Effects of Early Life Stress on Epigenetic Changes of the Glucocorticoid Receptor 17 Promoter during Adulthood

 ,
,
Abstract
1. Introduction
2. Results
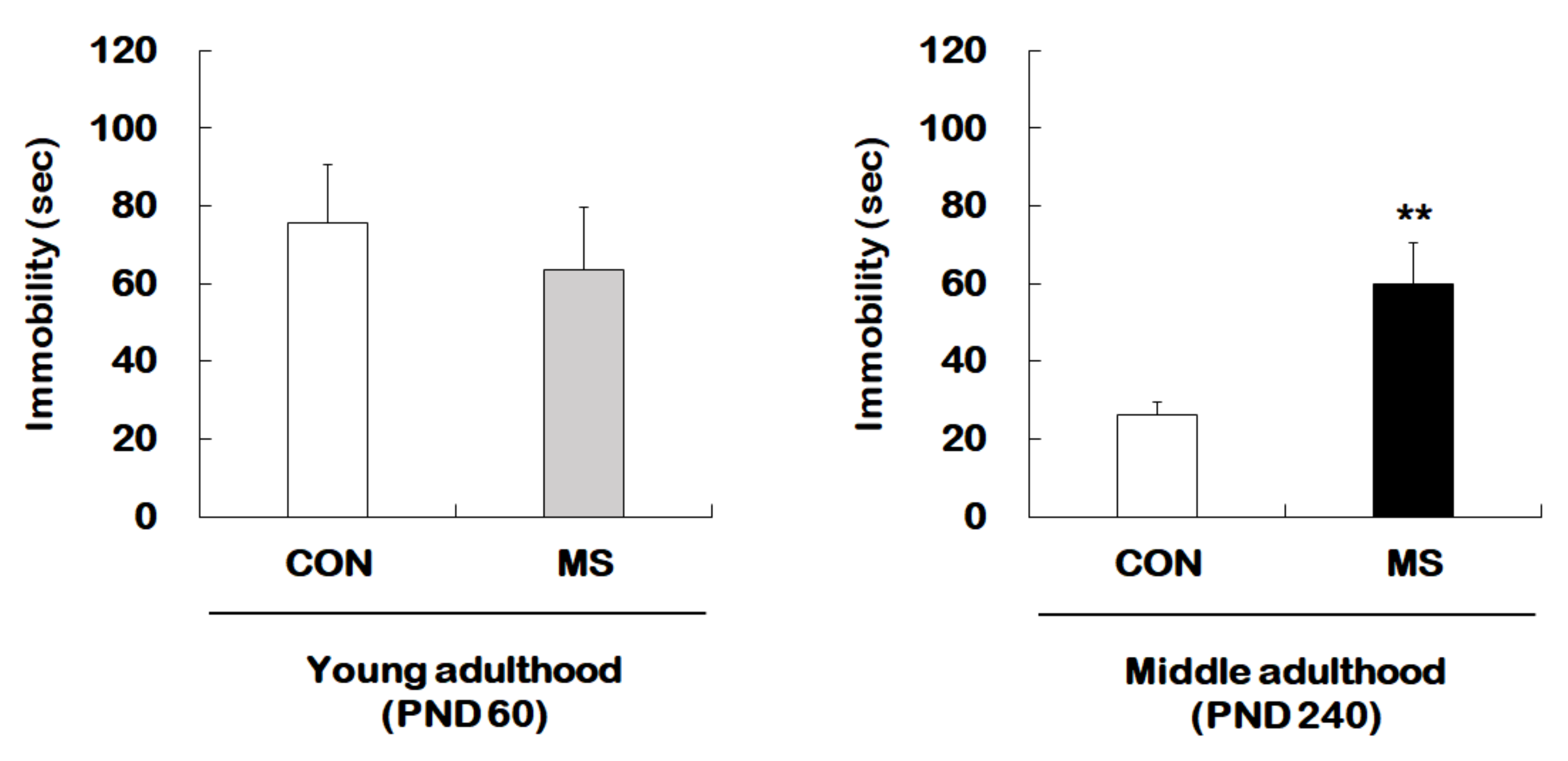
2.1. Behavioral Changes in the Forced Swimming Test
2.2. Changes in Total GR mRNA Expression
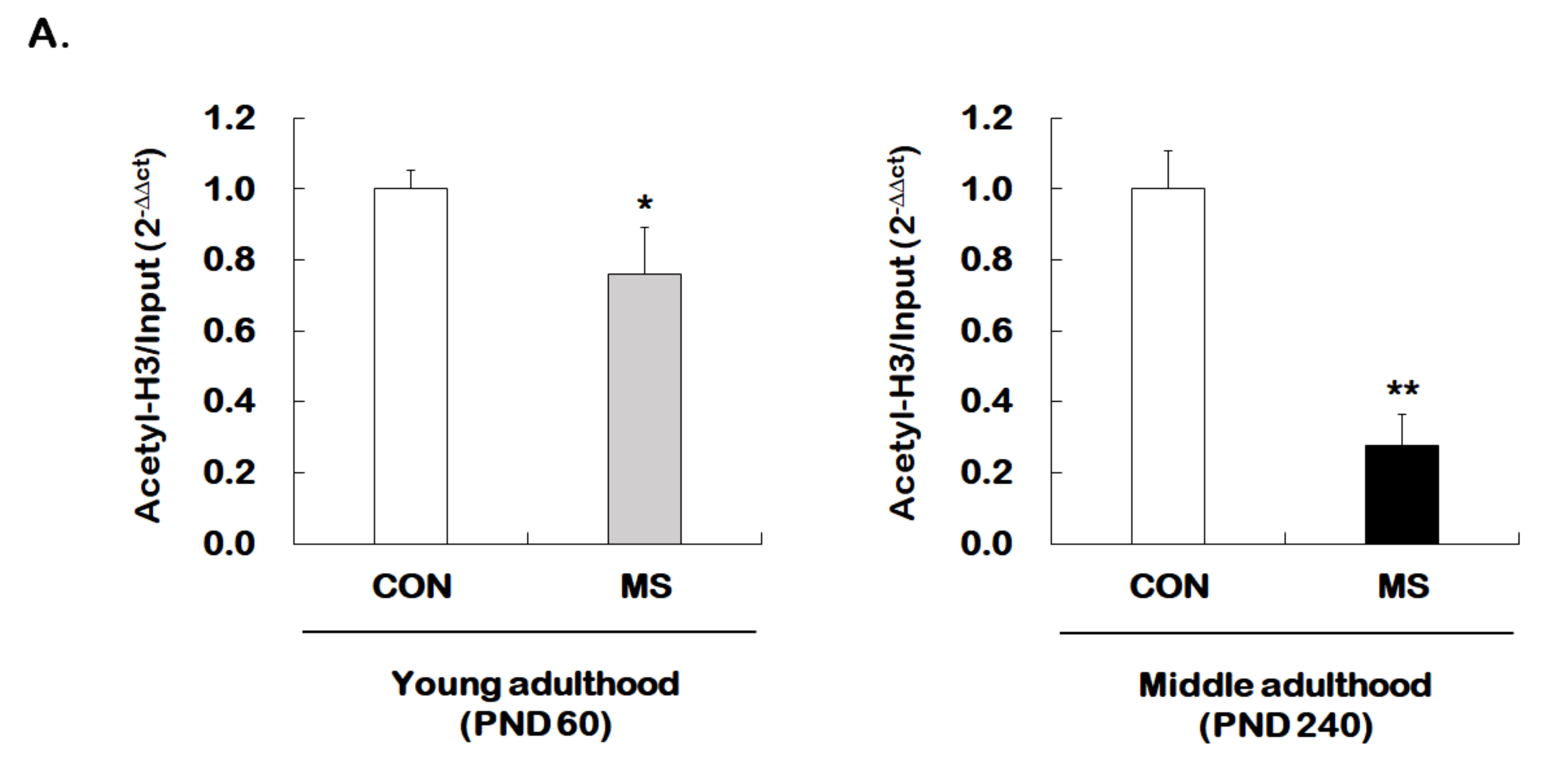
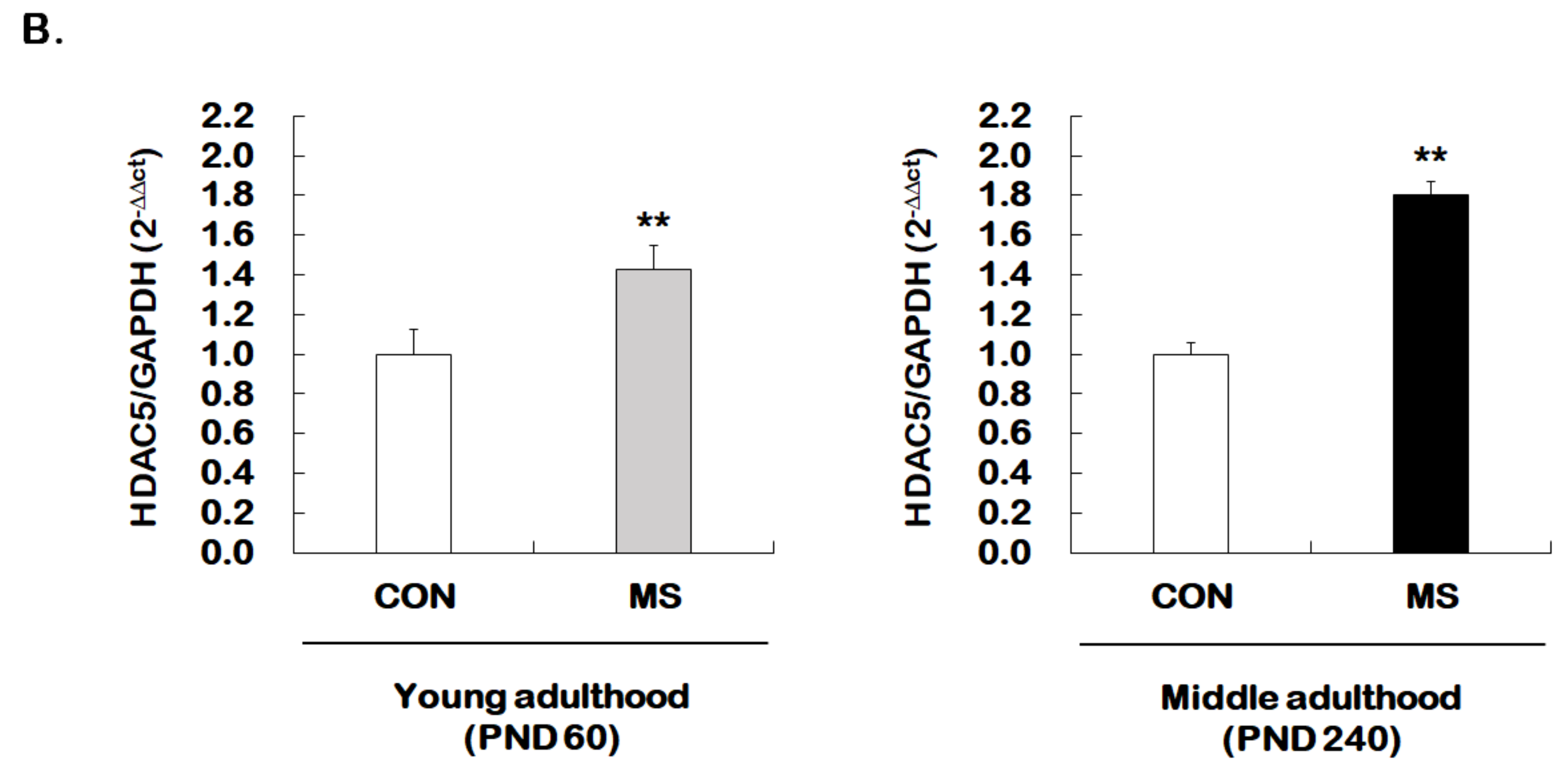
2.3. Changes of Histone H3 Acetylation of the GR Exon 17 Promoter
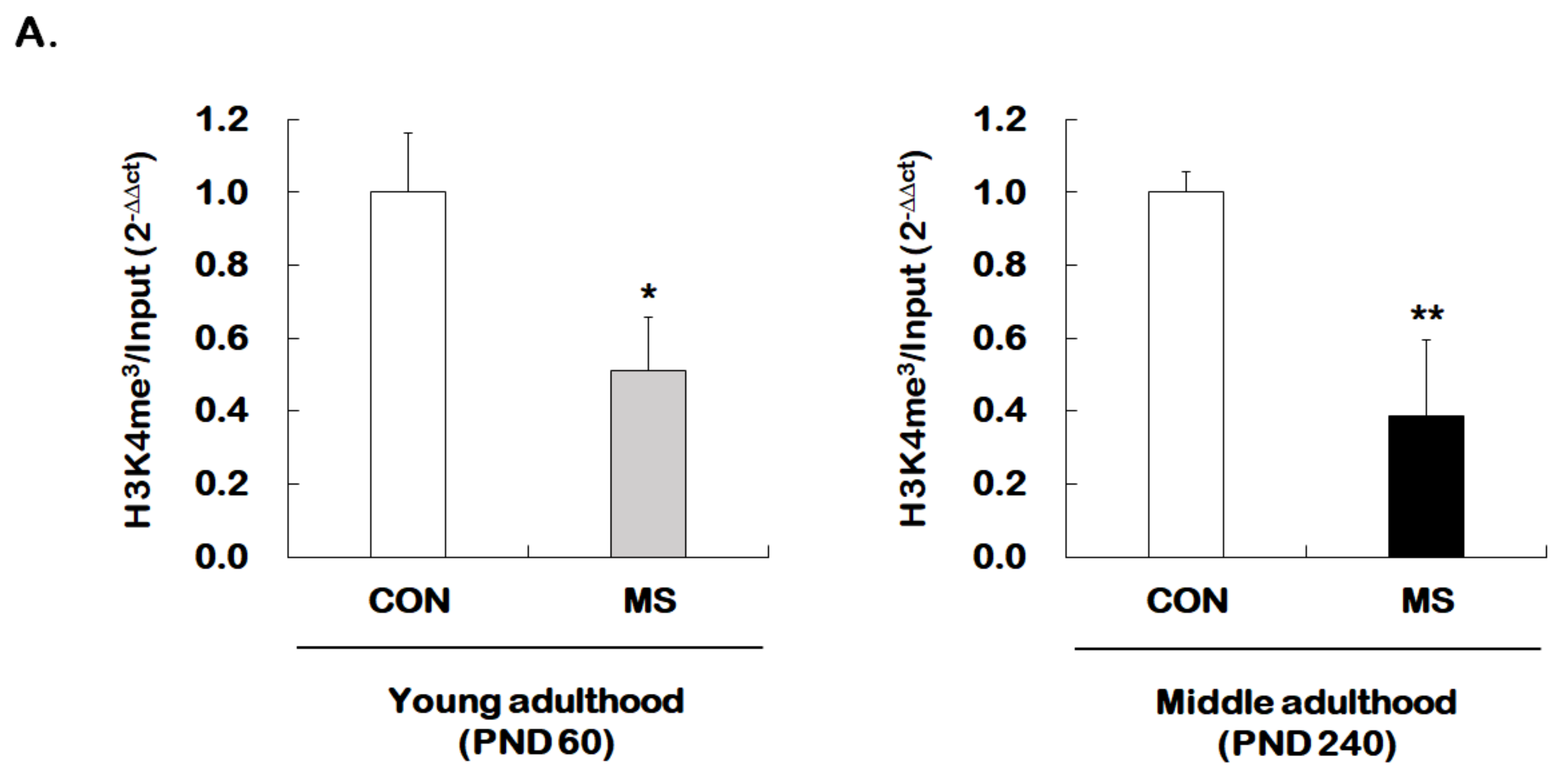
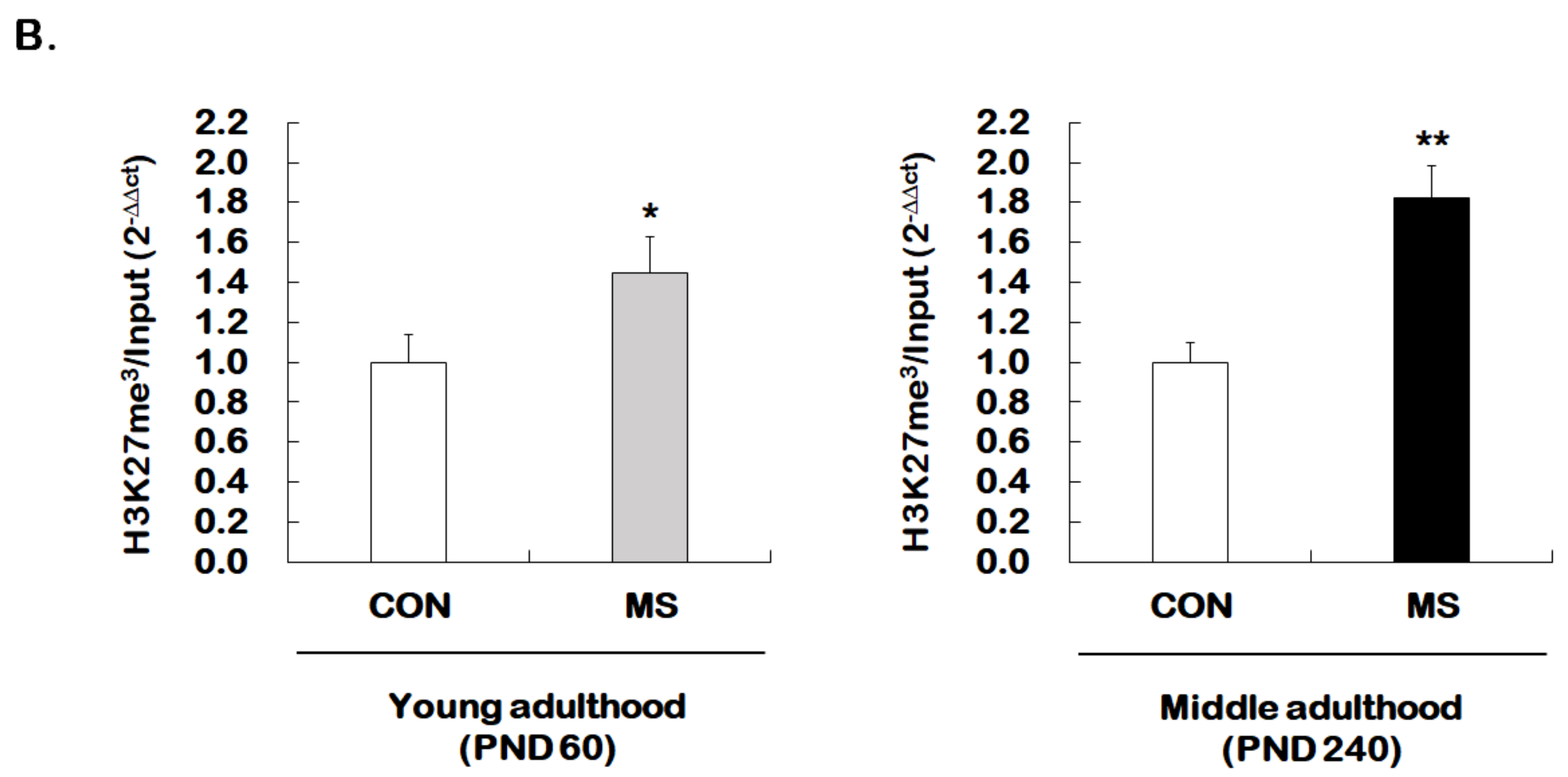
2.4. Changes of Histone H3 Methylation of the GR Exon 17 Promoter
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Maternal Separation (MS)
4.3. Forced Swimming Test (FST)
4.4. Measurement of mRNA Levels Using Quantitative Real-Time (qRT-PCR)
4.5. Chromatin Immunoprecipitation Assay (ChIP Assay)
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABN | Arched-back nursing |
| BDNF | Brain-derived neurotrophic factor |
| ChIP | Chromatin immunoprecipitation |
| CON | Control |
| DNA | Deoxyribo nucleic acid |
| ELS | Early life stress |
| FST | Forced swimming test |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GR | Glucocorticoid receptor |
| HATs | Histone acetyltransferases |
| HDACs | Histone deacetylases |
| H3 | Histone 3 |
| H3K4me3 | Tri-methylation of lysine 4 on histone H3 |
| H3K9 | lysine 9 on histone H3 |
| H3K27me3 | Tri-methylation of lysine 27 on histone H3 |
| H3K36me3 | lysine 36 on histone H3 |
| LG | Lick-groom |
| mRNA | messenger RNA |
| MS | Maternal separation |
| NGFIA | Nerve growth factor-inducible protein A |
| NR3C1 | GR gene |
| PND | Postnatal day |
| qRT-PCR | quantitative real-time PCR |
