Abstract
Osteosarcoma is the most common primary tumor of the skeletal system and is well-known to have an aggressive clinical outcome and high metastatic potential. The chemokine (C-X-C motif) ligand 13 (CXCL13) plays a vital role in the development of several cancers. However, the effect of CXCL13 in the motility of osteosarcoma cells remains uncertain. Here, we found that CXCL13 increases the migration and invasion potential of three osteosarcoma cell lines. In addition, CXCL13 expression was upregulated in migration-prone MG-63 cells. Vascular cell adhesion molecule 1 (VCAM-1) siRNA and antibody demonstrated that CXCL13 promotes migration via increasing VCAM-1 production. We also show that CXCR5 receptor controls CXCL13-mediated VCAM-1 expression and cell migration. Our study identified that CXCL13/CXCR5 axis facilitate VCAM-1 production and cell migration in human osteosarcoma via the phospholipase C beta (PLCβ), protein kinase C α (PKCα), c-Src, and nuclear factor-κB (NF-κB) signaling pathways. CXCL13 and CXCR5 appear to be a novel therapeutic target in metastatic osteosarcoma.
1. Introduction
Osteosarcoma is the most common primary bone tumor and is highly malignant [1]. Neoadjuvant and adjuvant chemotherapy for osteosarcoma has largely remained unchanged since the 1980s, with wide surgical excision and preservation of limb function [1]. However, despite such treatment, only 60–70% of patients without clinically evident metastasis at first diagnosis remain alive after 3 years [2]. Major challenges that reduce overall survival of patients with osteosarcoma include the development of chemoresistance and progression to lung metastasis [3,4,5]. Since the lung is the most frequent organ affected by metastasis in osteosarcoma [2], the development of therapeutic strategies for delaying or inhibiting lung metastasis progression remains critical for improving patient survival outcomes.
The metastatic process is a multi-stage cascade, whereby cancer characterized by the dissemination of tumor cells with proteolytic activity, enabling the tumor to invade various tissues [1]. Several different adhesion molecules regulate cancer migration and invasion during the metastatic process, including vascular cell adhesion molecule 1 (VCAM-1), an inducible surface glycoprotein from the immunoglobulin superfamily, which is involved in numerous biological activities [6]. VCAM-1 is highly expressed in metastatic cancer cells [7]. In osteosarcoma, levels of VCAM-1 expression correlate with disease stage and tumor progression [4], with higher levels of VCAM-1 expression enabling connective tissue growth factor to stimulate the migration of osteosarcoma cells and promote metastasis [8]. Thus, inhibiting VCAM-1 expression may be an effective strategy for treating osteosarcoma metastasis.
Chemokines are critical players in various cellular processes, contributing to the homing, migratory, angiogenic, and proliferative activities of cells [9]. They also regulate various cancer functions including progression, metastasis, apoptosis, and chemoresistance [10,11,12]. The chemokine (C-X-C motif) ligand 13 (CXCL13) plays a vital role in the development of several cancers (e.g., gastric, colorectal, breast and lung cancers), the promotion of angiogenesis, and the metastatic cascade [13,14,15,16]. Elevated levels of CXCL13 and its cognate receptor, CXCR5, are found in many types of cancer cells [16]. However, little is known about the CXCL13/CXCR5 signaling axis in osteosarcoma. Here, we report that the CXCL13/CXCR5 interaction promotes the migratory and invasive potential of human osteosarcoma cells, which in turn mediates the phospholipase C β (PLCβ), protein kinase C α (PKCα), c-Src, and nuclear factor-κB (NF-κB) pathways.
2. Results
2.1. CXCL13 Promotes Migration and Invasion of Human Osteosarcoma Cells
CXCL13 promotes the migration and metastases of cancer cells [16]. We first applied CXCL13 to human osteosarcoma cell lines and examined cell motility. Treatment of human osteosarcoma cell lines (MG-63, HOS, and U2OS) with CXCL13 (3–30 ng/mL) concentration-dependently facilitated migratory and invasive abilities (Figure 1A,B), but did not affect the viability of osteosarcoma cells (Figure 1C). Next, we investigated whether CXCL13 expression influences the migratory activity of osteosarcoma tumor cells. We previously established high-migration-prone sublines MG-63 (M2), MG-63 (M5), and MG-63 (M10) [17]. Our results in this study were similar to those in our previous experiment, which showed that migration-prone sublines MG-63 (M10) and MG-63 (M5) had higher cell motility than the original MG-63 cell line (Figure 1D). We also found that CXCL13 levels were upregulated in the high-migration-prone sublines (Figure 1E,F). Transfection of MG-63 (M10) cells with CXCL13 siRNA inhibited CXCL13 expression and cell migratory activity (Figure 1G–I). Transfecting MG-63 cells with CXCL13 siRNA did not significantly affect cell proliferation (Figure S1). Thus, these results indicate that CXCL13 promotes human osteosarcoma cell migration.
