Canonical, Non-Canonical and Atypical Pathways of Nuclear Factor кb Activation in Preeclampsia

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Results
2.1. Characterization of Study Population
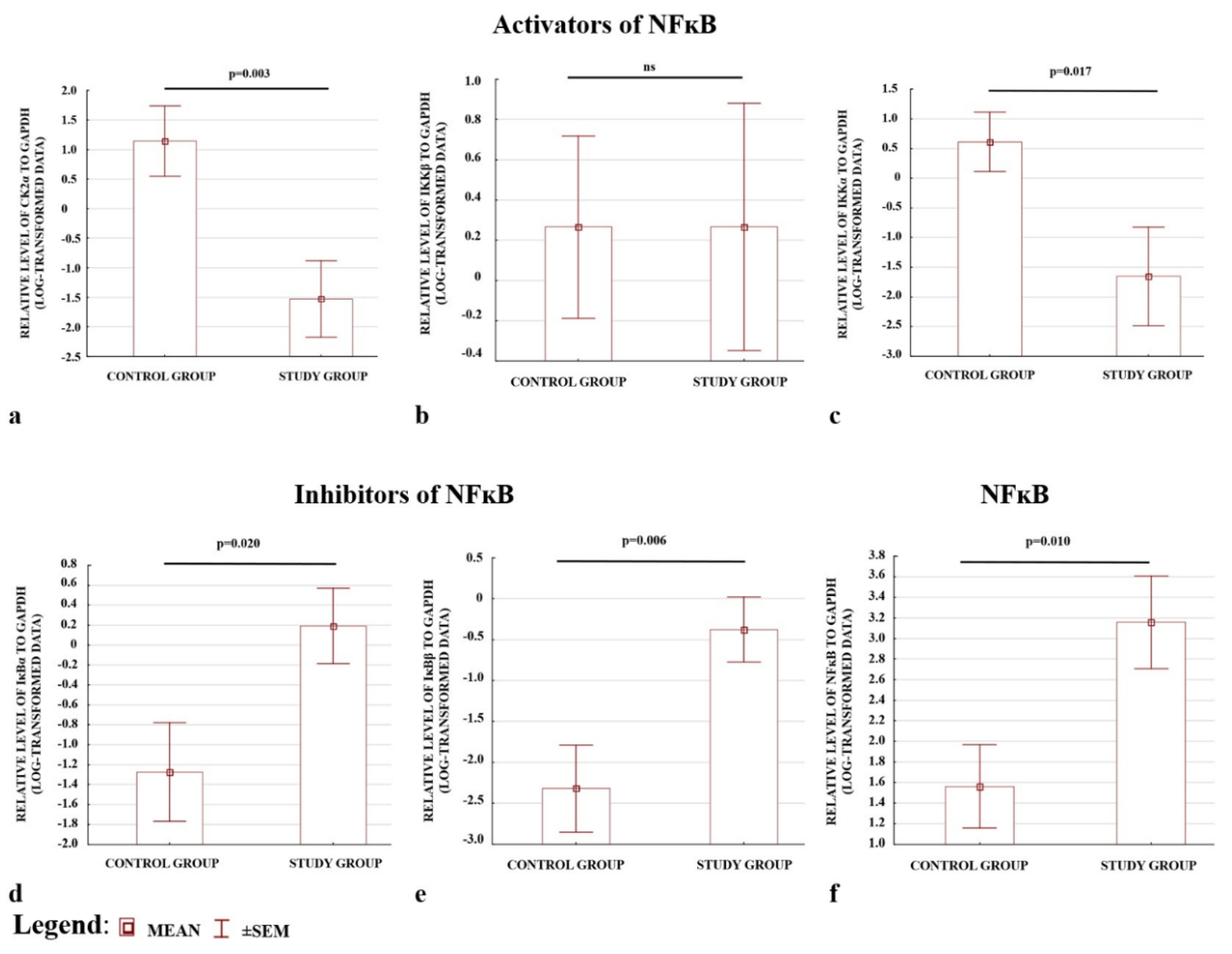
2.2. Preeclamptic Placentas Differ in the Expression Level of Activators and One of the Inhibitors of NFκB from Non-Complicated Cases
2.3. The Protein Levels of IκBα and IκBβ Inhibitors Were Higher, Whereas IKKα and CK2α Activators Were Lower in Preeclamptic Placentas
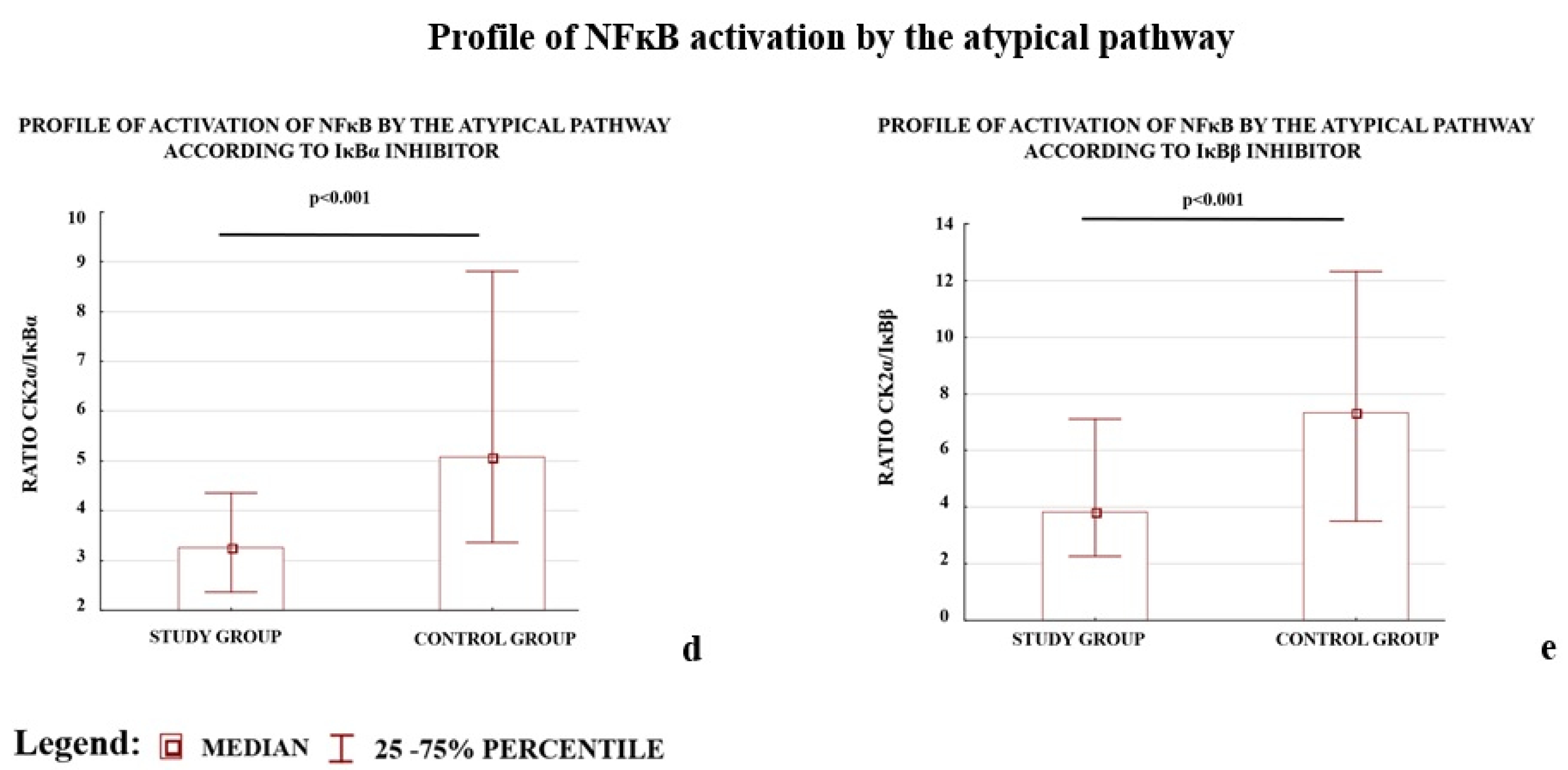
2.4. Western Blot Results Confirmed the ELISA Observations; the Levels of CK2α and IKKα Were Depleted, whereas IκBα and IκBβ Were Elevated in Preeclamptic Placentas
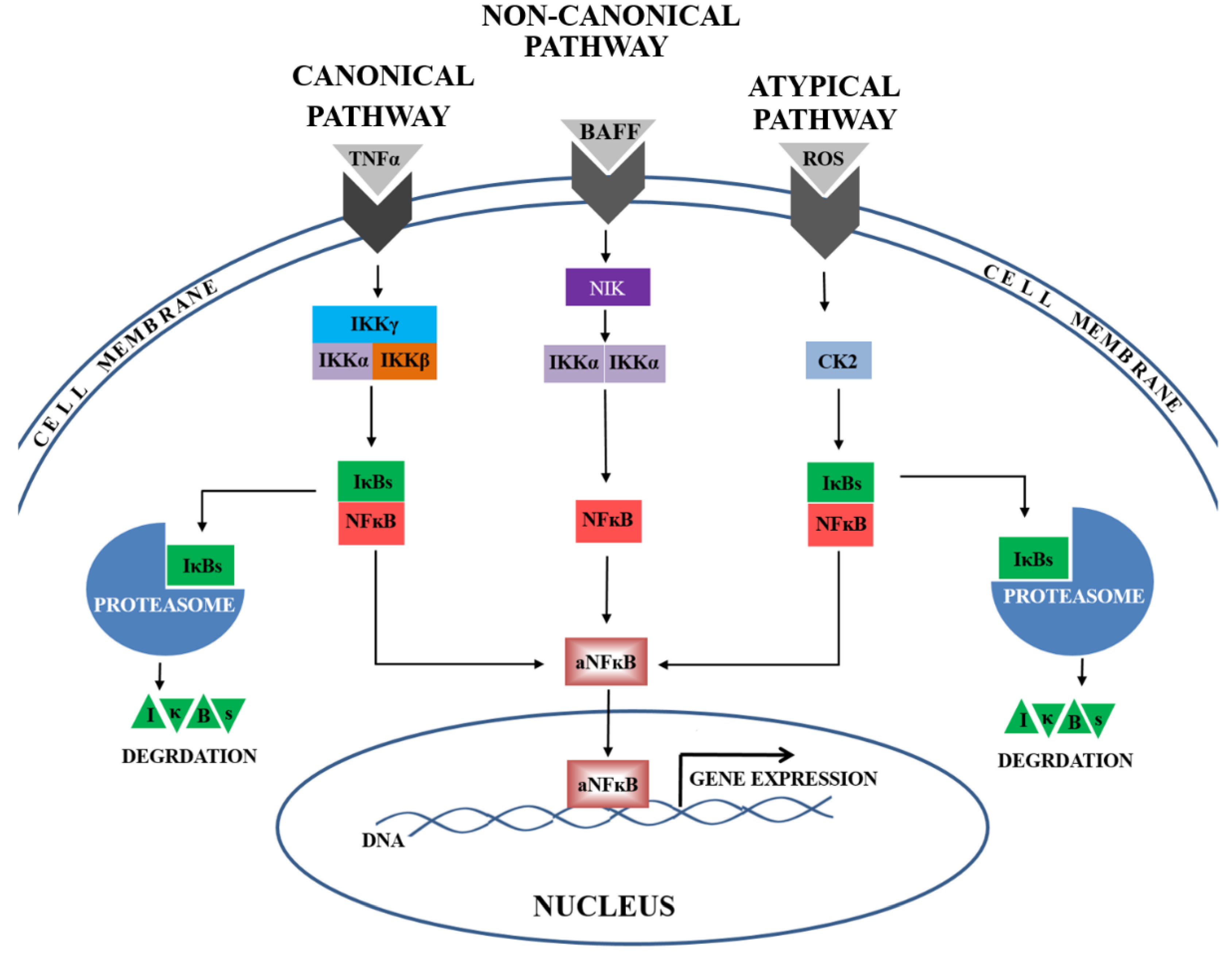
3. Discussion
4. Materials and Methods
4.1. Study and Control Groups
4.2. Collection of Placental Samples
4.3. Gene Expression Analysis
4.4. Protein Isolation
4.5. ELISA Analyses
4.6. Western Blot Analysis
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ahmed, R.; Dunford, J.; Mehran, R.; Robson, S.; Kunadian, V. Pre-eclampsia and future cardiovascular risk among women: A review. J. Am. Coll. Cardiol. 2014, 63, 1815–1822. [Google Scholar] [CrossRef] [PubMed]
- Hypertension in Pregnancy Report of the American College of Obstetricians and Gynecologists’ Task Force on Hypertension in Pregnancy Executive. Obstet. Gynecol. 2013, 122, 1122–1131. [CrossRef]
- Tranquilli, A.L.; Dekker, G.; Magee, L.; Roberts, J.; Sibai, B.M.; Steyn, W.; Zeeman, G.G.; Brown, M.A. The classification, diagnosis and management of the hypertensive disorders of pregnancy: A revised statement from the ISSHP. Pregnancy Hypertens. 2014, 4, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Serasanambati, M.; Chilakapati, S.R. Function of Nuclear Factor Kappa B (NF-kB) in Human Diseases—A Review. South Indian J. Biol. Sci. 2016, 2, 368. [Google Scholar] [CrossRef]
- Viatour, P.; Merville, M.P.; Bours, V.; Chariot, A. Phosphorylation of NF-κB and IκB proteins: Implications in cancer and inflammation. Trends Biochem. Sci. 2005, 30, 43–52. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, 1–14. [Google Scholar] [CrossRef]
- Brasier, A.R. The nuclear factor-B-interleukin-6 signalling pathway mediating vascular inflammation. Cardiovasc. Res. 2010, 86, 211–218. [Google Scholar] [CrossRef]
- Sun, S.-C. The non-canonical NF-kappaB pathway in immunity and inflammation. Nat. Rev. Immunol. 2017, 17, 545–558. [Google Scholar] [CrossRef]
- Janssens, S.; Tschopp, J. Signals from within: The DNA-damage-induced NF-κB response. Cell Death Differ. 2006, 13, 773–784. [Google Scholar] [CrossRef]