| RNA | Ribonucleic acid |
References
- Burgess, R.L.; Conger, R.D. Family interaction in abusive, neglectful, and normal families. Child Dev. 1978, 49, 1163–1173. [Google Scholar] [CrossRef]
- Macmillan, H.L.; Fleming, J.E.; Streiner, D.L.; Duku, E.K.; Wong, M.Y.-Y.; Lin, E.; Boyle, M.H.; Jamieson, E.; Walsh, C.A.; Beardslee, W.R. Childhood Abuse and Lifetime Psychopathology in a Community Sample. Am. J. Psychiatry 2001, 158, 1878–1883. [Google Scholar] [CrossRef] [PubMed]
- Bernet, C.Z.; Stein, M.B. Relationship of childhood maltreatment to the onset and course of major depression in adulthood. Depress. Anxiety 1999, 9, 169–174. [Google Scholar] [CrossRef]
- Klein, D.N.; Arnow, B.A.; Barkin, J.L.; Dowling, F.; Kocsis, J.H.; Leon, A.C.; Manber, R.; Rothbaum, B.O.; Trivedi, M.H.; Wisniewski, S.R.; et al. Early adversity in chronic depression: Clinical correlates and response to pharmacotherapy. Depress. Anxiety 2009, 26, 701–710. [Google Scholar] [CrossRef] [PubMed]
- McGowan, P.O.; Suderman, M.; Sasaki, A.; Huang, T.C.; Hallett, M.; Meaney, M.J.; Szyf, M. Broad Epigenetic Signature of Maternal Care in the Brain of Adult Rats. PLoS ONE 2011, 6, e14739. [Google Scholar] [CrossRef] [PubMed]
- Murgatroyd, C.; Patchev, A.V.; Wu, Y.; Micale, V.; Bockmühl, Y.; Fischer, D.; Holsboer, F.; Wotjak, C.T.; Almeida, O.F.; Spengler, D. Dynamic DNA methylation programs persistent adverse effects of early-life stress. Nat. Neurosci. 2009, 12, 1559–1566. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Lee, J.G.; Seo, M.K.; Ly, N.N.; Lee, C.H.; Cho, H.Y.; Hein, L.T.; Choi, A.J.; Kim, G.-M.; Kim, Y.H. Epigenetic modification of glucocorticoid receptor promoter I7 in maternally separated and restraint-stressed rats. Neurosci. Lett. 2017, 650, 38–44. [Google Scholar] [CrossRef]