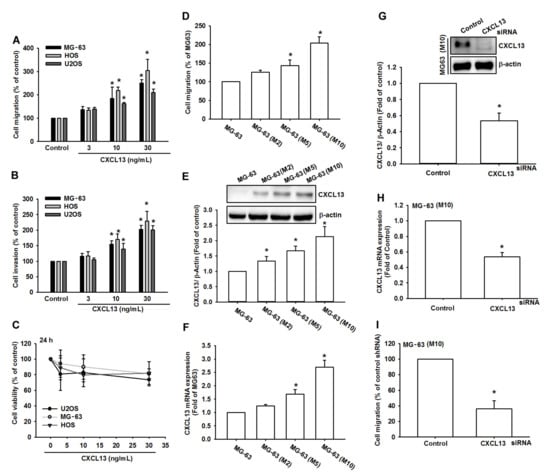
Figure 1.
The chemokine (C-X-C motif) ligand 13 (CXCL13) increases migration and invasion in human osteosarcoma. (A,B) Cells were incubated with CXCL13 (3–30 ng/mL) and the Transwell assay determined in vitro migratory and invasion activity after 24 h. (C) Cells were incubated with CXCL13 for 24 h and cell viability was examined by the MTT assay. (D–F) Migratory ability and CXCL13 expression of the indicated cells was examined by Transwell, Western blot and qPCR assays. (G–I) Cells were transfected with CXCL13 siRNA then stimulated with CXCL13; CXCL13 expression and migratory potential was examined by Western blot, qPCR, and Transwell assays. * p < 0.05 compared with the control group.
2.2. CXCL13 Promotes Osteosarcoma Cell Migration by Increasing VCAM-1 Expression
VCAM-1-dependent motility is essential for the development of tumor metastases [16,18]. We wondered if VCAM-1 also mediates the effects of CXCL13 upon cell migration. Incubation of osteosarcoma cells with CXCL13 increased VCAM-1 expression (Figure 2A). Transfection of cells with VCAM-1 siRNA reduced levels of VCAM-1 expression and CXCL13-induced promotion of osteosarcoma cell migration (Figure 2B–D), indicating that CXCL13 facilitates VCAM-1-dependent osteosarcoma cell migration.
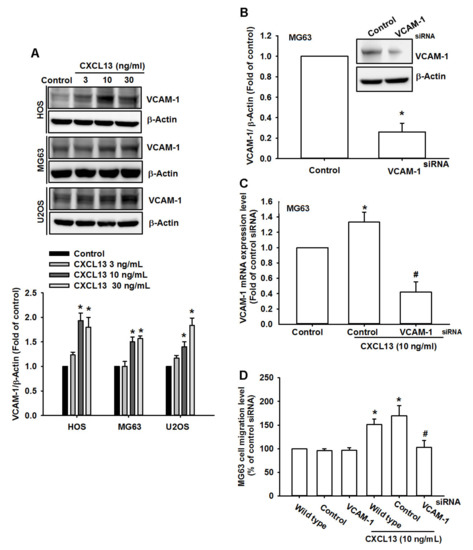
Figure 2.
CXCL13 promotes osteosarcoma migration by increasing vascular cell adhesion molecule 1 (VCAM-1) expression. (A) Cells were incubated with CXCL13 (3–30 ng/mL) and VCAM-1 expression was examined by Western blot. (B–D) MG-63 cells were transfected with VCAM-1 siRNA then stimulated with CXCL13; VCAM-1 expression and migratory potential was examined by Western blot, qPCR, and Transwell assays. * p < 0.05 compared with the control group; # p < 0.05 compared with the CXCL13-treated group.
2.3. The CXCL13/CXCR5 Axis Stimulates Osteosarcoma Cell Migration by Increasing VCAM-1 Expression
CXCR5 is a specific CXCL13 receptor that mediates CXCL13-regulated cancer functions [19]. Here, we found that transfecting cells with CXCR5 siRNA reversed the effects of CXCL13 upon cell migration and VCAM-1 expression (Figure 3A–C). Similarly, the CXCR5 receptor antibody reduced CXCL13-induced increases in cell migration and VCAM-1 production (Figure 3D–F). Therefore, the CXCL13/CXCR5 interaction regulates VCAM-1 synthesis and osteosarcoma cell migration. A comparison of levels of CXCR5 protein expression in all three osteosarcoma cell lines demonstrated that CXCR5 expression correlates with malignancy (Figure S2).
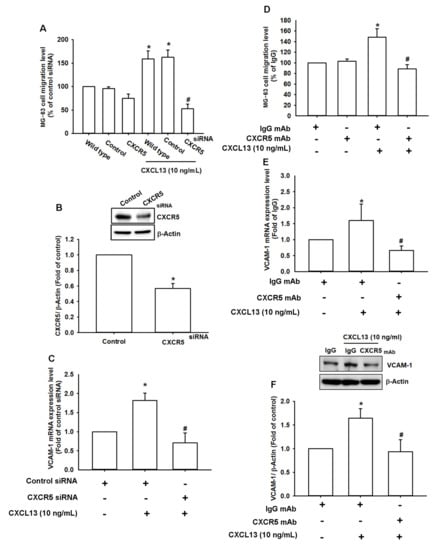
Figure 3.