- González-Ramos, R.; Defrère, S.; Devoto, L. Nuclear factor-kappaB: A main regulator of inflammation and cell survival in endometriosis pathophysiology. Fertil. Steril. 2012, 98, 520–528. [Google Scholar] [CrossRef]
- Huang, T.T.; Kudo, N.; Yoshida, M.; Miyamoto, S. A nuclear export signal in the N-terminal regulatory domain of IκBα controls cytoplasmic localization of inactive NF-κB/IκBα complexes. Proc. Natl. Acad. Sci. USA 2000, 97, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Nishikori, M. Classical and Alternative NF-.KAPPA.B Activation Pathways and Their Roles in Lymphoid Malignancies. J. Clin. Exp. Hematop. 2005, 45, 15–24. [Google Scholar] [CrossRef]
- Suyang, H.; Phillips, R.; Douglas, I.; Ghosh, S. Role of unphosphorylated, newly synthesized I kappa B beta in persistent activation of NF-kappa B. Mol. Cell. Biol. 1996, 16, 5444–5449. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Allport, V.; Sykes, A.; Lindstrom, T.; Slater, D.; Bennett, P. The effects of labour and of interleukin 1 beta upon the expression of nuclear factor kappa B related proteins in human amnion. Mol. Hum. Reprod. 2003, 9, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Bohuslav, J.; Chen, L.F.; Kwon, H.; Mu, Y.; Greene, W.C. p53 induces NF-κB activation by an IκB kinase-independent mechanism involving phosphorylation of p65 by ribosomal S6 kinase 1. J. Biol. Chem. 2004, 279, 26115–26125. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, C.Y.; Barberi, T.J.; Ghosh, P.; Longo, D.L. Phosphorylation of Re1A/p65 on serine 536 defines an IκBα- independent NF-κB pathway. J. Biol. Chem. 2005, 280, 34538–34547. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.M.; Escudero, C. The placenta in preeclampsia. Pregnancy Hypertens. 2012, 2, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.J. Pre-eclampsia: Is it all in the placenta? Malaysian J. Med. Sci. 2009, 16, 7–15. [Google Scholar]
- Soleymanlou, N.; Jurisica, I.; Nevo, O.; Ietta, F.; Zhang, X.; Zamudio, S.; Post, M.; Caniggia, I. Molecular evidence of placental hypoxia in preeclampsia. J. Clin. Endocrinol. Metab. 2005, 90, 4299–4308. [Google Scholar] [CrossRef]
- Conrad, K.P.; Bfnyo, D.F. Placental cytokines and the pathogenesis of preeclampsia. Am. J. Reprod. Immunol. 1997, 37, 240–249. [Google Scholar] [CrossRef]
- Lockwood, C.J.; Yen, C.F.; Basar, M.; Kayisli, U.A.; Martel, M.; Buhimschi, I.; Buhimschi, C.; Huang, S.J.; Krikun, G.; Schatz, F. Preeclampsia-related inflammatory cytokines regulate interleukin-6 expression in human decidual cells. Am. J. Pathol. 2008, 172, 1571–1579. [Google Scholar] [CrossRef] [PubMed]
