Equine Semen: What is the Effect on Sperm Cell Viability. Reprod. Domest. Anim. 2011, 46, 682–686. [Google Scholar] [CrossRef]
- Caballero, I.; Vazquez, J.M.; Garcia, E.M.; Parrilla, I.; Roca, J.; Calvete, J.J.; Sanz, L.; Martinez, E.A. Major proteins of boar seminal plasma as a tool for biotechnological preservation of spermatozoa. Theriogenology 2008, 70, 1352–1355. [Google Scholar] [CrossRef] [PubMed]
- Katila, T.; Kareskoski, M. Components of stallion seminal plasma and their influence on spermatozoa. Pferdeheilkunde 2006, 22, 193. [Google Scholar] [CrossRef]
- Al-Essawe, E.; Tsikis, G.; Labas, V.; Druart, X.; Wulf, M.; Aurich, C.; Morrell, J.M. Seminal plasma components differ between “good freezer” and “poor freezer” stallions. Anim. Reprod. Sci. 2016, 169, 113. [Google Scholar] [CrossRef]
- Calvete, J.J.; Nessau, S.; Mann, K.; Sanz, L.; Sieme, H.; Klug, E.; Topferpetersen, E. isolation and biochemical-characterization of stallion seminal-plasma proteins. Reprod. Domest. Anim. 1994, 29, 411–426. [Google Scholar] [CrossRef]
- Reinert, M.; Calvete, J.J.; Sanz, L.; Mann, K.; TopferPetersen, E. Primary structure of stallion seminal plasma protein HSP-7, a zona-pellucida-binding protein of the spermadhesin family. Eur. J. Biochem. 1996, 242, 636–640. [Google Scholar] [CrossRef]
- Magdaleno, L.; Gasset, M.; Varea, J.; Schambony, A.M.; Urbanke, C.; Raida, M.; Topfer-Petersen, E.; Calvete, J.J. Biochemical and conformational characterisation of HSP-3, a stallion seminal plasma protein of the cysteine-rich secretory protein (CRISP) family. FEBS Lett. 1997, 420, 179–185. [Google Scholar] [CrossRef]
- Ekhlasi-Hundrieser, M.; Schafer, B.; Kirchhoff, C.; Hess, O.; Bellair, S.; Muller, P.; Topfer-Petersen, E. Structural and molecular characterization of equine sperm-binding fibronectin-II module proteins. Mol. Reprod. Dev. 2005, 70, 45–57. [Google Scholar] [CrossRef]
- Schambony, A.; Gentzel, M.; Wolfes, H.; Raida, M.; Neumann, U.; Topfer-Petersen, E. Equine CRISP-3: Primary structure and expression in the male genital tract. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1998, 1387, 206–216. [Google Scholar] [CrossRef]
- Usuga, A.; Rojano, B.A.; Restrepo, G. Association of the cysteine-rich secretory protein-3 (CRISP-3) and some of its polymorphisms with the quality of cryopreserved stallion semen. Reprod. Fertil. Dev. 2018, 30, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Gandotra, V.K.; Cheema, R.S.; Bansal, A.K.; Kumar, A. Seminal Plasma Heparin Binding Proteins Improve Semen Quality by Reducing Oxidative Stress during Cryopreservation of Cattle Bull Semen. Asian Australas. J. Anim. Sci. 2016, 29, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Roncoletta, M.; Morani, E.D.C.; Esper, C.R.; Barnabe, V.H.; Franceschini, P.H. Fertility-associated proteins in Nelore bull sperm membranes. Anim. Reprod. Sci. 2006, 91, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Sarsaifi, K.; Haron, A.; Vejayan, J.; Yusoff, R.; Hani, H.; Omar, M.A.; Hong, L.W.; Yimer, N.; Ju, T.Y.; Othman, A.M. Two-dimensional polyacrylamide gel electrophoresis of Bali bull (Bos javanicus) seminal plasma proteins and their relationship with semen quality. Theriogenology 2015, 84, 956–968. [Google Scholar] [CrossRef] [PubMed]