- McCormick, J.A.; Lyons, V.; Jacobson, M.D.; Noble, J.; Diorio, J.; Nyirenda, M.; Weaver, S.; Ester, W.; Yau, J.L.; Meaney, M.J.; et al. 5’-Heterogeneity of Glucocorticoid Receptor Messenger RNA Is Tissue Specific: Differential Regulation of Variant Transcripts by Early-Life Events. Mol. Endocrinol. 2000, 14, 506–517. [Google Scholar] [CrossRef]
- Turecki, G.; Meaney, M.J. Effects of the Social Environment and Stress on Glucocorticoid Receptor Gene Methylation: A Systematic Review. Biol. Psychiatry 2016, 79, 87–96. [Google Scholar] [CrossRef]
- Sun, H.; Kennedy, P.J.; Nestler, E.J. Epigenetics of the Depressed Brain: Role of Histone Acetylation and Methylation. Neuropsychopharmacology 2013, 38, 124–137. [Google Scholar] [CrossRef]
- Borrelli, E.; Nestler, E.J.; Allis, C.D.; Sassone-Corsi, P. Decoding the Epigenetic Language of Neuronal Plasticity. Neuron 2008, 60, 961–974. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.K.; Howe, L.J. Histone acetylation: Truth of consequences? Biochem. Cell Biol. 2009, 87, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Weaver, I.C.G.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic programming by maternal behavior. Nat. Neurosci. 2004, 7, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Weaver, I.C.G.; D’Alessio, A.C.; Brown, S.E.; Hellstrom, I.C.; Dymov, S.; Sharma, S.; Szyf, M.; Meaney, M.J. The transcription factor nerve growth factor-inducible protein a mediates epigenetic programming: Altering epigenetic marks by immediate-early genes. J. Neurosci. 2007, 27, 1756–1768. [Google Scholar] [CrossRef]