- Myatt, L. Role of placenta in preeclampsia. Endocrine 2002, 19, 103–111. [Google Scholar] [CrossRef]
- Rani, S.; Chawla, D. Preeclampsia and Reactive Oxygen Species. Indian J. Pediatr. 2018, 85, 333–334. [Google Scholar] [CrossRef] [PubMed]
- Hladunewich, M.; Karumanchi, S.A.; Lafayette, R.A. Pathophysiology of the clinical manifestations of preeclampsia. Clin. J. Am. Soc. Nephrol. 2007, 2, 543–549. [Google Scholar] [CrossRef]
- Aban, M.; Cinel, L.; Arslan, M.; Dilek, U.; Kaplanoglu, M.; Arpaci, R.; Dilek, S. Expression of nuclear factor-kappa B and placental apoptosis in pregnancies complicated with intrauterine growth restriction and preeclampsia: An immunohistochemical study. Tohoku J. Exp. Med. 2004, 204, 195–202. [Google Scholar] [CrossRef]
- Roberts, J.M.; Hubel, C.A. Oxidative stress in preeclampsia. Am. J. Obstet. Gynecol. 2004, 190, 1177–1178. [Google Scholar] [CrossRef]
- Armistead, B.; Kadam, L.; Drewlo, S.; Kohan-Ghadr, H.R. The role of NFκB in healthy and preeclamptic placenta: Trophoblasts in the spotlight. Int. J. Mol. Sci. 2020, 21, 1775. [Google Scholar] [CrossRef]
- Silva Carmona, A.; Mendieta Zerón, H. NF-κB and SOD expression in preeclamptic placentas. Turk. J. Med. Sci. 2016, 46, 783–788. [Google Scholar] [CrossRef]
- Vaughan, J.E.; Walsh, S.W. Activation of NF-κB in Placentas of Women with Preeclampsia. Hypertens. Pregnancy 2012, 31, 243–251. [Google Scholar] [CrossRef]
- Li, F.-H.; Wang, Y.; Liu, X.-L.; Xu, Q. The silencing of ApoC3 suppresses oxidative stress and inflammatory responses in placenta cells from mice with preeclampsia via inhibition of the NF-kappaB signaling pathway. Biomed. Pharmacother. 2018, 107, 1377–1384. [Google Scholar] [CrossRef]
- Matthiesen, L.; Berg, G.; Ernerudh, J.; Ekerfelt, C.; Jonsson, Y.; Sharma, S. Immunology of preeclampsia. Chem. Immunol. Allergy 2005, 89, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Barkett, M.; Gilmore, T.D. Control of apoptosis by Rel/NF-κB transcription factors. Oncogene 1999, 18, 6910–6924. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-kappaB signaling in inflammation. Signal. Transduct. Target Ther. 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Liu, Z. Crosstalk of reactive oxygen species and NF-kappaB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Litang, Z.; Hong, W.; Weimin, Z.; Xiaohui, T.; Qian, S. Serum NF-κBp65, TLR4 as biomarker for diagnosis of preeclampsia. Open Med. 2017, 12, 399–402. [Google Scholar] [CrossRef]