The CXCL13/CXCR5 axis increases VCAM-1 expression and cell migration. MG-63 cells were transfected with CXCR5 siRNA (A–C) or pretreated with CXCR5 antibody (D,E), then stimulated with CXCL13; migratory potential and VCAM-1 expression was examined by Transwell, qPCR and Western blot assays. (F) MG-63 cells were pretreated with CXCR5 antibody then stimulated with CXCL13; VCAM-1 protein expression was examined by Western blot assay. * p < 0.05 compared with the control group; # p < 0.05 compared with the CXCL13-treated group.
2.4. The PLCβ, PKCα, c-Src, and NF-κB Pathways Are Involved in CXCL13-Induced VCAM-1 Expression and Cell Migration
The PLCβ, PKCα, and c-Src signaling pathways are commonly identified during cancer metastasis [20,21]. Incubation of all three osteosarcoma cell lines with PLCβ, PKCα, and c-Src inhibitors (U73122, GF109203X, and PP2, respectively) antagonized CXCL13-induced increases in cell migration and VCAM-1 production (Figure 4A–C and Figure S3A,B). Likewise, PLCβ, PKCα, and c-Src siRNAs reduced CXCL13-induced stimulation of osteosarcoma cell migration D and Figure S3C,D). Stimulation of the cells with CXCL13 promoted PLCβ, PKCα, and c-Src phosphorylation in a time-dependent manner (Figure 4E). These data suggest that CXCL13 promotes VCAM-1-dependent migration via the PLCβ, PKCα, and c-Src pathways.
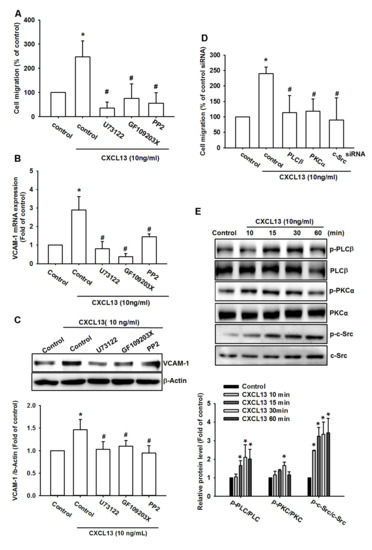
Figure 4.
The phospholipase C β (PLCβ), protein kinase C α (PKCα), and c-Src pathways are involved in CXCL13-promoted migration and VCAM-1 production. (A–C) MG-63 cells were pretreated with U73122, GF109203X, and PP2 then stimulated with CXCL13; migratory potential and VCAM-1 expression was examined by Transwell, qPCR and Western blot assays. (D) MG-63 cells were transfected with PLCβ, PKCα, and c-Src siRNA then stimulated with CXCL13; migratory potential was examined by the Transwell assay. (E) MG-63 cells were incubated with CXCL13 for the indicated time intervals and PLCβ, PKCα and c-Src phosphorylation was examined by Western blot. * p < 0.05 compared with the control group; # p < 0.05 compared with the CXCL13-treated group.
NF-κB is an important transcriptional factor that mediates cancer cell migration and metastasis [22,23]. Treating cells with NF-κB inhibitors (BAY 11-7082, PDTC, and TPCK) inhibited CXCL13-induced increases in cell migration and VCAM-1 expression (Figure 5A–C and Figure S3A,B). Transfection of cells with p65 siRNA also reduced CXCL13-induced increases in cell migration and VCAM-1 production (Figure 5D–F and Figure S3C,D). Treatment of osteosarcoma cells with CXCL13 facilitated IKK, IκBα, and p65 phosphorylation in a time-dependent manner (Figure 5G).
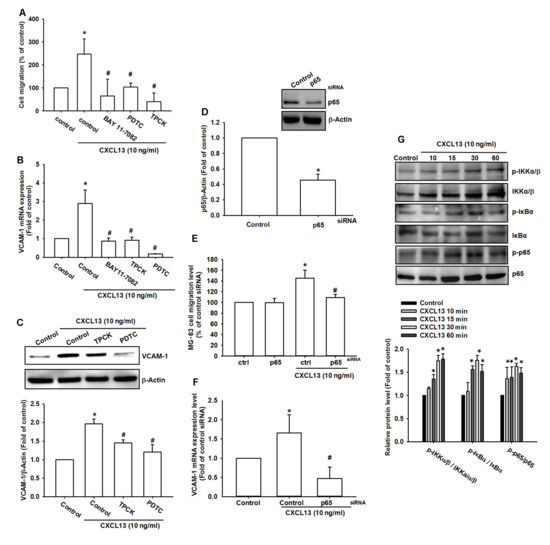
Figure 5.
The transcription factor nuclear factor-κB (NF-κB) is involved in CXCL13-induced cell migration and VCAM-1 production. MG-63 cells were pretreated with BAY 11-7082, PDTC, or TPCK (A–C), or transfected with p65 siRNA (D–F), then stimulated with CXCL13; cell migratory potential and VCAM-1 expression was examined by Transwell, qPCR, and Western blot assays. (G) MG-63 cells were incubated with CXCL13 for the indicated time intervals, and IKK, IκBα, and p65 phosphorylation was examined by Western blot. * p < 0.05 compared with the control group; # p < 0.05 compared with the CXCL13-treated group.