- Novak, S.; Smith, T.A.; Paradis, F.; Burwash, L.; Dyck, M.K.; Foxcroft, G.R.; Dixon, W.T. Biomarkers of in vivo fertility in sperm and seminal plasma of fertile stallions. Theriogenology 2010, 74, 956–967. [Google Scholar] [CrossRef] [PubMed]
- Jobim, M.I.M.; Trein, C.; Zirkler, H.; Gregory, R.M.; Sieme, H.; Mattos, R.C. Two-dimensional polyacrylamide gel electrophoresis of equine seminal plasma proteins and their relation with semen freezability. Theriogenology 2011, 76, 765–771. [Google Scholar] [CrossRef]
- Restrepo, G.; Rojano, B.; Usuga, A. Relationship of cysteine-rich secretory protein-3 gene and protein with semen quality in stallions. Reprod. Domest. Anim. 2019, 54, 39–45. [Google Scholar] [CrossRef]
- Doty, A.; Buhi, W.C.; Benson, S.; Scoggin, K.E.; Pozor, M.; Macpherson, M.; Mutz, M.; Troedsson, M.H.T. Equine CRISP3 Modulates Interaction Between Spermatozoa and Polymorphonuclear Neutrophils. Biol. Reprod. 2011, 85, 157–164. [Google Scholar] [CrossRef]
- Singh, A.; Brar, P.; Cheema, R. Effect of purified seminal plasma heparin binding proteins on in vitro acrosome reaction of frozen-thawed buffalo bull semen. Indian Vet. J. 2016, 93, 69–70. [Google Scholar]
- Srivastava, N.; Jerome, A.; Srivastava, S.K.; Ghosh, S.K.; Kumar, A. Bovine seminal PDC-109 protein: An overview of biochemical and functional properties. Anim. Reprod. Sci. 2013, 138, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bedford, S.J.; Jasko, D.J.; Graham, J.K.; Amann, R.P.; Squires, E.L.; Pickett, B.W. Effect of seminal extenders containing egg-yolk and glycerol on motion characteristics and fertility of stallion spermatozoa. Theriogenology 1995, 43, 955–967. [Google Scholar] [CrossRef]
- Celeghini, E.C.C.; de Arruda, R.P.; de Andrade, A.F.C.; Nascimento, J.; Raphael, C.F. Practical techniques for bovine sperm simultaneous fluorimetric assessment of plasma, acrosomal and mitochondrial membranes. Reprod. Domest. Anim. 2007, 42, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Heise, A.; Thompson, P.N.; Gerber, D. Influence of seminal plasma on fresh and post-thaw parameters of stallion epididymal spermatozoa. Anim. Reprod. Sci. 2011, 123, 192–201. [Google Scholar] [CrossRef] [PubMed]
- De Lazari, F.L.; Sontag, E.R.; Schneider, A.; Moura, A.A.A.; Vasconcelos, F.R.; Nagano, C.S.; Mattos, R.C.; Jobim, M.I.M.; Bustamante, I.C. Seminal plasma proteins and their relationship with sperm motility and morphology in boars. Andrologia 2019, 51. [Google Scholar] [CrossRef] [PubMed]
- Suárez, U.A.; Rojano, B.; Restrepo, G. Lyophilized seminal plasma can improve stallion semen freezability. Indian J. Anim. Sci. 2020, 90, 171–175. [Google Scholar]
- Quintero-Moreno, A.; Miro, J.; Rigau, A.T.; Rodriguez-Gil, J.E. Identification of sperm subpopulations with specific motility characteristics in stallion ejaculates. Theriogenology 2003, 59, 1973–1990. [Google Scholar] [CrossRef]