- King, A.E.; Critchley, H.O.; Kelly, R.W. The NF-kappaB pathway in human endometrium and first trimester decidua. Mol. Hum. Reprod. 2001, 7, 175–183. [Google Scholar] [CrossRef]
- Sakowicz, A.; Hejduk, P.; Pietrucha, T.; Nowakowska, M.; Płuciennik, E.; Pospiech, K.; Gach, A.; Rybak-Krzyszkowska, M.; Sakowicz, B.; Kaminski, M.; et al. Finding NEMO in preeclampsia. Am. J. Obstet. Gynecol. 2016, 214, 538.e1–538.e7. [Google Scholar] [CrossRef]
- Sakowicz, A.; Lisowska, M.; Biesiada, L.; Płuciennik, E.; Gach, A.; Rybak-Krzyszkowska, M.; Huras, H.; Sakowicz, B.; Romanowicz, H.; Piastowska-Ciesielska, A.W.; et al. Placental Expression of NEMO Protein in Normal Pregnancy and Preeclampsia. Dis. Markers 2019, 2019, 1–12. [Google Scholar] [CrossRef]
- Huang, W.-C.; Hung, M.-C. Beyond NF-kappaB activation: Nuclear functions of IkappaB kinase alpha. J. Biomed. Sci. 2013, 20, 3. [Google Scholar] [CrossRef]
- Haider, S.; Knofler, M. Human tumour necrosis factor: Physiological and pathological roles in placenta and endometrium. Placenta 2009, 30, 111–123. [Google Scholar] [CrossRef]
- Jia, X.-Q.; Xu, S.; Tian, M.-R.; Ma, Y.-Y. The relationship between inflammatory factor expression and blood pressure and urinary protein in the placenta of gestational hypertension rats. Exp. Ther. Med. 2018, 16, 3793–3798. [Google Scholar] [CrossRef] [PubMed]
- Benyo, D.F.; Smarason, A.; Redman, C.W.; Sims, C.; Conrad, K.P. Expression of inflammatory cytokines in placentas from women with preeclampsia. J. Clin. Endocrinol. Metab. 2001, 86, 2505–2512. [Google Scholar] [CrossRef] [PubMed]
- Prins, J.R.; Gomez-Lopez, N.; Robertson, S.A. Interleukin-6 in pregnancy and gestational disorders. J. Reprod. Immunol. 2012, 95, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Jiang, Y.-H.; Yang, C.-H.; Meng, X.-Q.; Hao, N.; Qi, Y.-Z. Establishment and Evaluation of Hypertensive Rat Model with Excessive Accumulation of Phlegm-dampness Syndrome. Zhongguo Zhong xi yi jie he za zhi Zhongguo Zhongxiyi jiehe zazhi Chin. J. Integr. Tradit. West. Med. 2016, 36, 222–228. (In Chinese) [Google Scholar]
- Rabalski, A.J.; Gyenis, L.; Litchfield, D.W. Molecular pathways: Emergence of protein kinase CK2 (CSNK2) as a potential target to inhibit survival and DNA damage response and repair pathways in cancer cells. Clin. Cancer Res. 2016, 22, 2840–2847. [Google Scholar] [CrossRef]
- Wang, G.; Unger, G.; Ahmad, K.A.; Slaton, J.W.; Ahmed, K. Downregulation of CK2 induces apoptosis in cancer cells—A potential approach to cancer therapy. Mol. Cell Biochem. 2005, 274, 77–84. [Google Scholar] [CrossRef]