- Francis, D.D.; Diorio, J.; Plotsky, P.M.; Meaney, M.J. Environmental Enrichment Reverses the Effects of Maternal Separation on Stress Reactivity. J. Neurosci. 2002, 22, 7840–7843. [Google Scholar] [CrossRef]
- Lippmann, M.; Bress, A.; Nemeroff, C.B.; Plotsky, P.M.; Monteggia, L.M. Long-term behavioural and molecular alterations associated with maternal separation in rats. Eur. J. Neurosci. 2007, 25, 3091–3098. [Google Scholar] [CrossRef]
- Plotsky, P.M.; Meaney, M.J. Early, postnatal experience alters hypothalamic corticotropin-releasing factor (CRF) mRNA, median eminence CRF content and stress-induced release in adult rats. Mol. Brain Res. 1993, 18, 195–200. [Google Scholar] [CrossRef]
- Ladd, C.O.; Huot, R.L.; Thrivikraman, K.V.; Nemeroff, C.B.; Plotsky, P.M. Long-term adaptations in glucocorticoid receptor and mineralocorticoid receptor mRNA and negative feedback on the hypothalamo-pituitary-adrenal axis following neonatal maternal separation. Biol. Psychiatry 2004, 55, 367–375. [Google Scholar] [CrossRef]
- Stankiewicz, A.M.; Swiergiel, A.H.; Lisowski, P. Epigenetics of stress adaptations in the brain. Brain Res. Bull. 2013, 98, 76–92. [Google Scholar] [CrossRef]
- Millstein, R.A.; Holmes, A. Effects of repeated maternal separation on anxiety- and depression-related phenotypes in different mouse strains. Neurosci. Biobehav. Rev. 2007, 31, 3–17. [Google Scholar] [CrossRef]
- Rüedi-Bettschen, D.; Zhang, W.; Russig, H.; Ferger, B.; Weston, A.; Pedersen, E.-M.; Feldon, J.; Pryce, C.R. Early deprivation leads to altered behavioural, autonomic and endocrine responses to environmental challenge in adult Fischer rats. Eur. J. Neurosci. 2006, 24, 2879–2893. [Google Scholar] [CrossRef] [PubMed]