We then used the NF-κB luciferase promoter plasmid to determine the possibility that the PLCβ, PKCα, and c-Src signaling pathways mediate the effects of CXCL13 upon NF-κB activation. Our results revealed that NF-κB luciferase activity was augmented by CXCL13 and this effect was prevented when we pretreated the cells with PLCβ, PKCα, and c-Src inhibitors. (Figure 6A). The chromatin immunoprecipitation (ChIP) assay also found that these pathway inhibitors reduced CXCL13-induced binding of p65 to the NF-κB binding site on the VCAM-1 promotor (Figure 6B). Activation of PLCβ, PKCα, and c-Src appears to be necessary for CXCL13-induced NF-κB activation in human osteosarcoma cells.
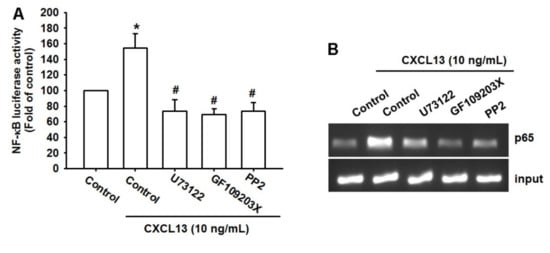
Figure 6.
The PLCβ, PKCα, and c-Src pathways regulate CXCL13-induced NF-κB activation. MG-63 cells were pretreated with U73122, GF109203X, or PP2, then stimulated with CXCL13; NF-κB luciferase activity and p65 binding to the NF-κB site on the VCAM-1 promoter was examined using the luciferase (A) and chromatin immunoprecipitation (ChIP) assays (B). * p < 0.05 compared with the control group; # p < 0.05 compared with the CXCL13-treated group.
3. Discussion
Osteosarcoma is the most common primary tumor of the skeletal system and is well-known to have an aggressive clinical outcome and high metastatic potential [24]. The high propensity for this disease to develop chemoresistance and progression to lung metastasis means that it is critical to develop an effective adjuvant therapy for inhibiting osteosarcoma metastasis. Chemokines are upregulated in several cancer types and can facilitate cancer cell proliferation, control apoptosis, survival, and metastasis. Although CXCL13-regulated cancer cell migration and metastasis has been reported in some cancers [16], the effect of CXCL13 in the motility of osteosarcoma cells remains uncertain. Numerous studies have shown that human physiological concentrations of CXCL13 are significantly increased in cancer patients compared with healthy controls [25]. Other cancer researchers have used treatment concentrations of CXCL13 in the range of 50–100 ng/mL [26,27], so the concentrations used in our study are physiologically relevant. Here, we found that CXCL13 increases the migration and invasion potential of three osteosarcoma cell lines. Interestingly, CXCL13 expression was upregulated in migration-prone MG-63 cells, indicating that CXCL13 levels are associated with the migratory ability of osteosarcoma cells. Treatment of MG-63 cells with inhibitors or transfection with their respective siRNAs did not significantly impact cell proliferation or migration as well as basal CXCL13 and CXCR5 expression, which suggests that these treatments did not affect cell viability (Figure S4). Moreover, we found that CXCL13 facilitates VCAM-1-dependent motility via the PLCβ, PKCα, c-Src, and NF-κB signaling cascades.
Chemokines and their receptors are capable of modulating the proliferative, angiogenic, migratory and invasive activities of cancer cells, so are critical to the progression and metastasis of cancers [28]. CXCL12 and its cell surface receptor CXCR4 combine to enhance the migration of breast cancer cells [29]. The CCL5/CCR5 interaction appears to modulate directional tumor migration and metastasis in different types of cancers, such as lung and breast cancer, as well as osteosarcoma [30,31,32]. Here, we found that CXCR5 siRNA and CXCR5 antibody markedly inhibited CXCL13-induced VCAM-1 expression and cell migration, suggesting that the CXCL13/CXCR5 interaction regulates VCAM-1-mediated migration of human osteosarcoma cells.
Many cellular functions are regulated by the activation of the PLCβ, PKCα, and c-Src signaling pathways [20,33], which also control adhesion molecule production and cell motility [21,34]. In this study, we found that CXCL13 facilitates PLCβ, PKCβ, and c-Src phosphorylation, while PLCβ, PKCα, and c-Src inhibitors reduce CXCL13-induced stimulation of VCAM-1 production and osteosarcoma cell migration. PLCβ, PKCα, and c-Src siRNAs confirmed the reversal of CXCL13-induced promotion of osteosarcoma cell migratory activity. The PLCβ, PKCα, and c-Src signaling pathways appear to play a role in CXCL13-facilitated VCAM-1-dependent osteosarcoma cell migration.