- Ferrusola, C.O.; Fernandez, L.G.; Morrell, J.M.; Sandoval, C.S.; Garcia, B.M.; Rodriguez-Martinez, H.; Tapia, J.A.; Pena, F.J. Lipid Peroxidation, Assessed with BODIPY-C11, Increases after Cryopreservation of Stallion Spermatozoa, is Stallion-dependent and is Related to Apoptotic-like Changes. Reprod. Domest. Anim. 2009, 44, 88. [Google Scholar] [CrossRef]
- Martinez-Pastor, F.; Jorge Tizado, E.; Julian Garde, J.; Anel, L.; de Paz, P. Statistical Series: Opportunities and challenges of sperm motility subpopulation analysis. Theriogenology 2011, 75, 783–795. [Google Scholar] [CrossRef]
- Taberner, E.; Morato, R.; Mogas, T.; Miro, J. Ability of Catalonian donkey sperm to penetrate zona pellucida-free bovine oocytes matured in vitro. Anim. Reprod. Sci. 2010, 118, 354–361. [Google Scholar] [CrossRef]
- Ferraz, M.; Morato, R.; Yeste, M.; Arcarons, N.; Pena, A.I.; Tamargo, C.; Hidalgo, C.O.; Muino, R.; Mogas, T. Evaluation of sperm subpopulation structure in relation to in vitro sperm-oocyte interaction of frozen-thawed semen from Holstein bulls. Theriogenology 2014, 81, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Gibb, Z.; Lambourne, S.R.; Aitken, R.J. The Paradoxical Relationship Between Stallion Fertility and Oxidative Stress. Biol. Reprod. 2014, 91, 2415–2426. [Google Scholar] [CrossRef] [PubMed]
- Caballero, I.; Parrilla, I.; Alminana, C.; del Olmo, D.; Roca, J.; Martinez, E.A.; Vazquez, J.M. Seminal Plasma Proteins as Modulators of the Sperm Function and Their Application in Sperm Biotechnologies. Reprod. Domest. Anim. 2012, 47, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Zigo, M.; Maňsková-Postlerov, P.; Zuidema, D.; Kerns, K.; Joňkov, V.; Tumov, L.; Bubeníckov, F.; Sutovsky, P. Porcine model for the study of sperm capacitation, fertilization and male fertility. Cell Tissue Res. 2020, 380, 237–262. [Google Scholar] [CrossRef] [PubMed]
- Visconti, P.E.; Galantino-Homer, H.; Moore, G.D.; Bailey, J.L.; Ning, X.P.; Fornes, M.; Kopf, G.S. The molecular basis of sperm capacitation. J. Androl. 1998, 19, 242–248. [Google Scholar] [PubMed]
- Therien, I.; Moreau, R.; Manjunath, P. Bovine seminal plasma phospholipid-binding proteins stimulate phospholipid efflux from epididymal sperm. Biol. Reprod. 1999, 61, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Moreau, R.; Manjunath, P. Characteristics of the cholesterol efflux induced by novel seminal phospholipid-binding proteins. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2000, 1487, 24–32. [Google Scholar] [CrossRef]
- Manjunath, P.; Villemure, M.; Lazure, C. Isolation and characterization of phospholipid-binding proteins from goat seminal plasma. Biol. Reprod. 2002, 66, 332. [Google Scholar]
- Sellem, E.; Broekhuijse, M.L.W.J.; Chevrier, L.; Camugli, S.; Schmitt, E.; Schibler, L.; Koenen, E.P.C. Use of combinations of in vitro quality assessments to predict fertility of bovine semen. Theriogenology 2015, 84, 1447–1454. [Google Scholar] [CrossRef]
- Ordonez-Leon, E.A.; Kjelland, M.E.; Moreno, J.F.; Welsh, T.H.; Randel, R.D.; Lammoglia, M.A.; Ducolomb, Y.; Romo, S. In vitro fertilization using frozen-thawed sexed semen treated with recombinant heparin-binding proteins. Reprod. Fertil. Dev. 2012, 24, 197–198. [Google Scholar] [CrossRef]