- Lambás-Señas, L.; Mnie-Filali, O.; Certin, V.; Faure, C.; Lemoine, L.; Zimmer, L.; Haddjeri, N. Functional correlates for 5-HT1A receptors in maternally deprived rats displaying anxiety and depression-like behaviors. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.K.; Ly, N.N.; Lee, C.H.; Cho, H.Y.; Choi, C.M.; Nhu, L.H.; Lee, J.G.; Lee, B.J.; Kim, G.-M.; Yoon, B.J.; et al. Early life stress increases stress vulnerability through BDNF gene epigenetic changes in the rat hippocampus. Neuropharmacology 2016, 105, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Seo, M.K.; Lee, J.G.; Hien, L.T.; Kim, Y.H. Effects of maternal separation and antidepressant drug on epigenetic regulation of the brain-derived neurotrophic factor exon I promoter in the adult rat hippocampus. Psychiatry Clin. Neurosci. 2018, 72, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Boyle, M.P.; Brewer, J.A.; Funatsu, M.; Wozniak, D.F.; Tsien, J.Z.; Izumi, Y.; Muglia, L.J. Acquired deficit of forebrain glucocorticoid receptor produces depression-like changes in adrenal axis regulation and behavior. Proc. Natl. Acad. Sci. USA 2005, 102, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Navailles, S.; Zimnisky, R.; Schmauss, C. Expression of glucocorticoid receptor and early growth response gene 1 during postnatal development of two inbred strains of mice exposed to early life stress. Dev. Neurosci. 2010, 32, 139–148. [Google Scholar] [CrossRef]