Several transcription factor-binding sites have been identified in the 5-regulatory region [4,35]. NF-κB is a vital transcriptional factor that mediates VCAM-1 transcriptional activity and cancer metastasis [35]. The results of our study show that all three NF-κB inhibitors (BAY 11-7082, PDTC, and TPCK) antagonized the stimulatory effects of CXCL13 upon VCAM-1 expression and cellular migratory activity, indicating the importance of NF-κB activation in these processes. Our data also show that CXCL13 facilitates p65 binding to the NF-κB binding site on the VCAM-1 promotor and NF-κB luciferase activity. The antagonistic effects of PLCβ, PKCα, and c-Src inhibitors upon CXCL13-mediated activities suggest that these are regulated by the PLCβ, PKCα, and c-Src pathways.
4. Materials and Methods
4.1. Materials
We purchased p-PLCβ, PLCβ, p-PKCα, PKCα, p-c-Src, c-Src, p-p65, p65, CXCL13, VCAM-1, p-IKK, IKK, p-IκBα, IκBα, and actin from GeneTex International Corporation (Hsinchu City, Taiwan). All ON-TARGETplus siRNAs were purchased from Dharmacon (Lafayette, CO, USA). CXCR5 mAb was purchased from R&D Systems (Minneapolis, MN, USA). Cell culture supplements were purchased from Invitrogen (Carlsbad, CA, USA). A Dual-Luciferase® Reporter Assay System was bought from Promega (Madison, WI, USA). qPCR primers and probes, as well as the Taqman® One-Step PCR Master Mix, were supplied by Applied Biosystems (Foster City, CA, USA). The PLCβ inhibitor (U73122), PKCα inhibitor (GF109203X), c-Src inhibitor (PP2), IκB inhibitor (TPCK), NF-κB inhibitor (PDTC), NF-κB inhibitor (BAY 11-7082), and other chemicals not already mentioned were supplied by Sigma-Aldrich (St. Louis, MO, USA).
4.2. Cell Culture
The human osteosarcoma cell lines (MG-63, HOS, and U2OS) were obtained from the American Type Cell Culture Collection (Manassas, VA, USA). MG-63 and HOS cells were cultured in MEM medium and U2OS cells were cultured in McCoy’s 5A medium. All cultured mediums were supplemented with 10% FBS and antibiotics then maintained in a humidified incubator at 37 °C in 5% CO2.
The migration-prone MG-63(M2), MG-63(M5), and MG-63(M10) cells were established from original MG-63 cells, as according to the methods described in our previous reports [4,17].
4.3. Western Blot Analysis
We applied the SDS-PAGE procedure to resolve extracted proteins, before transferring them to Immobilon® PVDF membranes. Analysis of the proteins was performed by Western blot, following the procedure detailed in our previous publications [36,37,38].
4.4. mRNA Quantification
Total RNA was extracted from osteosarcoma cells using TRIzol reagent. Quantitative real-time PCR (qPCR) analysis was conducted as according to our previous reports [39,40].
4.5. Migration and Invasion Assay
The migration and invasion assays were performed with Transwell plates (Costar, NY, USA), as according to our previous publications [17,41]. Migrated cells were fixed and stained with crystal violet, then manually counted under a microscope.
4.6. Luciferase Reporter Assay
Osteosarcoma cells were transfected for 24 h with κB-luciferase reporter gene construct, following the Lipofectamine™ 2000 Reagent protocol. The cells were then exposed to CXCL13 and the Promega luciferase activity kit measured luciferase activity [39,42,43].
4.7. Chromatin Immunoprecipitation (ChIP) Assay
DNA were extracted from osteosarcoma cells and immunoprecipitated using anti-p65 antibody. The SimpleChIP® Enzymatic Chromatin IP kit (Cell Signaling, MA, USA) was used to conduct ChIP assays. The purified DNA pellet was subjected to PCR testing, as detailed in our previous publication [4].
4.8. Statistics
All values represent mean ± standard deviation (S.D.). The two-tailed Student’s t-test assessed the significance of between-group differences, using a cut-off point of 0.05.
5. Conclusions
Our study has identified that the CXCL13/CXCR5 axis facilitates VCAM-1 production and the migration of human osteosarcoma cells via the PLCβ, PKCα, c-Src, and NF-κB signaling pathways (Figure 7). CXCL13 and CXCR5 appear to be novel therapeutic targets in metastatic osteosarcoma.
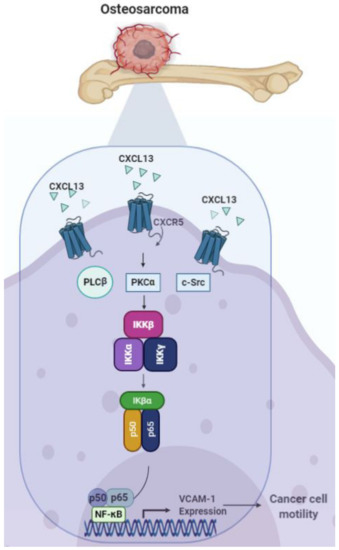
Figure 7.
Schema illustrating the effects of the CXCL13/CXCR5 axis on osteosarcoma cell migration. The CXCL13/CXCR5 interaction facilitates VCAM-1-dependent migration through the PLCβ, PKCα, c-Src, and NF-κB signaling pathways in human osteosarcoma cells.
Supplementary Materials
The following are available online at https://www.mdpi.com/1422-0067/21/17/6095/s1.