- Lee, S.W.; Song, Y.S.; Lee, S.Y.; Yoon, Y.G.; Lee, S.H.; Park, B.S.; Yun, I.; Choi, H.; Kim, K.; Chung, W.T.; et al. Downregulation of protein kinase CK2 activity facilitates tumor necrosis factor-alpha-mediated chondrocyte death through apoptosis and autophagy. PLoS ONE 2011, 6, e19163. [Google Scholar] [CrossRef]
- Tenório, M.B.; Ferreira, R.C.; Moura, F.A.; Bueno, N.B.; De Oliveira, A.C.M.; Goulart, M.O.F. Cross-Talk between Oxidative Stress and Inflammation in Preeclampsia. Oxid. Med. Cell Longev. 2019, 2019. [Google Scholar] [CrossRef]
- Aouache, R.; Biquard, L.; Vaiman, D.; Miralles, F. Oxidative Stress in Preeclampsia and Placental Diseases. Int. J. Mol. Sci. 2018, 19, 1496. [Google Scholar] [CrossRef]
- Abi Nahed, R.; Reynaud, D.; Lemaitre, N.; Lartigue, S.; Roelants, C.; Vaiman, D.; Benharouga, M.; Cochet, C.; Filhol, O.; Alfaidy, N. Protein kinase CK2 contributes to placental development: Physiological and pathological implications. J. Mol. Med. 2020, 98, 123–133. [Google Scholar] [CrossRef]
- Li, H.; Dakour, J.; Kaufman, S.; Guilbert, L.J.; Winkler-Lowen, B.; Morrish, D.W. Adrenomedullin is decreased in preeclampsia because of failed response to epidermal growth factor and impaired syncytialization. Hypertension 2003, 42, 895–900. [Google Scholar] [CrossRef] [PubMed]
- Lyall, F.; Robson, S.C.; Bulmer, J.N. Spiral artery remodeling and trophoblast invasion in preeclampsia and fetal growth restriction: Relationship to clinical outcome. Hypertension 2013, 62, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Jiang, S.; Chen, J.; Li, J.; Ao, L.; Zhang, Y. The Increased lncRNA MIR503HG in Preeclampsia Modulated Trophoblast Cell Proliferation, Invasion, and Migration via Regulating Matrix Metalloproteinases and NF-kappaB Signaling. Dis. Mark. 2019, 2019, 4976845. [Google Scholar] [CrossRef] [PubMed]
- Yung, H.W.; Atkinson, D.; Campion-Smith, T.; Olovsson, M.; Charnock-Jones, D.S.; Burton, G.J. Differential activation of placental unfolded protein response pathways implies heterogeneity in causation of early- and late-onset pre-eclampsia. J. Pathol. 2014, 234, 262–276. [Google Scholar] [CrossRef]
- Ryan, K.M.; Ernst, M.K.; Rice, N.R.; Vousden, K.H. Role of NF-κB in p53-mediated programmed cell death. Nature 2000, 404, 892–897. [Google Scholar] [CrossRef]
- Sharp, A.N.; Heazell, A.E.P.; Baczyk, D.; Dunk, C.E.; Lacey, H.A.; Jones, C.J.P.; Perkins, J.E.; Kingdom, J.C.P.; Baker, P.N.; Crocker, I.P. Preeclampsia is associated with alterations in the p53-pathway in villous trophoblast. PLoS ONE 2014, 9, e87621. [Google Scholar] [CrossRef]