- Daniels, W.M.; Fairbairn, L.R.; Van Tilburg, G.; McEvoy, C.R.; Zigmond, M.J.; Russell, V.A.; Stein, D.J. Maternal separation alters nerve growth factor and corticosterone levels but not the DNA methylation status of the exon 17 glucocorticoid receptor promoter region. Metab. Brain Dis. 2009, 24, 615–627. [Google Scholar] [CrossRef]
- Klengel, T.; Pape, J.; Binder, E.B.; Mehta, D. The role of DNA methylation in stress-related psychiatric disorders. Neuropharmacology 2014, 80, 115–132. [Google Scholar] [CrossRef]
- McGowan, P.O.; Sasaki, A.; D’Alessio, A.C.; Dymov, S.; LaBonte, B.; Szyf, M.; Turecki, G.; Meaney, M.J. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 2009, 12, 342–348. [Google Scholar] [CrossRef]
- Tsankova, N.M.; Berton, O.; Renthal, W.; Kumar, A.; Neve, R.L.; Nestler, E.J. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat. Neurosci. 2006, 9, 519–525. [Google Scholar] [CrossRef]
- Ferland, C.L.; Schrader, L.A. Regulation of histone acetylation in the hippocampus of chronically stressed rats: A potential role of sirtuins. Neuroscience 2011, 174, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Renthal, W.; Maze, I.; Krishnan, V.; Covington, H.E., III; Xiao, G.; Kumar, A.; Russo, S.J.; Graham, A.; Tsankova, N.; Kippin, T.E.; et al. Histone Deacetylase 5 Epigenetically Controls Behavioral Adaptations to Chronic Emotional Stimuli. Neuron 2007, 56, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Hobara, T.; Uchida, S.; Otsuki, K.; Matsubara, T.; Funato, H.; Matsuo, K.; Suetsugi, M.; Watanabe, Y. Altered gene expression of histone deacetylases in mood disorder patients. J. Psychiatr. Res. 2010, 44, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Weaver, I.C.G.; Champagne, F.A.; Brown, S.E.; Dymov, S.; Sharma, S.; Meaney, M.J.; Szyf, M. Reversal of Maternal Programming of Stress Responses in Adult Offspring through Methyl Supplementation: Altering Epigenetic Marking Later in Life. J. Neurosci. 