Author Contributions
J.-F.L., C.-W.L., Y.-C.F. and C.-H.T. conceived and designed the experiments, which were performed by J.-F.L., C.-Y.L., C.-C.C., T.-M.C., C.-K.H., Y.-L.H. and Y.-C.F. C.-W.L., C.-C.C. and T.-M.C. analyzed the data. C.-W.L., C.-K.H. and Y.-L.H. contributed reagents/materials/analysis tools. Y.-C.F. and C.-H.T. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the Ministry of Science and Technology of Taiwan (MOST 107-2320-B-039-019-MY3, MOST-108-2314-B-039-034-MY3 and 106-2314-B-341-001-MY3), China Medical University (CMU109-ASIA-01), Shin-Kong Wu Ho-Su Memorial Hospital (108-SKH-FJU-06) and Chang Gung Medical Research Program Foundation (grants CMRPF6K0041).
Acknowledgments
We would like to thank Iona J. MacDonald from China Medical University for her English language revision of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fan, T.M.; Roberts, R.D.; Lizardo, M.M. Understanding and Modeling Metastasis Biology to Improve Therapeutic Strategies for Combating Osteosarcoma Progression. Front. Oncol. 2020, 10, 13. [Google Scholar] [CrossRef]
- Isakoff, M.S.; Bielack, S.S.; Meltzer, P.; Gorlick, R. Osteosarcoma: Current Treatment and a Collaborative Pathway to Success. J. Clin. Oncol. 2015, 33, 3029–3035. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.C.; Cheng, S.P.; Han, C.K.; Huang, Y.L.; Wang, S.W.; Lee, J.J.; Lai, C.T.; Fong, Y.C.; Tang, C.H. Resistin enhances angiogenesis in osteosarcoma via the MAPK signaling pathway. Aging 2019, 11, 9767–9777. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.C.; Lee, C.W.; Chang, T.M.; Chen, P.C.; Liu, J.F. CXCL1/CXCR2 Paracrine Axis Contributes to Lung Metastasis in Osteosarcoma. Cancers 2020, 12, 459. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.K.; Tellez-Gabriel, M.; Heymann, D. Cancer stem cells in osteosarcoma. Cancer Lett. 2017, 386, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, M.; Bendas, G. Vascular cell adhesion molecule-1 (VCAM-1)--an increasing insight into its role in tumorigenicity and metastasis. Int. J. Cancer 2015, 136, 2504–2514. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.H.; Kim, Y.K.; Kim, M.R.; Jang, J.H.; Lee, S. Emerging Roles of Vascular Cell Adhesion Molecule-1 (VCAM-1) in Immunological Disorders and Cancer. Int. J. Mol. Sci. 2018, 19, 1057. [Google Scholar] [CrossRef]
- Wang, L.H.; Tsai, H.C.; Cheng, Y.C.; Lin, C.Y.; Huang, Y.L.; Tsai, C.H.; Xu, G.H.; Wang, S.W.; Fong, Y.C.; Tang, C.H. CTGF promotes osteosarcoma angiogenesis by regulating miR-543/angiopoietin 2 signaling. Cancer Lett. 2017, 391, 28–37. [Google Scholar] [CrossRef]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef]
- Do, H.T.T.; Lee, C.H.; Cho, J. Chemokines and their Receptors: Multifaceted Roles in Cancer Progression and Potential Value as Cancer Prognostic Markers. Cancers 2020, 12, 287. [Google Scholar] [CrossRef]
- Lien, M.Y.; Tsai, H.C.; Chang, A.C.; Tsai, M.H.; Hua, C.H.; Wang, S.W.; Tang, C.H. Chemokine CCL4 Induces Vascular Endothelial Growth Factor C Expression and Lymphangiogenesis by miR-195-3p in Oral Squamous Cell Carcinoma. Front. Immunol. 2018, 9, 412. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.T.; Huang, Y.L.; Tzeng, H.E.; Tsai, C.H.; Wang, S.W.; Tang, C.H. CCL5 promotes vascular endothelial growth factor expression and induces angiogenesis by down-regulating miR-199a in human chondrosarcoma cells. Cancer Lett. 2015, 357, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Yamakoshi, Y.; Tanaka, H.; Sakimura, C.; Deguchi, S.; Mori, T.; Tamura, T.; Toyokawa, T.; Muguruma, K.; Hirakawa, K.; Ohira, M. Immunological potential of tertiary lymphoid structures surrounding the primary tumor in gastric cancer. Int. J. Oncol. 2020, 1, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, A.; Ogino, T.; Kayama, H.; Okuzaki, D.; Nishimura, J.; Fujino, S.; Miyoshi, N.; Takahashi, H.; Uemura, M.; Matsuda, C.; et al. Human NKp44+ group 3 innate lymphoid cells associate with tumor-associated tertiary lymphoid structures in colorectal cancer. Cancer Immunol. Res. 2020, 6, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Yang, B.; Sun, Q.; Meng, J.; Zhao, X.; Du, S.; Li, X.; Jiao, S. PD-1 regulates CXCR5(+) CD4 T cell-mediated proinflammatory functions in non-small cell lung cancer patients. Int. Immunopharmacol. 2020, 82, 106295. [Google Scholar] [CrossRef]