- Pena, F.J.; O’Flaherty, C.; Rodriguez, J.M.O.; Cano, F.E.M.; Gaitskell-Phillips, G.L.; Gil, M.C.; Ferrusola, C.O. Redox Regulation and Oxidative Stress: The Particular Case of the Stallion Spermatozoa. Antioxidants 2019, 8, 567. [Google Scholar] [CrossRef] [PubMed]
- Watson, P.F. The causes of reduced fertility with cryopreserved semen. Anim. Reprod. Sci. 2000, 60, 481–492. [Google Scholar] [CrossRef]
- Gadella, B.M.; Harrison, R.A.P. The capacitating agent bicarbonate induces protein kinase A-dependent changes in phospholipid transbilayer behavior in the sperm plasma membrane. Development 2000, 127, 2407–2420. [Google Scholar] [PubMed]
- Cormier, N.; Sirard, M.A.; Bailey, J.L. Premature capacitation of bovine spermatozoa is initiated by cryopreservation. J. Androl. 1997, 18, 461–468. [Google Scholar] [PubMed]
- Kumaresan, A.; Siqueira, A.P.; Hossain, M.S.; Bergqvist, A.S. Cryopreservation-induced alterations in protein tyrosine phosphorylation of spermatozoa from different portions of the boar ejaculate. Cryobiology 2011, 63, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Bedford, J.M.; Mock, O.B.; Nagdas, S.K.; Winfrey, V.P.; Olson, G.E. Reproductive characteristics of the African pygmy hedgehog, Atelerix albiventris. J. Reprod. Fertil. 2000, 120, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Kadirvel, G.; Periasamy, S.; Kumar, S. Effect of Cryopreservation on Apoptotic-like Events and its Relationship with Cryocapacitation of Buffalo (Bubalus bubalis) Sperm. Reprod. Domest. Anim. 2012, 47, 143–150. [Google Scholar] [CrossRef]
- Vieira, L.A.; Gadea, J.; Garcia-Vazquez, F.A.; Aviles-Lopez, K.; Matas, C. Equine spermatozoa stored in the epididymis for up to 96 h at 4 degrees C can be successfully cryopreserved and maintain their fertilization capacity. Anim. Reprod. Sci. 2013, 136, 280–288. [Google Scholar] [CrossRef]
- Leemans, B.; Stout, T.A.E.; De Schauwer, C.; Heras, S.; Nelis, H.; Hoogewijs, M.; Van Soom, A.; Gadella, B.M. Update on mammalian sperm capacitation: How much does the horse differ from other species. Reproduction 2019, 157, R181–R197. [Google Scholar] [CrossRef]
- Pommer, A.C.; Rutllant, J.; Meyers, S.A. Phosphorylation of protein tyrosine residues in fresh and cryopreserved stallion spermatozoa under capacitating conditions. Biol. Reprod. 2003, 68, 1208–1214. [Google Scholar] [CrossRef]
- Lusignan, M.F.; Bergeron, A.; Crete, M.H.; Lazure, C.; Manjunath, P. Induction of epididymal boar sperm capacitation by pB1 and BSP-A1/-A2 proteins, members of the BSP protein family. Biol. Reprod. 2007, 76, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Therien, I.; Bleau, G.; Manjunath, P. Phosphatidylcholine-binding proteins of bovine seminal plasma modulate capacitation of spermatozoa by heparin. Biol. Reprod. 1995, 52, 1372–1379. [Google Scholar] [CrossRef]