2005, 25, 11045–11054. [Google Scholar] [CrossRef]
- Deussing, J.M. Animal models of depression. Drug Discov. Today Dis. Model. 2006, 3, 375–383. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Le Pichon, M.; Jalfre, M. Depression: A new animal model sensitive to antidepressant treatments. Nature 1977, 266, 730–732. [Google Scholar] [CrossRef]
- Onishchenko, N.; Karpova, N.; Sabri, F.; Castrn, E.; Ceccatelli, S.; Castrén, E. Long-lasting depression-like behavior and epigenetic changes of BDNF gene expression induced by perinatal exposure to methylmercury. J. Neurochem. 2008, 106, 1378–1387. [Google Scholar] [CrossRef]
- Vazquez, A.P.; Rye, N.; Ameri, M.; Mcsparron, B.; Smallwood, G.; Bickerdyke, J.; Rathbone, A.; Bailador, F.D.; Toledo-Rodriguez, M. Impact of voluntary exercise and housing conditions on hippocampal glucocorticoid receptor, miR-124 and anxiety. Mol. Brain 2015, 8, 40. [Google Scholar] [CrossRef]
- Licata, S.C.; Shinday, N.M.; Huizenga, M.N.; Darnell, S.B.; Sangrey, G.R.; Rudolph, U.; Rowlett, J.K.; Sadri-Vakili, G. Alterations in Brain-Derived Neurotrophic Factor in the Mouse Hippocampus Following Acute but Not Repeated Benzodiazepine Treatment. PLoS ONE 2013, 8, e84806. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Bank Number | Primer Sequence (5′-3′) | |
|---|---|---|---|
| quantitative real-time PCR (qRT-PCR) | |||
| Total GR [38] | NM_008173.3 | Forward | AGGCCGCTCAGTGTTTTCTA |