- Hussain, M.; Adah, D.; Tariq, M.; Lu, Y.; Zhang, J.; Liu, J. CXCL13/CXCR5 signaling axis in cancer. Life Sci. 2019, 227, 175–186. [Google Scholar] [CrossRef]
- Hou, C.H.; Lin, F.L.; Hou, S.M.; Liu, J.F. Cyr61 promotes epithelial-mesenchymal transition and tumor metastasis of osteosarcoma by Raf-1/MEK/ERK/Elk-1/TWIST-1 signaling pathway. Mol. Cancer 2014, 13, 236. [Google Scholar] [CrossRef]
- Chang, A.C.; Chen, P.C.; Lin, Y.F.; Su, C.M.; Liu, J.F.; Lin, T.H.; Chuang, S.M.; Tang, C.H. Osteoblast-secreted WISP-1 promotes adherence of prostate cancer cells to bone via the VCAM-1/integrin alpha4beta1 system. Cancer Lett. 2018, 426, 47–56. [Google Scholar] [CrossRef]
- Kazanietz, M.G.; Durando, M.; Cooke, M. CXCL13 and Its Receptor CXCR5 in Cancer: Inflammation, Immune Response, and Beyond. Front. Endocrinol. 2019, 10, 471. [Google Scholar] [CrossRef]
- Tsai, C.H.; Yang, D.Y.; Lin, C.Y.; Chen, T.M.; Tang, C.H.; Huang, Y.L. Sphingosine-1-phosphate suppresses chondrosarcoma metastasis by upregulation of tissue inhibitor of metalloproteinase 3 through suppressing miR-101 expression. Mol. Oncol. 2017, 11, 1380–1398. [Google Scholar] [CrossRef]
- Liu, J.F.; Fong, Y.C.; Chang, C.S.; Huang, C.Y.; Chen, H.T.; Yang, W.H.; Hsu, C.J.; Jeng, L.B.; Chen, C.Y.; Tang, C.H. Cyclooxygenase-2 enhances alpha2beta1 integrin expression and cell migration via EP1 dependent signaling pathway in human chondrosarcoma cells. Mol. Cancer 2010, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Lee, C.W.; Tsai, M.H.; Tang, C.H.; Chen, P.C.; Lin, L.W.; Lin, C.Y.; Lu, C.H.; Lin, Y.F.; Yang, S.H.; et al. Thrombospondin 2 promotes tumor metastasis by inducing matrix metalloproteinase-13 production in lung cancer cells. Biochem. Pharmacol. 2018, 155, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Tzeng, H.E.; Tang, C.H.; Wu, S.H.; Chen, H.T.; Fong, Y.C.; Lu, Y.C.; Chen, W.C.; Huang, H.D.; Lin, C.Y.; Wang, S.W. CCN6-mediated MMP-9 activation enhances metastatic potential of human chondrosarcoma. Cell Death Dis. 2018, 9, 955. [Google Scholar] [CrossRef] [PubMed]
- Arndt, C.A.; Crist, W.M. Common musculoskeletal tumors of childhood and adolescence. New Engl. J. Med. 1999, 341, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Curtaz, C.J.; Schmitt, C.; Herbert, S.L.; Feldheim, J.; Schlegel, N.; Gosselet, F.; Hagemann, C.; Roewer, N.; Meybohm, P.; Wockel, A.; et al. Serum-derived factors of breast cancer patients with brain metastases alter permeability of a human blood-brain barrier model. Fluids Barriers CNS 2020, 17, 31. [Google Scholar] [CrossRef] [PubMed]
- Jiao, F.; Sun, H.; Yang, Q.; Sun, H.; Wang, Z.; Liu, M.; Chen, J. Association of CXCL13 and Immune Cell Infiltration Signature in Clear Cell Renal Cell Carcinoma. Int. J. Med Sci. 2020, 17, 1610–1624. [Google Scholar] [CrossRef] [PubMed]
- El Haibi, C.P.; Sharma, P.K.; Singh, R.; Johnson, P.R.; Suttles, J.; Singh, S.; Lillard, J.W., Jr. PI3Kp110-, Src-, FAK-dependent and DOCK2-independent migration and invasion of CXCL13-stimulated prostate cancer cells. Mol. Cancer 2010, 9, 85. [Google Scholar] [CrossRef]
- Lazennec, G.; Richmond, A. Chemokines and chemokine receptors: New insights into cancer-related inflammation. Trends Mol. Med. 2011, 16, 133–144. [Google Scholar] [CrossRef]
- Muller, A.; Homey, B.; Soto, H.; Ge, N.; Catron, D.; Buchanan, M.E.; McClanahan, T.; Murphy, E.; Yuan, W.; Wagner, S.N.; et al. Involvement of chemokine receptors in breast cancer metastasis. Nature 2001, 410, 50–56. [Google Scholar] [CrossRef]