- Suzuki, K.; Asano, A.; Eriksson, B.; Niwa, K.; Nagai, T.; Rodriguez-Martinez, H. Capacitation status and in vitro fertility of boar spermatozoa: Effects of seminal plasma, cumulus-oocyte-complexes-conditioned medium and hyaluronan. Int. J. Androl. 2002, 25, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Vadnais, M.L.; Roberts, K.P. Effects of seminal plasma on cooling-induced capacitative changes in boar sperm. J. Androl. 2007, 28, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Leahy, T.; Rickard, J.P.; Bernecic, N.C.; Druart, X.; de Graaf, S.P. Ram seminal plasma and its functional proteomic assessment. Reproduction 2019, 157, R243–R256. [Google Scholar] [CrossRef] [PubMed]
- Druart, X.; Rickard, J.P.; Mactier, S.; Kohnke, P.L.; Kershaw-Young, C.M.; Bathgate, R.; Gibb, Z.; Crossett, B.; Tsikis, G.; Labas, V.; et al. Proteomic characterization and cross species comparison of mammalian seminal plasma. J. Proteom. 2013, 91, 13–22. [Google Scholar] [CrossRef]
- Davis, B.K. Timing of fertilization in mammals—Sperm cholesterol phospholipid ratio as a determinant of the capacitation interval. Proc. Natl. Acad. Sci. USA 1981, 78, 7560–7564. [Google Scholar] [CrossRef]
- Hoshi, K.; Aita, T.; Yanagida, K.; Yoshimatsu, N.; Sato, A. Variation in the cholesterol phospholipid ratio in human spermatozoa and its relationship with capacitation. Hum. Reprod. 1990, 5, 71–74. [Google Scholar] [CrossRef]
- Barańska, K.; Tischner, M. Evaluating Capacitation of Stallion Spermatozoa Obtained from the Mare’s Reproductive Tract. Biol. Reprod. 1995, 52, 707–712. [Google Scholar] [CrossRef]
- Manaskova, P.; Meszarosova, A.; Liberda, J.; Voburka, Z.; Ticha, M.; Jonakova, V. Aggregated forms of heparin-binding and non-heparin-binding proteins of boar seminal plasma and their binding properties. Folia Biol. 1999, 45, 193–201. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Fraction | Concentration | PI: Live Cells | PNA: Live Cells with Intact Acrosomes |
|---|---|---|---|
| Hep− | 125 µg/mL | 25.36 ± 3.1 | 48.77 ± 6.4 2 |
| 250 µg/mL | 22.03 ± 3.1 | 63.86 ± 6.4 | |
| 500 µg/mL | 23.10 ± 3.5 | 55.78 ± 7.1 2 | |
| C | 0 | 26.58 ± 3.3 | 62.73 ± 6.7 |
| Hep+ | 125 µg/mL | 25.05 ± 4.0 | 81.87 ± 8.2 1,b |
| 250 µg/mL | 27.94 ± 4.4 | 75.88 ± 9.0 b | |
| 500 µg/mL | 16.47 ± 5.7 | 44.77 ± 11.7 2,a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bubenickova, F.; Postlerova, P.; Simonik, O.; Sirohi, J.; Sichtar, J. Effect of Seminal Plasma Protein Fractions on Stallion Sperm Cryopreservation. Int. J. Mol. Sci. 2020, 21, 6415. https://doi.org/10.3390/ijms21176415
Bubenickova F, Postlerova P, Simonik O, Sirohi J, Sichtar J. Effect of Seminal Plasma Protein Fractions on Stallion Sperm Cryopreservation. International Journal of Molecular Sciences. 2020; 21(17):6415. https://doi.org/10.3390/ijms21176415
Chicago/Turabian StyleBubenickova, Filipa, Pavla Postlerova, Ondrej Simonik, Jitka Sirohi, and Jiri Sichtar. 2020. "Effect of Seminal Plasma Protein Fractions on Stallion Sperm Cryopreservation" International Journal of Molecular Sciences 21, no. 17: 6415. https://doi.org/10.3390/ijms21176415
APA StyleBubenickova, F., Postlerova, P., Simonik, O., Sirohi, J., & Sichtar, J. (2020). Effect of Seminal Plasma Protein Fractions on Stallion Sperm Cryopreservation. International Journal of Molecular Sciences, 21(17), 6415. https://doi.org/10.3390/ijms21176415