- Tan, B.; Tong, C.; Yuan, Y.; Xu, P.; Wen, L.; Zhang, C.; Zheng, Y.; Lin, L.; Zhu, F.; Gui, S.; et al. The regulation of trophoblastic p53 homeostasis by the p38-WIP1 feedback loop is disturbed in placentas from pregnancies complicated by preeclampsia. Cell Physiol. Biochem. 2019, 52, 315–335. [Google Scholar] [CrossRef]
- Levy, R.; Smith, S.D.; Yusuf, K.; Huettner, P.C.; Kraus, F.T.; Sadovsky, Y.; Nelson, D.M. Trophoblast apoptosis from pregnancies complicated by fetal growth restriction is associated with enhanced p53 expression. Am. J. Obstet. Gynecol. 2002, 186, 1056–1061. [Google Scholar] [CrossRef]
- Rao, P.; Hayden, M.S.; Long, M.; Scott, M.L.; West, A.P.; Zhang, D.; Oeckinghaus, A.; Lynch, C.; Hoffmann, A.; Baltimore, D.; et al. IκBβ acts to inhibit and activate gene expression during the inflammatory response. Nature 2010, 466, 1115–1119. [Google Scholar] [CrossRef]
- Ferreiro, D.U.; Komives, E.A. Molecular mechanisms of system control of NF-kappaB signaling by IkappaBalpha. Biochemistry 2010, 49, 1560–1567. [Google Scholar] [CrossRef]
- Verma, I.M.; Stevenson, J.K.; Schwarz, E.M.; Van Antwerp, D.; Miyamoto, S. Rel/NF-ĸB/IĸB family; intimate tales of association and dissociation. Genes Dev. 1995, 9, 2723–2735. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. Relative expression software tool (REST(C)) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time quantitative reverse transcription- PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Health and Human Services. Food and Drug Administration Guidance for Industry Bioanalytical Method Validation Guidance for Industry Bioanalytical Method Validation. 2018. Available online: https://www.fda.gov/media/70858/download (accessed on 24 May 2018).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Study Group N = 130 | Control Group N = 138 | p |
|---|---|---|---|
| Maternal age at the time of delivery (years) 1 | 31 (27–35) | 32 (30–35) | 0.056 |
| WBC (103/uL) 1 | 10.1 (8.9–12.1) | 10.1 (8.9–11.7) | 0.699 |
| RBC (106/uL) 1 | 4.2 (3.8–4.4) | 4.2 (3.9–4.4) | 0.365 |
| HB (g/dL) 1 | 12.4 (11.3–13.1) | 12.5 (11.8–13.1) | 0.298 |
| HCT (%) 2 | 35 ± 3.3 | 36 ± 2.7 | 0.011 |
| MCV (fL) 1 | 85.5 (83.3–88.0) | 87.2 (84.0–90.9) | 0.003 |
| MCHC (g/dL) 1 | 34.7 (33.8–35.3) | 34.1 (33.2–34.9) | <0.001 |
| PLT (103/uL) 2 | 195 ± 63 | 221 ± 55 | <0.001 |
| BMI (kg/m2) 1 | 26 (24–29) | 24 (22–28) | <0.001 |
| Newborn weight (g) 1 | 2130 (1500–3100) | 3370 (3100–3710) | <0.001 |
| Newborn length (cm) 1 | 48 (42–52) | 54 (52–56) | <0.001 |