- Huang, C.Y.; Fong, Y.C.; Lee, C.Y.; Chen, M.Y.; Tsai, H.C.; Hsu, H.C.; Tang, C.H. CCL5 increases lung cancer migration via PI3K, Akt and NF-kappaB pathways. Biochem. Pharmacol. 2009, 77, 794–803. [Google Scholar] [CrossRef]
- Velasco-Velazquez, M.; Jiao, X.; De La Fuente, M.; Pestell, T.G.; Ertel, A.; Lisanti, M.P.; Pestell, R.G. CCR5 antagonist blocks metastasis of basal breast cancer cells. Cancer Res. 2012, 72, 3839–3850. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.W.; Wu, H.H.; Liu, S.C.; Wang, P.C.; Ou, W.C.; Chou, W.Y.; Shen, Y.S.; Tang, C.H. CCL5 and CCR5 interaction promotes cell motility in human osteosarcoma. PLoS ONE 2012, 7, e35101. [Google Scholar]
- Chiu, Y.C.; Fong, Y.C.; Lai, C.H.; Hung, C.H.; Hsu, H.C.; Lee, T.S.; Yang, R.S.; Fu, W.M.; Tang, C.H. Thrombin-induced IL-6 production in human synovial fibroblasts is mediated by PAR1, phospholipase C, protein kinase C alpha, c-Src, NF-kappa B and p300 pathway. Mol. Immunol. 2008, 45, 1587–1599. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.T.; Tsou, H.K.; Tsai, C.H.; Kuo, C.C.; Chiang, Y.K.; Chang, C.H.; Fong, Y.C.; Tang, C.H. Thrombin enhanced migration and MMPs expression of human chondrosarcoma cells involves PAR receptor signaling pathway. J Cell. Physiol. 2010, 223, 737–745. [Google Scholar] [CrossRef]
- Sharma, R.; Sharma, R.; Khaket, T.P.; Dutta, C.; Chakraborty, B.; Mukherjee, T.K. Breast cancer metastasis: Putative therapeutic role of vascular cell adhesion molecule-1. Cell. Oncol. 2017, 40, 199–208. [Google Scholar] [CrossRef]
- Lee, H.P.; Chen, P.C.; Wang, S.W.; Fong, Y.C.; Tsai, C.H.; Tsai, F.J.; Chung, J.G.; Huang, C.Y.; Yang, J.S.; Hsu, Y.M.; et al. Plumbagin suppresses endothelial progenitor cell-related angiogenesis in vitro and in vivo. J. Funct. Foods 2019, 52, 537–544. [Google Scholar] [CrossRef]
- Lee, H.P.; Wang, S.W.; Wu, Y.C.; Tsai, C.H.; Tsai, F.J.; Chung, J.G.; Huang, C.Y.; Yang, J.S.; Hsu, Y.M.; Yin, M.C.; et al. Glucocerebroside reduces endothelial progenitor cell-induced angiogenesis. Food Agric. Immunol. 2019, 30, 1033–1045. [Google Scholar] [CrossRef]
- Liu, S.C.; Tsai, C.H.; Wu, T.Y.; Tsai, C.H.; Tsai, F.J.; Chung, J.G.; Huang, C.Y.; Yang, J.S.; Hsu, Y.M.; Yin, M.C.; et al. Soya-cerebroside reduces IL-1 beta-induced MMP-1 production in chondrocytes and inhibits cartilage degradation: Implications for the treatment of osteoarthritis. Food Agric. Immunol. 2019, 30, 620–632. [Google Scholar] [CrossRef]
- Yang, Y.C.; Chiou, P.C.; Chen, P.C.; Liu, P.Y.; Huang, W.C.; Chao, C.C.; Tang, C.H. Melatonin reduces lung cancer stemness through inhibiting of PLC, ERK, p38, beta-catenin, and Twist pathways. Environ. Toxicol. 2019, 34, 203–209. [Google Scholar] [CrossRef]
- Wang, M.; Chao, C.C.; Chen, P.C.; Liu, P.I.; Yang, Y.C.; Su, C.M.; Huang, W.C.; Tang, C.H. Thrombospondin enhances RANKL-dependent osteoclastogenesis and facilitates lung cancer bone metastasis. Biochem. Pharmacol. 2019, 166, 23–32. [Google Scholar] [CrossRef]
- Lee, H.P.; Wang, S.W.; Wu, Y.C.; Lin, L.W.; Tsai, F.J.; Yang, J.S.; Li, T.M.; Tang, C.H. Soya-cerebroside inhibits VEGF-facilitated angiogenesis in endothelial progenitor cells. Food Agric. Immunol. 2020, 31, 193–204. [Google Scholar] [CrossRef]
- Su, C.M.; Tang, C.H.; Chi, M.J.; Lin, C.Y.; Fong, Y.C.; Liu, Y.C.; Chen, W.C.; Wang, S.W. Resistin facilitates VEGF-C-associated lymphangiogenesis by inhibiting miR-186 in human chondrosarcoma cells. Biochem. Pharmacol. 2018, 154, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.J.; Lin, C.Y.; Tsai, C.H.; Huang, Y.L.; Tang, C.H. Glucose suppresses IL-1beta-induced MMP-1 expression through the FAK, MEK, ERK, and AP-1 signaling pathways. Environ. Toxicol. 2018, 33, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).