| Reverse | TACAGCTTCCACACGTCAGC | ||
| HDAC5 | NM_001284249.1 | Forward | CCATTGGAGATGTGGAATAC |
| Reverse | CAGTGGAGACAGATGTCCTT | ||
| GAPDH [39] | NG_007244.3 | Forward | AACAGCAACTCCCATTCTTC |
| Reverse | TGGTCCAGGGTTTCTTACTC | ||
| qRT-PCR for histone modification | |||
| GR exon 17 promoter | XM_006525663.4 | Forward | TTCCGTGCCATCCTGTA |
| Reverse | CCGAGTTTCTTTAGTTTCTCTT | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, M.K.; Kim, S.-g.; Seog, D.-H.; Bahk, W.-M.; Kim, S.-H.; Park, S.W.; Lee, J.G. Effects of Early Life Stress on Epigenetic Changes of the Glucocorticoid Receptor 17 Promoter during Adulthood. Int. J. Mol. Sci. 2020, 21, 6331. https://doi.org/10.3390/ijms21176331
Seo MK, Kim S-g, Seog D-H, Bahk W-M, Kim S-H, Park SW, Lee JG. Effects of Early Life Stress on Epigenetic Changes of the Glucocorticoid Receptor 17 Promoter during Adulthood. International Journal of Molecular Sciences. 2020; 21(17):6331. https://doi.org/10.3390/ijms21176331
Chicago/Turabian StyleSeo, Mi Kyoung, Seon-gu Kim, Dae-Hyun Seog, Won-Myong Bahk, Seong-Ho Kim, Sung Woo Park, and Jung Goo Lee. 2020. "Effects of Early Life Stress on Epigenetic Changes of the Glucocorticoid Receptor 17 Promoter during Adulthood" International Journal of Molecular Sciences 21, no. 17: 6331. https://doi.org/10.3390/ijms21176331
APA StyleSeo, M. K., Kim, S.-g., Seog, D.-H., Bahk, W.-M., Kim, S.-H., Park, S. W., & Lee, J. G. (2020). Effects of Early Life Stress on Epigenetic Changes of the Glucocorticoid Receptor 17 Promoter during Adulthood. International Journal of Molecular Sciences, 21(17), 6331. https://doi.org/10.3390/ijms21176331