| Week of delivery (week) 1 | 32 (29–33) (early preeclampsia) 38 (36–39) (late preeclampsia) | 39 (38–39) | <0.001 <0.001 |
| Primiparous n (%) 3 | 83 (64%) | 47 (34%) | <0.001 |
| History of miscarriage n (%) 3 | 23 (18%) | 25 (18%) | 0.928 |
| Male sex of the fetus n (%) 3 | 74 (57%) | 69 (50%) | 0.256 |
| Gene Symbol or Ratio | Study Group Median * (Interquartile Range) N = 130 | Control Group Median * (Interquartile Range) N = 138 | Fold Change | p ** |
|---|---|---|---|---|
| NFκB activators for the canonical, non-canonical and atypical pathways | ||||
| CHUK | 1.42 (0.81–2.45) | 2.38 (1.22–4.12) | −1.68 | <0.001 |
| IKBKB | 0.88 (0.45–1.66) | 1.44 (0.94–2.43) | −1.64 | <0.001 |
| CSNK2A1 | 1.07 (0.71–1.54) | 1.52 (0.97–2.46) | −1.42 | <0.001 |
| MAP3K14 | 1.48 (0.51–3.56) | 0.92 (0.50–3.26) | 1.61 | 0.393 |
| NFκB inhibitors | ||||
| IΚBA | 1.32 (0.83–1.78) | 1.07 (0.76–1.38) | 1.23 | 0.004 |
| IKBB | 1.02 (0.60–1.89) | 0.84 (0.53–1.71) | 1.21 | 0.098 |
| Profile of NFκB activation by the canonical pathway | ||||
| (CHUK+IKBKB)/IΚBA | 2.14 (1.27–4.28) | 3.80 (2.60–7.10) | −1.78 | <0.001 |
| (CHUK+IKBKB)/IKBB | 2.34 (1.05–4.19) | 4.29 (2.15–8.87) | −1.83 | <0.001 |
| Profile of NFκB activation by the non-canonical pathway | ||||
| CHUK *** | 1.42 (0.81–2.45) | 2.39 (1.22–4.12) | −1.68 | <0.001 |
| CHUK+MAP3K14 *** | 3.47 (1.87–5.91) | 3.44 (1.96–8.15) | 1.00 | 0.328 |
| Profile of NFκB activation by the atypical pathway | ||||
| CSNK2A1/IΚBA | 0.86 (0.53–1.47) | 1.62 (0.99–2.58) | −1.88 | <0.001 |
| CSNK2A1/IKBB | 1.11 (0.50–1.89) | 1.66 (0.80–3.23) | −1.50 | <0.001 |
| Gene Symbol | Study Group Median 1 (Interquartile Range) N = 114 | Control Group Median 1 (Interquartile Range) N = 133 | p2 |
|---|---|---|---|
| NFκB activators for the canonical, non-canonical and atypical 3 pathway | |||
| IKKα ** | 17.8 (11.6–25.5) | 21.3 (15.1–31.4) | 0.001 |
| IKKβ * | 11.6 (9.1–15.9) | 13.1 (8.7–22.0) | 0.147 |
| CK2α ** | 255.6 (191.3–403.5) | 343.1 (238.6–456.0) | <0.001 |
| NFκB inhibitors | |||
| IκBα ** | 84.2 (59.3–109.7) | 64.0 (39.9–91.2) | <0.001 |
| IκBβ ** | 64.9 (39.1–99.5) | 51.0 (30.6–78.3) | 0.004 |
| Unigene | GeneBank | Gene Symbol/ (Protein Symbol) | The Full Name of the Gene/Protein |
|---|---|---|---|
| NFκB activators for the canonical, non-canonical and atypical pathways | |||
| Hs.198998 | NM_001278 | CHUK/ (IKKα) 1,2 | Conserved Helix-loop-helix Ubiquitous Kinase or Inhibitor of Nuclear Factor Kappa-B Kinase Subunit Alpha/I-Kappa-B-Kinase Alpha |
| Hs.597664 | NM_001190720 | IKBKB/ (IKKβ) 1 | Inhibitor of Nuclear Factor Kappa B Kinase Subunit Beta I-Kappa-B-Kinase Beta |
| Hs.644056 | NM_001895 | CSNK2A1/ (CK2α) 3 | Casein Kinase 2 Alpha 1 |
| Hs.404183 | NM_003954 | MAP3K14/ (NIK) 2 | Mitogen-Activated Protein Kinase Kinase Kinase 14/NF-Kappa-Beta-Inducing Kinase |
| NFκB inhibitors | |||
| Hs.81328 | NM_020529 | IKBA(IκBα) 1,3 | Nuclear Factor of Kappa Light Polypeptide Gene Enhancer In B-Cells Inhibitor, Alpha/NF-Kappa-B Inhibitor Alpha |
| Hs.9731 | NM_001001716 | IKBB(IκBβ) 1,3 | Nuclear Factor of Kappa Light Polypeptide Gene Enhancer In B-Cells Inhibitor, Beta/NF-Kappa-B Inhibitor Beta |
| Housekeeping gene candidate for placental samples | |||
| Hs.520640 | NM_001101 | ACTB | Actin Beta |
| Hs.544577 | NM_001256799 | GAPDH * | Glyceraldehyde-3-Phosphate Dehydrogenase |
| Hs.546285 | NM_001002 | RPLP0 | Ribosomal Protein Lateral Stalk Subunit P0 |
| Hs.520348 | NM_021009 | UBC | Ubiquitin C |
| Hs.492407 | NM_001135699 | YWHAZ * | Tyrosine 3-Monooxygenase/Tryptophan 5-Monooxygenase Activation Protein Zeta |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakowicz, A.; Bralewska, M.; Pietrucha, T.; Habrowska-Górczyńska, D.E.; Piastowska-Ciesielska, A.W.; Gach, A.; Rybak-Krzyszkowska, M.; Witas, P.J.; Huras, H.; Grzesiak, M.; et al. Canonical, Non-Canonical and Atypical Pathways of Nuclear Factor кb Activation in Preeclampsia. Int. J. Mol. Sci. 2020, 21, 5574. https://doi.org/10.3390/ijms21155574
Sakowicz A, Bralewska M, Pietrucha T, Habrowska-Górczyńska DE, Piastowska-Ciesielska AW, Gach A, Rybak-Krzyszkowska M, Witas PJ, Huras H, Grzesiak M, et al. Canonical, Non-Canonical and Atypical Pathways of Nuclear Factor кb Activation in Preeclampsia. International Journal of Molecular Sciences. 2020; 21(15):5574. https://doi.org/10.3390/ijms21155574
Chicago/Turabian StyleSakowicz, Agata, Michalina Bralewska, Tadeusz Pietrucha, Dominika E Habrowska-Górczyńska, Agnieszka W Piastowska-Ciesielska, Agnieszka Gach, Magda Rybak-Krzyszkowska, Piotr J Witas, Hubert Huras, Mariusz Grzesiak, and et al. 2020. "Canonical, Non-Canonical and Atypical Pathways of Nuclear Factor кb Activation in Preeclampsia" International Journal of Molecular Sciences 21, no. 15: 5574. https://doi.org/10.3390/ijms21155574
APA StyleSakowicz, A., Bralewska, M., Pietrucha, T., Habrowska-Górczyńska, D. E., Piastowska-Ciesielska, A. W., Gach, A., Rybak-Krzyszkowska, M., Witas, P. J., Huras, H., Grzesiak, M., & Biesiada, L. (2020). Canonical, Non-Canonical and Atypical Pathways of Nuclear Factor кb Activation in Preeclampsia. International Journal of Molecular Sciences, 21(15), 5574. https://doi.org/10.3390/ijms21155574